Introduction
Pancreatic cancer is one of the most lethal
malignancies worldwide. Numerous factors, such as difficulty in
establishing an early diagnosis, recurrence, metastasis and
resistance to chemotherapy and radiotherapy, contribute to the poor
prognosis of this disease. Over the past 30 years, the long-term
survival rate of patients with pancreatic cancer has remained low
(as low as 6%) (1). A high rate of
metastasis is one of the principal causes of therapy failure in
patients with pancreatic cancer according to an autopsy series,
which reported that distant metastasis occurs in 90% of cases
(2). As the benefits from surgical
resection are limited for patients with early recurrence or distant
metastasis, and as chemotherapy, radiation therapy and palliative
care are insufficient to prevent metastasis, the identification of
novel therapeutic methods or targets for pancreatic cancer is
urgently required (3).
Gradients of the extracellular matrix (ECM) serve
important roles in supporting tumor cell proliferation, adhesion,
migration and survival (4). Matrix
metalloproteinase-1 (MMP1) degrades the ECM and basement membrane
components, resulting in tumor progression via the remodeling of
the ECM. Several factors contribute to the expression of MMP1,
including endogenous factors, such as polymorphisms and epigenetic
regulation in the promoter region of MMP1, as well as exogenous
factors, such as the tumor microenvironment. A variety of MMP1
promoter genotypic polymorphisms have been reported in various
ethnic populations, and their contributions to different disease
risks have been explored (5-7). In
the tumor microenvironment, a variety of inflammatory factors,
including interleukin-8 (IL-8) (8), IL-1β (9) and tumor necrosis factor-α (TNF-α)
(10), have been reported to
induce MMP1 expression in cancer cells.
Another important mechanism that influences the
expression of MMP1 is epigenetic modification. MicroRNAs
(miRs/miRNAs) are a group of small non-coding RNAs that have been
identified as important regulators of MMP gene expression at the
post-transcriptional level (11).
miRNA-mediated MMP regulation is considered to be a key factor
involved in neoplastic tumors, and is closely associated with the
clinicopathological features and outcomes of patients (12). miRNAs may serve as promising
biomarkers for predicting the therapeutic response or for
prognostic evaluation, and may additionally serve as potential
therapeutic targets for various types of cancer, including
pancreatic cancer (13-15). Several miRNAs have been identified
to target MMP1, such as miR-222 in hypertrophic scars (16), miR-526b in the skin (17), miR-let-7b in stem cells from apical
papilla (18) and miR-202-3p in
scleroderma fibrosis (19).
However, whether the miRNA modulation of MMP1 expression could
attenuate metastasis induced by inflammatory factors in pancreatic
cancer requires further investigation, as it is yet to be
completely elucidated.
In the present study, a systematical analysis,
combining in silico, in vitro, in vivo and
bioinformatics approaches, was performed to investigate the
interactions between miRNAs and MMP1. miRNA-623 was observed to
suppress MMP1 by targeting its 3′-untranslated region (3′-UTR),
attenuate IL-8-induced MMP1 expression, and inhibit pancreatic
cancer cell migration, invasion and metastasis in vitro and
in vivo.
Materials and methods
Cell lines
The 293T and human pancreatic cancer PANC-1 and
BXPC-3 cell lines were purchased from the American Type Culture
Collection and were incubated at 37°C in a humidified incubator
containing 5% CO2. The cells were cultured in DMEM
supplemented with 10% fetal bovine serum (FBS) (both from Thermo
Fisher Scientific, Inc.), 100 U/ml penicillin and 100 μg/ml
streptomycin (both Jiangsu KeyGEN BioTECH Corp., Ltd.).
Mice
A total of 20 female nude mice
(BALB/cJNju-Foxn1nu/Nju; age, 5 weeks; weight, 16±2.5 g) were
purchased from Nanjing Biomedical Research Institute (http://www.nbri-nju.com/en-us/about-animal-facility).
All the animals were raised under pathogen-free conditions at
20-26°C, 40-70% relative humidity, 15 times/h ventilation and a 12
h light/12 h dark cycle. The animals were provided with water and
food freely. All animal experiments were approved by the Animal
Care and Use Committee of Sun Yat-Sen University (Guangzhou,
China).
Reagents and chemicals
Synthetic miR-623 mimics
(5′-ACCCAACAGCCCCUGCAAGGGAU-3′), inhibitors
(5′-CAGUACUUUUGUGUAGUACAA-3′), miR- 623 mutant
(5′-ATAACTTGAAGGGGCTGTTGGGT-3′), mimics negative control (NC;
5′-UUCUCCGAACGUGUCACGUTTACGUGACACGUUCGGAGAATT-3′) and inhibitor NC
(5′-CAGUACUUUUGUGUAGUACAA-3′) miRNAs were purchased from Shanghai
GenePharma Co., Ltd. Recombinant human IL-8 (rIL-8) was purchased
from PeproTech, Inc. Rabbit-anti-human MMP1 (cat. no. 54376), ERK
(cat. no. 4695), phosphorylated (p-)ERK (cat. no. 4370),
Twist-related protein 1 (TWIST1; cat. no. 46702), zinc finger
protein SNAI1 (cat. no. 3879) and E-cadherin (cat. no. 14472)
polyclonal primary antibodies, mouse GAPDH (cat. no. 51332)
monoclonal primary antibody and horseradish peroxidase-conjugated
secondary antibodies (anti-rat cat. no. 7077; anti-rabbit cat. no.
7074) were purchased from Cell Signaling Technology, Inc.
Phalloidin was purchased from Sigma-Aldrich; Merck KGaA, and DAPI
was from Wuhan Boster Biological Technology, Ltd.
In silico analyses
The microRNA.org
(http://www.microrna.org/), PITA (http://genie.weizmann.ac.il/pubs/mir07/mir07_prediction.html),
TargetScan (Release 6.2, http://www.targetscan.org) and miRTar.human
(http://mirtar.mbc.nctu.edu.tw/human/)
databases were screened for potential miRNAs binding to the 3′-UTR
of the MMP1 transcript (acc. no. NM_002421). The minimum free
energy (MFE) of hybridization was calculated using the RNAhybrid
program (http://bibiserv2.cebitec.uni-bielefeld.de/rnahybrid).
Luciferase reporter assay
The pGL3-Control vector (Promega Corporation) and
the pGL3-CU vector were constructed according to the protocol
outlined in previous studies (20,21).
The sequence of the MMP1 3′-UTR was amplified via PCR using primers
listed in Table I. The PCR
products were digested with USER enzyme (New England BioLabs, Inc.)
and ligated into the pGL3-CU vector, resulting in a pGL3-MMP1
vector, according to the manufacturer's protocols. The authenticity
of pGL3-MMP1 vector was confirmed.
 | Table ISequences of primers and
oligonucleotides used in this study. |
Table I
Sequences of primers and
oligonucleotides used in this study.
| Assay | Name | Sequence
(5′-3′) |
|---|
| Luciferase reporter
gene assay | MMP1 3′-UTR
forward |
CCAAAGAAGGTGTTTTCCTG |
| MMP1 3′-UTR
reverse |
GAAAGTGTATAAAACAGTAG |
| hsa-miR-623
mutant |
ATAACTTGAAGGGGCTGTTGGGT |
|
Electrophoretic | dye-miR-623 |
/5IRD800CWN/rArUrCrCrCrUrUrGrCrArGrGrGrGrCrUrGrUr |
| mobility shift
assay | | UrGrGrGrU |
| cold-miR-623 |
/rArUrCrCrCrUrUrGrCrArGrGrGrGrCrUrGrUrUrGrGrGrU |
|
dye-miR-623-TAR |
/5Cy55/mUmCmAmCmCmCmUmGmGmAmUmAmGmGm |
| |
CmAmAmGmGmGmAmU |
| cold-negative
control |
/rUrCrArCrArArCrCrUrCrCrUrArGrArArArGrArGrUrArGrA |
| Quantitative
PCR | MMP1 forward |
ACACGCCAGATTTGCCAAGAGC |
| MMP1 reverse | GGAGAGTTGT
CCCGATGATCTCCCC |
| GAPDH forward |
GAAATCCCATCACCATCTTCCAGG |
| GAPDH reverse |
GAGCCCCAGCCTTCTCCATG |
| hsa-miR-623
forward |
ATCCCTTGCAGGGGCTGTTGGGT |
| U6 forward |
CTCGCTTCGGCAGCACA |
| miR universal
reverse |
AACGCTTCACGAATTTGCGT |
To verify the precise target of the miRNAs, the
Dual-Luciferase Reporter Assay system (Promega Corporation) was
used. Briefly, 293T cells were seeded in 96-well plates at a
density of 1×105 cells per well and cultured in complete
medium without antibiotics for 24 h. The luciferase reporter
plasmids pGL3-NC (Promega Corporation) and pGL3-MMP1 (100 ng/well)
were then co-transfected with a mimic NC, hsa-miR-623 mimic,
inhibitor NC, hsa-miR-623 inhibitor or hsa-miR-623 mutant (50 nM)
using Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Firefly and Renilla luciferase activities
were measured at 24 h post-transfection using the Dual-Luciferase
Reporter Assay kit. Firefly luciferase was normalized to
Renilla luciferase activity. Three independent experiments
were performed.
RNA electrophoretic mobility shift assay
(EMSA)
An RNA EMSA was performed using a LightShift
Chemiluminescent RNA EMSA kit (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. The binding buffer
mixture was prepared with 1X REMSA binding buffer supplemented with
5% glycerol, 200 mM KCl and 100 mM MgCl2. Next, 200 nmol
IRDye1800-labeled hsa-miR-623 and/or Cy5.5-labeled hsa-miR-623 TAR
(targeting sequences in the MMP1 3-'UTR) oligonucleotides
(Integrated DNA Technologies, Inc.) were added to the binding
buffer mixture. Cytoplasmic protein from PANC-1 cells was extracted
using NE-PER Nuclear and Cytoplasmic Extraction reagents (Thermo
Fisher Scientific, Inc.). The oligonucleotide sequences are listed
in Table I.
To confirm the binding specificity between
hsa-miR-623 and its target, 50-fold molar excess of
cold-hsa-miR-623 or cold-NC probe was added to the reaction system
5 min prior to the addition of the dye-labeled probes. A total of 2
μg PANC-1 cell cytoplasmic protein and 1 μg tRNA were
added to the hsa-miR-623/target mixtures to examine the RNA-protein
interactions. Following a 20-min incubation at room temperature,
the reaction mixture was loaded onto a 10% gel for PAGE and
separated by electrophoresis (100 V; 150 min) at 4°C. Finally, the
bands were detected using an Odyssey CLx Infrared Imaging system
(LI-COR Biosciences).
