Introduction
Colorectal cancer is one of the most common causes
of cancer-associated morbidity and mortality globally (1,2).
Despite advances in treatment, including surgery, chemotherapy and
radiotherapy, the prognosis of advanced colorectal cancer remains
poor (1,2). Therefore, development of an efficient
and safe anticancer agent is required to improve the prognosis of
patients with advanced colorectal cancer.
Reversine, a 2,6-diamino-substituted purine
analogue, was initially used as a dedifferentiation agent that
induces lineage reversal of C2C12 murine myoblasts cells to become
multipotent progenitor cells, which can subsequently
redifferentiate into osteoblasts, adipocytes, neural lineage cells
and cardiomyocytes under lineage-specific inducing conditions
(3-6). The activity of reversine as a
dedifferentiation agent may be applicable to cancer biology and
therapy.
The Aurora kinases, a family of mitotic
serine/threonine kinases, serve a critical role in regulating
cell-cycle progression, and aberrant expression of these kinases
has been reported in a broad range of human cancer types, including
breast, ovarian and gastric cancer (7-9).
Numerous studies demonstrated that reversine possesses anticancer
properties, causing cell growth arrest, cell cycle arrest,
apoptosis, polyploidy and autophagy via the inhibition of Aurora
kinases in various human cancer cell lines, including breast,
thyroid, kidney, cervix, lung and colon cancer (10-23).
Additionally, reversine inhibits tumor growth, as observed from
xenograft mice model experiments (22,23).
The results indicated that reversine may be used as an anticancer
agent.
However, it remains unclear whether reversine exerts
an anticancer effect in human colorectal cancer cells. The aim of
the present study was to investigate the impact of reversine on
tumor cell behavior and its association with intracellular
signaling pathways in human colorectal cancer cells.
Materials and methods
Cell culture and materials
Human colorectal carcinoma cell lines SW480
(ATCC® CCL-228™) and HCT116 (ATCC® CCL-247™)
were purchased from the American Type Culture Collection (Manassas,
VA, USA). The SW480 cells were cultured in Dulbecco’s modified
Eagle’s medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). The HCT116 cells were cultured in McCoy’s 5A medium (Welgene,
Inc., Gyeongsan, South Korea) supplemented with 10% fetal bovine
serum (Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin at 37°C in a humidified atmosphere of 5%
CO2. The pan-caspase inhibitor Z-VAD-FMK was purchased
from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). Inhibition of
Fas and death receptor 5 (DR5) was performed using the small
interfering RNA (siRNA). To transfect siRNA, SW480 and HCT116 cells
were seeded into 6-well plates at a density of 3×105
cells/well at 37°C such that they would be 40-50% confluent at the
time of transfection. On the next day, 20 µM siRNA was
transfected with 5 µl Lipofectamine® RNAiMAX
reagent (Invitrogen; Thermo Fisher Scientific, Inc.), according to
the manufacturer’s protocols. Further experiments were performed 24
h post transfection. Fas and DR5 siRNA were purchased from Santa
Cruz Biotechnology, Inc. (Dallas, TX, USA). Scrambled siRNA
(AllStars Negative Control siRNA; Qiagen GmbH, Hilden, Germany) was
used as a negative control. Reversine was purchased from Cayman
Chemical (Ann Arbor, MI, USA).
Cell proliferation assay
SW480 and HCT116 cells were seeded onto a 96-well
plate at a density of 1×104 cells/well and were
incubated for 24 h at 37°C. Cells were then treated with different
concentrations of reversine (0, 0.5, 1, 5, 10, and 20 µM)
with dimethyl sulfoxide (Sigma-Aldrich; Merck KGaA) for 24 and 48 h
at 37°C. Subsequently, the cells were treated with WST-1 reagent
(EZ-CYTOX; DoGen; Daeillab, Seoul, Korea) for 1 h at 37°C. After
the treatment, absorbance at 450 nm was measured using a microplate
reader (Infinite M200; Tecan Group Ltd., Mannedorf, Switzerland).
Each experiment was conducted in triplicate wells and was repeated
at least three times.
Calcein-acetomethoxy (AM) staining
SW480 and HCT116 cells were seeded onto a 96-well
plate at a density of 1×104 cells/well and incubated for
24 h at 37°C. Cells were then treated with different concentrations
of reversine (0, 0.5, 1, 5, 10 and 20 µM) with dimethyl
sulfoxide (Sigma-Aldrich; Merck KGaA) for 24 at 37°C. After the
reversine treatment, cells were stained with 1 µM Calcein-AM
solution (Invitrogen; Thermo Fisher Scientific, Inc.) for 60 min at
37°C. The cells were observed and imaged using a fluorescence
microscope (×200; DMI 3000B; Leica Microsystems GmbH, Wetzlar,
Germany).
Flow cytometric analysis
SW480 and HCT116 cells were seeded in a 6-well plate
at a density of 5×105 cells/well and were incubated for
24 h at 37°C prior to treatment with reversine. Subsequently, the
cells were treated with reversine (0, 1, 5 and 10 µM) for 24
at 37°C, and were resuspended in 100 µl of 1X binding buffer
(BD Biosciences, San Jose, CA, USA). These cells were incubated
with 7-amino-actinomycin D and Annexin V-APC (BD Biosciences) for
20 min at room temperature. To analyze the number of apoptotic
cells, a FACSCalibur flow cytometer (Becton-Dickinson and Company,
Franklin Lakes, NJ, USA) and WinMDI version 2.9 (The Scripps
Research Institute, San Diego, CA, USA) were used.
Cell cycle analysis
SW480 and HCT116 cells were seeded in a 6-well plate
at a density of 5×105 cells/well and were incubated for
24 h at 37°C. Subsequently, the cells were treated with reversine
(0 and 1 µM) for 24 h at 37°C, and were fixed with 70%
ethanol for 1 h at −20°C. These cells were resuspended in 100
µl of 1 U/ml RNase A (DNase free) for 20 min at room
temperature and incubated with 10 µg/ml of propidium iodide
(both from Sigma-Aldrich; Merck KGaA) for 30 min at room
temperature. To analyze the proportions of cell cycle, a
FACSCalibur flow cytometer and WinMDI version 2.9 were used.
