Introduction
Renal cell carcinoma (RCC) is a common type of
cancer worldwide and the incidence of this disease is increasing;
the incidence rate of RCC was 10.6/100,000 in 2001, which increased
to 12.4/100,000 in 2010 (1,2).
Genetic mutations are an important risk factor for the development
of RCC (3), which can be detected
at early stages. Systemic therapeutic agents, including sunitinib
and temsirolimus, have been used to treat RCC (4). The presence of smaller tumors leads
to fewer episodes of relapse and improved prognosis (5). Further investigation is required to
determine the molecular mechanisms underlying the development of
clear cell RCC (ccRCC), the most common type of RCC.
The phosphoinositide 3-kinase (PI3K)/protein kinase
B (Akt) signaling pathway regulates a series of cellular processes,
including the cell cycle, proliferation, apoptosis, autophagy and
metastasis in various types of cancer (6). The PI3K/Akt signaling pathway serves
its role by regulating numerous effectors, particularly factors of
the mitogen-activated protein kinase kinase (MEK)/extracellular
signal-regulated kinase (ERK) pathway (7). Due to its key roles in cancer,
several potential molecular targets of the PI3K/Akt signaling
pathway have been proposed in cancer therapy (8). Furthermore, the PI3K/Akt signaling
pathway has been demonstrated to be highly activated and serves a
critical role in ccRCC (9,10).
Homeobox A6 (HOXA6) encodes a transcription factor
that regulates gene expression and cellular differentiation. HOXA6
has been identified to serve important roles in cell proliferation,
differentiation and invasion (11,12).
It has been reported that HOXA6 may serve a role in a variety of
cancer types; however, the roles of HOXA6 in these cancer types
were inconsistent. HOXA6 was observed to be overexpressed in tumor
tissues and promoted the progression of cancer (13-15);
however, HOXA6 was also determined to be hypermethylated in certain
malignancies, indicating reduced expression (16,17).
In addition, HOXA6 expression has been reported to be suppressed in
breast cancer tissues (18). At
present, to the best of our knowledge, extensive research into the
roles of HOXA6 in ccRCC has not been conducted. Therefore, the
present study aimed to determine the expression profile of HOXA6 in
ccRCC. The results revealed that HOXA6 was downregulated in ccRCC;
however, the role of HOXA6 in ccRCC and its underlying mechanism
requires further investigation. In the present study, the role of
HOXA6 in cell proliferation and apoptosis was analyzed, and the
association between HOXA6 and the PI3K/Akt signaling pathway in
ccRCC was determined.
Materials and methods
Confirmation of HOXA6 expression and
survival analysis
Data of HOXA6 mRNA expression was obtained from the
GSE6344 dataset (19,20) of the Gene Expression Omnibus (GEO)
database (https://www.ncbi.nlm.nih.gov/geo/). The tissue samples
included in GSE6344 consisted of ten patient-matched normal renal
cortex and ccRCC tissues; five samples were of stage I and five
were of stage II ccRCC, according to the TNM staging system
(21). All patients were diagnosed
with localized disease; stage I tumors were <7 cm and stage II
tumors were >7 cm. Gene Expression Profiling Interactive
Analysis (GEPIA) (22) was
performed to determine the association between the mRNA expression
levels of HOXA6 and the survival of patients with ccRCC.
Tissue specimens
The present study was approved by the Ethics
Committee of The First Affiliated Hospital of Chongqing Medical
University (Chongqing, China; approval no. 20185501). Written
informed consent was obtained from all patients. A total of 25
pairs of ccRCC and adjacent non-tumor tissue specimens were
collected at the Department of Urology between June 2018 and August
2018. The distance between the ccRCC sample and the non-tumor
tissue sample was ≥2 cm. Among the 25 patients, 14 patients
underwent radical nephrectomy and 11 underwent partial nephrectomy
without any pre-operative treatment. The median age of the patients
was 53 years (range, 42-78 years). The clinicopathological features
of the patients are presented in Table
I. Pathological diagnosis of the specimens was based on the
pathological report of the Department of Pathology of Chongqing
Medical University.
 | Table IClinicopathological characteristics
of patients with ccRCC. |
Table I
Clinicopathological characteristics
of patients with ccRCC.
| Case no. | Age, years | Sex | Diagnosis | TNM stage | Tumor size, cm |
|---|
| 1 | 52 | Male | Right renal
ccRCC | T1aN0M0 | 2.7 |
| 2 | 66 | Male | Left renal
ccRCC | T2aN0M0 | 7.2 |
| 3 | 63 | Male | Right renal
ccRCC | T1aN0M0 | 2.4 |
| 4 | 60 | Male | Left renal
ccRCC | T1aN0M0 | 3.1 |
| 5 | 66 | Female | Right renal
ccRCC | T1aN0M0 | 2.6 |
| 6 | 74 | Male | Left renal
ccRCC | T1aN0M0 | 2.5 |
| 7 | 47 | Female | Left renal
ccRCC | T1aN0M0 | 3.2 |
| 8 | 68 | Male | Right renal
ccRCC | T1aN0M0 | 2.7 |
| 9 | 78 | Male | Right renal
ccRCC | T1aN0M0 | 2.0 |
| 10 | 67 | Male | Right renal
ccRCC | T2aN0M0 | 7.1 |
| 11 | 72 | Female | Left renal
ccRCC | T1aN0M0 | 2.1 |
| 12 | 49 | Male | Right renal
ccRCC | T1aN0M0 | 2.5 |
| 13 | 47 | Male | Left renal
ccRCC | T2aN0M0 | 8.6 |
| 14 | 44 | Male | Left renal
ccRCC | T1bN0M0 | 4.7 |
| 15 | 53 | Female | Left renal
ccRCC | T3aN0M0 | 4.0 |
| 16 | 64 | Male | Left renal
ccRCC | T3aN0M0 | 5.0 |
| 17 | 50 | Male | Left renal
ccRCC | T1bN0M0 | 5.0 |
| 18 | 47 | Male | Right renal
ccRCC | T1bN0M0 | 7.0 |
| 19 | 50 | Female | Left renal
ccRCC | T1aN0M0 | 3.8 |
| 20 | 50 | Female | Left renal
ccRCC | T1bN0M0 | 4.5 |
| 21 | 62 | Male | Left renal
ccRCC | T1bN0M0 | 6.5 |
| 22 | 43 | Male | Left renal
ccRCC | T1aN0M0 | 2.5 |
| 23 | 42 | Male | Left renal
ccRCC | T1aN0M0 | 3.0 |
| 24 | 53 | Male | Left renal
ccRCC | T1bN0M0 | 5.0 |
| 25 | 66 | Female | Right renal
ccRCC | T3aN0M0 | 3.5 |
Immunohistochemistry (IHC)
Tissues were fixed with 10% formalin at room
temperature for 4-6 h. Subsequently, paraffin-embedded tissue
sections (5-µm) were dewaxed and rehydrated in 100, 95, 85,
75 and 50% ethanol, boiled at 100°C in 0.01 mol/l sodium citrate
buffer (pH 6.0) for 15 min for antigen retrieval and incubated with
3% hydrogen peroxide for 20 min to inhibit endogenous peroxidase
activity. Samples were blocked with normal goat serum (reagent B in
the SP-9001 kit; OriGene Technologies, Inc.) at 37°C for 30 min and
incubated with rabbit anti-HOXA6 (cat. no. bs-11294R, 1:400; BIOSS)
overnight at 4°C. A horseradish peroxidase (HRP)-conjugated goat
anti-rabbit IgG working solution (reagent C in the SP-9001 kit;
OriGene Technologies, Inc.) was then applied for 30 min at 37°C.
