Introduction
Alternative splicing, also referred to as
differential splicing, refers to the process of generating various
isoforms of an mRNA precursor by several approaches, i.e., by
selecting diverse splice site combinations. The resulting protein
products may exhibit distinguishable or antagonistic functional and
structural characteristics, or they may yield various phenotypes in
the same cell type due to differences in their expression levels
(1,2).
The splicing isoforms of certain genes may serve as
drivers for the development of tumors (3-7).
Additionally, the genomes of tumor cells have more extensive
alternative splicing mechanisms compared with normal cells. In most
types of tumors, the expression products of genes may vary due to
changes in alternative splicing, which is associated with tumor
development, metastasis (8-12)
and resistance to treatment (7,13-15).
Additionally, alternative splicing events in tumors may act as
diagnostic and prognostic molecular biomarkers (16-19),
and they may also be used as targets for cancer treatment (20-24).
Alternative splicing events are regulated by
splicing factors that act as oncogenes or tumor suppressor genes
(25). The development of drugs
targeting splicing factors is a new research focus in cancer
therapy (26-32), and several studies have attempted
to distinguish cancers based on the presence of abnormal RNA
splicing (10,33). Furthermore, disturbances of
alterative splicing are reportedly involved in the development of
cancers. In addition, several survival-associated splicing events
that are unique to cancer have been identified (34). RNA-binding proteins, which function
as regulatory elements of splicing events, have also been
associated with cancer development (35-37).
Hepatocellular carcinoma (HCC) is the fifth most
common type of cancer with the third highest mortality rate
worldwide. Its occurrence has been associated with viral infection,
aflatoxin exposure, and other factors; in addition, a number of
studies have demonstrated that alternative splicing also plays an
important role in HCC (38-43).
However, these studies focused on the associations between the
alternative splicing events of only a few genes and HCC, whereas
only few studies have analyzed alternative splicing events in HCC
using large datasets. Tremblay et al (44) analyzed RNA sequencing (RNA-Seq)
data from The Cancer Genome Atlas (TCGA) and found a large number
of differential alternative splicing events between HCC and
corresponding paracancerous tissues. Hepatitis B and C virus
infections also affect alternative splicing events in HCC (44). Accordingly, alternative splicing is
considered to play an important role in the occurrence of HCC.
However, to the best of our knowledge, no study has yet examined
the association between mRNA splicing and the prognosis of HCC
based on TCGA SpliceSeq data. Therefore, by using TCGA SpliceSeq
data, the present study analyzed the associations among alternative
splicing events, splicing factors, and the survival of patients
with HCC, with the aim of identifying splicing events that may
serve as new molecular targets for the prognosis of HCC.
Materials and methods
Assortment of alternative splicing event
data
The alternative splicing event profiles of HCC
patients were downloaded from TCGA SpliceSeq (45), which is a resource for the
investigation of transcript splicing patterns and splicing event
details based on TCGA covering quantified introns or exons.
Information on percent-splice-in (PSI), the ratio of normalized
read counts indicating the inclusion of a transcript element over
the total normalized reads for that event, was collected from the
database. PSI was calculated as the ratio of reading densities of
inclusions to the sum of the reading densities of inclusion and
exclusion. PSI values range from 0 to 100%. Only samples with PSI
values >90% were downloaded. Simultaneously, clinical data were
also obtained from TCGA. A total of 7 different alternative
splicing events were obtained: Exon skips (ESs), retained introns
(RIs), mutually exclusive exons (MEs), alternate donor sites (ADs),
alternate acceptor sites (AAs), alternate promoters (APs), and
alternate terminators(ATs).
Survival analysis and production of a
prognostic signature
A total of 371 HCC patients in TCGA were used to
select survival-related alternative splicing events. The
association between alternative splicing events and overall
survival (OS) was evaluated using univariate Cox regression
analysis. The most significant prognostic alternative splicing
events (P<0.0001) were subjected to multivariate Cox regression
analysis. The area under the curve (AUC) of a time-dependent
receiver operating characteristic (ROC) curve was also calculated,
which has been used widely to compare the ability of prognostic
predictors. All survival analyses were conducted using the
'survival' and 'survival ROC' packages in R software. The OS rates
of patients with HCC grouped by high- and low-risk alternative
splicing events were plotted using Kaplan-Meier plots and the
differences between groups were analyzed using log-rank tests.
UpSet plot and gene-annotation enrichment
analysis
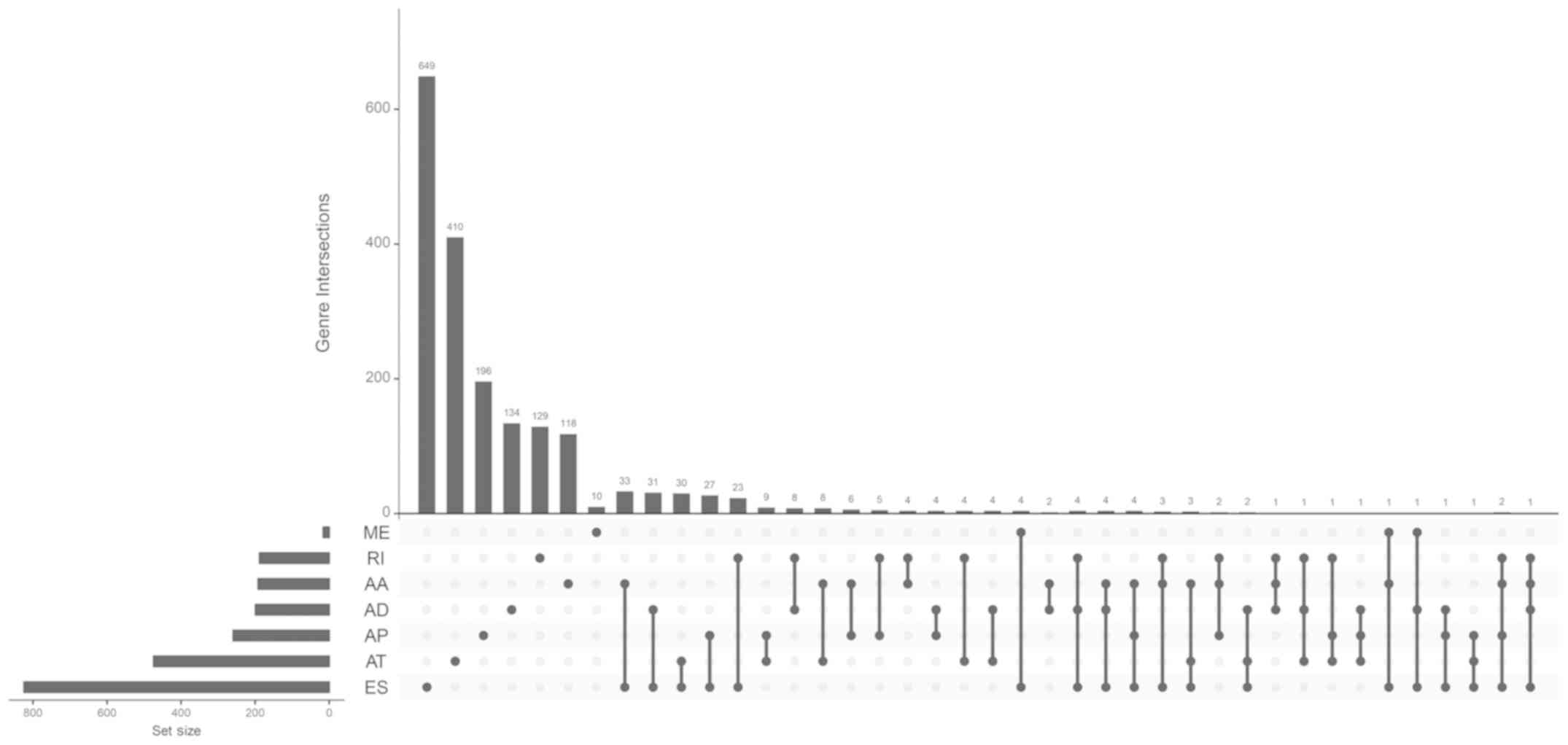
An UpSet plot, a novel visualization technique for
the quantitative analysis of interactive sets, was used to analyze
the intersections between the 7 types of alternative splicing.
Gene-annotation enrichment analyses were used in the
'clusterProfiler' package in R to annotate and visualize biological
process terms and Kyoto Encyclopedia of Genes and Genomes (KEGG)
pathways of survival-associated alternative splicing genes based on
the criterion of an adjusted P-value of <0.05.
Splicing factor genes
Splicing factor genes were collected from the
SpliceAid 2 database (http://193.206.120.249/splicing_tissue.html).
Subsequently, the expression profiles of splicing factor genes were
extracted from TCGA. Cox univariate regression analysis was
performed, and genes significantly associated with OS (P<0.001)
were retained. An analysis of the 32-gene splicing factors was also
incorporated by using the ProgGene (http://genomics.jefferson.edu/proggene/) database
(46). The database helps users
perform a combined analysis for a list of genes and generate a
prognostic signature based on the imputed genes using Cox
proportional hazard analysis. The 32 splicing factors were input
and the TCGA database was selected for survival analysis. Student
t-test analysis was conducted to estimate the differences of
splicing factors between tumor and non-tumor samples.
Splicing correlation network construction
and hub splicing factor genes
The correlation between the expression of
prognosis-related splicing factor genes and the PSI values of the
most significant survival-associated alternative splicing events
were analyzed by Pearson's tests. The splicing events-factors axes
were submitted to correlation network construction for P-values
<0.05 and a Pearson's correlation coefficient >0.3. The
correlation plots were generated by Cytoscape (version 3.5.1). Hub
splicing factor genes were identified according to the numerical
degree of each node and edge. The expression level of hub splicing
factors between tumor and non-tumor tissues were calculated based
on data from TCGA. The data were transformed to the form of
log2(count+1) and visually displayed by OriginPro 2017 (OriginLab
Corp.). Furthermore, calibration plots were generated to evaluate
model calibration by using 'rms' package in R software (https://CRAN.R-project.org/package=rms).
Results
Identification of mRNA splicing event
profiles in HCC
The alternative splicing event profiles of 371 HCC
patients were obtained from TCGA SpliceSeq. In total, 2,6210 mRNA
splicing events in 7,727 genes were collected, which comprised
8,524 ESs in 4,120 genes, 1,306 RIs in 1,879 genes, 4,359 APs in
1,775 genes, 7,736 ATs in 3,379 genes, 1,675 ADs in 1,205genes,
1,936 AAs in 1,451 genes, and 102 MEs in 101 genes. These results
also indicated that one gene may have several types of mRNA
splicing events, and ES was the predominant type.