DNA fragmentation
Reversine-treated cells were incubated with cell
lysate buffer (1% NP-40 in 20 mM EDTA, and 50 mM Tris-HCl; pH 7.5)
for 30 min on ice. The samples were then centrifuged at 12,000 × g
for 30 min at 4°C. RNase A was added to the supernatant and
incubated for 2 h at 56°C. Proteinase K was added to the
supernatant and the mixture was incubated for 2 h at 37°C. An equal
volume of isopropanol was added and the mixture was incubated at
-80°C overnight to precipitate the genomic DNA. The genomic DNA was
loaded onto 2% agarose gel and was stained with 0.1% ethidium
bromide for 10 min at 37°C. The DNA was visualized under
ultraviolet light transilluminator.
Western blotting
Reversine-treated cells were lysed in
Radioimmunoprecipitation Assay extraction solution with Halt™
Phosphatase inhibitor and Halt™ Protease inhibitor cocktails
(Thermo Fisher Scientific, Inc.) for 30 min on ice. The protein
concentration was measured using a Bicinchoninic Acid protein assay
(Thermo Fisher Scientific, Inc.). Equal amounts of 20 µg
proteins were separated using 10% SDS-PAGE and transferred to a
polyvinylidene fluoride membrane (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The membranes was blocked with 5% bovine serum
albumin (Sigma-Aldrich; Merck KGaA) for 1 h at room temperature and
then incubated overnight at 4°C with primary antibodies at 1:1,000
dilution. Primary antibodies against the following proteins were
used: Cleaved poly(ADP-ribose) polymerase (PARP; cat. no. 5625),
cleaved caspase-3 (cat. no. 9664), cleaved caspase-7 (cat. no.
8438), cleaved caspase-8 (cat. no. 9746), second
mitochondria-derived activator of caspase/direct inhibitor of
apoptosis-binding protein with low pI (SMAC/DIABLO; cat. no. 2954),
B-cell lymphoma-extra large (Bcl-xL; cat. no. 2764), Fas (cat. no.
4233), DR5 (cat. no. 8074), p21 (cat. no. 2947), p27 (cat. no.
2552) and p57 (cat. no. 2557) (Cell Signaling Technology, Inc.,
Danvers, MA, USA), and GAPDH (cat. no. FL-335; Santa Cruz
Biotechnology, Inc.). The membranes were washed four time with
TBS-0.1% Tween-20 and were incubated with a horseradish peroxidase
(HRP)-conjugated secondary antibody at 1:2,000 dilution for 1 h at
room temperature. The HRP-conjugated anti-rabbit (cat. no. 7074)
and anti-mouse (cat. no. 7076) secondary antibodies were purchased
from Cell Signaling Technology, Inc. Protein bands were developed
using an Enhanced Chemiluminescent reagent (GE Healthcare Life
Sciences, Little Chalfont, UK) and the luminescent image analyzer
LAS-4000 (Fujifilm, Tokyo, Japan) was used to analyze the
bands.
Statistical analysis
The results are presented as the mean ± standard
deviation. Differences between groups were determined by Student’s
t-test. The statistical software program used was SPSS (Version
18.0; SPSS, Inc., Chicago, IL, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
Reversine inhibits the growth of human
colorectal cancer cells
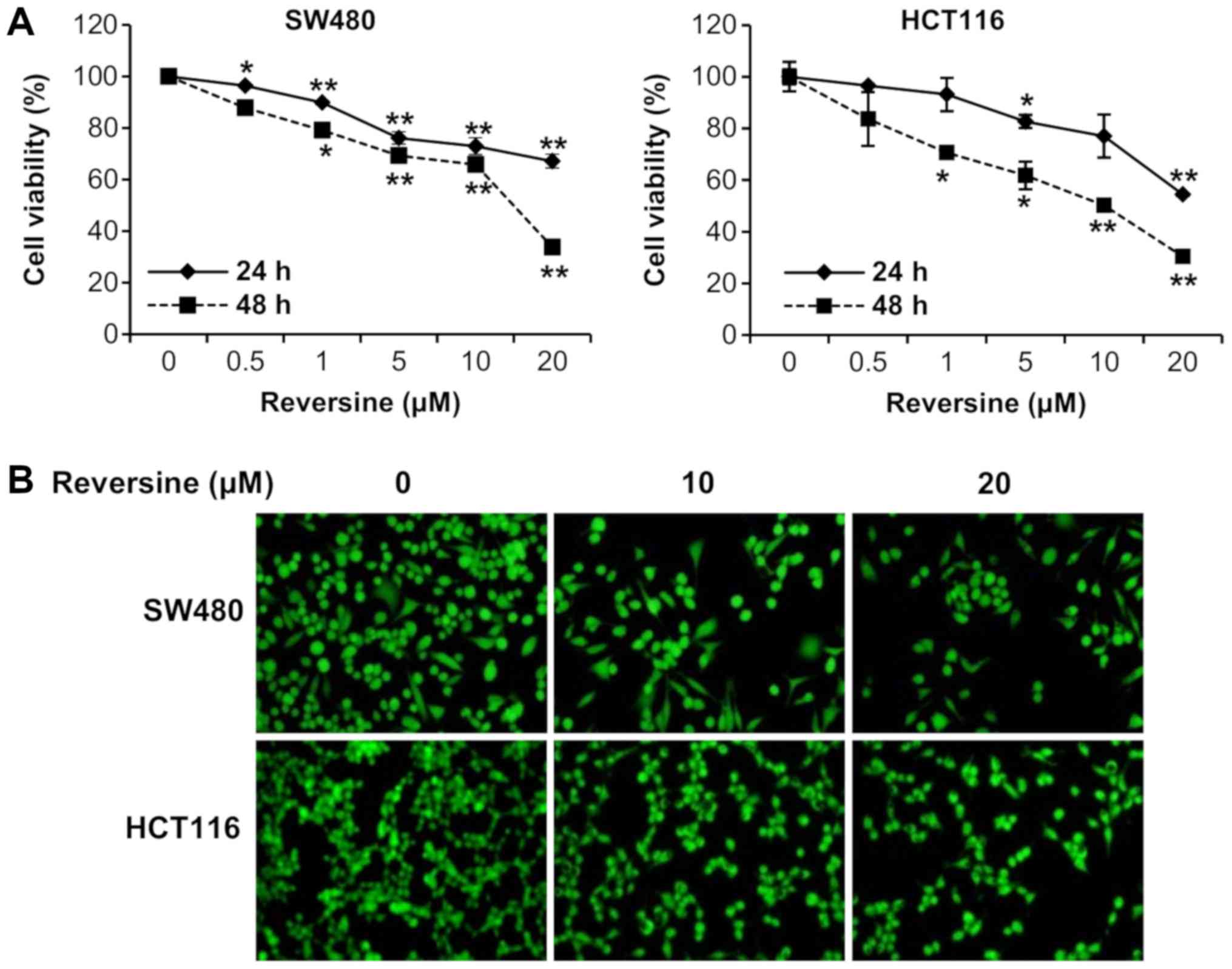
Firstly, the effects of reversine on the growth of
the human colorectal cancer cell lines SW480 and HCT116 were
investigated. The human colorectal cancer cells were exposed to
different concentrations of reversine (0-20 µM) for 24 and
48 h. The effects of reversine against the cells were determined
with a cell proliferation assay, followed by observation under
inverted microscopy. The cell proliferation assays and inverted
microscopy demonstrated that reversine treatment significantly
inhibited cell growth of SW480 and HCT116 cells, compared with the
control cells (P<0.05) (Fig. 1A and
B). These results indicated that reversine inhibits
proliferation in human colorectal cancer cells.