Subsequently, sections were stained at room temperature with
3,3′-diaminobenzidene (OriGene Technologies, Inc.) for 10 sec and
hematoxylin (OriGene Technologies, Inc.) for 30 sec at room
temperature. Negative control specimens were incubated without
anti-HOXA6.
The score criterion was set according to the
intensity and the extent of section staining (percentage of
positive cells). The intensity scores were as follows: 0, no
staining; 1, weak; 2, moderate; and 3, intense. The scores for the
extent of staining were as follows: 0, 0%; 1, 1-25%; 2, 26-50%; 3,
51-75%; and 4, 76-100%. The IHC scores were calculated by
multiplying the intensity scores with the extent of staining scores
(0-12). IHC scores <3 were defined as low expression, while
scores ≥4 were defined as high expression. A light microscope was
used to capture the images (magnification, ×200 and ×400). The
ccRCC subtype and IHC scores were confirmed independently by two
pathologists.
Cell culture and transfection
The human ccRCC cell lines, 786-O and 769-P, were
purchased from the American Type Culture Collection and cultured in
RPMI-1640 medium (HyClone; GE Healthcare) containing 10% fetal
bovine serum (PAN-Biotech) at 37°C in 5% CO2.
The plasmids were supplied by GeneCopoeia, Inc.,
including HOXA6-expressing plasmid, empty vector (termed vector;
pReceiver-M03), and plasmids containing short hairpin (sh)HOXA6
(5′-CCTTGTTTCTACCAACAGTCC-3′) and non-targeting shRNA (NC;
5′-GCTTCGCGCCGTAGT CTTA-3′). 786-O and 769-P cells were transfected
using Lipofectamine 2000® (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer’s protocol. Cells
were seeded into a 6-well plate at a density of ~2×105
cells per well, and a total of 4 µg plasmids were added for
transfection at 50-75% confluence.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
At 48 h post-transfection, total RNA was extracted
from cells using TRIzol® reagent (Thermo Fisher
Scientific, Inc.) and reverse transcribed with the GoScript™
Reverse Transcription system (Promega Corporation) at 25°C for 5
min, 42°C for 60 min, 70°C for 15 min and 4°C until the subsequent
step. The primers used for qPCR were as follows (15): HOXA6 forward, 5′-TACACGCGCTACCAGA
CAC-3′ and reverse, 5′-GCGTGGAATTGATGAGCTTG TTT-3′, and β-actin
forward, 5′-TGGAACGGTGAAGGTGA CAG-3′ and reverse,
5′-AACAACGCATCTCATATTTG GAA-3′. qPCR was performed using the GoTaq
qPCR Master mix (Promega Corporation), with the following
thermocycling conditions: 95°C for 10 min, 40 cycles of 95°C for 15
sec and 60°C for 1 min, followed by 95°C for 15 sec, 60°C for 1 min
and 95°C for 15 sec, according to the manufacturer’s protocol. Data
were analyzed using the 2−∆∆Cq method (23).
Western blotting
At 72 h post-transfection, total proteins were
extracted from cells using radioimmunoprecipitation assay lysis
buffer (Beyotime Institute of Biotechnology) and the protein
concentration was measured via the bicinchoninic acid method.
Proteins (40 µg) were separated by 10% SDS-PAGE and
transferred to a polyvinylidene difluoride (PVDF) membrane.
Subsequently, the PVDF membrane was blocked with 5% skim milk at
room temperature for 2 h and incubated with primary antibodies at
4°C overnight. The membrane was then incubated with an
HRP-conjugated secondary antibody (1:3,000, cat. no. bs-0295G;
BIOSS) at 37°C for 2 h. Bands were visualized using an enhanced
chemiluminescence kit (Beyotime Institute of Biotechnology). The
grayscale value of the images was analyzed using Fusion FX Spectra
software (version 7; Vilber). The primary antibodies were as
follows: Anti-HOXA6 (1:800; cat. no. bs-11294R; BIOSS), anti-PI3K
(1:1,000; cat. no. 4257; Cell Signaling Technology, Inc.), anti-Akt
(1:1,000; cat. no. 4691; Cell Signaling Technology, Inc.),
anti-phosphorylated (p)-Akt (1:1,000; cat. no. 9271; Cell Signaling
Technology, Inc.), anti-PTEN (1:1,000; cat. no. 9188; Cell
Signaling Technology, Inc.), anti-MEK (1:1,000; cat. no. 8727; Cell
Signaling Technology, Inc.), anti-ERK (1:1,000; cat. no. 4695; Cell
Signaling Technology, Inc.), anti-p-ERK (1:1,000; cat. no. 4370;
Cell Signaling Technology, Inc.), anti-Bax (1:1,000; cat. no.
ab32503; Abcam), anti-Bcl-2 (1:1,000; cat. no. ab32124; Abcam),
anti-cleaved-PARP (1:1,000; cat. no. ab32561; Abcam),
anti-cleaved-caspase3 (1:1,000; cat. no. ab2302; Abcam),
anti-caspase3 (1:1,000; cat. no. ab44976; Abcam), anti-β-actin
(1:1,000; cat. no. ab8227; Abcam).
Cell Counting Kit-8 (CCK-8) assay
CCK-8 regent (Dojindo Molecular Technologies, Inc.)
was used to detect cell proliferation (24). At 48 h post-transfection, cells
were seeded in a 96-well plate (4×103 cells/well). At
each time point for detection (24, 48 and 72 h), cells were
incubated with CCK-8 reagent at 37°C for 2 h and the absorbance at
450 nm was measured.
Colony formation assay
At 48 h post-transfection, cells were seeded in a
6-well plate (500 cells/well), cultured at 37°C in 5%
CO2 for 2 weeks, then fixed using 4% paraformaldehyde at
room temperature for 15 min and stained with 0.5% gentian violet
(Beyotime Institute of Biotechnology) at room temperature for 15
min. Clones consisting of ≥50 cells were counted under a light
microscope (magnification, ×40) (25,26).
5-Ethynyl-2′-deoxyuridine (EdU)
staining
Cell proliferation was also analyzed using a
Cell-Light EdU Apollo643 in vitro kit (cat. no. C10310-2;
Guangzhou RiboBio Co., Ltd.), according to the manufacturer’s
protocol (27,28). A fluorescence microscope (Olympus
Corporation) was used for analysis and images were obtained
(magnification, ×100).
Caspase-Glo assay
The activities of caspases-3, -7, -8 and -9 were
measured using Caspase-Glo 3/7, 8 and 9 kits (cat. nos. G8090,
G8200 and G8210, respectively; Promega Corporation), according to
the manufacturer’s protocol. Transfected cells were seeded
(5×103 cells/well) into a 96-well plate and incubated
with Caspase-Glo reagents for 30 min. The luminescence was then
detected with a microplate reader.