Incubation with rIL-8
PANC-1 and BXPC-3 cells were seeded at a density of
6×105 cells per well in 6-well plates and cultured for
12 h to allow cells to fully attach. The medium was then replaced
with serum-free medium, and rIL-8 was added to a final
concentration of 100 ng/ml (22,23),
followed by incubation for 6, 12, 24, 36 and 48 h. The cells were
then harvested for further analyses.
Cell transfection
Synthetic miR-623 mimics, inhibitors and NC miRNAs
were transfected into PANC-1 and BXPC-3 cells using Lipofectamine
2000 reagent (Invitrogen; Thermo Fisher Scientific, Inc.). PANC-1
and BXPC-3 cells were seeded at a density of 6×105 cells
per well in 6-well plates, 3 μl transfection reagent was
added into each well, and the final concentration of microRNA
(mimic NC, mimic, inhibitor NC or inhibitor) was 20 nmol/l in a
2-ml final volume. At 6 h following incubation, the medium was
changed to fresh complete medium, and the cells were harvested 48 h
post-transfection. The transfection efficiency was determined by
reverse transcription-quantitative PCR (RT-qPCR).
For stable transfections, 3 μg miR-623-GFP or
NC-GFP vector were packaged into lentiviral virus vectors,
according to the manufacturer's protocol (Vigene Biosciences,
Inc.). The PANC-1 cells were then transfected with the lentiviruses
and selected with 2 μg/ml puromycin (Sigma-Aldrich; Merck
KGaA). The transfection efficiency was evaluated by RT-qPCR.
RT-qPCR
Total RNA was extracted from PANC-1 and BXPC-3 cells
using the miRNeasy Mini kit (Qiagen GmbH). The RNA concentration
and purity were determined according to the A260/280 values, using
a NanoDrop spectrophotometer (NanoDrop Technologies; Thermo Fisher
Scientific, Inc.). A High-Capacity cDNA Reverse Transcription kit
(Applied Biosystems; Thermo Fisher Scientific, Inc.) or
miScript® II RT kit (Qiagen GmbH) was used for
first-strand cDNA synthesis. The samples were reverse transcribed
at 37°C for 15 min and heated to 85°C for 5 sec to inactivate the
reverse transcriptase.
The qPCR was performed on an ABI Prism 7900 Sequence
Detection system (Applied Biosystems; Thermo Fisher Scientific,
Inc.) using a QuantiFast SYBR1 Green RT-PCR kit (Qiagen GmbH). The
reactions were preheated for 2 min at 50°C and denatured for 2 min
at 90°C, followed by 40 cycles of denaturation at 95°C for 15 sec
and annealing/extension at 60°C for 15 sec. The qPCR results,
recorded as threshold cycle numbers (Ct), were calculated by the
2−ΔΔCq method with normalization against GAPDH mRNA as
an internal control (24). The
relative expression levels of MMP1 and hsa-miR-623 were normalized
to the housekeeping gene GAPDH and U6 small nuclear RNA,
respectively. The primers used in the RT-qPCR were obtained from
Integrated DNA Technologies, Inc. and their sequences are listed in
Table I.
Western blot analysis
Protein extracts from PANC-1 and BXPC-3 cells were
prepared using RIPA lysis buffer with protease inhibitors and
quantified using a bicinchoninic acid protein assay kit (Pierce;
Thermo Fisher Scientific, Inc.). A total of 20 μg protein
from each experimental group was separated on a polyacrylamide gel
containing 10% sodium dodecyl sulfate and subsequently transferred
to a 0.45-μm polyvinylidene difluoride membrane (EMD
Millipore). The membrane was then blocked with 5% non-fat milk at
room temperature for 1 h. Following an incubation with the primary
polyclonal primary antibodies MMP1 (1:1,000 dilution), TWIST1
(1:500 dilution), SNAI1 (1:1,000 dilution), E-cadherin (1:400
dilution), ERK (1:1,000 dilution), p-ERK (1:2,000 dilution) or
GAPDH (1:2,000 dilution) overnight at 4°C and incubation with the
secondary antibodies (1:2,000 dilution) for 1 h at room
temperature, protein bands were detected using enhanced
chemiluminescence reagents (Thermo Fisher Scientific, Inc.) and
visualized after exposure using the Tanon-5200 Multi-imaging system
(Tanon Science and Technology Co., Ltd.). ImageJ version 1.8.0
software (National Institutes of Health) was used to analyze the
images.
Immunofluorescence assay
Following transfection with synthetic miR-623
mimics, inhibitors and NC miRNAs, and co-culturing with IL-8,
PANC-1 and BXPC-3 cells were fixed with 4% formaldehyde for 20 min
at room temperature. The cells were then stained with 5
μg/ml phalloidin for 30 min, counterstained with 1
μg/ml DAPI for 10 min at room temperature, and finally
visualized using a fluorescence scanning microscope at ×400
magnification.
Wound-healing assay
PANC-1 and BXPC-3 cells were transfected with
synthetic miR-623 mimics, inhibitors and NC miRNAs, and a
wound-healing assay was performed 48 h after transfection. When the
cells reached 100% confluence, a wound was created by manually
scraping the cell monolayer with a 200-μl pipette tip. At 0
and 24 h after incubation, images were captured using a light
microscope (Zeiss GmbH) at ×40 magnification, and analyzed
quantitatively by ImageJ version 1.8.0 software.
Transwell invasion assay
An invasion assay was performed using 8-μm
pore, 24-well Transwell chambers coated with Matrigel (Corning Life
Sciences). PANC-1 and BXPC-3 cells were transfected with synthetic
miR-623 mimics, inhibitors and NC miRNAs. At 48 h after
transfection, the cells were harvested, 5×104 cells in
200 μl serum-free medium were reseeded into the top chamber,
and the bottom chamber was filled with 750 μl DMEM
containing 10% FBS. Cells that migrated to the underside of the
membrane were stained with 0.1% crystal violet (Phygene Life
Sciences Co., Ltd.) for 30 min at room temperature, followed by
imaging and counting under a light microscope (Zeiss GmbH) at ×40
magnification. All experiments were performed in triplicate and
repeated ≥3 times.
In vivo efficacy studies
To investigate the effect of hsa-miR-623 on the
metastasis of pancreatic cancer cells, 1×106 PANC-1
cells stably expressing miR-623 or NC were injected into the tail
veins of mice, which were divided into 4 groups (n=5):
PANC-1Lenti-GFP, PANC-1Lenti-miR-623-GFP,
PANC-1Lenti-GFP+IL-8 and
PANC-1Lenti-miR623-GFP+IL-8. The rIL-8 was administered
via intraperitoneal injection at 100 ng/animal once a week for 5
weeks. The weight, activity and appearance of the mice were
monitored every 3 days. If the body weight of the animal dropped
markedly in a short time, the subcutaneous fat disappeared, the
animals exhibited spinal curvature, and the activity and reaction
ability decreased, euthanasia was carried out in advance, otherwise
euthanasia was carried out at 5 weeks. The mice were anaesthetized
by intraperitoneal injection of 0.5% pentobarbital sodium salt
(CAS, 57-33-0; Beijing Huayehuanyu Chemical Co., Ltd.) at a dosage
of 100 mg/kg, and then placed in transparent glass containers. 100%
carbon dioxide was introduced into the bottles with the filling
rate of 10-30% volume per min. The death of the mice was confirmed
when no spontaneous breathing was detected for 2-3 min and they
exhibited no blinking reflex. Subsequently, the livers and lungs of
the animals were dissected and imaged under a fluorescence scanning
microscope. All dissected organs were fixed overnight with 4%
formalin at room temperature and then embedded in paraffin. The
livers and lungs were sliced to 5-μm thickness and stained
with hematoxylin for 15 min and eosin for 1 min at room
temperature. Immunochemical assays were performed according to the
protocols of the immunohistochemical staining kit manufacturer
(cat. no. SA1022; Wuhan Boster Biological Technology, Ltd.), using
the aforementioned primary antibodies incubated overnight at 4°C.
The numbers of liver and lung metastases were independently
calculated and assessed by two pathologists.
The Cancer Genome Atlas (TCGA) and Gene
Expression Omnibus (GEO) dataset analysis
The clinicopathological characteristics, mRNA
expression levels of MMP1 and IL-8, and the hsa-miR-623 levels in
pancreatic ductal adenocarcinoma (PDAC) tissues were obtained from
TCGA database (https://cancergenome.nih.gov/), which contained 172
patients with PDAC. As there are only 4 normal samples included in
TCGA database, the present study also searched other GEO
(https://www.ncbi.nlm.nih.gov/geo/)
public datasets for further data, including datasets GES28735 (45
paired PDAC and adjacent non-tumor tissues), GSE32678 and GSE32676
(data for miRNA and mRNA expression levels in 25 PDAC tissues)
(25,26).
Statistical analysis
All statistical analyses were performed using SPSS
version 17.0 software (SPSS, Inc.). The results are presented as
the mean ± standard deviation. Pearson's correlation analysis was
used to analyze the correlation between MMP1 mRNA levels and
hsa-miR-623 levels, and between MMP1 mRNA levels and IL-8 mRNA
levels in the original data from human PDAC tissues published in
the GEO or TCGA databases. One-way analysis of variance was used to
compare data from multiple groups, and the Bonferroni method was
used to determine the significant differences between the groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Identification of potential miRNAs that
regulate MMP1
The screening results from all four databases
suggested that several miRNAs may target the 3′-UTR of the MMP1
transcript. The correlation between the expression of miRNA and the
expression of MMP1 in human PDAC tissues was analyzed, and the
results are presented in Table
II. According to our previous studies, the value of MFE ≤-20
kcal/mol was considered to be essential for the miRNA/mRNA
hybridization (20,21,27-29).
As presented in Table II, the MFE
of hybridization between hsa-miR-623 and its cognate target at the
3′-UTR of MMP1 was -27 kcal/mol. Based on the database analysis,
compared with adjacent non-tumor tissues, the expression of MMP1 in
PDAC tumor tissues was increased, whereas the expression of
hsa-miR-623 was decreased, and a negative correlation was observed
between their expression (r=−0.402, P=0.046; Fig. 1A). Based on these results,
hsa-miR-623 was identified as a candidate miRNA for further
investigation.