Reversine induces apoptosis and cell
cycle arrest in human colorectal cancer cells
In order to determine whether or not reversine
induced apoptosis and cell cycle arrest in human colorectal cancer
cells, flow cytometric analyses and a DNA fragmentation assay were
performed. Human colorectal cancer cells were exposed to different
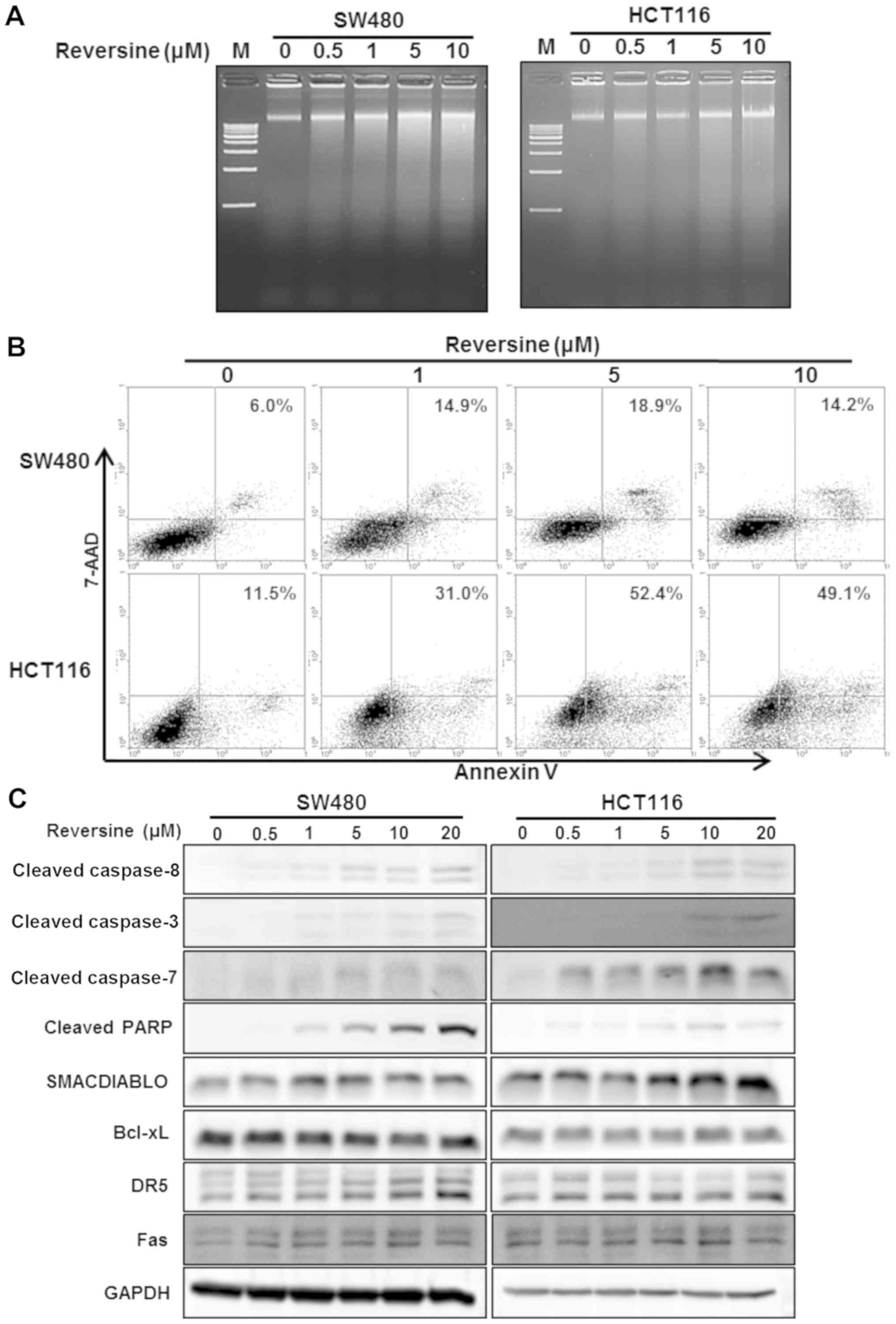
concentrations of reversine (0-20 µM) for 24 h. Reversine
treatment (0.5-10 µM) induced an increase in DNA
fragmentation in SW480 and HCT116 cells, compared with non-treated
control (Fig. 2A). The proportion
of early and late apoptotic cells within the group treated with
reversine was increased, compared with the non-treated control, in
SW480 and HCT116 cells (Fig. 2B).
To determine the activation of caspases, key enzymes of apoptosis,
the caspase-specific activities were further investigated. The
expression of cleaved PARP, caspase-3, -7 and -8 was upregulated in
the SW480 and HCT116 cells following reversine treatment, in a
dose-dependent manner (Fig. 2C).