Terminal deoxynucleotidyl transferase
dUTP nick end labeling (TUNEL)
Transfected cells were seeded onto a climbing slide
(5×103 cells/well), cultured overnight and stained with
a TUNEL reagent kit (In Situ Cell Death Detection kit, POD;
cat. no. 11684817910; Roche Diagnostics) (29) at 37°C for 1-2 h and DAPI at 37°C
for 5 min. Images were obtained using a fluorescence microscope
(magnification, ×100; Olympus Corporation).
Flow cytometry
To analyze the cell cycle, cells were suspended in
PBS buffer at 48 h post-transfection, fixed in 75% ethanol at 4°C
overnight and stained with propidium iodide (PI) at room
temperature for 30 min. For the analysis of cell apoptosis,
harvested cells were suspended in PBS and stained using an Annexin
V-fluorescein isothiocyanate/PI kit (cat. no. 556570, FITC Annexin
V Apoptosis Detection kit II, BD Biosciences). Following staining,
cells were immediately detected using a flow cytometer
(FACSCalibur; BD Biosciences), and analyzed with ModFit LT™ V3.3
(Verity Software House, Inc.) for cell cycle analysis and FlowJo 10
(FlowJo LLC) for apoptosis analysis (30,31).
Cell migration
Cell migration was evaluated using Transwell and
wound-healing assays with the 786-O cell line. For the Transwell
assay, 5×104 cells suspended in 200 µl RPMI-1640
medium was added to the upper chamber and 700 µl complete
medium was added to the lower chamber. After incubation for 48 h,
the migrated cells were fixed. The fixed cells were then stained
with crystal violet at room temperature for 15 min. The stained
cells were viewed and counted in five random fields of view under
an inverted phase contrast microscope (magnification, ×200). For
the wound-healing assay, the transfected cells were seeded in a
6-well plate (5×105 cells/well). At confluence, the cell
monolayer was scraped off and maintained in RPMI-1640. After 0, 24
and 48 h of incubation, an inverted microscope (magnification,
×100) was used to capture the images.
Statistical analysis
Data were analyzed using SPSS 22.0 (IBM Corp.) and
three independent experiments were performed. The data are
presented as the mean ± standard deviation. CCK-8 and wound-healing
results were statistically analyzed using two-way ANOVA. All other
data were compared using unpaired Student’s t-test. P<0.05 was
considered to indicate a statistically significant difference.
Results
HOXA6 is downregulated in ccRCC
specimens
The GSE6344 database contains ten paired ccRCC and
normal tissues. The mRNA expression levels of HOXA6 were extracted
from this database and analyzed. The results revealed that HOXA6
mRNA expression was significantly increased in normal tissues
compared with in carcinoma tissues (Fig. 1A). Subsequently, GEPIA online
analysis demonstrated that patients with lower expression levels of
HOXA6 exhibited significantly poorer survival (Fig. 1B). Additionally, a total of 25
paired ccRCC and adjacent non-tumor specimens were analyzed by IHC.
The majority of normal specimens (15/25) exhibited high expression
levels of HOXA6, whereas the majority of tumor specimens (23/25)
demonstrated low HOXA6 protein expression levels. HOXA6 was notably
expressed in the nucleus of renal tubular epithelial cells.
Furthermore, the protein expression levels of HOXA6 in normal
tissues were significantly increased compared with in paired
carcinoma tissues (Fig. 1C). These
results suggest that HOXA6 protein expression is downregulated in
ccRCC, which is in accordance with the findings of GSE6344
analysis.
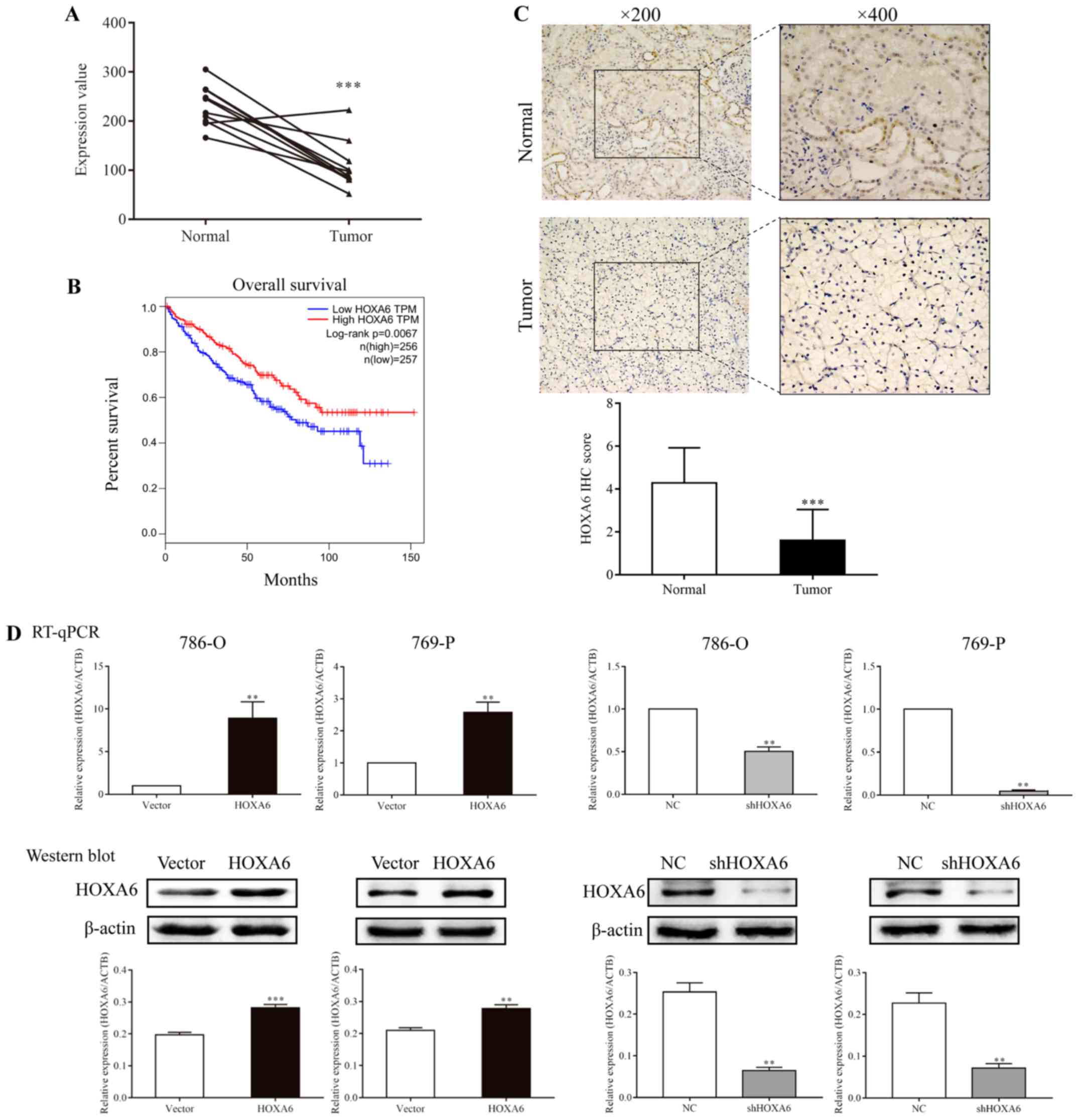 | Figure 1Expression of HOXA6 in carcinoma
tissues and transfected cell lines, and survival analysis. (A) The
mRNA expression levels of HOXA6 in ten paired ccRCC tumor and
normal tissues extracted from GSE6344. ***P<0.001 vs.