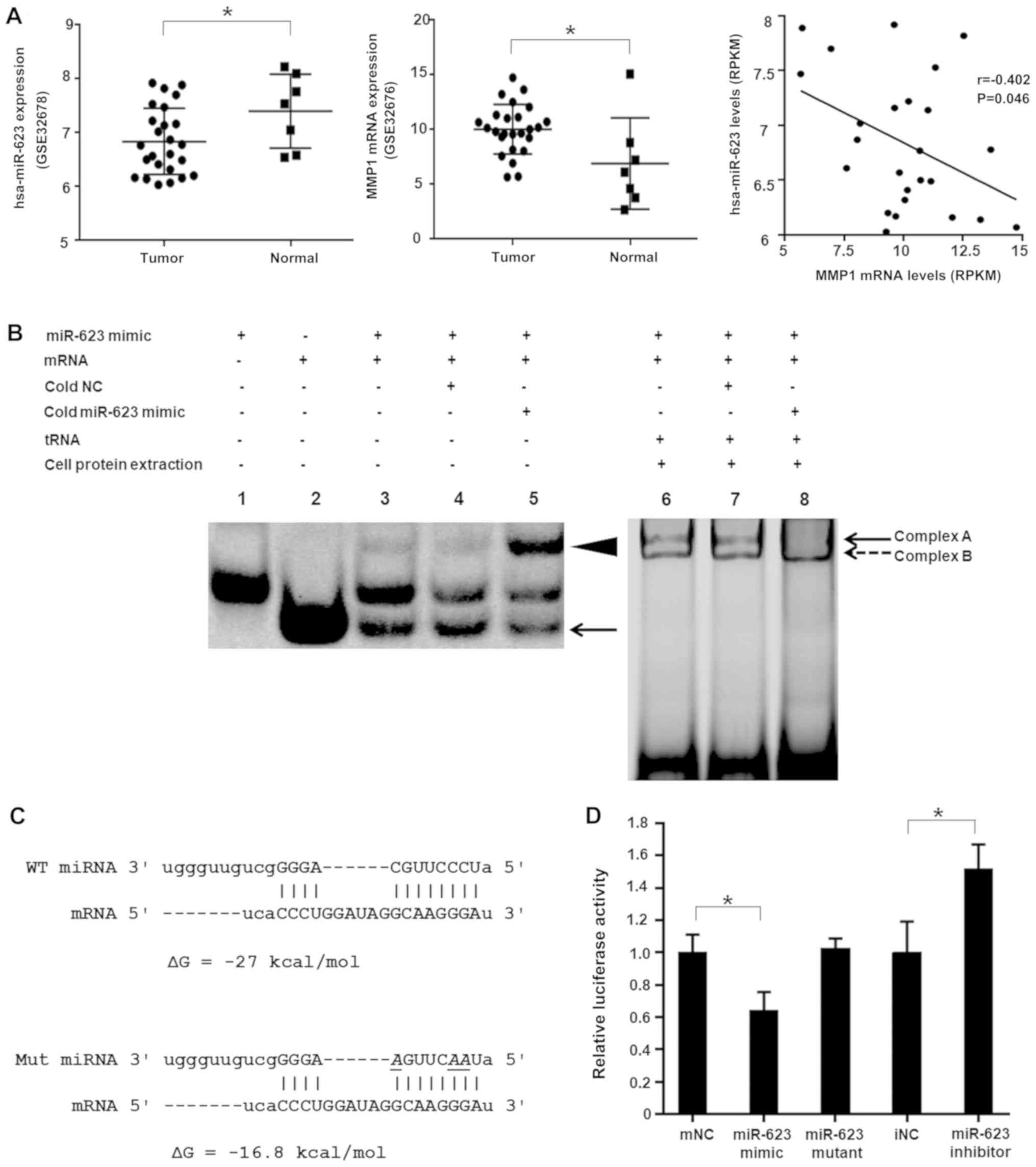 | Figure 1hsa-miRNA-623 modulates MMP1 by
targeting its 3′-UTR. (A) The expression of MMP1 in PDAC tumor
tissues was increased; however, the expression of hsa-miR-623 was
decreased compared with adjacent non-tumor tissues, and the levels
of hsa-miR-623 were negatively correlated with MMP1 mRNA levels in
human PDAC tissues (datasets GSE32678 and GSE32676).
*P<0.05. (B) RNA electrophoretic mobility shift
assays. In lane 4 the cold NC did not affect hybridization, and in
lane 5 hybridization was inhibited, as most of the mRNA was bound
by cold hsa-miR-623. The arrowhead indicates miR/mRNA
hybridization, and the arrow indicates free mRNA. miR/mRNA/protein
complex formation is evident in lanes 6-8 (complexes A and B) and
was not affected by cold NC, yet was inhibited by a 50-fold molar
excess of cold hsa-miR-623 (lane 8). (C) In silico analyses
of the interaction between hsa-miRNA-623 and its target sequence
within the 3′-UTR of MMP1 mRNA. The free energy of the predicted
hybrid complex formed by hsa-miR-623 and its target was -27
kcal/mol. When 3 nucleotides in the hsa-miR-623 sequence were
altered (italic, underlined), the predicted free energy of
hybridization changed to -16.8 kcal/mol. (D) hsa-miR-623
specifically regulated luciferase activity produced by the reporter
gene plasmid containing the 3′-UTR of MMP1 in 293T cells, whereas
mutant hsa-miR-623 did not have a significant influence on
luciferase activity. The data are presented as the mean ± standard
deviation (n=3). *P<0.05. miR, microRNA; MMP1, matrix
metal-loproteinase-1; 3′-UTR, 3′-untranslated region; RPKM, Reads
Per Kilobase Million; NC, negative control; mNC, mimic NC; iNC,
inhibitor NC. |
| Table IICandidate miRs targeting the
3′-untranslated region of MMP1 and expression correlation in
pancreatic ductal adenocarcinoma tissuesa. |
Table II
Candidate miRs targeting the
3′-untranslated region of MMP1 and expression correlation in
pancreatic ductal adenocarcinoma tissuesa.
| Correlation
| Free energyb, kcal/mol | Positionc |
|---|
| miR name | R | P-value |
|---|
| hsa-miR-623 | −0.4025 | 0.046 | −27.0 | 8158-8179 |
| hsa-miR-127-5p | −0.293 | 0.155 | −22.0 | 8046-8063 |
| hsa-miR-188-5p | 0.050 | 0.811 | −28.2 | 8159-8180 |
| hsa-miR-190b | −0.2661 | 0.199 | −21.6 | 8057-8077 |
|
hsa-miR-1915-5p | −0.1671 | 0.425 | −27.8 | 8159-8177 |
| hsa-miR-330-5p | 0.0296 | 0.888 | −24.2 | 8032-8053 |
| hsa-miR-361-5p | −0.030 | 0.888 | −24.0 | 7838-7859 |
| hsa-miR-526a | −0.223 | 0.283 | −20.6 | 7828-7849 |
|
hsa-miR-518c-5p | −0.030 | 0.888 | −19.0 | 8006-8026 |
|
hsa-miR-518f-5p | −0.308 | 0.134 | −19.6 | 7828-7849 |
|
hsa-miR-519a-5p | −0.337 | 0.099 | −20.6 | 7828-7849 |
|
hsa-miR-518e-5p | −0.101 | 0.631 | −20.6 | 7828-7849 |
| hsa-miR-523-5p | −0.337 | 0.099 | −20.6 | 7828-7849 |
| hsa-miR-558 | 0.177 | 0.397 | −23.3 | 8115-8133 |
| hsa-miR-769-3p | 0.237 | 0.253 | −24.4 | 8029-8050 |
| hsa-miR-638 | −0.104 | 0.620 | −26.3 | 8028-8049 |
| hsa-miR-936 | −0.173 | 0.408 | −20.5 | 8220-8241 |
hsa-miR-623 directly interacts with its
target within the MMP1 3′-UTR
RNA EMSA was performed to explore whether direct
binding occurs between hsa-miR-623 and its target within the MMP1
3′-UTR. As illustrated in Fig. 1B,
complexes formed between hsa-miR-623 and their cognate MMP1 mRNA
target sequences in vitro (lanes 3-5, the duplex is
indicated by an arrowhead). Competition assays indicated that the
complex of hsa-miR-623 and its cognate targets was
sequence-specific, as the complex formation was inhibited by a
50-fold molar excess of cold hsa-miR-623 (lane 5); however, was not
influenced by a 50-fold molar excess of unlabeled NC
oligonucleotide (lane 4). PANC-1 cytoplasmic extracts were
subsequently used to observe the formation of miRNA/mRNA/protein
complexes (lanes 6-8, complexes A and B). Taken together, the
competition assays revealed that complex A was inhibited by a
50-fold molar excess of cold hsa-miR-623 (lane 8), suggesting that
hsa-miR-623 directly and specifically interacts with its target
within the MMP1 3′-UTR.
hsa-miR-623 suppresses MMP1 3′-UTR
luciferase reporter gene activity, MMP1 endogenous expression, and
the migration and invasion of pancreatic cancer cells
Luciferase reporter assays were performed to explore
whether hsa-miR-623 could regulate the 3′-UTR of MMP1. The mutant
hsa-miR-623 was designed with 3 cytosines replaced by adenines;
consequently, the free energy of hybridization between the mutated
miRNA and its target mRNA changed from -27 to -16.8 kcal/mol
(Fig. 1C). Following dual
transfection, the luciferase activity produced by the reporter gene
plasmid was significantly downregulated by the wild-type
hsa-miR-623 mimic (0.64±0.11; P<0.05) and increased by the
hsa-miR-623 inhibitor (1.52±0.15; P<0.05). Conversely, the
mutant hsa-miR-623 mimic failed to influence the luciferase
activity in the 293T cells (Fig.
1D). These results indicate that hsa-miR-623 modulates its
target in the 3′-UTR of MMP1 in a direct and specific manner.
Following synthetic miRNA transfection, mimics increased and
inhibitors decreased hsa-miR-623 expression levels in the PANC-1
and BXPC-3 cells (Fig. 2A and D).
Consistent with the results from the EMSA and luciferase assay, the
hsa-miR-623 mimic suppressed endogenous MMP1 expression at the mRNA
and protein levels, whereas the hsa-miR-623 inhibitor led to an
increase in MMP1 expression (Fig. 2B,
C, E and F). Furthermore, the hsa-miR-623 mimic resulted in a
decrease in the migration and invasion ability of PANC-1 and BXPC-3
cells, whereas the hsa-miR-623 inhibitor displayed the opposite
effects (all P<0.05; Fig. 3A and
B).