Additionally, whether reversine treatment resulted in modulation of
apoptosis-regulatory proteins was further examined. As depicted in
Fig. 2C, reversine treatment
resulted in an increase in the levels of pro-apoptotic protein
SMAC/DIABLO in SW480 and HCT116 cells, but the levels of the
anti-apoptotic protein Bcl-xL were not altered in response to
reversine treatment. In addition, the effect of reversine on death
receptor signaling pathways was investigated, and it was observed
that the expression of DR5 and Fas proteins was upregulated by
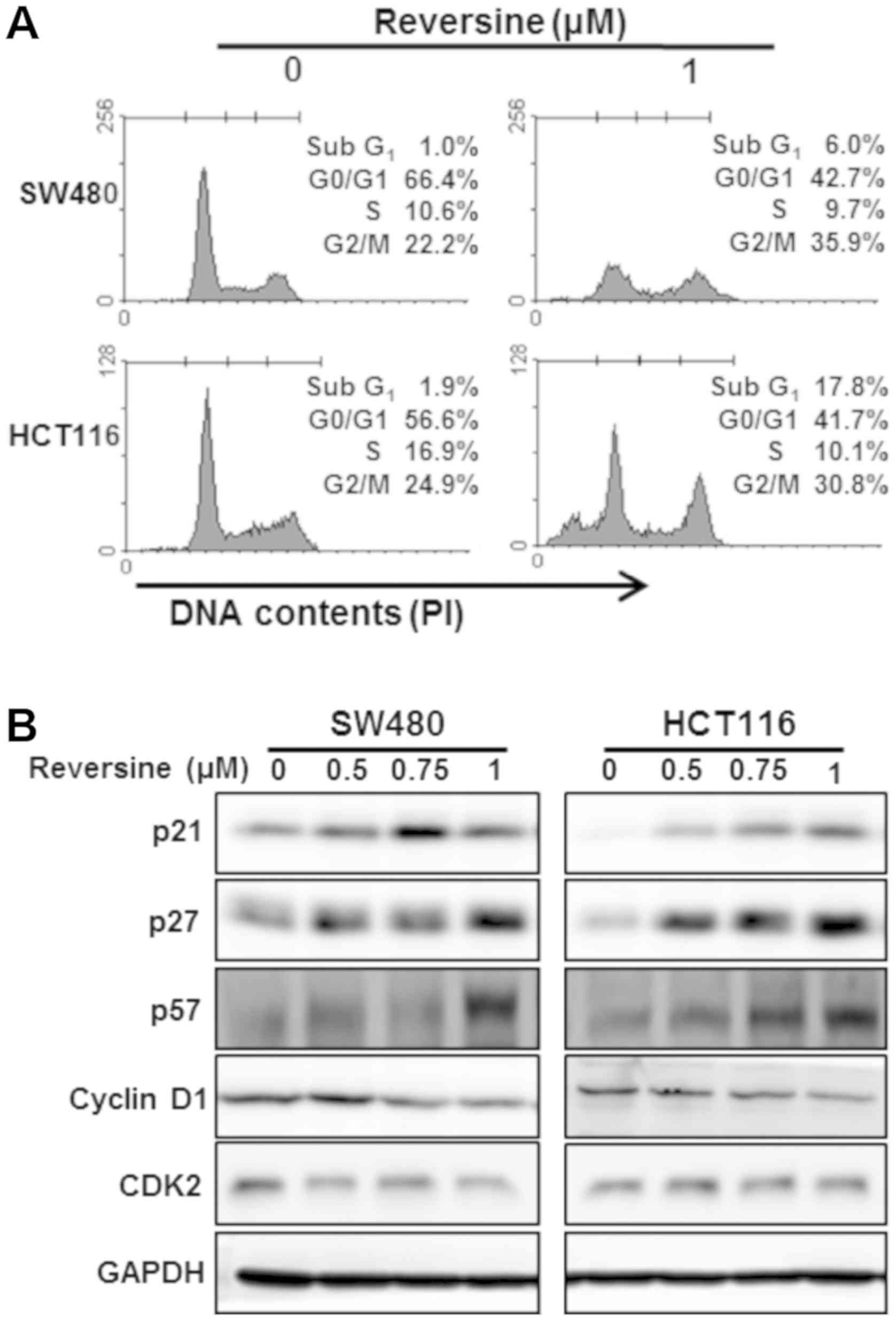
reversine treatment in SW480 and HCT116 cells (Fig. 2C). Reversine treatment resulted in
cell cycle arrest in the subG1 and G2/M phase of SW480 and HCT116
cells (Fig. 3A). Subsequently, the
effects of reversine treatment on various CDK inhibitors (CDKIs)
involved in cell cycle arrest in human colorectal cancer cells were
evaluated. The p21, p27 and p57 protein levels were notably
increased by reversine treatment in SW480 and HCT116 cells
(Fig. 3B). Cyclins and CDKs were
negatively regulated by CDKIs. Additionally, we examined the
effects of reversine on the expression levels of cyclin D1 and
CDK2. The cyclin D1 protein level was notably decreased by
reversine treatment in SW480 and HCT116 cells. The CDK2 protein
level was notably decreased in SW480 cells but was not altered in
response to reversine treatment in HCT116 cells (Fig. 3B).