Prognosis-related alternative splicing
events in HCC
The association of these alternative splicing events
with OS was studied by univariate survival analysis, and a total of
3,082 survival-associated alternative splicing events were detected
in HCC (P<0.05). We observed that one gene may have two or more
events that are significantly associated with survival. Thus, an
intersection visualization plot was generated via UpSet (Fig. 1). The top significant
survival-associated genes in HCC (P<0.0001) were piped to
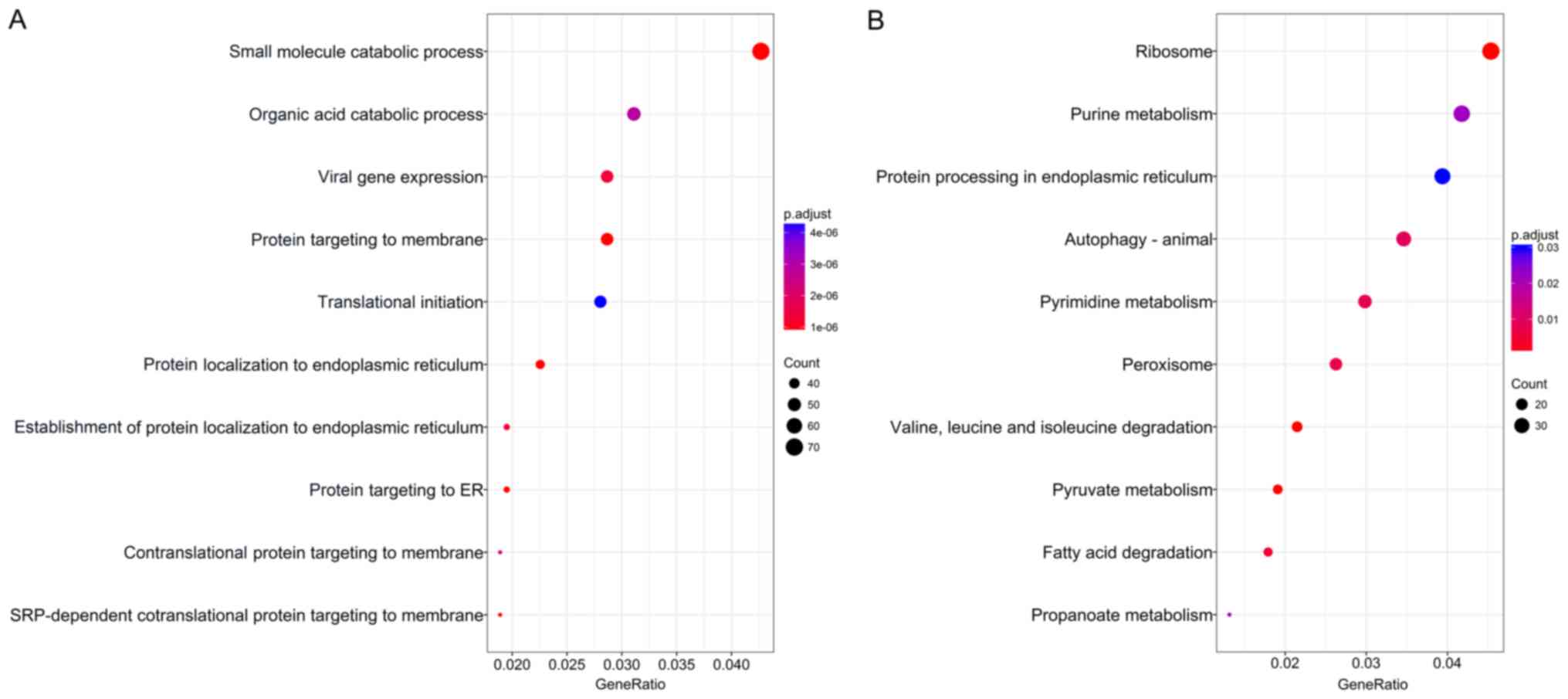
annotate and visualize biological process terms and KEGG pathways.
The biological process terms of these genes were most enriched in
small molecule catabolic process, organic acid catabolic process,
and viral gene expression (Fig.
2). KEGG pathway analysis indicated that these genes were
mainly enriched in ribosome, purine metabolism, protein processing
in endoplasmic reticulum, and autophagy.
Prognostic indicators for HCC
patients
The top significant survival-associated alternative
splicing events in the 7 types of alternative splicing were
selected as candidate prognostic factors. Multivariate Cox
regression with prognostic model construction was then applied to
develop a computational model for HCC prognosis based on each
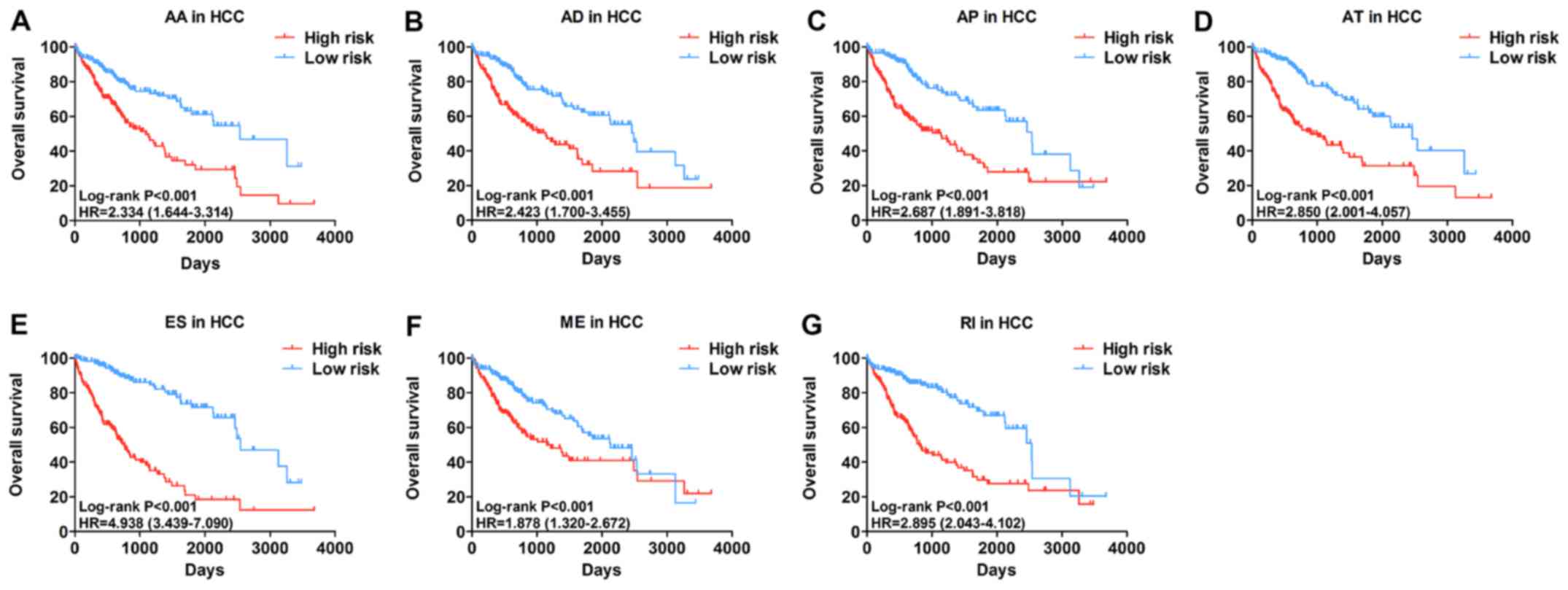
individual splicing type. HCC patients were divided into subgroups
with significantly different prognoses (Fig. 3) based on the 7 types of
alternative splicing (Table I).
These independent prognostic alternative splicing events were
analyzed further to build the final prognostic computational model
using a PI for HCC. Time-dependent ROC curve analyses were also
applied to compare the efficiency of these prognostic models
(Figs. 4 and 5). The final PI containing the most
significant candidate alternative splicing events exhibited the
best performance at distinguishing favorable or poor survival in
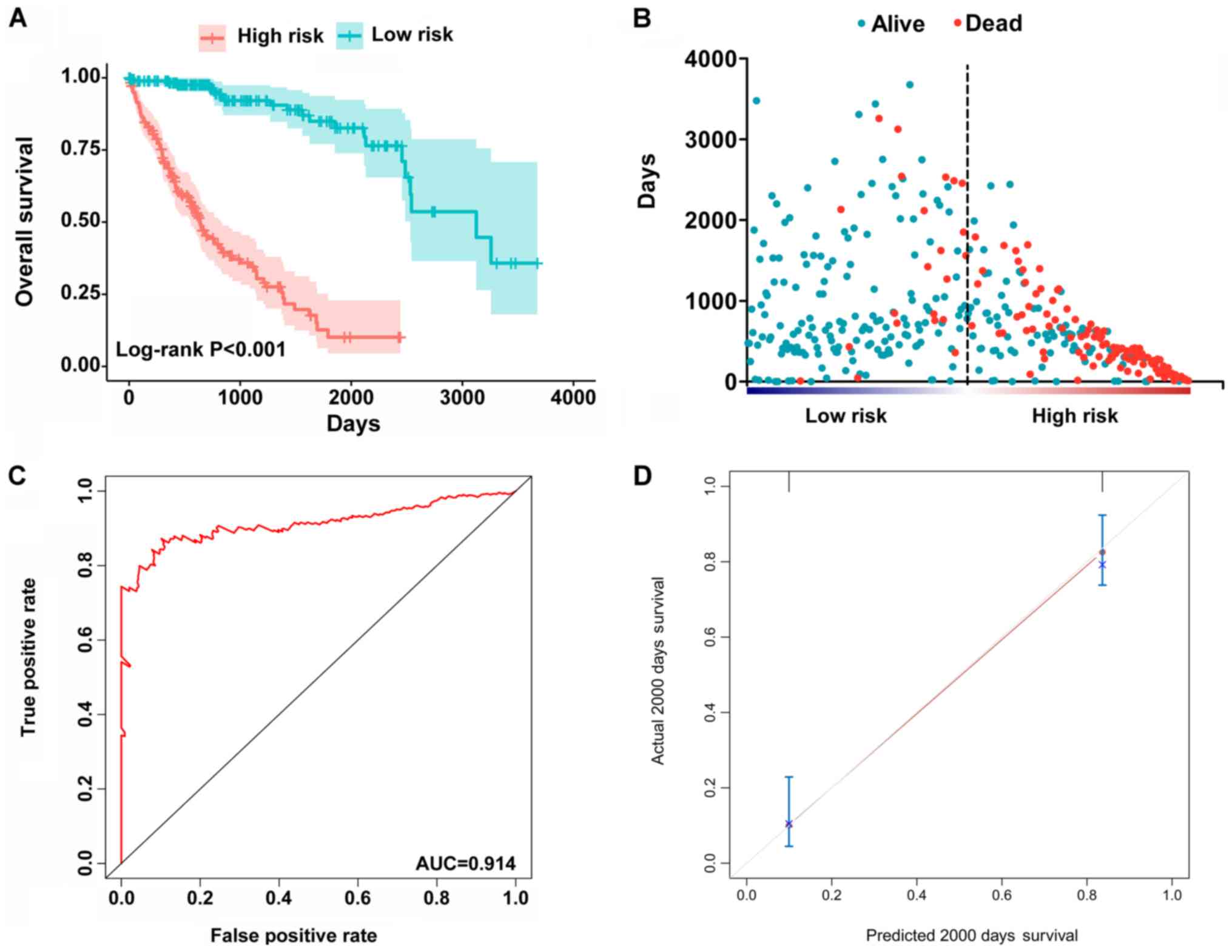
patients (Fig. 6). Patients in the
high-risk group had significantly worse survival compared with
those in the low-risk group [hazard ratio (HR)=12.904, 95%
confidence interval (CI): 7.513-22.161, P<0.001; Fig. 6A and B]. Furthermore, the PI
remained an independent prognostic indicator for HCC patients in
multivariate analyses after other clinicopathological
characteristics were adjusted (HR=16.541, 95% CI: 7.660-35.717,
P<0.001; Table II). ROC curves
confirmed that the final prognostic predictor with all types of
alternative splicing had the highest efficiency (Fig. 6C) with an AUC of 0.914. The
specific genes involved in the final models are listed in Table III. The calibration curves
exhibited good consistency between the predicted and actual
survival probability (Fig.