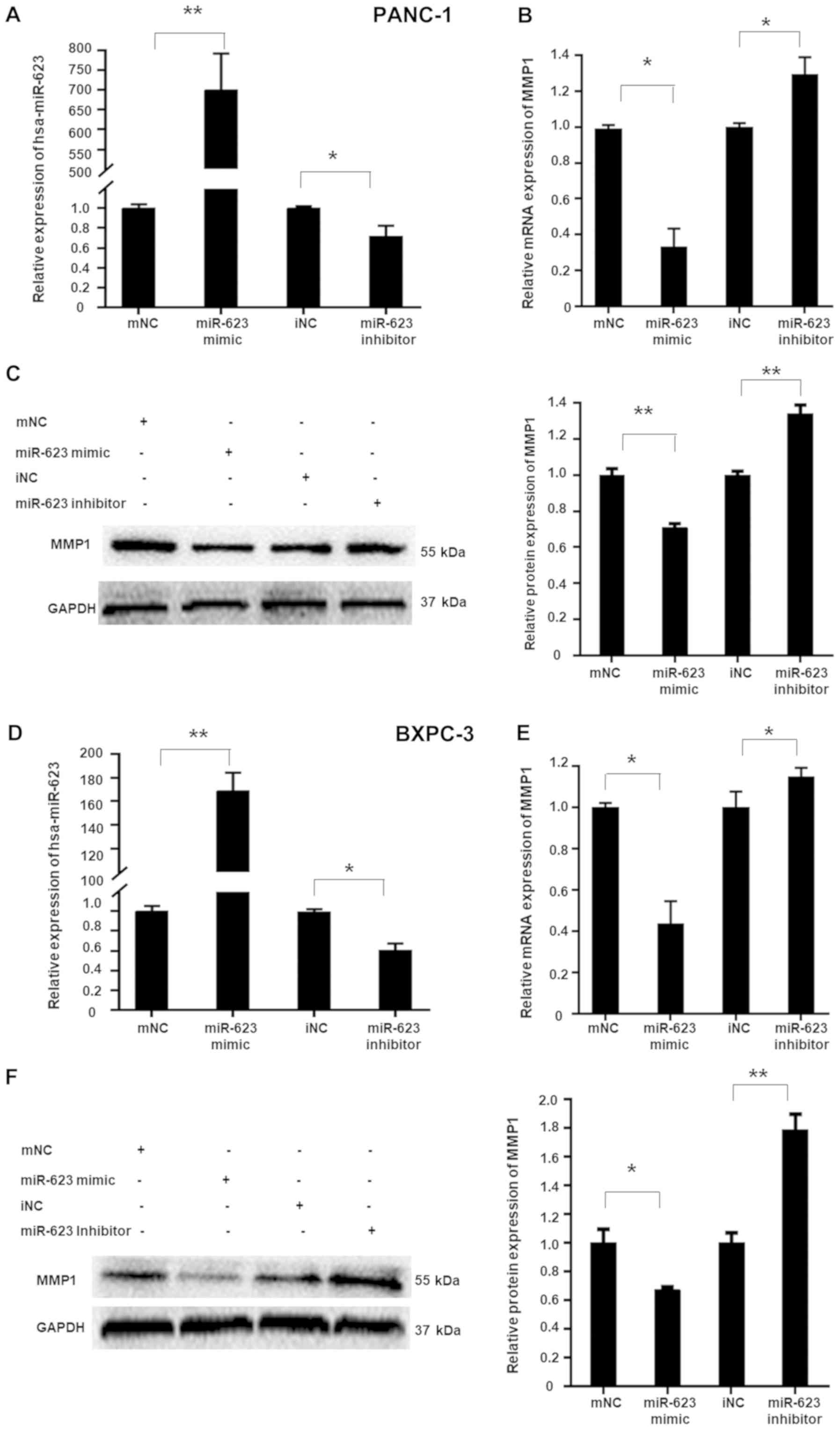 | Figure 2hsa-miR-623 suppresses endogenous
MMP1 expression in PANC-1 and BXPC-3 cells. Cells were transiently
transfected using 50 nmol/l hsa-miR-623 mimic, mNC, hsa-miR-623
inhibitor or iNC. Transfection of the hsa-miR-623 mimic
significantly increased hsa-miR-623 levels in (A) PANC-1 and (D)
BXPC-3 cells, whereas the hsa-miR-623 inhibitor decreased the
hsa-miR-623 levels. The hsa-miR-623 mimic suppressed, whereas the
hsa-miR-623 inhibitor upregulated, endogenous MMP1 expression in (B
and C) PANC-1 and (E and F) BXPC-3 cells. The data are presented as
the mean ± standard deviation (n=3). *P<0.05,
**P<0.001. miR, microRNA; MMP1, matrix
metalloproteinase-1; mNC, mimic negative control; iNC, inhibitor
negative control. |
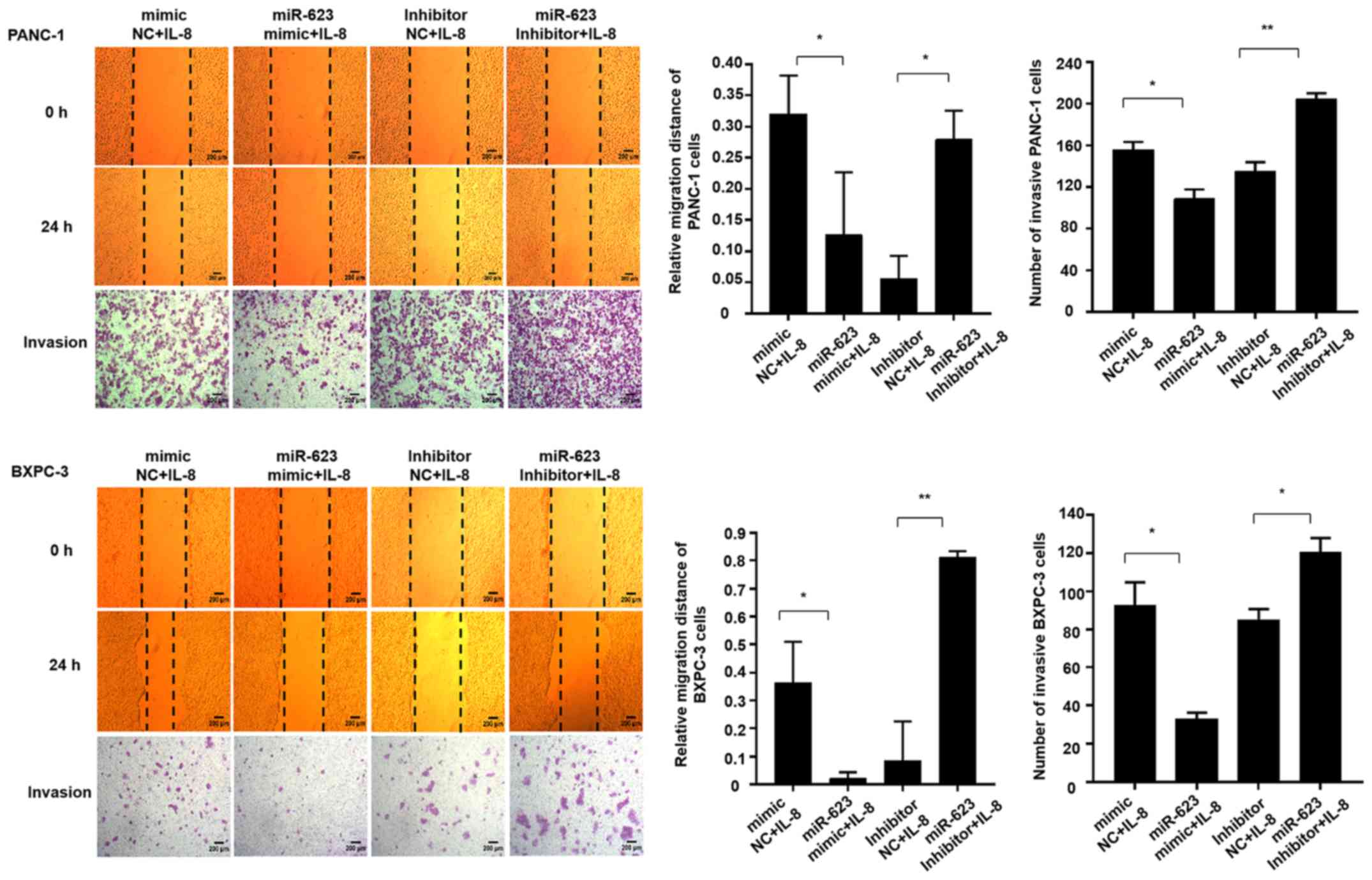
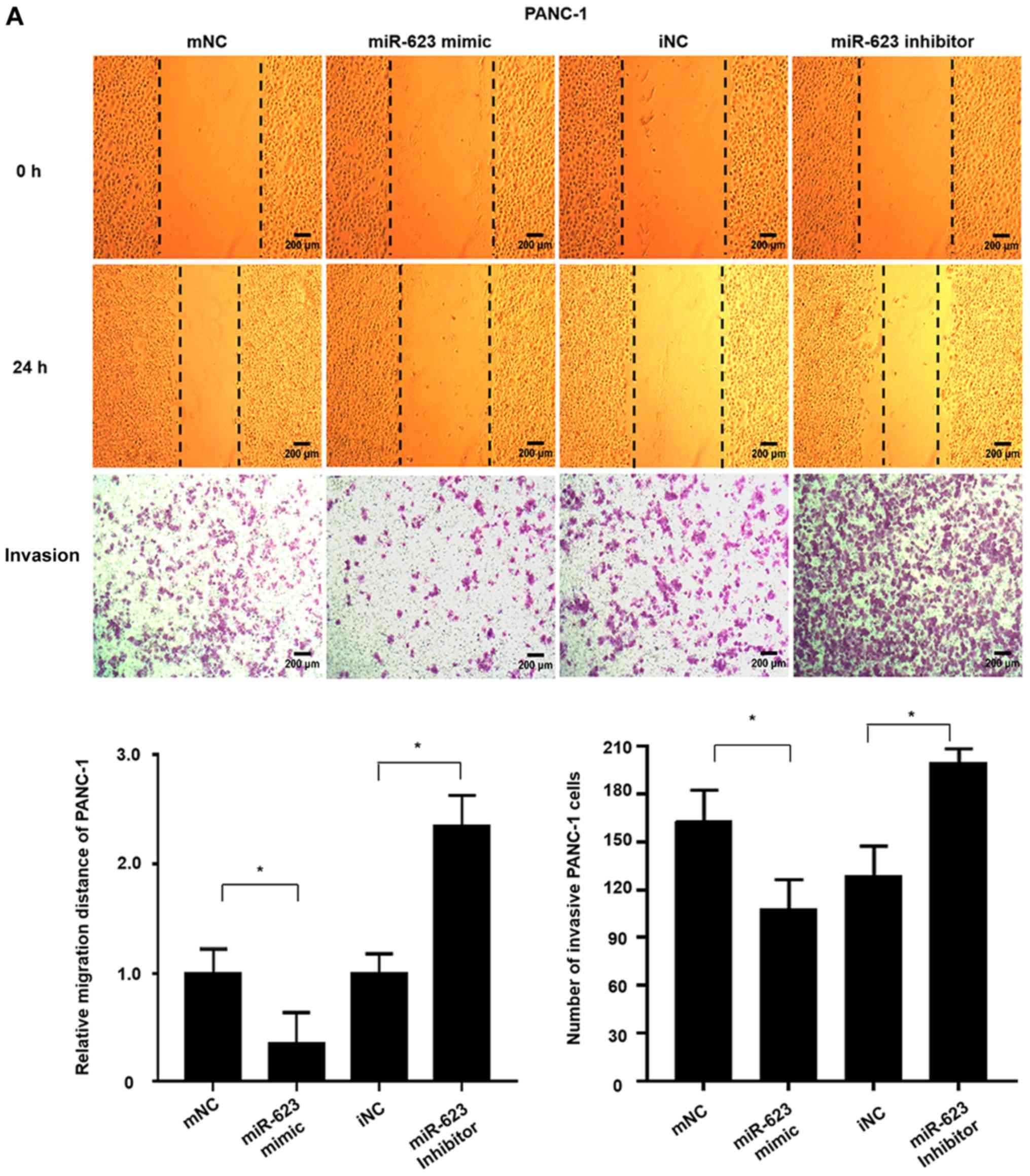 | Figure 3hsa-miR-623 inhibits the migration
and invasion of PANC-1 and BXPC-3 cells. Cells were transiently
transfected with 50 nmol/m hsa-miR-623 mimic, mNC, hsa-miR-623
inhibitor or iNC. A wound-healing assay was performed to
investigate the migratory characteristics of the cells, and a
Transwell assay was used to investigate the invasive
characteristics. The hsa-miR-623 mimic inhibited, whereas the
hsa-miR-623 inhibitor markedly enhanced the migration and invasion
of (A) PANC-1 cells. The data are presented as the mean ± standard
deviation (n=3). Magnification, ×40. *P<0.05. miR,
microRNA; mNC, mimic negative control; iNC, inhibitor negative
control. hsa-miR-623 inhibits the migration and invasion of PANC-1
and BXPC-3 cells. Cells were transiently transfected with 50 nmol/m
hsa-miR-623 mimic, mNC, hsa-miR-623 inhibitor or iNC. A
wound-healing assay was performed to investigate the migratory
characteristics of the cells, and a Transwell assay was used to
investigate the invasive characteristics. The hsa-miR-623 mimic
inhibited, whereas the hsa-miR-623 inhibitor markedly enhanced the
migration and invasion of (B) BXPC-3 cells. The data are presented
as the mean ± standard deviation (n=3). Magnification, ×40.