Pan-caspase inhibitor attenuates the
reversine-induced inhibition of cell growth and induction of
apoptosis in human colorectal cancer cells
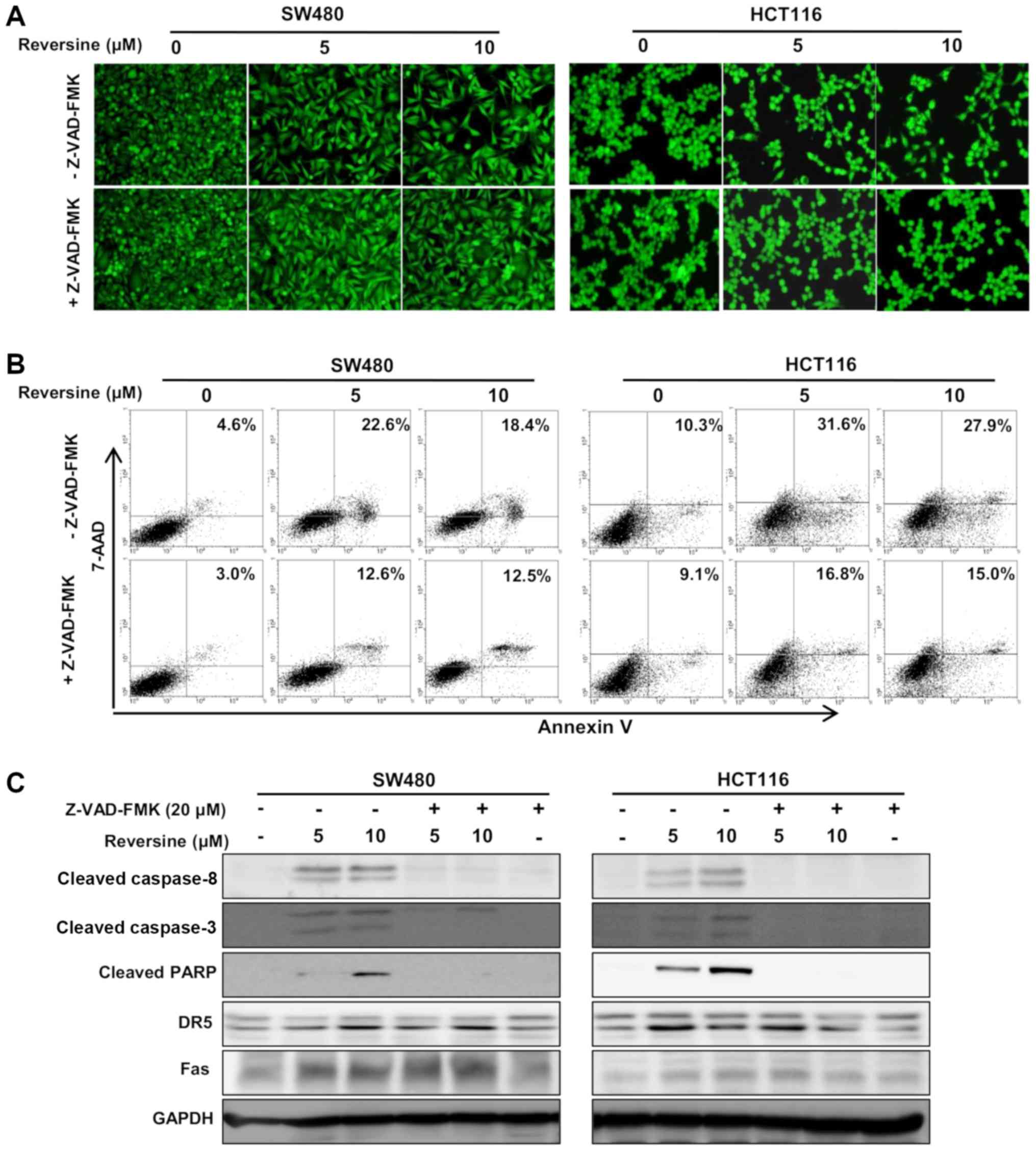
The pan-caspase inhibitor Z-VAD-FMK (20 µM)
was used to determine whether cell growth was inhibited and
apoptosis was induced by reversine treatment. The inhibition of
cell growth by reversine treatment was reversed in SW480 and HCT116
cells by Z-VAD-FMK treatment (Fig.
4A). The increase in the number of early and late apoptotic
cells by reversine treatment was also attenuated in SW480 and
HCT116 cells by Z-VAD-FMK treatment (Fig. 4B). Furthermore, Z-VAD-FMK abrogated
the reversine-induced PARP, and caspase-3 and -8 activation.
However, Z-VAD-FMK did not notably affect reversine-induced DR5 and
Fas protein expression (Fig. 4C).
Therefore, reversine treatment exerts inhibition of cell growth and
promotion of apoptosis via direct activation of caspases in human
colorectal cancer cells.
Impact of reversine on Fas and DR5
signaling pathways in human colorectal cancer cells
To confirm whether Fas and DR5 signaling pathways
are directly upregulated by reversine treatment, SW480 and HCT116
cells were transfected with Fas and DR5 siRNAs for 24 h, and then 5
µM reversine was added. To determine whether activation of
Fas and DR5 is associated with reversine-induced cell apoptosis,
flow cytometry was conducted. The increase in the number of early
and late apoptotic cells by reversine treatment was attenuated
following inhibition of Fas and DR5 by siRNA in SW480 and HCT116
cells (Fig. 5A). Inhibition of Fas
and DR5 by siRNA suppressed reversine-induced cell cycle arrest of
the subG1 and G2/M phase (Fig.
5B). Inhibition of Fas and DR5 by siRNAs suppressed
reversine-induced upregulation of the Fas and DR5 proteins.
Reversine-induced caspase-8 and -3, and PARP activation was
attenuated following inhibition of Fas and DR5 by siRNA (Fig. 5C). These results indicated that
reversine-induced apoptosis and cell cycle arrest in human
colorectal cancer cells is mediated by activation of the Fas and
DR5 signaling pathways.