6D).
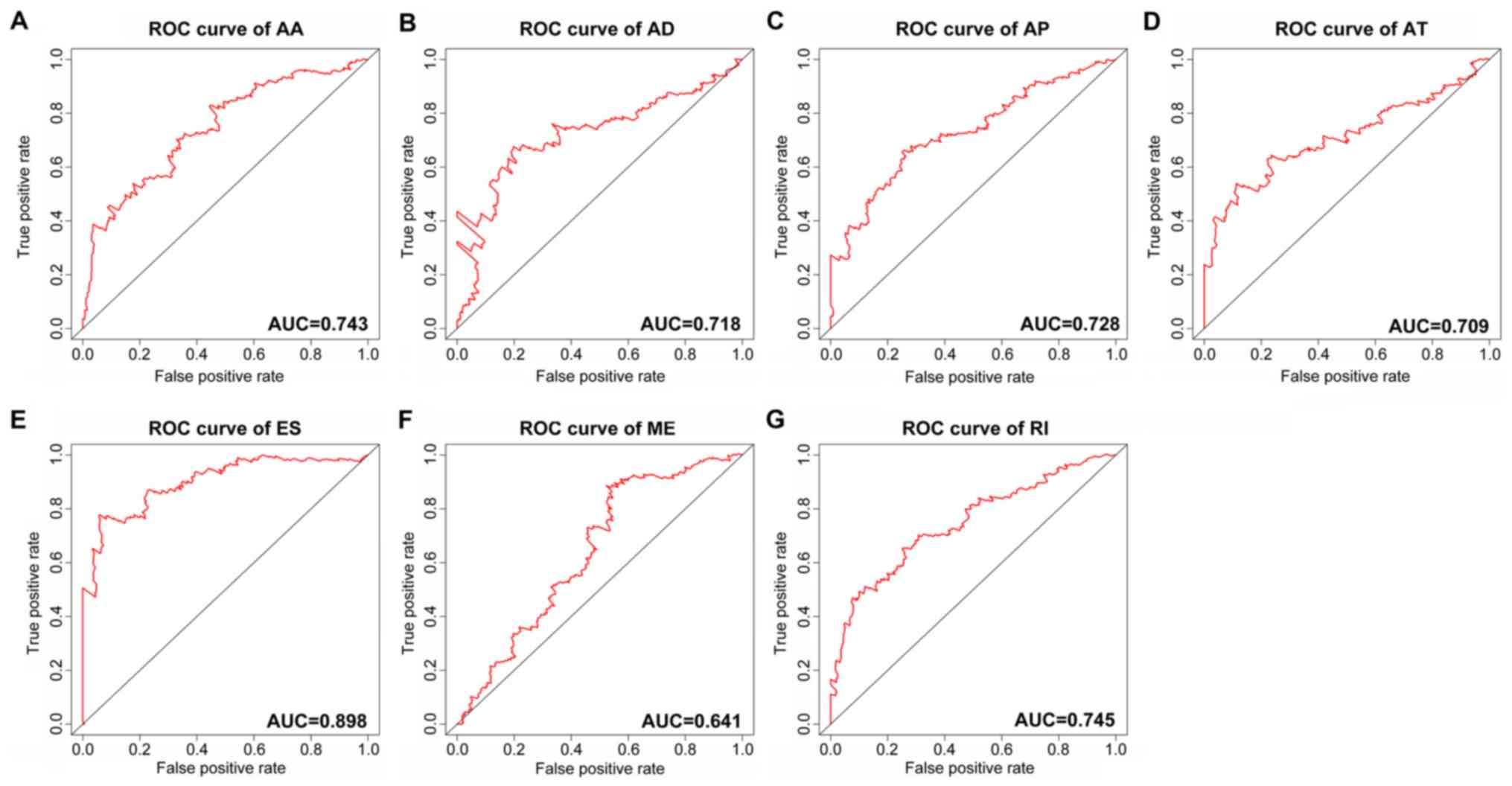 | Figure 4ROC curves of the PI built using the
7 types of alternative splicing in HCC. (A) AA; (B) AD; (C) AP; (D)
AT; (E) ES; (F) ME; and (G) RI. ROC, receiver operating
characteristic; AUC, area under the ROC curve; PI, prognostic
index; HCC, hepatocellular carcinoma; AA, alternate acceptor sites;
AD, alternate donor sites; AP, alternate promoters; AT, alternate
terminators; ES, exon skipping; ME, mutually exclusive exons; RI,
retained introns. |
 | Table IPrognostic signatures based on each
type of splicing event. |
Table I
Prognostic signatures based on each
type of splicing event.
| Type of splicing
event | Algorithm | HR (95% CI) | ROC curve |
|---|
| Alternate acceptor
sites (AA) | 'SWI5-87732-AA' *
0.03740 + 'FDPS-8074-AA' * 0.02940 + 'COMT-61101-AA' * 0.31109 | 2.334
(1.644-3.314) | 0.743 |
| Alternate donor
sites (AD) | 'VPS28-85610-AD' *
1.70e-01 + 'UBE2D3-70148-AD' * (−1.57e-02) + 'TXNDC17-38768-AD' *
9.27e-02 + 'TPT1-25803-AD' * 1.62e+01 + 'SUMF2-79789-AD' * 3.92e-02
+ 'SMIM19-83740-AD' * 2.69e-02 + 'RPL26-39180-AD' * 6.06e-01 +
'NDUFB11-88904-AD' * 4.87e-02 + 'IRF3-50997-AD' * (−8.62e-03) +
'F10-26354-AD' * 2.21e-02 + 'CCDC90B-18085-AD' * 2.02e-02 +
'C1orf159-13-AD' * (−1.12e-02) + 'ADCK4-49949-AD' (-4.56e-02) | 2.423
(1.700-3.455) | 0.718 |
| Alternate promoters
(AP) | 'RPL27-41172-AP' *
0.21696 + 'RCL1-85779-AP' * −0.00550 + 'NUDT6-70523-AP' * 0.01129 +
'MXI1-13080-AP' * 0.00857 + 'DUSP22-75126-AP' * 0.07876 +
'CYB5A-45803-AP' * 0.49517 + 'CHPF-57719-AP' * 0.07805 | 2.687
(1.891-3.818) | 0.728 |
| Alternate
terminators (AT) | 'ZSCAN20-1679-AT' *
0.01077 + 'ZNF706-84737-AT' * 0.06539 +'WWOX-37672-AT' * −0.03181 +
'NUCB2-14523-AT' * −0.07215 +'NEIL3-71227-AT' * 0.01306 +
'MORN1-252-AT' * 0.00837 + 'FRMD4A-10810-AT' * 4.59555 +
'CRISPLD2-37867-AT' * 0.24507 + 'ACAT1-18599-AT' * 0.69847 | 2.850
(2.001-4.057) | 0.709 |
| Exon skips
(ES) | 'USF2-49098-ES' *
0.03030 + 'TRAPPC6A-50409-ES' −0.03711 + 'TMEM150A-54305-ES'
−0.01822 + 'TMEM120B-24894-ES' * 0.00897 + 'TCF3-46542-ES' *
−0.02326 + 'SSR4-90499-ES' * 0.95791 + 'SMIM7-48191-ES' * 0.26393 +
'SHBG-39015-ES' * −0.02339 'PTRH1-87649-ES' * −0.64609 +
'PPP6R2-62825-ES' * −0.03581 + 'PPP2CB-83304-ES' * −3.17592 +
'PGAP3-40670-ES' * 0.06043 + 'PCYT2-44234-ES' * 0.12079 +
'OCEL1-48244-ES' * −0.04796 + 'SOC2-52105-ES' * 0.01667 +
'IRF3-50995-ES' * −0.03249 + 'IRF3-50990-ES' * 0.04115 +
'FKBP8-48448-ES' * −0.71033 + 'ECHDC2-3037-ES' * 0.04796 +
'CYTH1-43890-ES' * 0.01764 + 'CHRD-67959-ES' * −0.01020 +
'C16orf13-32924-ES' * 0.03301 + 'C14orf2-29536-ES' * 0.38460 +
'BAX-50838-ES' * −0.35118 + 'ARHGEF10L-862-ES' * 0.01344 +
'AFMID-94694-ES' * −0.01010 + 'ACAA1-64027-ES' * −0.71363 | 4.938
(3.439-7.090) | 0.898 |
| Mutually exclusive
exons (ME) |
'SLC39A14-140283-ME' * (−0.00563) +
'H2AFY-96931-ME' * 0.02881 | 1.878
(1.320-2.672) | 0.641 |
| Retained introns
(RI) | 'RPL13-38091-RI' *
0.62076 + 'ROMO1-59223-RI` * 0.07039 + 'NUDT22-16590-RI' * 0.01621
+ 'NAA60-33527-RI' * (−0.00942) + 'MRPL52-26642-RI' * 0.01287 +
'MDK-15571-RI' * 0.09757 + 'MARVELD2-72372-RI' * (−2.62393) | 2.895
(2.043-4.102) | 0.745 |
| Table IIUnivariate and multivariate analyses
of OS in HCC patients from TCGA by Cox regression analysis. |
Table II
Univariate and multivariate analyses
of OS in HCC patients from TCGA by Cox regression analysis.
| Univariate analysis
| Multivariate
analysis
|
|---|
| Variables | HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| Age (≥60/<60
years) | 1.212
(0.854-1.720) | 0.281 | | |
| Sex
(male/female) | 0.817
(0.573-1.164) | 0.262 | | |
| Pathological stage
(III-IV/I-II) | 2.446
(1.687-3.545) | <0.001 | 1.018
(0.138-7.527) | 0.986 |
| T stage
(T3-T4/T1-T2) | 2.537
(1.783-3.609) | <0.001 | 1.757
(0.238-12.981) | 0.581 |
| Lymph node
metastasis (yes/no) | 1.999
(0.490-8.161) | 0.334 | | |
| Distant metastasis
(yes/no) | 4.033
(1.267-12.834) | 0.018 | 0.755
(0.179-3.177) | 0.701 |
| Histological grade
(G3-G4/G1-G2) | 1.119
(0.780-1.604) | 0.542 | | |
| Tumor status during
follow-up (tumor/tumor-free) | 2.366
(1.623-3.447) | <0.001 | 1.567
(0.979-2.508) | 0.061 |
| Vascular invasion
(micro+macro/none) | 1.351
(0.892-2.047) | 0.155 | | |
| Relative family
cancer history (yes/no) | 1.181
(0.818-1.704) | 0.375 | | |
| Prognostic index
(high/low) | 12.904
(7.513-22.161) | <0.001 | 16.541
(7.660-35.717) | <0.001 |
| Table IIIPrognostic index for HCC patients
based on alternative splicing events. |
Table III
Prognostic index for HCC patients
based on alternative splicing events.