*P<0.05. miR, microRNA; mNC, mimic negative control;
iNC, inhibitor negative control. |
hsa-miR-623 attenuates IL-8-induced MMP1
expression, and the migration and invasion of pancreatic cancer
cells
In rheumatoid arthritis, MMP1 can be induced by
TNF-α and IL-1β (30,31), however, whether MMP1 can be induced
by inflammatory factors in PDAC cells remains unclear. First, the
correlation was investigated between the expression of MMP1 mRNA
and 3 commonly studied inflammatory factors, IL-1β, IL-6 and IL-8.
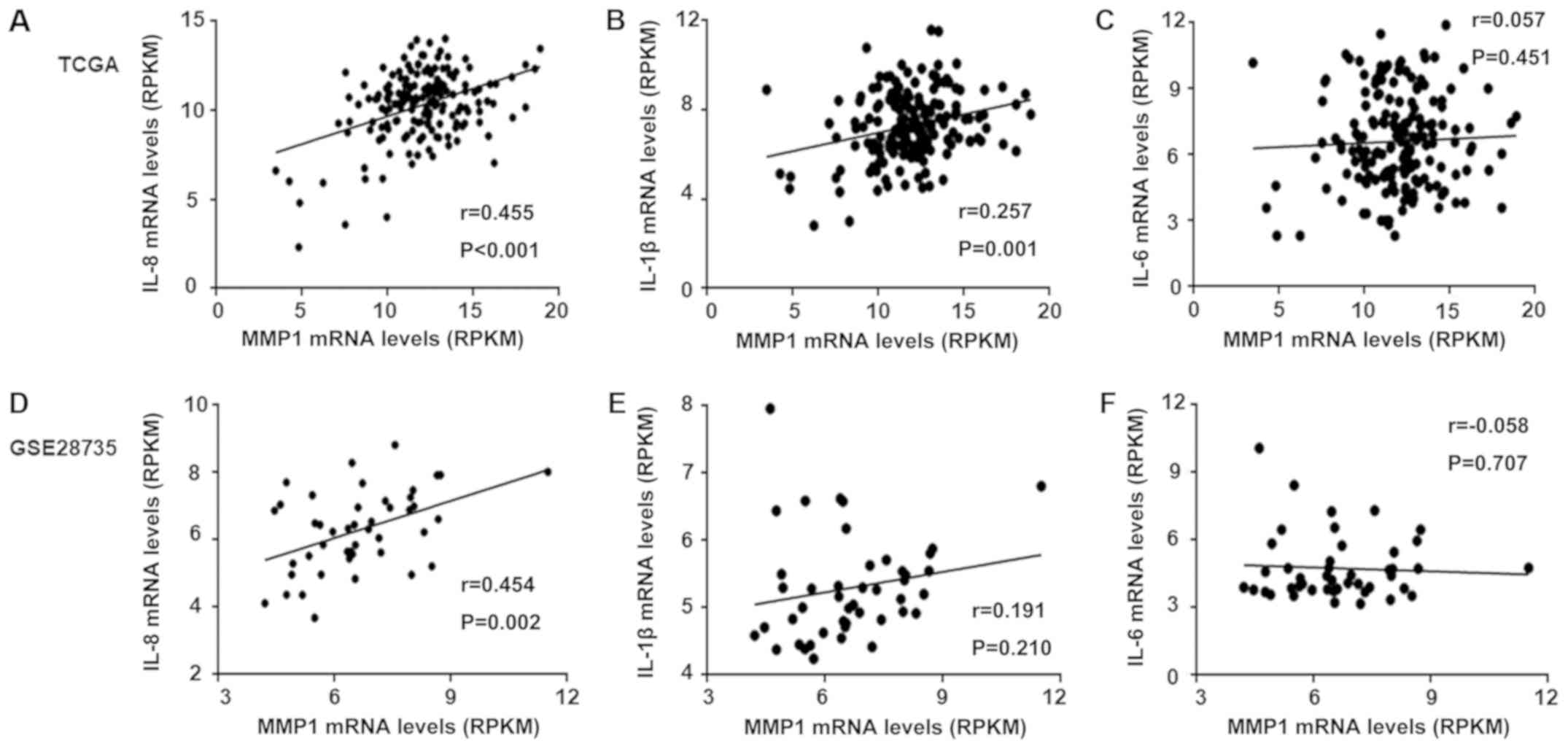
The results based on TCGA database (Fig. 4A-C) revealed that the correlation
between IL-8 and MMP1 (r=0.445) was higher than that with IL-1β
(r=0.257), and both were statistically significant. However, only
IL-8 exhibited significant correlation with MMP1 expression in the
GEO dataset GSE28735 (Fig. 4D, E and
F). Based on the results of the analysis from the two
databases, IL-8 was selected as an inducer of MMP1. As presented in
Fig. 5A, at 24, 36 and 48 h of
co-culturing with rIL-8, the expression of MMP1 was significantly
upregulated (all P<0.05). When the hsa-miR-623 mimic was
transfected into PANC-1 and BXPC-3 cells prior to treatment with
rIL-8, the induction of MMP1 was attenuated, whereas the
hsa-miR-623 inhibitor transfection further increased the MMP1
expression (Fig. 5B). In addition,
the expressions of TWIST1 and SNAI1 were increased following IL-8
treatment for 6, 12 and 24 h, and E-cadherin was decreased at 12
and 24 h (Fig. 5C). As the ERK
signaling pathway is commonly involved in mechanisms underlying
IL-8 function, and exerts pro-tumor activities, western blot assays
were subsequently performed to investigate whether hsa-miR-623
could suppress the phosphorylation of ERK. As hypothesized, the
transfection of hsa-miR-623 mimic not only decreased ERK
phosphorylation, but also decreased IL-8-induced TWIST1 and SNAI1
expression, and reversed the downregulation of E-cadherin. By
contrast, transfection of the hsa-miR-623 inhibitor exhibited the
opposite results (Fig. 5D). As
indicated by the phalloidin staining (Fig. 5E), the morphology of the miR-623
mimic-transfected PANC-1 and BXPC-3 cells displayed relatively
tight cell-cell adhesion, whereas miR-623 inhibitor-transfected
cells exhibited many new spike-like filopodia at their edges,
indicating a spindle-like and fibroblastic phenotype. These results
suggest that upregulation of miR-623 results in inhibition of the
epithelial-mesenchymal transition (EMT). Furthermore, the
hsa-miR-623 mimic attenuated IL-8-induced migration and invasion of
PANC-1 and BXPC-3 cells, whereas miR-623 inhibition resulted in
more aggressive features (Fig.
6).
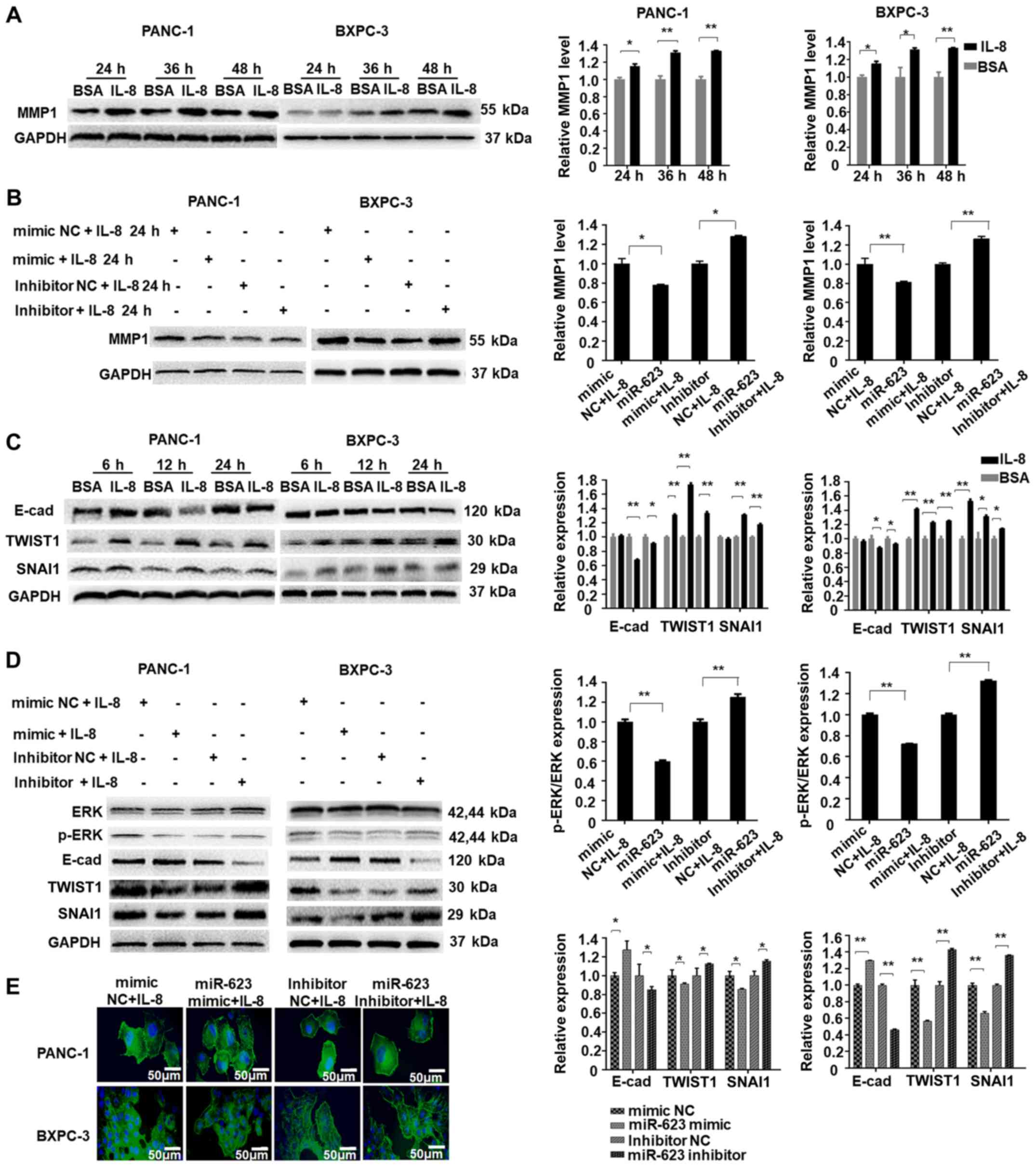 | Figure 5hsa-miR-623 attenuates IL-8-induced
MMP1 expression and the epithelial-mesenchymal transition in
pancreatic cancer cells. (A) Treatment with recombinant human IL-8
increased the levels of MMP1, whereas (B) hsa-miR-623 inhibitor led
to the upregulation of MMP1 levels. (C) Recombinant IL-8 treatment
increased the expression of TWIST1 and SNAI1, and decreased the
expression of E-cad in PANC-1 and BXPC-3 cells. (D) The hsa-miR-623
mimic inhibited the IL-8-induced upregulation of the
epithelial-mesenchymal transition by suppressing ERK
phosphorylation. (E) Cells were stained with phalloidin (green) and
counterstained with DAPI (blue). Magnification, ×400.