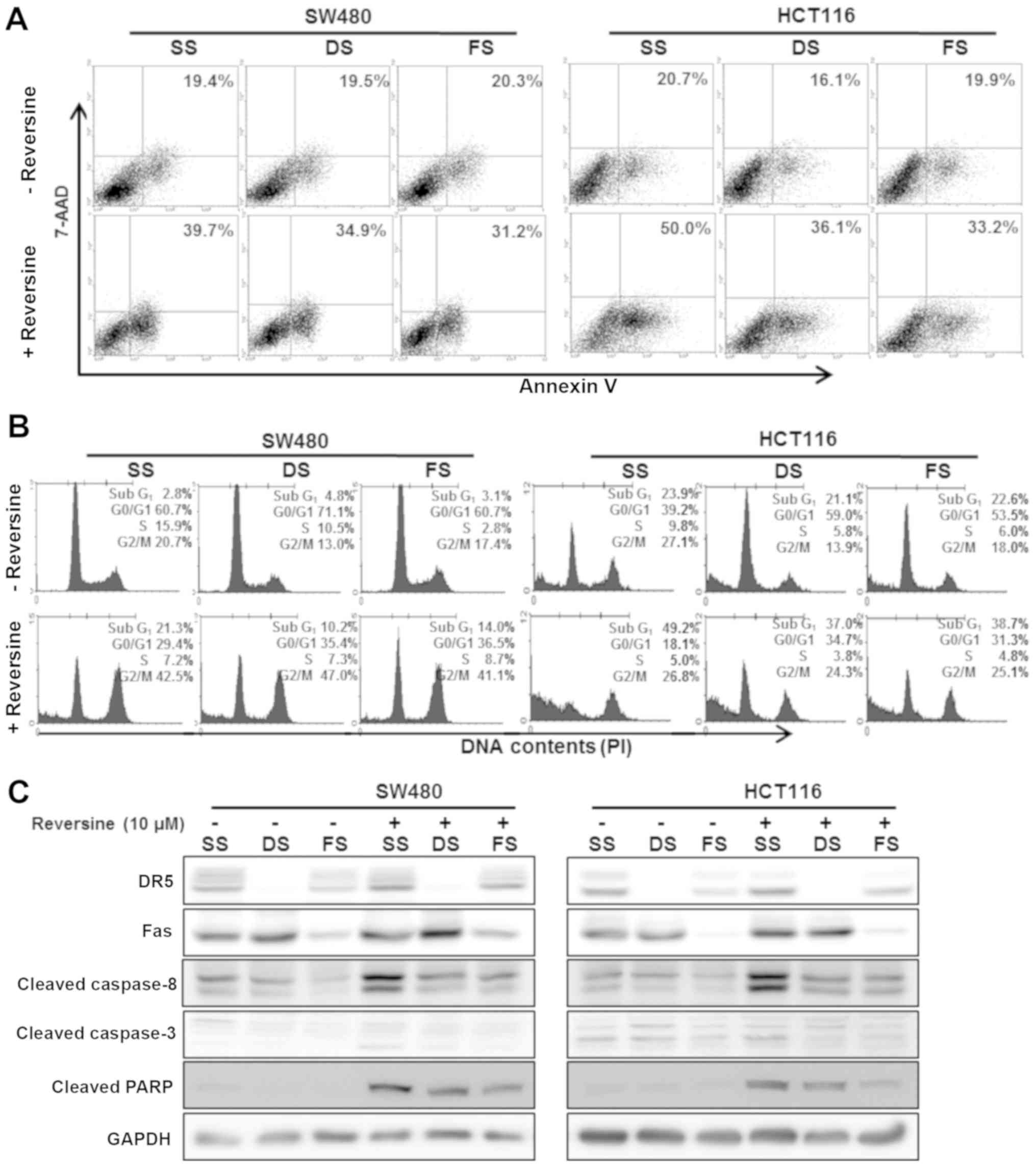 | Figure 5Impact of reversine on Fas and DR5
signaling pathways in human colorectal cancer cells. SW480 and
HCT116 cells were transfected with Fas and DR5 siRNA for 24 h and
then exposed to the indicated concentrations of reversine for 24 h.
(A) Treated cells were stained with Annexin V and the number of
apoptotic cells was counted. (B) Treated cells were stained with PI
and subjected to flow cytometric analysis to determine the cell
distribution at each phase of cell cycle. (C) Protein levels were
detected by western blotting. GAPDH was used as a loading control.
siRNA, small interfering RNA; SS, scrambled siRNA, DS, death
receptor 5 siRNA; FS, Fas siRNA; Cas, caspase; PARP,
poly(ADP-ribose) polymerase; DR5, death receptor 5; PI, propidium
iodide; 7-AAD, 7-amino-actinomycin D. |
Discussion
Reversine, a synthetic purine analogue, induces
dedifferentiation of murine myoblasts to multipotent progenitor
cells, which can redifferentiate into other different cell types
(3-6). Previously, reversine has been
demonstrated to exert anticancer effects via induction of
apoptosis, cell cycle arrest and autophagy in numerous human cancer
types, including breast, lung and thyroid cancer (10-23).
Thus, the impact of reversine on tumor cell behavior and its
association with intracellular signaling pathways in human
colorectal cancer cells were investigated.
Apoptosis is an essential mechanism for cell
morphogenesis, cell turnover and removal of damaged cells. Balance
between cell growth and apoptosis is crucial to ensure normal
development and homeostasis (24-26).
However, deregulation of cell growth and apoptosis is responsible
for cancer development, progression and resistance to treatment
(24-26). In the present study, reversine
inhibited proliferation and induced apoptosis in human colorectal
cancer cells. Recently, reversine was demonstrated to suppress the
migration and invasion of human colorectal cancer RKO cells by
inhibiting the c-Jun NH2-terminal kinase signaling pathway, which
serves a crucial role in cell proliferation, cell cycle progression
and apoptosis (21). These results
indicated that reversine may contribute to altering the invasive
and oncogenic phenotypes of human colorectal cancer cells.
Apoptosis involves the activation of caspases, which
participate in enzymatic cascades that terminate in cellular
disassembly, and is a complex process that results from multiple
genetic alterations in pro- and anti-apoptotic genes. Thus, the
effects of reversine on apoptosis in human colorectal cancer cells
were investigated. Reversine treatment induced apoptosis by
activating PARP, and caspase-3, -7 and -8, and enhancing the levels
of the pro-apoptotic protein SMAC/DIABLO. Furthermore, the
pan-caspase inhibitor was used to determine the association between
reversine-induced apoptosis and caspase activation. The present
study demonstrated that the pan-caspase inhibitor abrogated the
reversine-induced increase in the number of early and late
apoptotic cells, and caspase-3, -8 and PARP activation. Therefore,
reversine treatment induces apoptosis by directly inducing caspase
activity in human colorectal cancer cells. These results were
consistent with those of previous studies on other human cancer
types (10-23).
Cell cycle, cell proliferation and cell apoptosis
are associated with the circadian clock, which controls the rhythms
of various physiological processes, including cell division, cell
migration and metabolism (27,28).
The disruption of the circadian clock is associated with cancer
development and progression (27,28).
Recently, reversine and reversine-associated molecules were
indicated to cause cell cycle arrest in the G2/M phase and
polyploidy in the HCT116 colorectal cancer cell line (14). In the present study, reversine
treatment induced cell cycle arrest at the subG1 and G2/M phases of
human colorectal cancer cells.