| Symbol | AS ID | Splice type | Coefficient | z score | P-value |
|---|
| ZFAND6 | 32171 | AA | −8.51E-01 | −2.83 | 0.0047 |
| SWI5 | 87732 | AA | 8.52E-01 | 2.46 | 0.01392 |
| BIRC2 | 18447 | AA | −1.28E+01 | −4.66 | 3.20E-06 |
| RPS14 | 74096 | AD | −2.55E+00 | −4.38 | 1.20E-05 |
| PGRMC2 | 70577 | AD | −3.42E+01 | −4.58 | 4.70E-06 |
| CCDC90B | 18085 | AD | 8.30E-01 | 2.57 | 0.0102 |
| APOC1 | 50361 | AD | 4.11E+00 | 3.32 | 0.00089 |
| RTN4 | 53584 | AP | −4.26E-01 | −3.29 | 0.001 |
| RPL23A | 39967 | AP | −9.34E+01 | −2.57 | 0.01003 |
| RPL23A | 39965 | AP | −3.06E+00 | −3.44 | 0.00059 |
| RCL1 | 85779 | AP | 4.21E-01 | 2.92 | 0.00347 |
| NUDT6 | 70523 | AP | 2.19E-01 | 2.16 | 0.03054 |
| NUDT6 | 70521 | AP | 6.16E-01 | 3.07 | 0.00216 |
| MXI1 | 13080 | AP | 1.53E-01 | 1.49 | 0.13594 |
| MXI1 | 13079 | AP | 4.76E-01 | 2.67 | 0.00752 |
| IAH1 | 52628 | AP | 1.41E+01 | 4.98 | 6.30E-07 |
| DUSP22 | 75126 | AP | −2.38E-01 | −1.76 | 0.07903 |
| DUSP22 | 75125 | AP | 9.50E-01 | 3.53 | 0.00041 |
| CYB5A | 45803 | AP | 1.29E+00 | 3.29 | 0.00099 |
| CHPF | 57718 | AP | −1.64E+00 | −2.21 | 0.02698 |
| CARKD | 26251 | AP | −2.49E-01 | −1.71 | 0.08786 |
| IAH1 | 52629 | AP | 4.82E-01 | 3 | 0.00273 |
| ZSCAN20 | 1679 | AT | −7.88E-01 | −3.12 | 0.00182 |
| ZNF706 | 84738 | AT | −1.42E+01 | −5.76 | 8.40E-09 |
| WWOX | 37672 | AT | −5.47E+00 | −7.13 | 9.70E-13 |
| TROAP | 21551 | AT | 3.63E-01 | 1.56 | 0.11927 |
| RAPH1 | 57075 | AT | 1.48E+00 | 1.83 | 0.0673 |
| RAPH1 | 57074 | AT | 5.54E-01 | 2.8 | 0.00504 |
| NUCB2 | 14523 | AT | −9.05E+00 | −2.22 | 0.0262 |
| FRMD4A | 10810 | AT | 5.09E+00 | 5.4 | 6.50E-08 |
| CRISPLD2 | 37867 | AT | 8.61E-01 | 3.72 | 0.0002 |
| CLSPN | 1731 | AT | −8.03E-01 | −1.74 | 0.08206 |
| BCAM | 50347 | AT | −3.64E-01 | −1.6 | 0.11049 |
| ACAT1 | 18599 | AT | 2.88E+00 | 4.6 | 4.30E-06 |
| XPC | 63523 | ES | 2.96E-01 | 1.79 | 0.0731 |
| WIPI2 | 78656 | ES | −3.47E-01 | −1.98 | 0.04758 |
| TRAPPC6A | 50409 | ES | −3.27E+00 | −4.69 | 2.70E-06 |
| TPRA1 | 66613 | ES | −4.71E+00 | −1.84 | 0.06575 |
| TMEM150A | 54305 | ES | 1.66E+00 | 2.85 | 0.00443 |
| TMEM141 | 88206 | ES | −3.62E-01 | −3.7 | 0.00022 |
| TCF3 | 46542 | ES | −1.57E+00 | −4.66 | 3.10E-06 |
| STRA13 | 44266 | ES | 2.97E+00 | 4.89 | 1.00E-06 |
| SSR4 | 90499 | ES | 2.39E+02 | 6.3 | 3.00E-10 |
| SLC7A9 | 48907 | ES | 3.61E-01 | 3.17 | 0.0015 |
| SLC25A45 | 16828 | ES | 3.71E-01 | 2.05 | 0.04004 |
| SLAIN2 | 69214 | ES | −6.78E-01 | −2.89 | 0.00385 |
| SHBG | 39015 | ES | −8.49E-01 | −7.07 | 1.50E-12 |
| PTRH1 | 87649 | ES | −6.58E+01 | −5.47 | 4.40E-08 |
| PPP2CB | 83304 | ES | −5.52E+02 | −5.72 | 1.10E-08 |
| PGAP3 | 40670 | ES | 2.10E+01 | 3.32 | 0.00089 |
| OCEL1 | 48244 | ES | −5.74E+00 | −2.97 | 0.00298 |
| MTA1 | 29647 | ES | −3.93E-01 | −2.25 | 0.02464 |
| MPPE1 | 44648 | ES | −3.09E-01 | −2.34 | 0.01921 |
| MPND | 46796 | ES | −6.87E+00 | −2.12 | 0.034 |
| LTBP3 | 16865 | ES | −4.93E-01 | −1.87 | 0.06167 |
| JOSD2 | 51205 | ES | −5.21E+00 | −2.51 | 0.0119 |
| ISOC2 | 52106 | ES | −5.98E+00 | −3.29 | 0.00099 |
| IRF3 | 50991 | ES | −7.04E+01 | −2.94 | 0.00327 |
| IRF3 | 50990 | ES | 5.08E-01 | 2.41 | 0.01599 |
| GUK1 | 10185 | ES | −2.01E+01 | −3.77 | 0.00016 |
| ECHDC2 | 3037 | ES | 6.27E-01 | 3.96 | 7.50E-05 |
| DGKZ | 15546 | ES | −8.54E-01 | −2.5 | 0.01235 |
| CSAD | 21952 | ES | 8.72E-01 | 3.91 | 9.30E-05 |
| CLASRP | 50393 | ES | −1.15E+00 | −2.99 | 0.00281 |
| CHRD | 67959 | ES | −5.44E-01 | −3.86 | 0.00011 |
| C17orf49 | 38825 | ES | 2.01E+01 | 3.52 | 0.00043 |
| C16orf13 | 32921 | ES | 2.25E+00 | 2.39 | 0.01699 |
| C16orf13 | 32919 | ES | −3.45E+00 | −3.98 | 6.80E-05 |
| BAX | 50838 | ES | −4.99E+01 | −3.86 | 0.00011 |
| ARHGEF10L | 862 | ES | 4.31E-01 | 1.83 | 0.06684 |
| APOC1 | 99361 | ES | −2.72E-01 | −3.56 | 0.00037 |
| AFMID | 94694 | ES | 2.79E-01 | 1.98 | 0.04811 |
| AFMID | 94690 | ES | 4.05E-01 | 2.62 | 0.00888 |
| AFMID | 43800 | ES | −1.21E+00 | −4.21 | 2.50E-05 |
| AFMID | 43798 | ES | 7.48E-01 | 3.04 | 0.00236 |
| AFMID | 43795 | ES | −5.04E-01 | −4.01 | 6.00E-05 |
| ACY1 | 65150 | ES | 3.56E+01 | 1.76 | 0.07779 |
| SLC39A14 | 140283 | ME | 3.82E-01 | 2.9 | 0.00372 |
| H2AFY | 96931 | ME | 2.26E+00 | 3.09 | 0.00198 |
| CYP4F3 | 48101 | ME | −3.19E-01 | −2.94 | 0.00327 |
| VPS28 | 85600 | RI | −7.11E-01 | −2.25 | 0.02443 |
| ROMO1 | 59223 | RI | 7.85E-01 | 2.64 | 0.00822 |
| NUDT22 | 16590 | RI | 2.71E+00 | 5.83 | 5.70E-09 |
| MRPL52 | 26642 | RI | 1.11E+00 | 4.03 | 5.60E-05 |
Survival-associated splicing factor
genes
A total of 71 splicing factor genes were identified,
and the associations between the expression profiles of these genes
and OS were assessed based on the data obtained from TCGA,
resulting in the identification of 32 prognosis-related splicing
factor genes. Interestingly, overexpression of all these 32 genes
may represent a risk factor for the survival of patients with HCC,
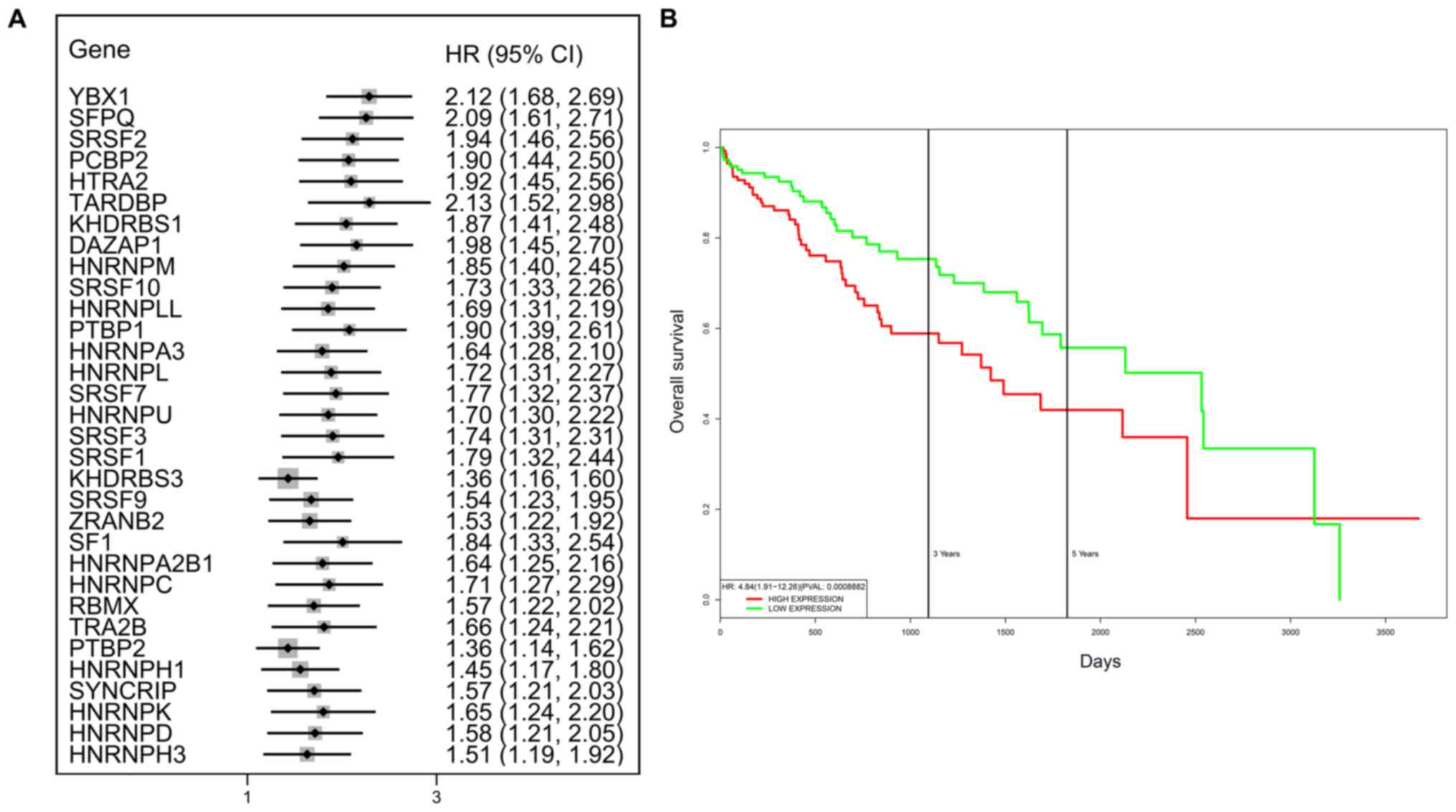
as all HRs were >1 (P<0.05) (Fig. 7A). We found that the integration of
the 32 SFs could also provide an effective survival risk
stratification for HCC patients (Fig.