Normal. (B) Survival analysis of patients with ccRCC, which was
performed using Gene Expression Profiling Interactive Analysis. (C)
The expression of HOXA6 protein in ccRCC and normal tissues as
detected by immunohistochemistry. ***P<0.001 vs.
Normal. (D) Expression of HOXA6 in transfected cell lines.
**P<0.01 vs. vector or NC. ***P<0.001
vs. vector. ccRCC, clear cell renal cell carcinoma; HOXA6, homeobox
A6; RT-qPCR, reverse transcription-quantitative polymerase chain
reaction; vector, vector-only control; NC, negative control;
shHOXA6, HOXA6 short hairpin RNA; ACTB, β-actin; TPM, transcripts
per kilobase of exon model per million mapped reads. |
Detection of HOXA6 expression in
transfected cells
786-O and 769-P cells were transfected with HOXA6
plasmid, empty vector, NC or shHOXA6, and HOXA6 expression was
determined by RT-qPCR and western blotting. The results revealed
that the expression of HOXA6 in 786-O and 769-P cells
overexpressing HOXA6 was increased compared with the vector groups
(RT-qPCR, P=0.0032 and P=0.0058; western blotting, P<0.001 and
P=0.0020, respectively). Compared with the corresponding NC groups
of 786-O and 769-P cells, HOXA6 expression in the shHOXA6 groups
was significantly lower (RT-qPCR, P=0.0036 and P=0.0050; western
blotting, P=0.0019 and P=0.0030, respectively; Fig. 1D). These results indicated that
HOXA6 expression was successfully promoted and inhibited in the
respective groups of transfected cells.
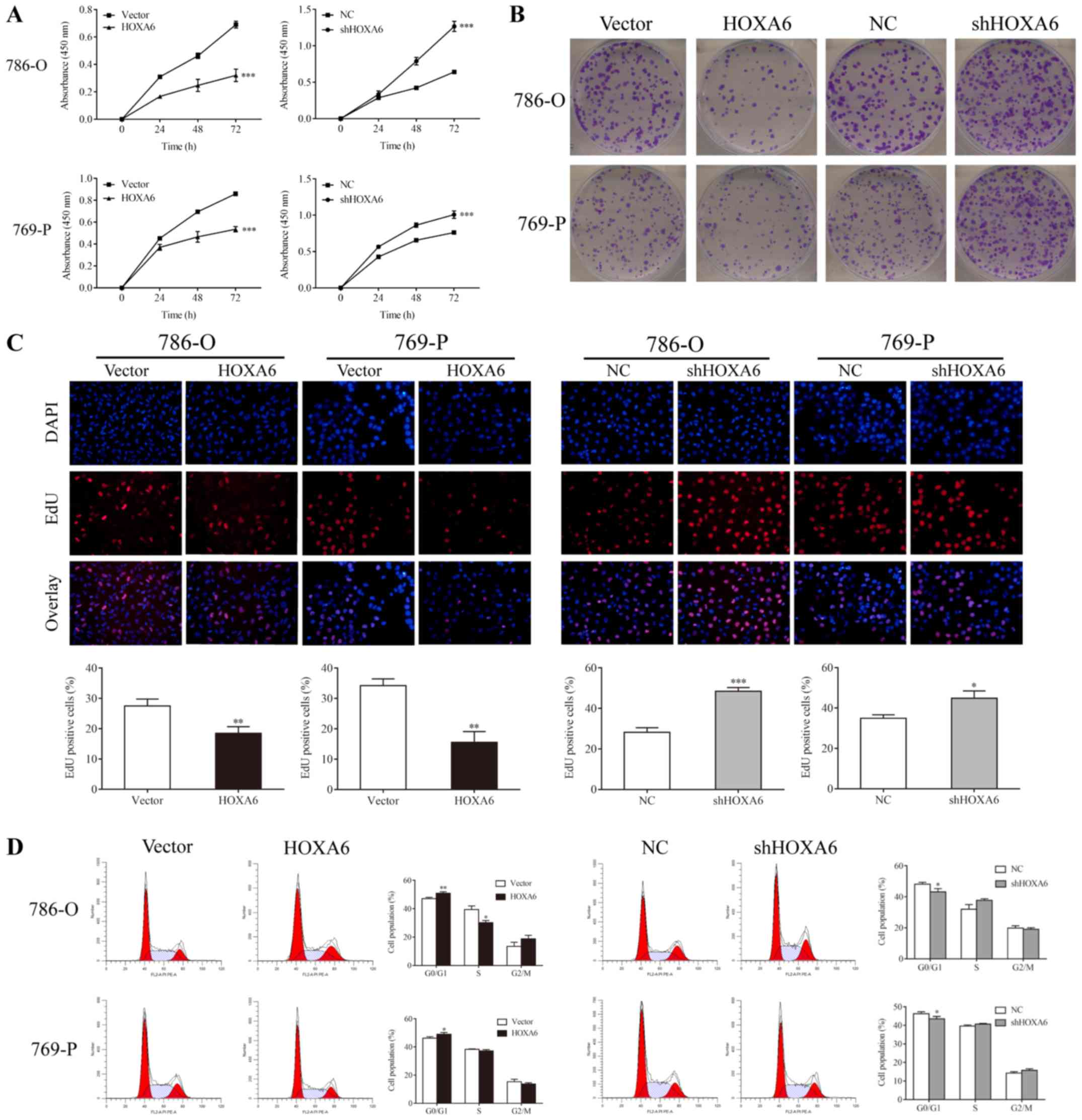
HOXA6 inhibits the proliferation of 786-O
and 769-P cells
CCK-8, colony formation, EdU and flow cytometry
assays were performed to determine the proliferation of transfected
786-O and 769-P cells. The CCK-8 assay revealed that
HOXA6-overexpressing cells exhibited reduced proliferation compared
with the vector groups (P<0.001), whereas the proliferation of
cells expressing shHOXA6 significantly increased compared with the
NC groups (P<0.001; Fig. 2A).