The cell cycle progression is regulated by a complex
system involving cyclins, CDKs and CDKIs. CDKs are serine/threonine
kinases that are modulated by interaction with positive effectors,
cyclins and negative regulators, CDKIs. Until now, two families of
CDKIs have been identified: The Cip/Kip family; and the INK family
(27-29). The Cip/Kip family includes p21, p27
and p57. The INK family includes p16, p15, p18 and p19 (27-29).
In the present study, reversine treatment induced cell cycle arrest
in the subG1 and G2/M phases via the upregulation of p21, p27 and
p57, and the downregulation of cyclin D1. Previously, reversine
induced cell growth arrest and polyploidy by upregulating p21,
cyclin D3 and CDK6, while downregulating cyclin B1 and CDK1
(7). Among the numerous reported
CDKIs, aberrant expression of p21, p27 and p57 has been
demonstrated in various human cancer cells and their downregulation
has been associated with tumor progression in various human cancer
types, including pancreas, breast, lung and colorectal cancer,
indicating their role as tumor suppressors (30-32).
Therefore, reversine may contribute to inhibit colorectal cancer
progression via cell cycle arrest by upregulation of members of the
Cip/Kip family.
The Fas/FasL and DR5 signaling pathways are the
major pathways that regulate apoptosis (33-35).
DR5 is located on the surface of cancer cells, and stimulation of
this death receptor activates the caspase-8-dependent apoptotic
pathway (33-35). Fas-associated DR5 protein and
caspase-8 are essential for anticancer agent-induced cell apoptosis
in numerous human cancer types, including gastric and colorectal
cancer (36-38). In the present study, the expression
of Fas, DR5, caspase-8 and -3, and PARP proteins was upregulated by
reversine treatment in human colorectal cancer cells.
Reversine-induced apoptosis and cell cycle arrest were inhibited by
inhibition of Fas and DR5 via siRNA. These results indicated that
reversine induced caspase-dependent apoptosis and cell cycle arrest
by activation of the Fas and DR5 signaling pathways.
Overall, in the present study, reversine treatment
suppressed tumor progression by inhibition of cell proliferation,
induction of apoptosis and cell cycle arrest through upregulation
of the Fas and DR5 signaling pathways in human colorectal cancer
cells. The present study indicated that reversine may be used as a
novel anticancer agent in human colorectal cancer.
Funding
This study was supported by research funds from the
Research Institute of Clinical Medicine, Chonnam National
University Hwasun Hospital in 2017 (grant no. HCRI 17912-1),
Republic of Korea, and partly the National Research Foundation of
Korea grant (grant no. NRF-2017R1A2B4004703) funded by the Korean
government (Ministry of Science, ICT and Future Planning), Republic
of Korea.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors’ contributions
YEJ was involved in the conceptualization of the
study. YLP, SYH, SYP, JHC, MWJ, DSM and HSK were involved in data
curation, were responsible for formal analysis and provided
resources. YLP, SYH and YEJ were involved in the investigative part
of the study, were involved in the methodology and were involved in
the writing, reviewing and editing of the manuscript. YEJ was
involved in project administration, and was involved in the writing
of the manuscript and original draft preparation.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014.
|
|
2
|
Choi Y, Sateia HF, Peairs KS and Stewart
RW: Screening for colorectal cancer. Semin Oncol. 44:34–44.
2017.
|
|
3
|
Chen S, Zhang Q, Wu X, Schultz PG and Ding
S: Dedifferentiation of lineage-committed cells by a small
molecule. J Am Chem Soc. 126:410–411. 2004.
|
|
4
|
Chen S, Takanashi S, Zhang Q, Xiong W, Zhu
S, Peters EC, Ding S and Schultz PG: Reversine increases the
plasticity of lineage-committed mammalian cells. Proc Natl Acad Sci
USA. 104:10482–10487. 2007.
|
|
5
|
Lee EK, Bae GU, You JS, Lee JC, Jeon YJ,
Park JW, Park JH, Ahn SH, Kim YK, Choi WS, et al: Reversine
increases the plasticity of lineage-committed cells toward
neuroectodermal lineage. J Biol Chem. 284:2891–2901. 2009.
|
|
6
|
Pikir BS, Susilowati H, Hendrianto E and
Abdulrantam F: Reversin increase the plasticity of bone
marrow-derived mesenchymal stem cell for generation of
cardiomyocyte in vitro. Acta Med Indones. 44:23–27. 2012.
|
|
7
|
Yan M, Wang C, He B, Yang M, Tong M, Long
Z, Liu B, Peng F, Xu L, Zhang Y, et al: Aurora-A Kinase: A Potent
Oncogene and Target for Cancer Therapy. Med Res Rev. 36:1036–1079.
2016.
|
|
8
|
Tang A, Gao K, Chu L, Zhang R, Yang J and
Zheng J: Aurora kinases: Novel therapy targets in cancers.
Oncotarget. 8:23937–23954. 2017.
|
|
9
|
Malumbres M and Pérez de Castro I: Aurora
kinase A inhibitors: Promising agents in antitumoral therapy.
Expert Opin Ther Targets. 18:1377–1393. 2014.
|
|
10
|
Hsieh TC, Traganos F, Darzynkiewicz Z and
Wu JM: The 2,6-disubstituted purine reversine induces growth arrest
and polyploidy in human cancer cells. Int J Oncol. 31:1293–1300.
2007.
|
|
11
|
Lu CH, Liu YW, Hua SC, Yu HI, Chang YP and
Lee YR: Autophagy induction of reversine on human follicular
thyroid cancer cells. Biomed Pharmacother. 66:642–647. 2012.
|
|
12
|
Piccoli M, Palazzolo G, Conforti E,
Lamorte G, Papini N, Creo P, Fania C, Scaringi R, Bergante S,
Tringali C, et al: The synthetic purine reversine selectively
induces cell death of cancer cells. J Cell Biochem. 113:3207–3217.
2012.
|
|
13
|
Kuo CH, Lu YC, Tseng YS, Shi CS, Chen SH,
Chen PT, Wu FL, Chang YP and Lee YR: Reversine induces cell cycle
arrest, polyploidy, and apoptosis in human breast cancer cells.