*P<0.05, **P<0.001. IL, interleukin;
MMP1, matrix metalloproteinase-1; miR, microRNA; E-cad, E-cadherin;
TWIST1, Twist-related protein 1; SNAI1, zinc finger protein SNAI1;
p-ERK, phosphorylated ERK; NC, negative control. |
hsa-miR-623 inhibits pancreatic cancer
cell metastasis in vivo
A xenograft nude mouse model was established to
further investigate whether hsa-miR-623 could suppress distant
metastasis of PDAC in vivo. The transfection efficiency
evaluated by qPCR demonstrated that Lenti-miR-623-GFP significantly
increased the expression of miR-623 compared with NC transfection
(Fig. 7A). The transfected PANC-1
cells were injected into the tail vein of nude mice to mimic the
spread of cancer cells throughout the blood. All animals survived
until the end time point of the experiment. As presented in
Fig. 7B and C, heightened
metastasis was observed in mice injected with rIL-8; however, when
hsa-miR-623 was overexpressed in PANC-1 cells, metastasis was
markedly inhibited in mice with or without IL-8 treatment. Compared
with that in the control mice, the mean fluorescence density of
lungs and livers in the hsa-miR-623 overexpression groups was
markedly lower (P<0.05), indicating decreased metastasis of
cancer cells. In addition, the number of lung and liver metastasis
nodules was also significantly decreased in the
hsa-miR-623-overexpression group compared with those in the control
group (Fig. 7C and D).
Furthermore, the expression of MMP1, TWIST1 and SNAI1 protein in
liver metastatic nodules in the hsa-miR-623 overexpression group
was also significantly decreased (Fig.
7E). These results were consistent with the in vitro
results, indicating that hsa-miR-623 attenuates IL-8-induced PDAC
metastasis in vitro and in vivo.
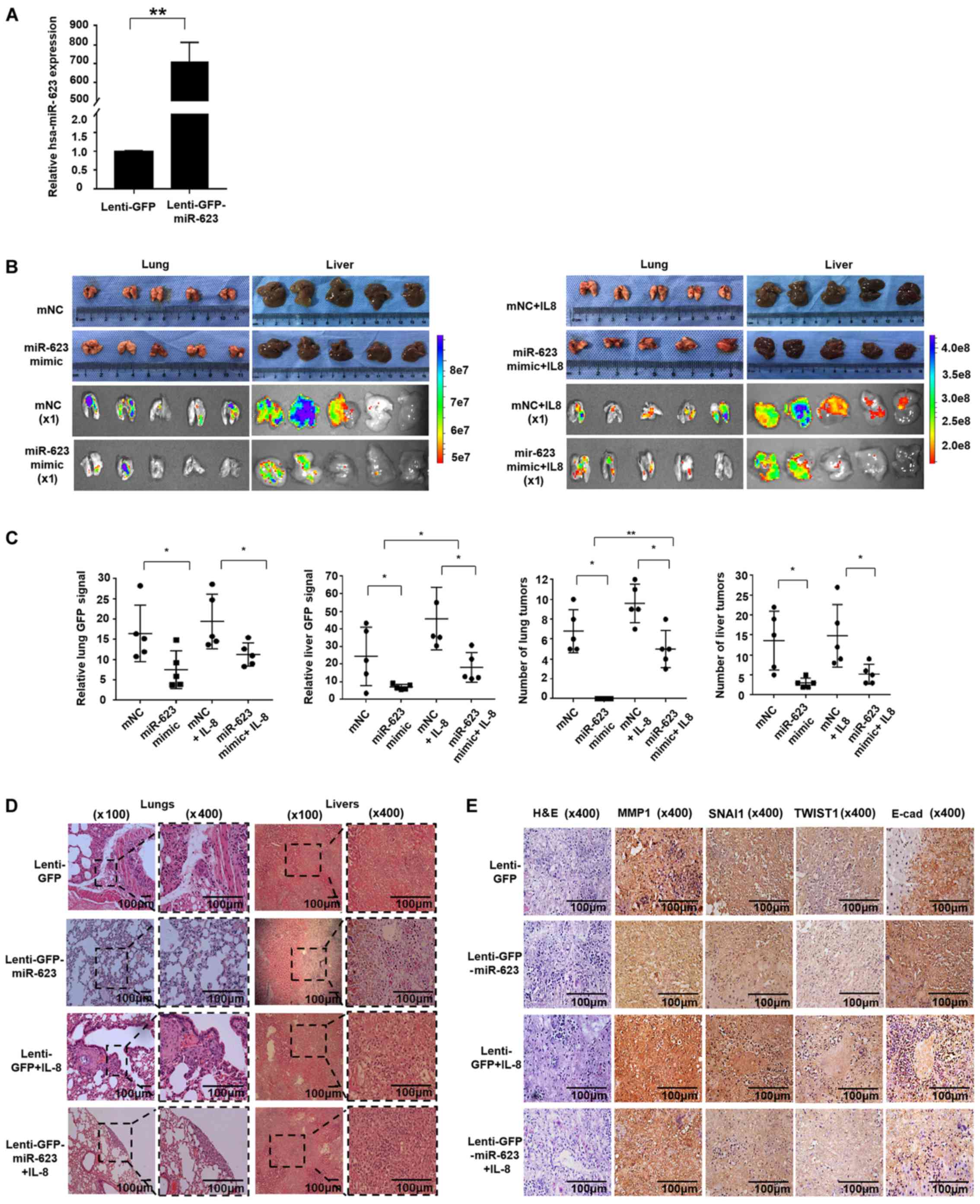 | Figure 7hsa-miR-623 inhibits pancreatic
cancer cell metastasis in vivo. A total of 2×107
PANC-1 cells pre-transfected with lenti-GFP or lenti-miR-623-GFP
were injected into the tail veins of 5-week-old nude mice.
Recombinant human IL-8 was administered via intraperitoneal
injection at a dosage of 100 ng/animal once a week for a period of
5 weeks. (A) The transfection efficiency was evaluated by reverse
transcription-quantitative PCR, and the results revealed that the
lenti-miR-623-GFP increased the expression of miR-623 to
706.43±73.17 times that of the lenti-GFP transfection (P<0.001).
Increased levels of distant metastasis were observed in mice that
had been treated with IL-8. The livers and lungs were isolated, and
metastatic lesions were evaluated by (B-C) fluorescence scanning
and (D) tissue section HE staining. Mice injected with PANC-1
lenti-miR-623-GFP cells exhibited decreased liver and lung
metastasis compared with those injected with PANC-1 lenti-GFP cells
with or without IL-8 treatment. (E) Immunochemical staining of
tumor sections. The expression of MMP1, TWIST1 and SNAI1 protein in
liver metastatic nodules from the hsa-miR-623-overexpression group
was decreased, and the E-cad protein expression increased. The data
are presented as the mean ± standard deviation (n=3).
*P<0.05, **P<0.001. miR, microRNA; GFP,
green fluorescent protein; IL, interleukin; HE, hematoxylin and
eosin; MMP1, matrix metalloproteinase-1; TWIST1, Twist-related
protein 1; SNAI1, zinc finger protein SNAI1; E-cad, E-cadherin;
mNC, mimic negative control. |
Discussion
The overexpression of MMP1 is associated with a poor
prognosis in pancreatic and breast cancer, and is a novel biomarker
for monitoring hepatocellular carcinoma in patients who have
received liver transplants (32).
MMP1 is a remarkable MMP family member due to its function as an
agonist of protease-activated receptor 1 (PAR1) (33), which is involved in a plethora of
pathophysiological processes, such as arterial stenosis and
atherosclerosis (34,35). In cancer, MMP1 activates PAR1 and
exerts several pro-tumor functions, including i) promoting tumor
metastasis via transendothelial migration by exerting enhanced
endothelial permeability (36,37),
ii) inducing protein activation to promote endothelial cell
angiogenesis (38), and iii) being
involved with the association between perineural invasion and
post-operative relapse in patients with PDAC. Our previous study
demonstrated that MMP1 mediates pancreatic cancer cell perineural
invasion in the early stages of disease via the MMP1/PAR1/substance
P/neurokinin 1 receptor paracrine loop (39). However, clinical trials using MMP
inhibitors combined with gemcitabine were unsuccessful in patients
with advanced pancreatic cancer (40,41).
Therefore, exploring the mechanism of MMP1 and selective MMP1
inhibitors rather than targeting all MMPs may be effective for the
treatment of cancer in the early stage of disease.