7B). HCC patients with high-risk scores for the splicing
factors-based classifier had worse OS compared with those who had
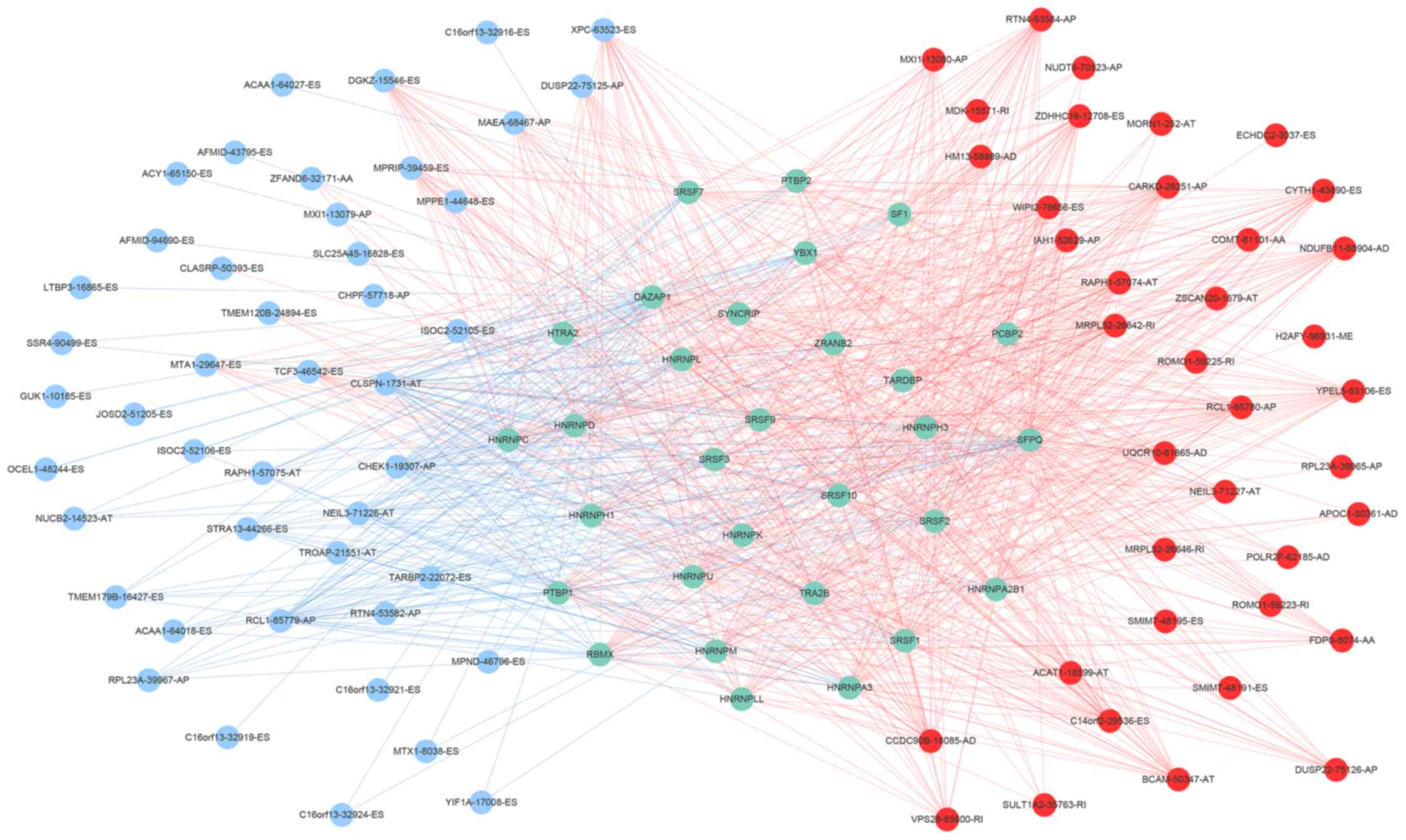
low-risk scores. Next, we characterized the splicing-regulatory
network of the 152 (P<0.0001) most significant
survival-associated alternative splicing events and 32
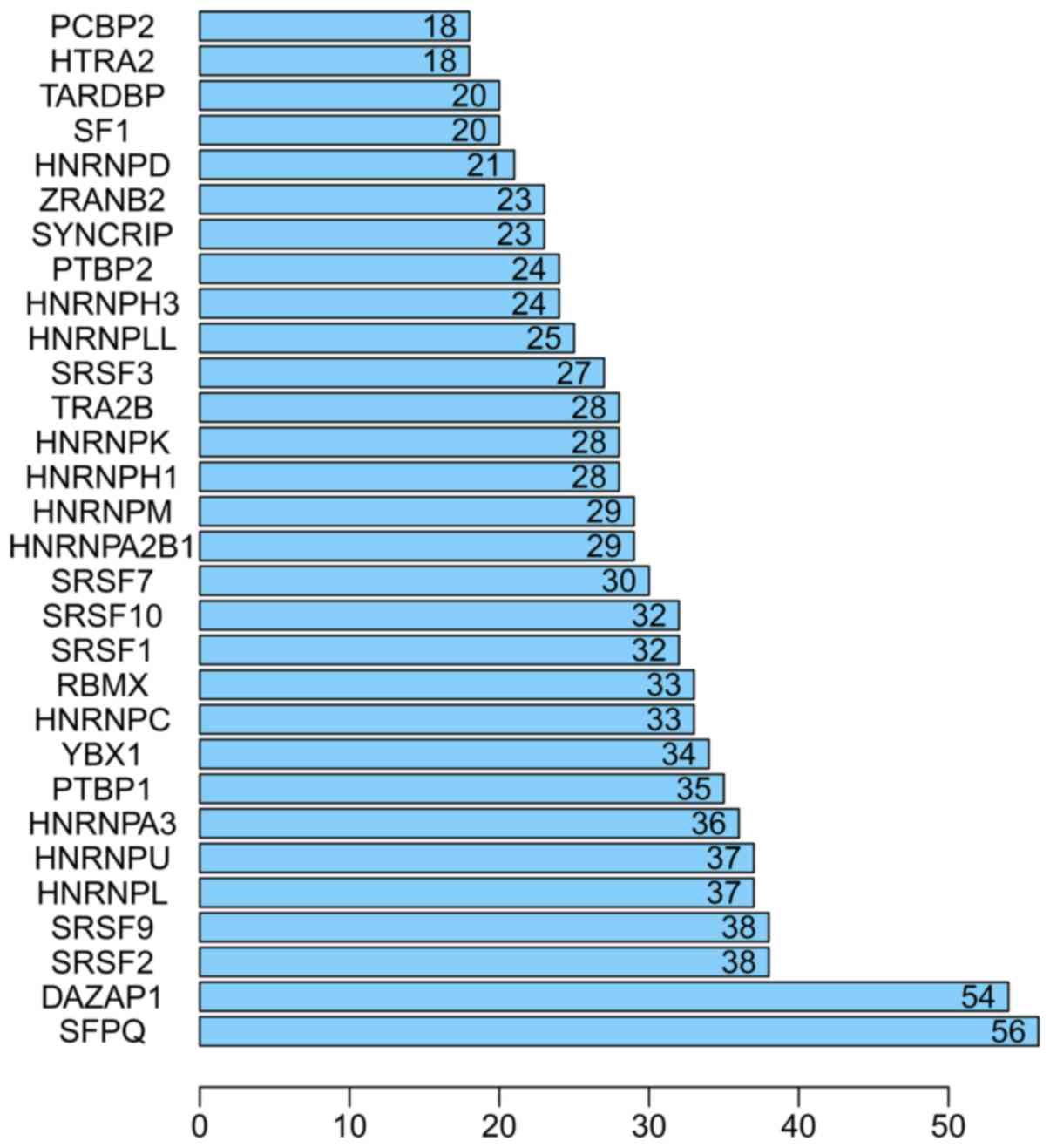
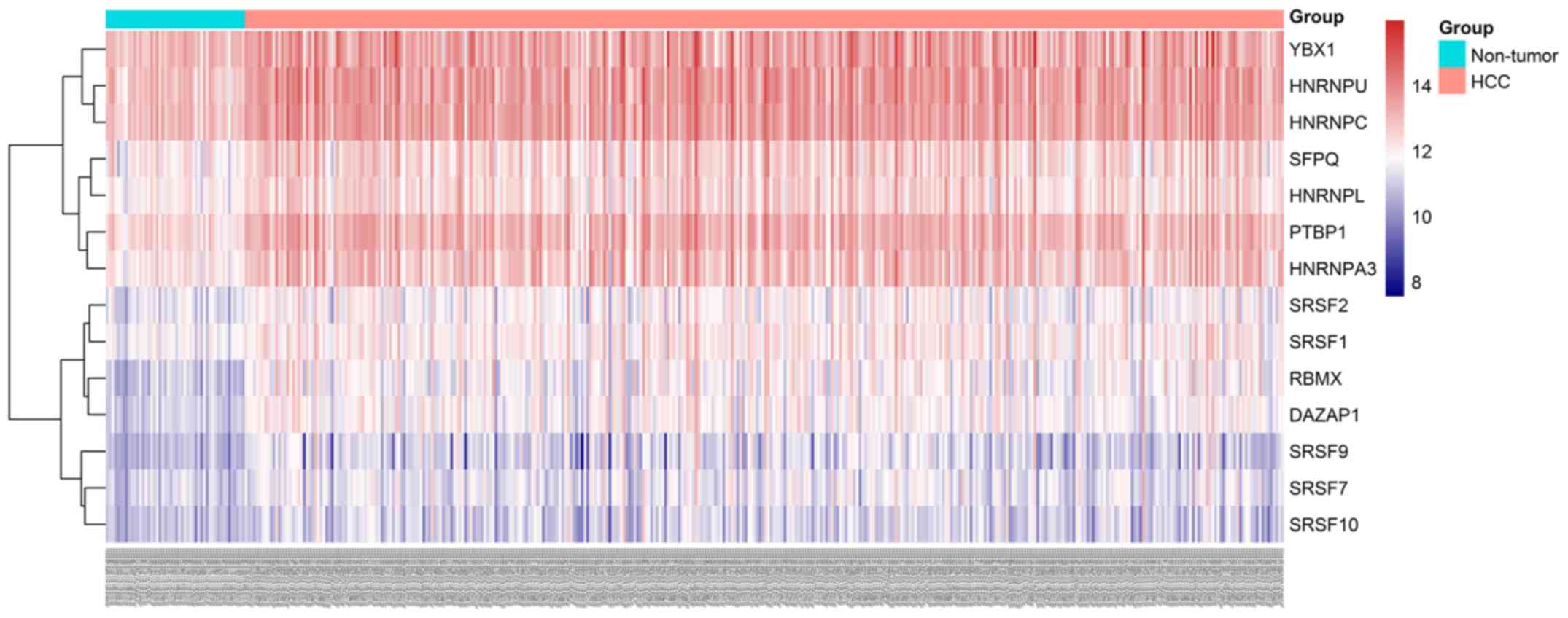
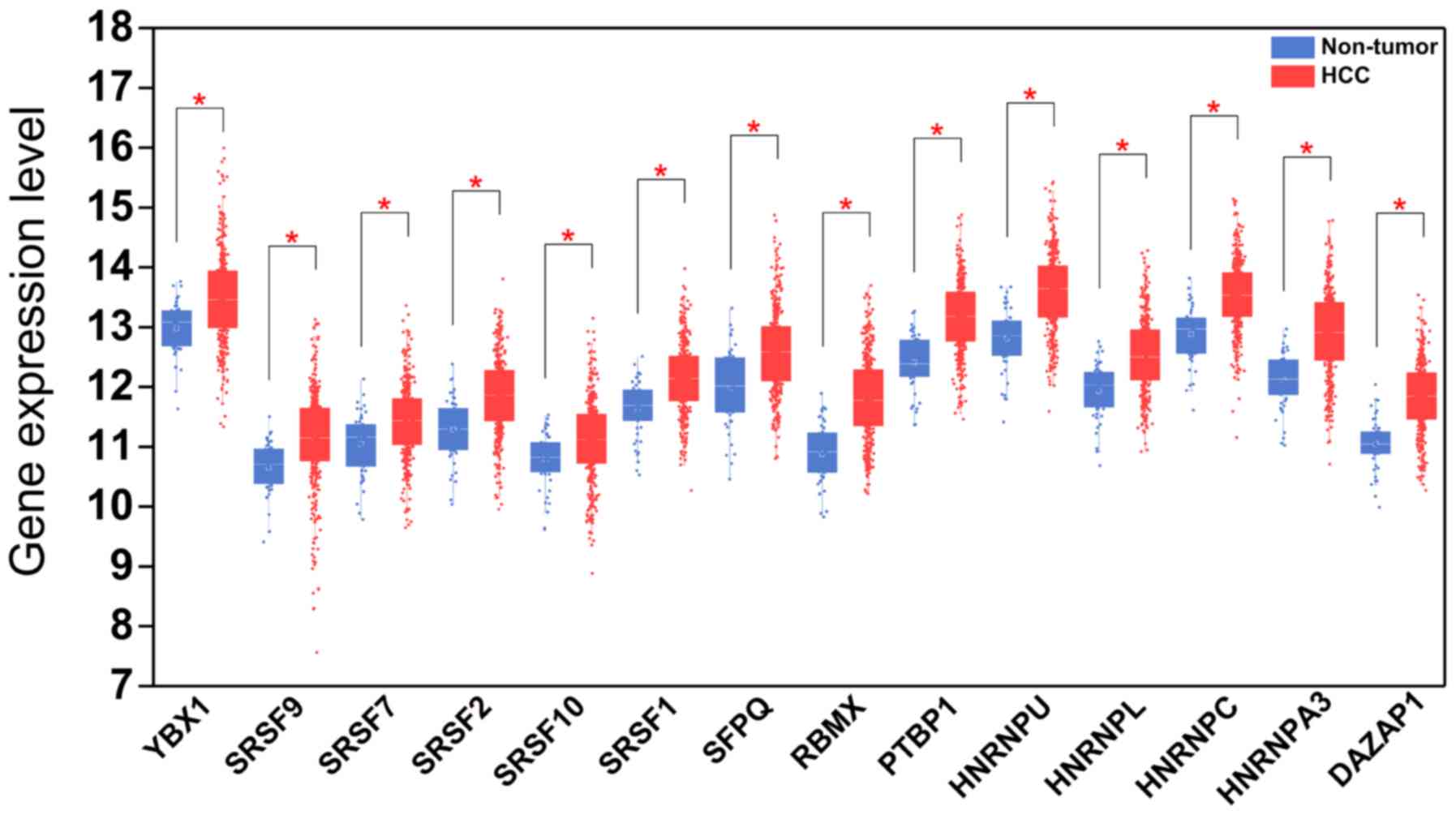
prognosis-related splicing factor genes in HCC (Fig. 8). Additionally, 14 core interacting
genes were identified in the splicing-regulated network:
SRSF7, SRSF10, SRSF1, RBMX,
HNRNPC, YBX1, PTBP1, HNRNPA3,
HNRNPU, HNRNPL, SRSF9, SRSF2,
DAZAP1 and SFPQ (Fig.
9). Furthermore, these genes were found to not only possess
prognostic significance, but to also have indispensable functions
in the initiation of HCC (Figs.
10 and 11).
Discussion
In the present study, the PSI value and clinical
outcome of patients with HCC were integrated using a novel
methodology. Furthermore, prognostic signatures were developed
based on splicing events to promote the development of precision
treatment. To determine the underlying mechanism, we investigated
the potential regulatory associations between prognosis-related
splicing factors and splicing events, which provided deeper
insights into the phenotype of splicing events in HCC.
With the advantage of high-throughput RNA-Seq, TCGA
provides multiple sources for the investigation of whole-genome or
transcriptome analyses, including the exploration of genome
splicing events. Previously, Xue et al (47) and Zhu et al (48) conducted SpliceSeq analyses using
TCGA (49) to generate alterative
splicing profiles for non-small-cell lung cancer and ovarian cancer
patients, in order to develop prognostic models with several
splicing events. SpliceSeq is a Java program providing a clear view
of the inclusion level of each exon and splice junction. Recently,
Ryan et al (45) extended
the methodology of SpliceSeq and calculated the PSI for each
potential splicing event across 33 types of cancer to establish
TCGA SpliceSeq database; the PSI data for HCC generated in the
present study were obtained from this database. Hence, to the best
of our knowledge, this is the first group to integrate PSI values
from SpliceSeq with TCGA clinical prognostic parameters for the
comprehensive investigation of the prognostic value of alternative
splicing events in HCC. This algorithm will facilitate the
discovery of novel splicing events that may be used for the
prognosis of HCC and provide insights into the role of splicing
events at the genome-wide level.
On the basis of the analysis of the tumor tissue
data of 371 HCC cases in TCGA SpliceSeq, a large number of
alternative splicing events were identified. These events do not
only affect protein diversity by altering the amino acid sequences
of the translated products (e.g., ES can delete a portion of the
coding sequence), but they can also affect protein function by
altering the yield of the translated products (e.g., AP can change
the efficiency of translation). For example, the functions of the