In addition, fewer colonies in the HOXA6 overexpression groups
(786-O, 95.67±11.71; 769-P, 143.33±10.02) were observed compared
with the vector groups (786-O, 199.33±23.25, P=0.0066; 769-P,
189.33±17.04, P=0.0238); however, the number of colonies was
significantly increased in the shHOXA6 groups (786-O, 306.00±22.72;
769-P, 338.33±28.04) compared with the corresponding NC groups
(786-O, 196.33±17.67, P=0.0034; 769-P, 213.00±13.11, P=0.0071;
Fig. 2B). Additionally, EdU
staining revealed that, compared with the vector groups (786-O,
27.53±2.27%; 769-P, 34.24±1.28%), the proliferation of
HOXA6-overexpressing cells (786-O, 18.44±2.26%, P=0.0079; 769-P,
15.52±2.07%, P=0.0015) was significantly inhibited. By contrast,
the proliferation of the shHOXA6 groups (786-O, 48.41±1.94%; 769-P,
44.86±3.60%) was promoted compared with the corresponding NC groups
(786-O, 28.19±2.28%, P<0.001; 769-P, 34.99±1.68%, P=0.0141;
Fig. 2C). Furthermore, the cell
cycle of transfected cells was analyzed by PI staining and flow
cytometry. Compared with the vector groups, the number of
G0/G1 phase cells was significantly increased
in the HOXA6 overexpression groups (786-O, P=0.0072; 769-P,
P=0.0380). By contrast, the number of G0/G1
phase cells was significantly decreased in the shHOXA6 expression
groups compared with the corresponding NC groups (786-O, P=0.0332;
769-P, P=0.0460; Fig. 2D). These
results indicate that HOXA6 suppresses cell proliferation by
inhibiting the cell cycle at the G0/G1 phase
in ccRCC.
HOXA6 induces the apoptosis of 786-O and
769-P cells
Caspase-Glo, TUNEL and flow cytometry assays were
performed to analyze the apoptosis of transfected cells. A
Caspase-Glo assay was performed to detect the activities of
caspases-3/7, -8 and -9. Compared with the vector group, the
activities of caspases-3/7, -8 and -9 (were significantly increased
in the HOXA6-overexpressing 786-O and 769-P cell groups. By
contrast, the activities of caspases-3/7 were significantly reduced
in the shHOXA6-expressing 786-O and 769-P cell groups compared with
the corresponding NC groups (Fig.
3A). The TUNEL assay also indicated that the apoptotic rates of
the HOXA6 overexpression groups (786-O, 30.80±1.34%; 769-P;
26.72±1.28%) were significantly increased compared with the vector
groups (786-O, 25.31±2.41%, P=0.0384; 769-P, 23.69±0.86%,
P=0.0332). However, the apoptotic rates were significantly
decreased in the shHOXA6 groups (786-O, 17.61±2.82%; 769-P,
12.83±2.39%) compared with the corresponding NC groups (786-O,
24.31±1.30%, P=0.0372; 769-P, 22.92±0.58%, P=0.0141; Fig. 3B). In addition, flow cytometry was
performed to analyze the apoptosis of transfected cells. The
results revealed that, compared with the vector group, upregulation
of HOXA6 significantly induced cell apoptosis (786-O, P=0.0067;
769-P, P=0.0027), while downregulation of HOXA6 suppressed
apoptosis compared with the corresponding NC groups (786-O,
P=0.0475; 769-P, P=0.0418; Fig.
3C). These results demonstrated that HOXA6 could induce the
apoptosis of ccRCC cells.
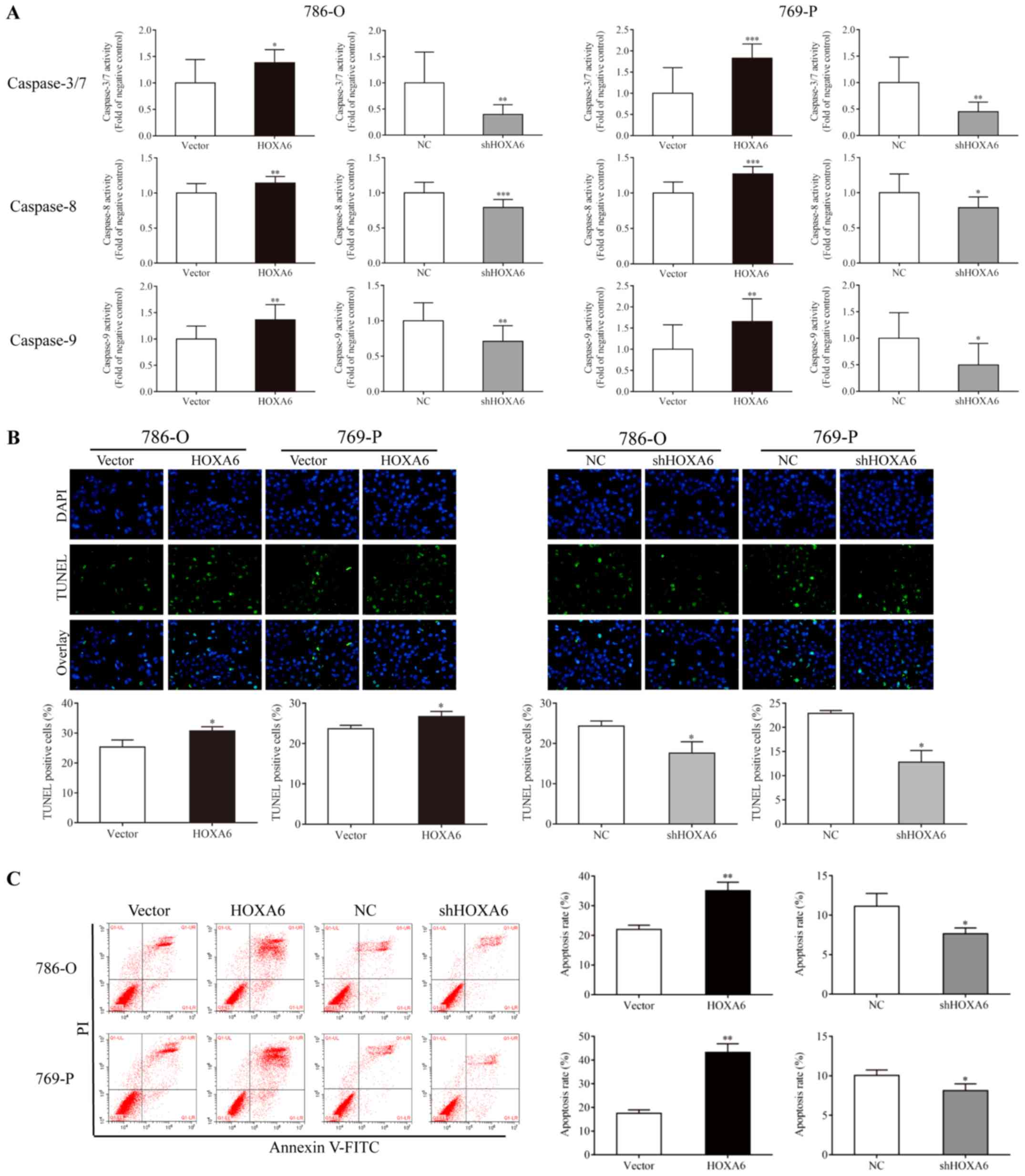 | Figure 3Cell apoptosis as analyzed via
Caspase-Glo, TUNEL and flow cytometry assays. (A) Caspase-Glo
analysis was performed to detect the apoptosis of cells.