Ras mutations are detected in >90% of pancreatic
cancer cases at a relatively early stage. Mutated Ras results in
the continuous activation of the mitogen-activated protein kinase
(MAPK) pathway (ERK, JNK and p38 MAPK) and regulates the
transcription of transcription factor AP-1-mediated genes. A
previous study demonstrated that constitutive expression of MMP1 in
human pancreatic cancer cell lines was mediated by activation of
the JNK/AP-1 or ERK/AP-1 pathways, which could be suppressed by a
specific inhibitor of either JNK or MAPK kinase (42). In addition to constitutive
expression, MMP1 can be induced by cytokines that activate the MAPK
pathway, which has been well studied in rheumatoid arthritis and
joint damage (43). Similar to
inflammatory diseases, MMP1 expression in neoplastic cells is also
inducible, and exerts a pivotal function in the progression of
cancer (44). Taking into
consideration that MMP1 expression is affected by several factors,
it is conceivable that modulators that inhibit MMP1 expression at
the transcriptional and post-transcriptional levels will exert
potent anti-invasive effects on pancreatic cancer cells.
Mature miRNAs recognize the 3′-UTR of their target
mRNA, subsequently forming miRNA/mRNA/protein complexes that
incorporate into the RNA-induced silencing complex, which
ultimately results in mRNA cleavage (45). The present study screened miRNAs
that potentially modulate MMP1 and analyzed the correlation between
miRNAs and MMP1 mRNA in PDAC tissues, discovering that only the
expression of hsa-miR-623 exhibited a negative correlation with
MMP1 mRNA expression. By contrast, other miRNAs that have been
reported to target MMP1 in different types of non-tumor diseases
exhibited no correlation with MMP1 mRNA expression. These findings
may be explained by the heterogeneity of pancreatic cancer cells.
An RNA EMSA was subsequently used to demonstrate the direct and
specific interaction between hsa-miRNA-623 and its target within
the 3′-UTR of MMP1, and a luciferase reporter assay confirmed the
downregulation of MMP1 in response to hsa-miR-623 targeting its
3′-UTR. miRNAs also regulate gene expression indirectly, by weaving
a complex regulation network in cancer, as supported by the western
blotting results following cell transfection. The upregulation of
hsa-miRNA-623 not only decreased MMP1 protein expression, but also
suppressed the phosphorylation of ERK, which is part of one of the
essential pathways for AP1-dependent MMP1 transcription (42). Although the exact mechanism whereby
hsa-miRNA-623 inhibits the phosphorylation of ERK requires further
investigation, the results of the present study indicate that
hsa-miR-623 may downregulate MMP1 expression directly by
post-transcriptional degradation and indirectly by transcriptional
inhibition.
A previous study reported that hsa-miR-623 inhibits
the proliferation and metastasis of human lung adenocarcinoma by
targeting X-ray repair cross-complementing protein 5 (46). However, overexpression of
hsa-miR-623 exhibited no apparent influence on PANC-1 or BXPC-3
cell proliferation (data not shown); therefore, in the present
study, the primary focus was on the anti-metastatic effects of
hsa-miR-623 on PDAC cells. The EMT is a crucial driver of cancer
cell migration and invasion, which is regulated by various
molecular mechanisms, including pro-inflammatory factors and MMP
activity (47). SNAI1 and TWIST1
are two important transcription factors, referred to as EMT
transcription factors (EMT-TFs), which exert pivotal functions in
the process of EMT (48).
Exogenous factors that induce the expression of SNAI1 or TWIST1
(IL-8 and IL-6) and endogenous regulators (miRNAs) contribute to
cancer metastasis (49,50). The results of the present study
demonstrated that hsa-miR-623 is involved in this cellular/factor
network: First, hsa-miR-623 downregulated MMP1 by targeting the
3′-UTR. Secondly, IL-8 induced the expression of MMP1, SNAI1 and
TWIST1, whereas hsa-miR-623 attenuated the induction effect of IL-8
via the underlying mechanism of ERK phosphorylation suppression.
Since SNAI1 and TWIST1 are EMT-TFs that inhibit the expression of
E-cadherin, one of the most important EMT-regulating genes, through
E-boxes in the promoter region (51), hsa-miR-623 upregulated E-cadherin
expression and inhibited EMT. Finally, hsa-miR-623 inhibited PANC-1
and BXPC-3 cell invasion in vitro and in vivo.
IL-8 has been reported to induce MMPs, particularly
MMP2 and 9, which are overexpressed in various types of cancer,
whereas studies investigating IL-8 and MMP1 are limited. It appears
that IL-8 and MMP1 are mutual promoting factors, and IL-8 induces
MMP1 expression via IL-8/androgen receptor signaling in bladder
cancer cells (8); in addition,
IL-8/C-X-C chemokine receptor types 1 and 2 (CXCR1/2) activates
ERK1/2 (52) and may upregulate
MMP1 expression via ERK/AP-1 signaling. On the other hand, in
ovarian cancer cells, MMP1 induces IL-8 production in a
PAR1-dependent manner; subsequently, secreted IL-8 activates
CXCR1/2 receptors on endothelial cells, thereby causing
angiogenesis and metastasis (53).
A database analysis verified a positive correlation
between IL-8 and MMP1, and a negative correlation between MMP1 and
the expression of hsa-miR-623 in PDAC tissues. Therefore, the
MMP1-PAR1/IL-8-CXCR1/2 paracrine pathway may be therapeutically
relevant for the treatment of pancreatic cancer (22,54).
As a direct suppressor of MMP1, hsa-miR-623 may consequently block
this paracrine loop and therefore exert antitumor activity by
modulating multiple steps.
In summary, the present study demonstrated that
hsa-miR-623 binds to a specific sequence located within the 3′-UTR
of MMP1 and suppresses MMP1 expression. In addition, hsa-miR-623
suppressed the IL-8-induced expression of MMP1 and several EMT
functional proteins, consequently inhibiting pancreatic cancer
metastasis. These results may provide insights pertaining to a
novel epigenetic mechanism for regulating the expression of the
MMP1 gene and suggest that hsa-miR-623 may be a novel adjuvant
therapeutic target to prevent pancreatic cancer metastasis.
Funding
This study was supported by the National Natural
Science Foundation of China (grant nos. 81872348 and 81502503) and
the Natural Science Foundation of Guangdong Province, China (grant
no. 2016A030310191).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YuC, SP and CH carried out the experimental work,
data collection and interpretation. HC and YL participated in the
animal experiments. HS, YS and LZ participated in the study design
and drafted the manuscript. YiC carried out data collection and
analysis. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Animal
Care and Use Committee of Sun Yat-Sen University, Guangzhou, China
(approval no. 180604).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
ECM
|
extracellular matrix
|
|
EMT-TFs
|
epithelial- mesenchymal transition
transcription factors
|
|
EMSA
|
electrophoretic mobility shift
assay
|
|
FBS
|
fetal bovine serum
|
|
IL-8
|
interleukin-8
|
|
MMP1
|
matrix metalloproteinase-1
|
|
MFE
|
minimum free energy
|
|
miR/miRNA
|
microRNA
|
|
NC
|
negative control
|
|
PDAC
|
pancreatic ductal adenocarcinoma
|
|
PAR1
|
protease-activated receptor 1
|
|
TNF-α
|
tumor necrosis factor-α
|
|
TCGA
|
The Cancer Genome Atlas
|
|
3′-UTR
|
3′-untranslated region
|
Acknowledgments
Not applicable.
References
|
1
|
Kamisawa T, Wood LD, Itoi T and Takaori K:
Pancreatic cancer. Lancet. 388:73–85. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kamisawa T, Isawa T, Koike M, Tsuruta K
and Okamoto A: Hematogenous metastases of pancreatic ductal
carcinoma. Pancreas. 11:345–349. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Conroy T, Bachet JB, Ayav A, Huguet F,
Lambert A, Caramella C, Maréchal R, Van Laethem JL and Ducreux M:
Current standards and new innovative approaches for treatment of
pancreatic cancer. Eur J Cancer. 57:10–22. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bonnans C, Chou J and Werb Z: Remodelling
the extracellular matrix in development and disease. Nat Rev Mol
Cell Biol. 15:786–801. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shen TC, Chang WS, Tsai CW, Chao CY, Lin
YT, Hsiao CL, Hsu CL, Chen WC, Hsia TC and Bau DT: The contribution
of matrix metalloproteinase-1 promoter genotypes in Taiwan lung
cancer risk. Anticancer Res. 38:253–257. 2018.
|
|
6
|
Padala C, Tupurani MA, Puranam K, Gantala
S, Shyamala N, Kondapalli MS, Gundapaneni KK, Mudigonda S, Galimudi
RK, Kupsal K, et al: : Synergistic effect of collagenase-1 (MMP1),
stromelysin-1 (MMP3) and gelatinase-B (MMP9) gene polymorphisms in
breast cancer. PLoS One. 12:e01844482017. View Article : Google Scholar
|
|
7
|
Hu W, Ye Y, Yin Y, Sang P, Li L, Wang J,
Wan W, Li R, Bai X, Xie Y, et al: Association of matrix
metalloprotease 1, 3, and 12 polymorphisms with rheumatic heart
disease in a Chinese Han population. BMC Med Genet. 19:272018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ou Z, Wang Y, Liu L, Li L, Yeh S, Qi L and
Chang C: Tumor microenvironment B cells increase bladder cancer
metastasis via modulation of the IL-8/androgen receptor (AR)/MMPs
signals. Oncotarget. 6:26065–26078. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang G, Im HJ and Wang JH: Repetitive
mechanical stretching modulates IL-1beta induced COX-2, MMP-1
expression, and PGE-2 production in human patellar tendon
fibroblasts. Gene. 363:166–172. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li R, Hebert JD, Lee TA, Xing H,
Boussommier-Calleja A, Hynes RO, Lauffenburger DA and Kamm RD:
Macrophage-secreted TNFα and TGFβ1 influence migration speed and
persistence of cancer cells in 3D tissue culture via independent
pathways. Cancer Res. 77:279–290. 2017. View Article : Google Scholar
|
|
11
|
Croce CM: Oncogenes and cancer. N Engl J
Med. 358:502–511. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rozario T and DeSimone DW: The
extracellular matrix in development and morphogenesis: A dynamic
view. Dev Biol. 341:126–140. 2010. View Article : Google Scholar :
|
|
13
|
Marchesi F, Regazzo G, Palombi F,
Terrenato I, Sacconi A, Spagnuolo M, Donzelli S, Marino M, Ercolani
C, Di Benedetto A, et al: Serum miR-22 as potential non-invasive
predictor of poor clinical outcome in newly diagnosed, uniformly
treated patients with diffuse large B-cell lymphoma: An explorative
pilot study. J Exp Clin Cancer Res. 37:952018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Samir N, Matboli M, El-Tayeb H, El-Tawdi
A, Hassan MK, Waly A, El-Akkad HAE, Ramadan MG, Al-Belkini TN,
El-Khamisy S, et al: Competing endogenous RNA network crosstalk
reveals novel molecular markers in colorectal cancer. J Cell
Biochem. 119:6869–6881. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Frampton AE, Castellano L, Colombo T,
Giovannetti E, Krell J, Jacob J, Pellegrino L, Roca-Alonso L, Funel
N, Gall TM, et al: Integrated molecular analysis to investigate the
role of microRNAs in pancreatic tumour growth and progression.