Breast Cancer. 21:358–369. 2014.
|
|
14
|
Bosco B, Defant A, Messina A, Incitti T,
Sighel D, Bozza A, Ciribilli Y, Inga A, Casarosa S and Mancini I:
Synthesis of 2,6-diamino-substituted purine derivatives and
evaluation of cell cycle arrest in breast and colorectal cancer
cells. Molecules. 23:E19962018.
|
|
15
|
Cheng L, Wang H, Guo K, Wang Z, Zhang Z,
Shen C, Chen L and Lin J: Reversine, a substituted purine, exerts
an inhibitive effect on human renal carcinoma cells via induction
of cell apoptosis and polyploidy. OncoTargets Ther. 11:1025–1035.
2018.
|
|
16
|
Qin HX, Yang J, Cui HK, Li SP, Zhang W,
Ding XL and Xia YH: Synergistic antitumor activity of reversine
combined with aspirin in cervical carcinoma in vitro and in vivo.
Cytotechnology. 65:643–653. 2013.
|
|
17
|
Rodrigues Alves AP, Machado-Neto JA,
Scheucher PS, Paiva HH, Simões BP, Rego EM and Traina F: Reversine
triggers mitotic catastrophe and apoptosis in K562 cells. Leuk Res.
48:26–31. 2016.
|
|
18
|
Lu YC, Lee YR, Liao JD, Lin CY, Chen YY,
Chen PT and Tseng YS: Reversine induced multinucleated cells, cell
apoptosis and autophagy in human non-small cell lung cancer cells.
PLoS One. 11:e01585872016.
|
|
19
|
Lee YR, Wu WC, Ji WT, Chen JY, Cheng YP,
Chiang MK and Chen HR: Reversine suppresses oral squamous cell
carcinoma via cell cycle arrest and concomitantly apoptosis and
autophagy. J Biomed Sci. 19:92012.
|
|
20
|
D’Alise AM, Amabile G, Iovino M, Di
Giorgio FP, Bartiromo M, Sessa F, Villa F, Musacchio A and Cortese
R: Reversine, a novel Aurora kinases inhibitor, inhibits colony
formation of human acute myeloid leukemia cells. Mol Cancer Ther.
7:1140–1149. 2008.
|
|
21
|
Jemaà M, Abassi Y, Kifagi C, Fezai M,
Daams R, Lang F and Massoumi R: Reversine inhibits Colon Carcinoma
Cell Migration by Targeting JNK1. Sci Rep. 8:118212018.
|
|
22
|
Hua SC, Chang TC, Chen HR, Lu CH, Liu YW,
Chen SH, Yu HI, Chang YP and Lee YR: Reversine, a 2,6-disubstituted
purine, as an anti-cancer agent in differentiated and
undifferentiated thyroid cancer cells. Pharm Res. 29:1990–2005.
2012.
|
|
23
|
Bijian K, Lougheed C, Su J, Xu B, Yu H, Wu
JH, Riccio K and Alaoui-Jamali MA: Targeting focal adhesion
turnover in invasive breast cancer cells by the purine derivative
reversine. Br J Cancer. 109:2810–2818. 2013.
|
|
24
|
Kiechle FL and Zhang X: Apoptosis:
Biochemical aspects and clinical implications. Clin Chim Acta.
326:27–45. 2002.
|
|
25
|
Schultz DR and Harrington WJ Jr:
Apoptosis: Programmed cell death at a molecular level. Semin
Arthritis Rheum. 32:345–369. 2003.
|
|
26
|
Llambi F and Green DR: Apoptosis and
oncogenesis: Give and take in the BCL-2 family. Curr Opin Genet
Dev. 21:12–20. 2011.
|
|
27
|
Lim S and Kaldis P: Cdks, cyclins and
CKIs: Roles beyond cell cycle regulation. Development.
140:3079–3093. 2013.
|
|
28
|
Soták M, Sumová A and Pácha J: Cross-talk
between the circadian clock and the cell cycle in cancer. Ann Med.
46:221–232. 2014.
|
|
29
|
Bonelli P, Tuccillo FM, Borrelli A,
Schiattarella A and Buonaguro FM: CDK/CCN and CDKI alterations for
cancer prognosis and therapeutic predictivity. BioMed Res Int.
2014:3610202014.
|
|
30
|
Wang K, Baldwin GS, Nikfarjam M and He H:
p21-activated kinase signalling in pancreatic cancer: New insights
into tumour biology and immune modulation. World J Gastroenterol.
24:3709–3723. 2018.
|
|
31
|
Bencivenga D, Caldarelli I, Stampone E,
Mancini FP, Balestrieri ML, Della Ragione F and Borriello A:
p27Kip1 and human cancers: A reappraisal of a still enigmatic
protein. Cancer Lett. 403:354–365. 2017.
|
|
32
|
Kobatake T, Yano M, Toyooka S, Tsukuda K,
Dote H, Kikuchi T, Toyota M, Ouchida M, Aoe M, Date H, et al:
Aberrant methylation of p57KIP2 gene in lung and breast cancers and
malignant mesotheliomas. Oncol Rep. 12:1087–1092. 2004.
|
|
33
|
Villa-Morales M and Fernández-Piqueras J:
Targeting the Fas/FasL signaling pathway in cancer therapy. Expert
Opin Ther Targets. 16:85–101. 2012.
|
|
34
|
Mocellin S: Targeting death receptors to
fight cancer: From biological rational to clinical implementation.
Curr Med Chem. 17:2713–2728. 2010.
|
|
35
|
Tschumi BO, Dumauthioz N, Marti B, Zhang
L, Schneider P, Mach JP, Romero P and Donda A: CART cells are prone
to Fas- and DR5-mediated cell death. J Immunother Cancer.
6:712018.
|
|
36
|
Wang X, Xue Q, Wu L, Wang B and Liang H:
Dasatinib promotes TRAIL-mediated apoptosis by upregulating
CHOP-dependent death receptor 5 in gastric cancer. FEBS Open Bio.
8:732–742. 2018.
|
|
37
|
Shlyakhtina Y, Pavet V and Gronemeyer H:
Dual role of DR5 in death and survival signaling leads to TRAIL
resistance in cancer cells. Cell Death Dis. 8:e30252017.
|
|
38
|
Karbasi A, Borhani N, Daliri K, Kazemi B
and Manoochehri M: Downregulation of external death receptor genes
FAS and DR5 in colorectal cancer samples positive for human
papillomavirus infection. Pathol Res Pract. 211:444–448. 2015.
|