*P<0.05, **P<0.01,
***P<0.001 vs. vector or NC. (B) Cells stained with
TUNEL and DAPI were considered as apoptotic, *P<0.05
vs. vector or NC (magnification, ×200). (C) Cell apoptosis as
analyzed by flow cytometry. *P<0.05 vs. NC,
**P<0.01 vs. vector .TUNEL, terminal deoxynucleotidyl
transferase dUTP nick end labeling; HOXA6, homeobox A6; vector,
vector-only control; NC, negative control; shHOXA6, HOXA6 short
hairpin RNA; PI, propidium iodide; FITC, fluorescein
isothiocyanate. |
HOXA6 inhibits the PI3K/Akt signaling
pathway
The expression levels of PI3K, Akt, p-Akt,
phosphatase and tensin homolog (PTEN), MEK, ERK, p-ERK, B-cell
lymphoma 2 (Bcl-2)-associated X protein (Bax), Bcl-2, caspase-3,
cleaved caspase-3 and cleaved poly (ADP-ribose) polymerase (PARP)
were analyzed by western blotting. The expression levels of PI3K,
p-Akt, MEK, p-ERK and Bcl-2 were significantly suppressed in the
HOXA6 overexpression groups, but were significantly upregulated
following downregulation of HOXA6. The expression levels of PTEN,
Bax, cleaved caspase-3 and cleaved PARP were significantly
increased following HOXA6 overexpression compared with the vector
group, but were significantly decreased in the shHOXA6 groups
compared with the corresponding NC group (Fig. 4). These results suggest that HOXA6
inhibits cell proliferation and induces apoptosis via suppression
of the PI3K/Akt signaling pathway.
 | Figure 4Western blotting for the analysis of
proteins involved in the PI3K/Akt signaling pathway. Compared with
the vector group, the expression levels of PI3K, p-Akt, MEK, p-ERK
and Bcl-2 were suppressed in the HOXA6 group, but the expression
levels of PTEN, Bax, cleaved caspase-3 and cleaved PARP were
increased. Opposing results were observed in the shHOXA6 group
compared with the NC group. *P<0.05,
**P<0.01, ***P<0.001 vs. vector or NC.
Akt, protein kinase B; PI3K, phosphoinositide 3-kinase; p,
phosphorylated; PTEN, phosphatase and tensin homolog; MEK,
mitogen-activated protein kinase kinase; ERK, extracellular
signal-regulated kinase; Bax, B-cell lymphoma 2-associated X
protein; Bcl-2, B-cell lymphoma 2; PARP, poly (ADP-ribose)
polymerase; HOXA6, homeobox A6; vector, vector-only control; NC,
negative control; shHOXA6, HOXA6 short hairpin RNA. |
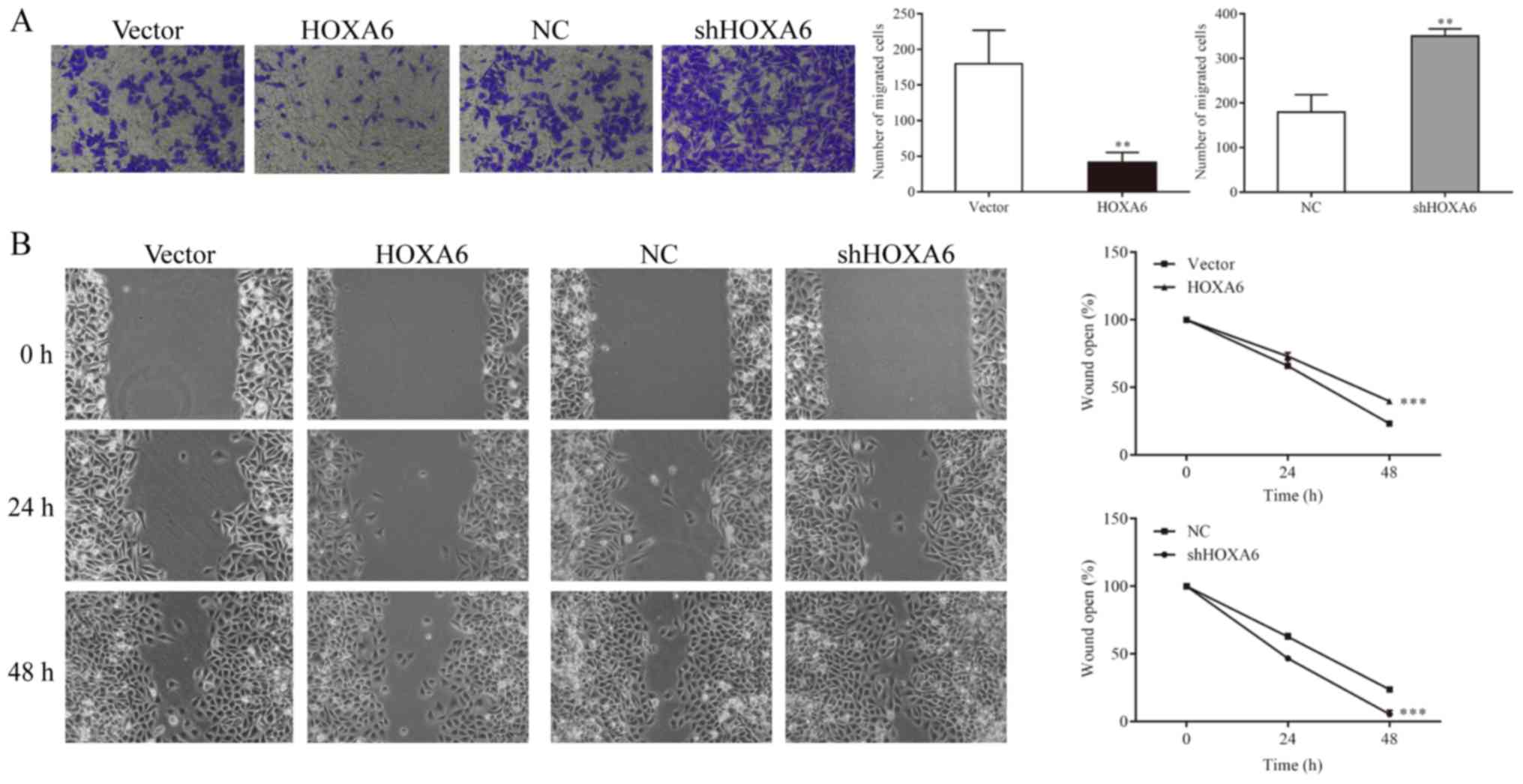
HOXA6 inhibits the migration of 786-O
cells
Cell migration was detected by Transwell and
wound-healing assays. The Transwell assay results indicated that
significantly fewer migrated cells were observed in the HOXA6
overexpression group compared with the vector group (P=0.0082). In
addition, the number of migrated cells was significantly increased
in the shHOXA6 group compared with the NC group (P=0.0023; Fig.5A). The wound-healing assay results
revealed that compared with the vector group, the migration ability
was significantly reduced in the HOXA6-overexpressing cells
(P<0.001). By contrast, the migration ability was significantly
increased in the shHOXA6-expressing cells compared with the NC
group (P<0.001; Fig. 5B).
Discussion
It has been reported that HOXA6 is upregulated in
certain types of cancer and exhibits a role as an oncogene
(13-15); however, studies have demonstrated
that the CpG islands of HOXA6 are hypermethylated in malignant
tumors (16,17). In addition, downregulated HOXA6
expression has been reported in breast cancer tissues (18); therefore, conflicting observations
have been reported. In the present study, the expression of HOXA6
in ccRCC was downregulated. To further investigate this finding,
the expression of HOXA6 in ccRCC microarrays from a GEO database
was analyzed. The expression of HOXA6 mRNA was downregulated in
cancer tissues in the GSE6344 dataset. To further verify these
results, HOXA6 protein expression in ccRCC tissues was determined
by IHC. The results revealed reduced expression of HOXA6 in cancer
tissues compared with normal tissues. Furthermore, survival
analysis was performed via GEPIA, which was based on data of The
Cancer Genome Atlas. This analysis demonstrated that low expression
of HOXA6 was associated with poor survival. The aforementioned
results indicate that HOXA6 may serve a protective role in ccRCC.