Lancet. 385(Suppl 1): S372015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang Y, Lin X, Zhang L, Hong W and Zeng
K: MicroRNA-222 regulates the viability of fibroblasts in
hypertrophic scars via matrix metalloproteinase 1. Exp Ther Med.
15:1803–1808. 2018.PubMed/NCBI
|
|
17
|
Kim KH, Jung JY, Son ED, Shin DW, Noh M
and Lee TR: miR-526b targets 3′ UTR of MMP1 mRNA. Exp Mol Med.
47:e1782015. View Article : Google Scholar
|
|
18
|
Wang Y, Pang X, Wu J, Jin L, Yu Y, Gobin R
and Yu J: MicroRNA hsa-let-7b suppresses the odonto/osteogenic
differentiation capacity of stem cells from apical papilla by
targeting MMP1. J Cell Biochem. 119:6545–6554. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou B, Zhu H, Luo H, Gao S, Dai X, Li Y
and Zuo X: MicroRNA-202-3p regulates scleroderma fibrosis by
targeting matrix metalloproteinase 1. Biomed Pharmacother.
87:412–418. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu D, Green B, Marrone A, Guo Y, Kadlubar
S, Lin D, Fuscoe J, Pogribny I and Ning B: Suppression of CYP2C9 by
microRNA hsa-miR-128-3p in human liver cells and association with
hepatocellular carcinoma. Sci Rep. 5:85342015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen Y, Zeng L, Wang Y, Tolleson WH, Knox
B, Chen S, Ren Z, Guo L, Mei N, Qian F, et al: The expression,
induction and pharmacological activity of CYP1A2 are
post-transcriptionally regulated by microRNA hsa-miR-132-5p.
Biochem Pharmacol. 145:178–191. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen L, Fan J, Chen H, Meng Z, Chen Z,
Wang P and Liu L: The IL-8/CXCR1 axis is associated with cancer
stem cell-like properties and correlates with clinical prognosis in
human pancreatic cancer cases. Sci Rep. 4:59112014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jaillon S, Peri G, Delneste Y, Frémaux I,
Doni A, Moalli F, Garlanda C, Romani L, Gascan H, Bellocchio S, et
al: The humoral pattern recognition receptor PTX3 is stored in
neutrophil granules and localizes in extracellular traps. J Exp
Med. 204:793–804. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-ΔΔC(T)) method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
25
|
Zhang G, Schetter A, He P, Funamizu N,
Gaedcke J, Ghadimi BM, Ried T, Hassan R, Yfantis HG, Lee DH, et al:
: DPEP1 inhibits tumor cell invasiveness, enhances chemosensitivity
and predicts clinical outcome in pancreatic ductal adenocarcinoma.
PLoS One. 7:e315072012. View Article : Google Scholar
|
|
26
|
Donahue TR, Tran LM, Hill R, Li Y,
Kovochich A, Calvopina JH, Patel SG, Wu N, Hindoyan A, Farrell JJ,
et al: Integrative survival-based molecular profiling of human
pancreatic cancer. Clin Cancer Res. 18:1352–1363. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zeng L, Chen Y, Wang Y, Yu LR, Knox B,
Chen J, Shi T, Chen S, Ren Z, Guo L, et al: : MicroRNA
hsa-miR-370-3p suppresses the expression and induction of CYP2D6 by
facilitating mRNA degradation. Biochem Pharmacol. 140:139–149.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang Y, Yu D, Tolleson WH, Yu LR, Green B,
Zeng L, Chen Y, Chen S, Ren Z, Guo L, et al: A systematic
evaluation of microRNAs in regulating human hepatic CYP2E1. Biochem
Pharmacol. 138:174–184. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jin Y, Yu D, Tolleson WH, Knox B, Wang Y,
Chen S, Ren Z, Deng H, Guo Y and Ning B: MicroRNA hsa-miR-25-3p
suppresses the expression and drug induction of CYP2B6 in human
hepatocytes. Biochem Pharmacol. 113:88–96. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Trenkmann M, Brock M, Gay RE, Michel BA,
Gay S and Huber LC: Tumor necrosis factor α-induced microRNA-18a
activates rheumatoid arthritis synovial fibroblasts through a
feedback loop in NF-κB signaling. Arthritis Rheum. 65:916–927.
2012. View Article : Google Scholar
|
|
31
|
Yoon HY, Lee EG, Lee H, Cho IJ, Choi YJ,
Sung MS, Yoo HG and Yoo WH: Kaempferol inhibits IL-1β-induced
proliferation of rheumatoid arthritis synovial fibroblasts and the
production of COX-2, PGE2 and MMPs. Int J Mol Med. 32:971–977.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sánchez-Lorencio MI, Saenz L, Ramirez P,
Villalba-López F, de la Orden V, Mediero-Valeros B, Revilla Nuin B,
Gonzalez MR, Cascales-Campos PA, Ferreras-Martínez D, et al: Matrix
metalloproteinase 1 as a novel biomarker for monitoring
hepatocellular carcinoma in liver transplant patients. Transplant
Proc. 50:623–627. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Trivedi V, Boire A, Tchernychev B,
Kaneider NC, Leger AJ, O'Callaghan K, Covic L and Kuliopulos A:
Platelet matrix metalloprotease-1 mediates thrombogenesis by
activating PAR1 at a cryptic ligand site. Cell. 137:332–343. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Austin KM, Nguyen N, Javid G, Covic L and
Kuliopulos A: Noncanonical matrix
metalloprotease-1-protease-activated receptor-1 signaling triggers
vascular smooth muscle cell dedifferentiation and arterial
stenosis. J Biol Chem. 288:23105–23115. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rana R, Huang T, Koukos G, Fletcher EK,
Turner SE, Shearer A, Gurbel PA, Rade JJ, Kimmelstiel CD, Bliden
KP, et al: Noncanonical matrix metalloprotease 1-protease-activated
receptor 1 signaling drives progression of atherosclerosis.
Arterioscler Thromb Vasc Biol. 38:1368–1380. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Juncker-Jensen A, Deryugina EI, Rimann I,
Zajac E, Kupriyanova TA, Engelholm LH and Quigley JP: Tumor MMP-1
activates endothelial PAR1 to facilitate vascular intravasation and
metastatic dissemination. Cancer Res. 73:4196–4211. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Goerge T, Barg A, Schnaeker EM, Poppelmann
B, Shpacovitch V, Rattenholl A, Maaser C, Luger TA, Steinhoff M and
Schneider SW: Tumor-derived matrix metalloproteinase-1 targets
endothelial proteinase-activated receptor 1 promoting endothelial
cell activation. Cancer Res. 66:7766–7774. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gopal SK, Greening DW, Zhu HJ, Simpson RJ
and Mathias RA: Transformed MDCK cells secrete elevated MMP1 that
generates LAMA5 fragments promoting endothelial cell angiogenesis.
Sci Rep. 6:283212016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Huang C, Li Y, Guo Y, Zhang Z, Lian G,
Chen Y, Li J, Su Y, Li J, Yang K, et al: MMP1/PAR1/SP/NK1R
paracrine loop modulates early perineural invasion of pancreatic
cancer cells. Theranostics. 8:3074–3086. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bramhall SR, Rosemurgy A, Brown PD, Bowry
C and Buckels JA; Marimastat Pancreatic Cancer Study Group:
Marimastat as first-line therapy for patients with unresectable
pancreatic cancer: A randomized trial. J Clin Oncol. 19:3447–3455.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bramhall SR, Schulz J, Nemunaitis J, Brown
PD, Baillet M and Buckels JA: A double-blind placebo-controlled,
randomised study comparing gemcitabine and marimastat with
gemcitabine and placebo as first line therapy in patients with
advanced pancreatic cancer. Br J Cancer. 87:161–167. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Endo H, Watanabe T, Sugioka Y, Niioka M,
Inagaki Y and Okazaki I: Activation of two distinct MAPK pathways
governs constitutive expression of matrix metalloproteinase-1 in
human pancreatic cancer cell lines. Int J Oncol. 35:1237–1245.
2009.PubMed/NCBI
|
|
43
|
Maksymowych WP, van der Heijde D, Allaart
CF, Landewé R, Boire G, Tak PP, Gui Y, Ghahary A, Kilani R and
Marotta A: 14-3-3η is a novel mediator associated with the
pathogenesis of rheumatoid arthritis and joint damage. Arthritis
Res Ther. 16:R992014. View
Article : Google Scholar
|
|
44
|
Ha NH, Park DG, Woo BH, Kim DJ, Choi JI,
Park BS, Kim YD, Lee JH and Park HR: Porphyromonas gingivalis
increases the invasiveness of oral cancer cells by upregulating
IL-8 and MMPs. Cytokine. 86:64–72. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gregory RI, Chendrimada TP, Cooch N and
Shiekhattar R: Human RISC couples microRNA biogenesis and
posttranscriptional gene silencing. Cell. 123:631–640. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wei S, Zhang ZY, Fu SL, Xie JG, Liu XS, Xu
YJ, Zhao JP and Xiong WN: Hsa-miR-623 suppresses tumor progression
in human lung adenocarcinoma. Cell Death Dis. 8:e28292017.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Nieto MA, Huang RY, Jackson RA and Thiery
JP: Emt: 2016. Cell. 166:21–45. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Onder TT, Gupta PB, Mani SA, Yang J,
Lander ES and Weinberg RA: Loss of E-cadherin promotes metastasis
via multiple downstream transcriptional pathways. Cancer Res.
68:3645–3654. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Dominguez C, David JM and Palena C:
Epithelial-mesenchymal transition and inflammation at the site of
the primary tumor. Semin Cancer Biol. 47:177–184. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ding XM: MicroRNAs: Regulators of cancer
metastasis and epithelial-mesenchymal transition (EMT). Chin J
Cancer. 33:140–147. 2014. View Article : Google Scholar :
|
|
51
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial- mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Nasser MW, Raghuwanshi SK, Grant DJ, Jala
VR, Rajarathnam K and Richardson RM: Differential activation and
regulation of CXCR1 and CXCR2 by CXCL8 monomer and dimer. J
Immunol. 183:3425–3432. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Agarwal A, Tressel SL, Kaimal R, Balla M,
Lam FH, Covic L and Kuliopulos A: Identification of a
metalloprotease-chemokine signaling system in the ovarian cancer
microenvironment: Implications for antiangiogenic therapy. Cancer
Res. 70:5880–5890. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Queiroz KC, Shi K, Duitman J, Aberson HL,
Wilmink JW, van Noesel CJ, Richel DJ and Spek CA:
Protease-activated receptor-1 drives pancreatic cancer progression
and chemoresistance. Int J Cancer. 135:2294–2304. 2014. View Article : Google Scholar : PubMed/NCBI
|