common clinical tumor suppressor genes TP53, ARID1A,
PTEN and PI3K, and the proto-oncogenes MET and
NOTCH1, can be changed by alternative splicing events
(21). The common
apoptosis-related genes Bcl-X and MLC1 can also
express different products due to the occurrence of alternative
splicing, showing two distinct functions of promoting apoptosis and
inhibiting apoptosis (3,4,50,51).
In the currently popular molecular targeting therapy for tumors,
the target sequence can be lost from cancer cells due to
alternative splicing, leading to drug resistance (7,13,21).
In the present study, ES was the most common of the 7 alternative
splicing events in HCC. Previous studies have demonstrated that the
frequency of alternative splicing events is not the same in
different cancers, but the frequency of ES is generally the highest
(52). Although the mechanism has
not yet been fully elucidated, evidence has shown that the
occurrence of splicing events is not random. Chen et al
(53) analyzed RNA-Seq data from
30 types of tumors and found that the frequency of alternative
splicing events is markedly higher in tumors compared with that in
corresponding normal tissues. These alternative splicing events are
often accompanied by premature termination codons, and the
probability of a premature termination codon due to alternative
splicing is markedly higher in a tumor suppressor gene rather than
in a proto-oncogene in cancer tissue. In the gene functional
enrichment analysis, it was observed that genes with
survival-associated splicing events were significant correlated
with 'small molecule catabolic processes', 'organic acid processes'
and 'viral gene expression'. The gene functional enrichment
analysis results revealed that these genes may be involved in the
metabolism of HCC. The liver is the metabolic center of the body.
Hence, the onset and progression of HCC is also inseparable from
metabolic abnormalities. Previously, Tremblay et al reported
unique alternative splicing patterns in HBV-associated HCC,
HCV-associated HCC, HBV&HCV-associated HCC, and virus-free HCC
based on TCGA dataset. They also found the signatures of genes for
which AS is dysregulated in different types of HCC (44). These findings also validated that
the process of splicing is closely associated with viral infection
status.
Accordingly, alternative splicing events may be
driving the occurrence and progression of cancer. Therefore, the
Cox regression method was used to analyze the association between
alternative splicing events and the survival of HCC patients. The
predictive power of alternative splicing on the prognosis of HCC
was analyzed by considering the ROC curve. The results demonstrated
that several alternative splicing events were associated with the
survival of HCC patients. Thus, we attempted for the first time to
use multiple alternative splicing events for the prognosis of HCC
patients and obtained satisfactory results. To date, a number of
studies have demonstrated that an alternative splicing event alone
may be used as a biomarker for tumor diagnosis and prognosis.