Furthermore, following the promotion or suppression of HOXA6
expression in ccRCC cells, alterations in cell proliferation and
apoptosis were analyzed in the present study. CCK-8, colony
formation, EdU staining and flow cytometry assays indicated that
HOXA6 may suppress cell proliferation and inhibit the cell cycle at
the G0/G1 phase. Caspase-Glo, TUNEL and flow
cytometry assays further revealed that HOXA6 may promote cell
apoptosis, and the activities of caspases-3/7, -8 and -9. However,
a limitation of the present study is that a rescue experiment of
apoptosis by HOXA6 was not performed.
The PI3K/Akt signaling pathway serves an important
role in ccRCC for the regulation of cell proliferation,
differentiation and survival. PTEN is a crucial inhibitor of the
PI3K/Akt signaling pathway and functions as a tumor suppressor
(32-34). In the present study, overexpression
of HOXA6 promoted the expression of PTEN, leading to reductions in
PI3K expression and Akt phosphorylation. p-Akt has a series of
downstream effectors and is involved in various sub-pathways,
including the mammalian target of rapamycin, glycogen synthase
kinase 3, Bcl-2-associated death promoter, ERK, nuclear factor-κB
and the c-Jun N-terminal kinase signaling pathways (35). The progression of the cell cycle
from the G1 to S-phase is dependent on the activation of
ERK (36). MEK is the upstream
regulator of ERK, and can induce proliferation and inhibit
apoptosis; thus, MEK may be a potential target of cancer therapy
(37,38). The present study revealed that
upregulation of HOXA6 led to reductions in the levels of p-Akt,
increased the expression of MEK and suppressed the phosphorylation
of ERK. The ERK signaling pathway has been reported to induce the
anti-apoptotic properties of Bcl-2 (39). This supports the results of the
present study as the expression of Bcl-2 was suppressed but that of
Bax was increased, which leads to a decrease of the Bcl-2/Bax ratio
following the upregulation of HOXA6. A decrease in the Bcl-2/Bax
ratio has been negatively associated with cell apoptosis (40,41).
Caspase-3 is the most important executioner of apoptosis, while
PARP is an indicator of caspase-3 activation (42-44).
The present study revealed that the protein expression levels of
caspase-3 and PARP were upregulated in the HOXA6 group. These
findings demonstrated that HOXA6 was involved in regulating cell
proliferation and apoptosis, which may occur via the PI3K/Akt/ERK
cascade in ccRCC cells.
In the present study, the expression of HOXA6 was
suppressed in ccRCC tissues, which was associated with poor
survival. In addition, it was identified that HOXA6 may be involved
in regulating cell proliferation and apoptosis in ccRCC. Analysis
of the pathways associated with these processes revealed that HOXA6
may be involved in the PI3K/Akt signaling pathway; however, the
mechanism of how it regulates this pathway requires further
investigation. The underlying mechanism of HOXA6 downregulation in
ccRCC remains unknown; however, HOXA6 has been reported to be
hypermethylated in meningiomas (17). Thus, further investigation is
required to determine whether HOXA6 is hypermethylated in ccRCC and
the additional roles of HOXA6 in this disease.
In summary, the present study reported that the
expression of HOXA6 was downregulated in ccRCC and was negatively
associated with the survival of patients. HOXA6 expression was
determined to be associated with reduced cell proliferation and
increased apoptosis, which may occur via inhibition of the
PI3K/Akt/ERK pathway. These results indicate that HOXA6 may serve a
role in the regulation of ccRCC; however, the underlying mechanism
requires further investigation.
Funding
The present study was supported by a grant from the
Foundation of Chongqing Science and Technology Commission (grant
no. cstc2015shmszx0466).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors’ contributions
FW made substantial contributions to the study
conception, design, acquisition of data, analysis and
interpretation of the data, and was involved in drafting the
manuscript. SW and HT took part in the acquisition, analysis and
interpretation of the data. WH and XG were involved in study
conception, study design and revising the manuscript critically for
important intellectual content. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Written informed consent was signed by all patients
and ethics approval was obtained from the Ethics Committee of the
First Affiliated Hospital of Chongqing Medical University
(Chongqing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
ccRCC
|
clear cell renal cell carcinoma
|
|
IHC
|
immunohistochemistry
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
CCK-8
|
Cell Counting Kit-8
|
|
EdU
|
5-ethynyl-2′-deoxyuridine
|
|
TUNEL
|
terminal deoxynucleotidyl trans-ferase
dUTP nick end labeling
|
Acknowledgments
The authors would like to thank Dr Lixue Chen (The
First Affiliated Hospital of Chongqing Medical University,
Chongqing, China) for providing technical support.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
King SC, Pollack LA, Li J, King JB and
Master VA: Continued increase in incidence of renal cell carcinoma
especially in young patients and high grade disease: United States
2001 to 2010. J Urol. 191:1665–1670. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lipworth L, Tarone RE and McLaughlin JK:
The epidemiology of renal cell carcinoma. J Urol. 176:2353–2358.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Banegas MP, Harlan LC, Mann B and Yabroff
KR: Renal cell cancer: A shift in approaches for treatment of
advanced disease in the United States. J Natl Compr Canc Netw.
12:1271–1279. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sunela KL, Lehtinen ET, Kataja MJ, Kujala
PM, Soimakallio S and Kellokumpu-Lehtinen PL: Development of renal
cell carcinoma (RCC) diagnostics and impact on prognosis. BJU Int.
113:228–235. 2014. View Article : Google Scholar
|
|
6
|
Roy B, Pattanaik AK, Das J, Bhutia SK,
Behera B, Singh P and Maiti TK: Role of PI3K/Akt/mTOR and MEK/ERK
pathway in Concanavalin A induced autophagy in HeLa cells. Chem
Biol Interact. 210:96–102. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ersahin T, Tuncbag N and Cetin-Atalay R:
The PI3K/AKT/mTOR interactive pathway. Mol Biosyst. 11:1946–1954.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Polivka J Jr and Janku F: Molecular
targets for cancer therapy in the PI3K/AKT/mTOR pathway. Pharmacol
Ther. 142:164–175. 2014. View Article : Google Scholar
|
|
9
|
Guo H, German P, Bai S, Barnes S, Guo W,
Qi X, Lou H, Liang J, Jonasch E, Mills GB, et al: The PI3K/AKT
pathway and renal cell carcinoma. J Genet Genomics. 42:343–353.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu F, Wu S and Gou X: Identification of
biomarkers and potential molecular mechanisms of clear cell renal
cell carcinoma. Neoplasma. 65:242–252. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dickson GJ, Kwasniewska A, Mills KI,
Lappin TR and Thompson A: Hoxa6 potentiates short-term hemopoietic
cell proliferation and extended self-renewal. Exp Hematol.