However, due to the heterogeneity of tumors, using a single
molecule as a marker is often insufficient. Therefore, the
combination of multiple molecular events for the prognosis of HCC
may improve the sensitivity of diagnosis and prognosis. This
follows a trend in current clinical research. Similarly, previous
studies have reported several prognostic signatures based on
different types of molecular events. For example, Qiao et al
proposed an eight-gene signature for HCC patients' survival
prediction (54). However, the AUC
of their model only reached 0.77. Liao et al also developed
an effective prognostic index (PI) for predicting the survival of
HCC patients based on microRNAs (55). However, the AUC also only reached
0.687. Hence, our PI exhibits better discriminatory power. In the
present study, the discriminatory power and model calibration were
estimated to comprehensively evaluate the model accuracy. More
importantly, the prognostic signature based on splicing events may
provide novel insight into the monitoring of HCC patients from
alternative splicing events. The splicing events included in the PI
may provide promising therapeutic targets for HCC patients.
As mentioned above, alternative splicing events do
not occur randomly, but they are rather regulated by multiple
cellular factors, including splicing factors (56,57).
Splicing factors can activate or inhibit the occurrence of splicing
events by identifying regulatory sequences in exons or introns. DNA
mutations, DNA methylation and aberrant histone modifications may
all affect the recognition of splicing sites by splicing factors,
leading to changes in the occurrence of splicing events (10,58-67).
The roles of common anomalies, such as DNA mutations, in cancer can
be rationally explained by selectivity. Mutations within introns
and synonymous mutations in exons, which were previously believed
not to affect the function of genes, have gradually been found to
affect gene function by affecting splicing events (10,58,60).
Splicing factors play a crucial regulatory role in the occurrence
of splicing events, and mutations in their sequences or changes in
their expression levels may affect splicing events (21). Changes in the expression levels of
splicing factors are usually accompanied by changes in splicing
events, but the pattern is not yet clear. Both the upregulation and
downregulation of their expression can cause cancer or inhibit
tumor growth by altering splicing events (25). Analysis of the association between
splicing factors and survival in patients with HCC revealed that
the expression levels of 32 splicing factors were correlated with
survival time, with high expression being associated with shorter
survival. It was hypothesized that the expression levels of these
factors may be regulated by the similar events. For example, such
an effect of the MYC oncogene has been reported (22). To elucidate the mechanism
underlying the effect of splicing factors on the survival of
patients with HCC, we analyzed the correlations between the
expression levels of these 32 splicing factors and survival-related
alternative splicing events. Surprisingly, the alternative splicing
events that were positively correlated with survival were also
positively correlated with the expression levels of multiple
splicing factors, and most alternative splicing events that were
negatively correlated with survival were also negatively correlated
with the expression levels of the splicing factors. This finding
indicates that the increase in the expression of multiple splicing
factors can affect the survival of patients with HCC by
synergistically regulating the alternative splicing events of
genes.
However, there were several limitations to the
present study. First, this study was based on an individual source
of data from TCGA, without validation from independent cohorts,
which will be carried out by our group in the future with clinical
samples. Furthermore, the biological roles of the splicing events
require further validation. In addition, the present study mainly
focused on the splicing status of mRNAs. In fact, the splicing
events of other types of RNA, including circular RNAs, also play
important roles in the processes of tumors (68). Further research and effective
arithmetic are required to broaden its application range (69). Finally, classic splicing factors
were utilized in the present study, whereas it is possible that
other RNA-binding proteins may also affect splicing events. Hence,
the association between splicing events and a complete repertoire
of RNA-binding proteins from established sources in the human
genome should be further investigated.
Our study preliminarily demonstrated that
alternative splicing events and the expression levels of splicing
factors play important roles in the progression of HCC. On the
basis of these results, it may be considered that splicing factors
affect the occurrence and progression of HCC by regulating
alternative splicing events. Some special alternative splicing
events may affect the prognosis and progression of HCC by being
regulated by their corresponding splicing factors. The constructed
computational prognostic model based on HCC-specific alternative
splicing events may be used as a molecular marker for the prognosis
of HCC.
Currently, certain anticancer drugs targeting
splicing factors, such as E7107 and FR901464 (29,70),
have been developed, but they are not widely used in clinical
practice due to their prominent side effects (21). The identified HCC-specific
alternative splicing events may be used as molecular markers for
the diagnosis and prognosis of HCC, and they can help physicians
develop more precise targeted therapies. This approach has the
potential to be widely applied in the field of HCC research.
Acknowledgments
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 31760319), the
Natural Science Foundation of Guangxi, China, the Guangxi Degree
and Postgraduate Education Reform and Development Research
Projects, China (grant no. JGY2019xxx), the Promoting Project of
Basic Capacity for Young and Middle-aged University Teachers in
Guangxi (grant no. 2017KY0111), the Innovation Project of Guangxi
Graduate Education (grant no. YCBZ2017045), the Guangxi Medical
University Training Program for Distinguished Young Scholars, the
Medical Excellence Award Funded by the Creative Research
Development Grant from the First Affiliated Hospital of Guangxi
Medical University, and the Guangxi Zhuang Autonomous Region Health
and Family Planning Commission Self-financed Scientific Research
Project (grant no. Z20180979).
Availability of data and materials
The datasets used and analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
HW, SP and GC participated in the conception and
design of the study. ZP, RH, BL, JM, XH and YD performed the
statistical analysis and were involved in the preparation of the
figures and tables. HW, ZP and RH reviewed the results and
participated in the discussion of the data. HW, ZP, SP and GC
prepared and revised the manuscript. All the authors have read and
approved the final version of this manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Breitbart RE, Andreadis A and Nadal-Ginard
B: Alternative splicing: A ubiquitous mechanism for the generation
of multiple protein isoforms from single genes. Annu Rev Biochem.
56:467–495. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Blencowe BJ: Alternative splicing: New
insights from global analyses. Cell. 126:37–47. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Takehara T, Liu X, Fujimoto J, Friedman SL
and Takahashi H: Expression and role of Bcl-xL in human
hepatocellular carcinomas. Hepatology. 34:55–61. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bingle CD, Craig RW, Swales BM, Singleton
V, Zhou P and Whyte MK: Exon skipping in Mcl-1 results in a bcl-2
homology domain 3 only gene product that promotes cell death. J
Biol Chem. 275:22136–22146. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Todaro M, Gaggianesi M, Catalano V,
Benfante A, Iovino F, Biffoni M, Apuzzo T, Sperduti I, Volpe S,
Cocorullo G, et al: CD44v6 is a marker of constitutive and
reprogrammed cancer stem cells driving colon cancer metastasis.
Cell Stem Cell. 14:342–356. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zheng S, Kim H and Verhaak RGW: Silent
mutations make some noise. Cell. 156:1129–1131. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Salton M, Kasprzak WK, Voss T, Shapiro BA,
Poulikakos PI and Misteli T: Inhibition of vemurafenib-resistant
melanoma by interference with pre-mRNA splicing. Nat Commun.
6:71032015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bechara EG, Sebestyén E, Bernardis I,
Eyras E and Valcárcel J: RBM5, 6, and 10 differentially regulate
NUMB alternative splicing to control cancer cell proliferation. Mol
Cell. 52:720–733. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ghigna C, Giordano S, Shen H, Benvenuto F,
Castiglioni F, Comoglio PM, Green MR, Riva S and Biamonti G: Cell
motility is controlled by SF2/ASF through alternative splicing of
the Ron protooncogene. Mol Cell. 20:881–890. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jung H, Lee D, Lee J, Park D, Kim YJ, Park
WY, Hong D, Park PJ and Lee E: Intron retention is a widespread
mechanism of tumor-suppressor inactivation. Nat Genet.
47:1242–1248. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Goldstein LD, Lee J, Gnad F, Klijn C,
Schaub A, Reeder J, Daemen A, Bakalarski CE, Holcomb T, Shames DS,
et al: Recurrent loss of NFE2L2 exon 2 is a mechanism for Nrf2
pathway activation in human cancers. Cell Rep. 16:2605–2617. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kornblihtt AR: Epigenetics at the base of
alternative splicing changes that promote colorectal cancer. J Clin
Invest. 127:3281–3283. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Poulikakos PI, Persaud Y, Janakiraman M,
Kong X, Ng C, Moriceau G, Shi H, Atefi M, Titz B, Gabay MT, et al:
RAF inhibitor resistance is mediated by dimerization of aberrantly
spliced BRAF(V600E). Nature. 480:387–390. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sotillo E, Barrett DM, Black KL, Bagashev
A, Oldridge D, Wu G, Sussman R, Lanauze C, Ruella M, Gazzara MR, et
al: Convergence of acquired mutations and alternative splicing of
CD19 enables resistance to CART-19 immunotherapy. Cancer Discov.
5:1282–1295. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang BD, Ceniccola K, Hwang S, Andrawis R,
Horvath A, Freedman JA, Olender J, Knapp S, Ching T, Garmire L, et
al: Alternative splicing promotes tumour aggressiveness and drug
resistance in African American prostate cancer. Nat Commun.
8:159212017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sebestyén E, Zawisza M and Eyras E:
Detection of recurrent alternative splicing switches in tumor
samples reveals novel signatures of cancer. Nucleic Acids Res.
43:1345–1356. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Trincado JL, Sebestyén E, Pagés A and
Eyras E: The prognostic potential of alternative transcript
isoforms across human tumors. Genome Med. 8:852016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shen S, Wang Y, Wang C, Wu YN and Xing Y:
SURVIV for survival analysis of mRNA isoform variation. Nat Commun.
7:115482016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Parsons DW, Li M, Zhang X, Jones S, Leary
RJ, Lin JC, Boca SM, Carter H, Samayoa J, Bettegowda C, et al: The
genetic landscape of the childhood cancer medulloblastoma. Science.
331:435–439. 2011. View Article : Google Scholar :
|
|
20
|
Ghigna C, De Toledo M, Bonomi S, Valacca
C, Gallo S, Apicella M, Eperon I, Tazi J and Biamonti G:
Pro-metastatic splicing of Ron proto-oncogene mRNA can be reversed:
Therapeutic potential of bifunctional oligonucleotides and indole
derivatives. RNA Biol. 7:495–503. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee SC and Abdel-Wahab O: Therapeutic
targeting of splicing in cancer. Nat Med. 22:976–986. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Koh CM, Bezzi M, Low DH, Ang WX, Teo SX,
Gay FP, Al-Haddawi M, Tan SY, Osato M, Sabò A, et al: MYC regulates
the core pre-mRNA splicing machinery as an essential step in
lymphomagenesis. Nature. 523:96–100. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Havens MA and Hastings ML:
Splice-switching antisense oligonucleotides as therapeutic drugs.
Nucleic Acids Res. 44:6549–6563. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Salton M and Misteli T: Small molecule
modulators of Pre-mRNA splicing in cancer therapy. Trends Mol Med.
22:28–37. 2016. View Article : Google Scholar :
|
|
25
|
Dvinge H, Kim E, Abdel-Wahab O and Bradley
RK: RNA splicing factors as oncoproteins and tumour suppressors.
Nat Rev Cancer. 16:413–430. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bonnal S, Vigevani L and Valcárcel J: The
spliceosome as a target of novel antitumour drugs. Nat Rev Drug
Discov. 11:847–859. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Webb TR, Joyner AS and Potter PM: The
development and application of small molecule modulators of SF3b as
therapeutic agents for cancer. Drug Discov Today. 18:43–49. 2013.