37:322–333. e3232009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Guo YB, Shao YM, Chen J, Xu SB, Zhang XD,
Wang MR and Liu HY: Effect of overexpression of HOX genes on its
invasive tendency in cerebral glioma. Oncol Lett. 11:75–80. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang H, Liu Y, Yan L, Zhang M, Yu X, Du
W, Wang S, Li Q, Chen H, Zhang Y, et al: Increased levels of the
long noncoding RNA, HOXA-AS3, promote proliferation of A549 cells.
Cell Death Dis. 9:7072018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu S, Wu F and Jiang Z: Effect of HOXA6 on
the proliferation, apoptosis, migration and invasion of colorectal
cancer cells. Int J Oncol. 52:2093–2100. 2018.PubMed/NCBI
|
|
15
|
Eoh KJ, Kim HJ, Lee J-Y, Nam EJ, Kim S,
Kim SW and Kim YT: Upregulation of homeobox gene is correlated with
poor survival outcomes in cervical cancer. Oncotarget.
8:84396–84402. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xavier FC, Destro MF, Duarte CM and Nunes
FD: Epigenetic repression of HOXB cluster in oral cancer cell
lines. Arch Oral Biol. 59:783–789. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Galani V, Lampri E, Varouktsi A, Alexiou
G, Mitselou A and Kyritsis AP: Genetic and epigenetic alterations
in meningiomas. Clin Neurol Neurosurg. 158:119–125. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hur H, Lee JY, Yun HJ, Park BW and Kim MH:
Analysis of HOX gene expression patterns in human breast cancer.
Mol Biotechnol. 56:64–71. 2014. View Article : Google Scholar
|
|
19
|
Gumz ML, Zou H, Kreinest PA, Childs AC,
Belmonte LS, LeGrand SN, Wu KJ, Luxon BA, Sinha M, Parker AS, et
al: Secreted frizzled-related protein 1 loss contributes to tumor
phenotype of clear cell renal cell carcinoma. Clin Cancer Res.
13:4740–4749. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tun HW, Marlow LA, von Roemeling CA,
Cooper SJ, Kreinest P, Wu K, Luxon BA, Sinha M, Anastasiadis PZ and
Copland JA: Pathway signature and cellular differentiation in clear
cell renal cell carcinoma. PLoS One. 5:e106962010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Edge SB, Byrd DR, Compton CC, Fritz AG,
Greene FL and Trotti A: AJCC Cancer Staging Manual. 7th edition.
Springer; New York, NY: pp. 547–560. 2010
|
|
22
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45(W1):
W98–W102. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
24
|
Ishiyama M, Miyazono Y, Sasamoto K, Ohkura
Y and Ueno K: A highly water-soluble disulfonated tetrazolium salt
as a chro-mogenic indicator for NADH as well as cell viability.
Talanta. 44:1299–1305. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Franken NA, Rodermond HM, Stap J, Haveman
J and van Bree C: Clonogenic assay of cells in vitro. Nat Protoc.
1:2315–2319. 2006. View Article : Google Scholar
|
|
26
|
Rafehi H, Orlowski C, Georgiadis GT,
Ververis K, El-Osta A and Karagiannis TC: Clonogenic assay:
Adherent cells. J Vis Exp. (49): 25732011.PubMed/NCBI
|
|
27
|
Salic A and Mitchison TJ: A chemical
method for fast and sensitive detection of DNA synthesis in vivo.
Proc Natl Acad Sci USA. 105:2415–2420. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pierzyńska-Mach A, Szczurek A, Cella
Zanacchi F, Pennacchietti F, Drukała J, Diaspro A, Cremer C,
Darzynkiewicz Z and Dobrucki JW: Subnuclear localization, rates and
effectiveness of UVC-induced unscheduled DNA synthesis visualized
by fluorescence widefield, confocal and super-resolution
microscopy. Cell Cycle. 15:1156–1167. 2016. View Article : Google Scholar
|
|
29
|
Gavrieli Y, Sherman Y and Ben-Sasson SA:
Identification of programmed cell death in situ via specific
labeling of nuclear DNA fragmentation. J Cell Biol. 119:493–501.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Szeberenyl J: Analysis of the cell cycle
by flow cytometry. Biochem Mol Biol Educ. 35:153–154. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Riccardi C and Nicoletti I: Analysis of
apoptosis by propidium iodide staining and flow cytometry. Nat
Protoc. 1:1458–1461. 2006. View Article : Google Scholar
|
|
32
|
Worby CA and Dixon JE: Pten Annu Rev
Biochem. 83:641–669. 2014. View Article : Google Scholar
|
|
33
|
Hopkins BD, Hodakoski C, Barrows D, Mense
SM and Parsons RE: PTEN function: The long and the short of it.
Trends Biochem Sci. 39:183–190. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wyatt LA, Filbin MT and Keirstead HS: PTEN
inhibition enhances neurite outgrowth in human embryonic stem
cell-derived neuronal progenitor cells. J Comp Neurol.
522:2741–2755. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hemmings BA and Restuccia DF: PI3K-PKB/Akt
pathway. Cold Spring Harb Perspect Biol. 4:a0111892012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Meloche S and Pouysségur J: The ERK1/2
mitogen-activated protein kinase pathway as a master regulator of
the G1- to S-phase transition. Oncogene. 26:3227–3239. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhao Y and Adjei AA: The clinical
development of MEK inhibitors. Nat Rev Clin Oncol. 11:385–400.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Neuzillet C, Tijeras-Raballand A, de
Mestier L, Cros J, Faivre S and Raymond E: MEK in cancer and cancer
therapy. Pharmacol Ther. 141:160–171. 2014. View Article : Google Scholar
|
|
39
|
Yang T, Xu F, Sheng Y, Zhang W and Chen Y:
A targeted proteomics approach to the quantitative analysis of
ERK/Bcl-2-mediated anti-apoptosis and multi-drug resistance in
breast cancer. Anal Bioanal Chem. 408:7491–7503. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zeren T, Inan S, Vatansever HS and Sayhan
S: Significance of apoptosis related proteins on malignant
transformation of ovarian tumors: A comparison between Bcl-2/Bax
ratio and p53 immunoreactivity. Acta Histochem. 116:1251–1258.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Samarghandian S, Nezhad MA and Mohammadi
G: Role of caspases, Bax and Bcl-2 in chrysin-induced apoptosis in
the A549 human lung adenocarcinoma epithelial cells. Anticancer
Agents Med Chem. 14:901–909. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Brentnall M, Rodriguez-Menocal L, De
Guevara RL, Cepero E and Boise LH: Caspase-9, caspase-3 and
caspase-7 have distinct roles during intrinsic apoptosis. BMC Cell
Biol. 14:322013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Boulares AH, Yakovlev AG, Ivanova V,
Stoica BA, Wang G, Iyer S and Smulson M: Role of poly(ADP-ribose)
polymerase (PARP) cleavage in apoptosis. Caspase 3-resistant PARP
mutant increases rates of apoptosis in transfected cells. J Biol
Chem. 274:22932–22940. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mullen P: PARP cleavage as a means of
assessing apoptosis. Methods Mol Med. 88:171–181. 2004.
|