View Article : Google Scholar :
|
|
28
|
Yokoi A, Kotake Y, Takahashi K, Kadowaki
T, Matsumoto Y, Minoshima Y, Sugi NH, Sagane K, Hamaguchi M, Iwata
M, et al: Biological validation that SF3b is a target of the
antitumor macrolide pladienolide. FEBS J. 278:4870–4880. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kotake Y, Sagane K, Owa T, Mimori-Kiyosue
Y, Shimizu H, Uesugi M, Ishihama Y, Iwata M and Mizui Y: Splicing
factor SF3b as a target of the antitumor natural product
pladienolide. Nat Chem Biol. 3:570–575. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fan L, Lagisetti C, Edwards CC, Webb TR
and Potter PM: Sudemycins, novel small molecule analogues of
FR901464, induce alternative gene splicing. ACS Chem Biol.
6:582–589. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xargay-Torrent S, López-Guerra M, Rosich
L, Montraveta A, Roldán J, Rodríguez V, Villamor N, Aymerich M,
Lagisetti C, Webb TR, et al: The splicing modulator sudemycin
induces a specific antitumor response and cooperates with ibrutinib
in chronic lymphocytic leukemia. Oncotarget. 6:22734–22749. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wan L, Yu W, Shen E, Sun W, Liu Y, Kong J,
Wu Y, Han F, Zhang L, Yu T, et al: SRSF6-regulated alternative
splicing that promotes tumour progression offers a therapy target
for colorectal cancer. Gut. 68:118–129. 2019. View Article : Google Scholar
|
|
33
|
Dvinge H and Bradley RK: Widespread intron
retention diversifies most cancer transcriptomes. Genome Med.
7:452015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hashemikhabir S, Budak G and Janga SC:
ExSurv: A web resource for prognostic analyses of exons across
human cancers using clinical transcriptomes. Cancer Inform.
15(Suppl 2): 17–24. 2016.PubMed/NCBI
|
|
35
|
Neelamraju Y, Gonzalez-Perez A,
Bhat-Nakshatri P, Nakshatri H and Janga SC: Mutational landscape of
RNA-binding proteins in human cancers. RNA Biol. 15:115–129. 2018.
View Article : Google Scholar :
|
|
36
|
Wang ZL, Li B, Luo YX, Lin Q, Liu SR,
Zhang XQ, Zhou H, Yang JH and Qu LH: Comprehensive Genomic
Characterization of RNA-Binding Proteins across Human Cancers. Cell
Rep. 22:286–298. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kechavarzi B and Janga SC: Dissecting the
expression landscape of RNA-binding proteins in human cancers.
Genome Biol. 15:R142014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Luo C, Cheng Y, Liu Y, Chen L, Liu L, Wei
N, Xie Z, Wu W and Feng Y: SRSF2 regulates alternative splicing to
drive hepatocellular carcinoma development. Cancer Res.
77:1168–1178. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li X, Qian X, Peng LX, Jiang Y, Hawke DH,
Zheng Y, Xia Y, Lee JH, Cote G, Wang H, et al: Corrigendum: A
splicing switch from ketohexokinase-C to ketohexokinase-A drives
hepatocellular carcinoma formation. Nat Cell Biol. 18:7092016.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chiu YT, Wong JK, Choi SW, Sze KM, Ho DW,
Chan LK, Lee JM, Man K, Cherny S, Yang WL, et al: Novel pre-mRNA
splicing of intronically integrated HBV generates oncogenic chimera
in hepatocellular carcinoma. J Hepatol. 64:1256–1264. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Luo ZL, Cheng SQ, Shi J, Zhang HL, Zhang
CZ, Chen HY, Qiu BJ, Tang L, Hu CL, Wang HY, et al: A splicing
variant of Merlin promotes metastasis in hepatocellular carcinoma.
Nat Commun. 6:84572015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bayliss J, Lim L, Thompson AJ, Desmond P,
Angus P, Locarnini S and Revill PA: Hepatitis B virus splicing is
enhanced prior to development of hepatocellular carcinoma. J
Hepatol. 59:1022–1028. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yuan JH, Liu XN, Wang TT, Pan W, Tao QF,
Zhou WP, Wang F and Sun SH: The MBNL3 splicing factor promotes
hepatocellular carcinoma by increasing PXN expression through the
alternative splicing of lncRNA-PXN-AS1. Nat Cell Biol. 19:820–832.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tremblay MP, Armero VE, Allaire A,
Boudreault S, Martenon-Brodeur C, Durand M, Lapointe E, Thibault P,
Tremblay-Létourneau M, Perreault JP, et al: Global profiling of
alternative RNA splicing events provides insights into molecular
differences between various types of hepatocellular carcinoma. BMC
Genomics. 17:6832016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ryan M, Wong WC, Brown R, Akbani R, Su X,
Broom B, Melott J and Weinstein J: TCGASpliceSeq a compendium of
alternative mRNA splicing in cancer. Nucleic Acids Res.
44:D1018–D1022. 2016. View Article : Google Scholar :
|
|
46
|
Goswami CP and Nakshatri H: PROGgeneV2:
Enhancements on the existing database. BMC Cancer. 14:9702014.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xue L, Xie L and Song X and Song X:
Identification of potential tumor-educated platelets RNA biomarkers
in non-small-cell lung cancer by integrated bioinformatical
analysis. J Clin Lab Anal. 32:e224502018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhu J, Chen Z and Yong L: Systematic
profiling of alternative splicing signature reveals prognostic
predictor for ovarian cancer. Gynecol Oncol. 148:368–374. 2018.
View Article : Google Scholar
|
|
49
|
Ryan MC, Cleland J, Kim R, Wong WC and
Weinstein JN: SpliceSeq: A resource for analysis and visualization
of RNA-Seq data on alternative splicing and its functional impacts.
Bioinformatics. 28:2385–2387. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Boise LH, González-García M, Postema CE,
Ding L, Lindsten T, Turka LA, Mao X, Nuñez G and Thompson CB:
bcl-x, a bcl-2-related gene that functions as a dominant regulator
of apoptotic cell death. Cell. 74:597–608. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Bae J, Leo CP, Hsu SY and Hsueh AJ:
MCL-1S, a splicing variant of the antiapoptotic BCL-2 family member
MCL-1, encodes a proapoptotic protein possessing only the BH3
domain. J Biol Chem. 275:25255–25261. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Li Y, Sun N, Lu Z, Sun S, Huang J, Chen Z
and He J: Prognostic alternative mRNA splicing signature in
non-small cell lung cancer. Cancer Lett. 393:40–51. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Chen L, Tovar-Corona JM and Urrutia AO:
Increased levels of noisy splicing in cancers, but not for
oncogene-derived transcripts. Hum Mol Genet. 20:4422–4429. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Qiao GJ, Chen L, Wu JC and Li ZR:
Identification of an eight-gene signature for survival prediction
for patients with hepatocellular carcinoma based on integrated
bioinformatics analysis. PeerJ. 7:e65482019. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Liao X, Zhu G, Huang R, Yang C, Wang X,
Huang K, Yu T, Han C, Su H and Peng T: Identification of potential
prognostic microRNA biomarkers for predicting survival in patients
with hepatocellular carcinoma. Cancer Manag Res. 10:787–803. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wahl MC, Will CL and Lührmann R: The
spliceosome: Design principles of a dynamic RNP machine. Cell.
136:701–718. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Turunen JJ, Niemelä EH, Verma B and
Frilander MJ: The significant other: Splicing by the minor
spliceosome. Wiley Interdiscip Rev RNA. 4:61–76. 2013. View Article : Google Scholar :
|
|
58
|
Supek F, Miñana B, Valcárcel J, Gabaldón T
and Lehner B: Synonymous mutations frequently act as driver
mutations in human cancers. Cell. 156:1324–1335. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Sterne-Weiler T and Sanford JR: Exon
identity crisis: Disease-causing mutations that disrupt the
splicing code. Genome Biol. 15:2012014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Diederichs S, Bartsch L, Berkmann JC,
Fröse K, Heitmann J, Hoppe C, Iggena D, Jazmati D, Karschnia P,
Linsenmeier M, et al: The dark matter of the cancer genome:
Aberrations in regulatory elements, untranslated regions, splice
sites, non-coding RNA and synonymous mutations. EMBO Mol Med.
8:442–457. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Singh S, Narayanan SP, Biswas K, Gupta A,
Ahuja N, Yadav S, Panday RK, Samaiya A, Sharan SK and Shukla S:
Intragenic DNA methylation and BORIS-mediated cancer-specific
splicing contribute to the Warburg effect. Proc Natl Acad Sci USA.
114:11440–11445. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Gelfman S, Cohen N, Yearim A and Ast G:
DNA-methylation effect on cotranscriptional splicing is dependent
on GC architecture of the exon-intron structure. Genome Res.
23:789–799. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Shukla S, Kavak E, Gregory M, Imashimizu
M, Shutinoski B, Kashlev M, Oberdoerffer P, Sandberg R and
Oberdoerffer S: CTCF-promoted RNA polymerase II pausing links DNA
methylation to splicing. Nature. 479:74–79. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Yuan H, Li N, Fu D, Ren J, Hui J, Peng J,
Liu Y, Qiu T, Jiang M, Pan Q, et al: Histone methyltransferase
SETD2 modulates alternative splicing to inhibit intestinal
tumorigenesis. J Clin Invest. 127:3375–3391. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Ding X, Liu S, Tian M, Zhang W, Zhu T, Li
D, Wu J, Deng H, Jia Y, Xie W, et al: Activity-induced histone
modifications govern Neurexin-1 mRNA splicing and memory
preservation. Nat Neurosci. 20:690–699. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Sharma A, Nguyen H, Geng C, Hinman MN, Luo
G and Lou H: Calcium-mediated histone modifications regulate
alternative splicing in cardiomyocytes. Proc Natl Acad Sci USA.
111:E4920–E4928. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Kim S, Kim H, Fong N, Erickson B and
Bentley DL: Pre-mRNA splicing is a determinant of histone H3K36
methylation. Proc Natl Acad Sci USA. 108:13564–13569. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zhang XO, Dong R, Zhang Y, Zhang JL, Luo
Z, Zhang J, Chen LL and Yang L: Diverse alternative back-splicing
and alternative splicing landscape of circular RNAs. Genome Res.
26:1277–1287. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Feng J, Chen K, Dong X, Xu X, Jin Y, Zhang
X, Chen W, Han Y, Shao L, Gao Y, et al: Genome-wide identification
of cancer-specific alternative splicing in circRNA. Mol Cancer.
18:352019. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Kaida D, Motoyoshi H, Tashiro E, Nojima T,
Hagiwara M, Ishigami K, Watanabe H, Kitahara T, Yoshida T, Nakajima
H, et al: Spliceostatin A targets SF3b and inhibits both splicing
and nuclear retention of pre-mRNA. Nat Chem Biol. 3:576–583. 2007.
View Article : Google Scholar : PubMed/NCBI
|