Introduction
The World Health Organization (WHO) estimates that
one third of the population worldwide is infected with
Mycobacterium tuberculosis, and that one tenth of the
infected population will develop active tuberculosis (1). Phagocytic cells, such as macrophages,
dendritic cells (DCs) and neutrophils, are among the first cells to
recognize M. tuberculosis within the host. Macrophages and
DCs recognize M. tuberculosis by pattern recognition
receptors (PRRs), such as toll-like receptors (TLRs) (2,3).
TLR2, 4 and 9 can mediate the in vitro recognition of M.
tuberculosis. In their absence (particularly of TLR2), the
cytokine response of macrophages or DCs to M. tuberculosis
will be reduced (4–6). The apoptosis of macrophages in M.
tuberculosis infection is considered to play an important role
in the pathogenesis of tuberculosis. However, the mechanism
underlying this apoptosis remains unclear.
Lipoproteins, which are post-translationally
modified proteins, are generally considered to be potent immune
modulators. Mycobacteria are rich in lipoproteins: there are ~100
open reading frames that possess the characteristic amino-terminal
acylation motif of lipoproteins in the M. tuberculosis
genome (7). The M.
tuberculosis 19-kDa lipoprotein (P19) is cell wall-associated
and immunodominant, and has been shown to stimulate the
proliferation of CD4+ T cells and the release of
interleukin 2 (IL-2), interferon-γ (IFN-γ) and IL-12 (8,9). In
addition, P19 also acts as an adhesin, interacting with the mannose
receptor, and promotes the phagocytosis of mycobacteria (10). Moreover, P19 plays a critical role
in inducing the death of macrophages through both extrinsic
(TLR2-dependent apoptotic pathway) and intrinsic (mitochondrial
pathway) mechanisms (11).
Although the mechanism of apoptosis of
mycobacteria-infected macrophages remains to be elucidated, it is
clear that the activation of inflammation and apoptosis is involved
in this process. Curcumin, a widely used dietary pigment and spice,
possesses potent antioxidant, anti-inflammatory and antitumor
activities (12,13). Synthetic curcumin analogs and
chalcone derivatives are considered as active compounds against
M. tuberculosis, but their mechanism of action is not fully
understood (14,15). Curcumin can suppress cell growth by
inhibiting various kinases, such as protein kinase C (PKC), c-Jun
NH2-terminal kinase (JNK), and the epidermal growth
factor (EGF) receptor kinase (16–18).
Curcumin can also inhibit TLR2 gene expression and protein
functions possibly via an oxidative process (19). Overall, the mechanism of the
anti-tuberculosis effect of curcumin remains unclear and needs to
be further elucidated.
In the present study, we tested the hypothesis that
curcumin exerts a protective effect against P19-induced
inflammation and apoptosis in human macrophages by inhibiting the
TLR2-induced and mitogen-activated protein kinase (MAPK)-dependent
signaling pathways. Our results demonstrated that both P19 and
curcumin induce the apoptosis of macrophages. Furthermore, our
study indicated that low doses of curcumin can inhibit the
apoptosis induced by P19 in macrophages by activating the JNK
pathway.
Materials and methods
Reagents and antibodies
Curcumin was purchased from Sigma-Aldrich (St.
Louis, MO, USA), was dissolved in dimethylsulfoxide (DMSO), and
stored at −20°C. The final concentration of DMSO in all experiments
was <0.1%. The JNK inhibitor SP600125 and the p38 inhibitor
SB203580 were purchased from Cayman Chemical Co. (Ann Arbor, MI,
USA). Cell culture reagents, medium, fetal bovine serum (FBS),
L-glutamine, and antibiotics were purchased from Thermo Fisher
Scientific (Gibco®; Waltham, MA, USA). The antibodies
targeting total extracellular signal-regulated kinase (ERK),
phospho-ERK (Thr202/Tyr204), total JNK, phospho-JNK
(Thr183/Tyr185), total p38, phospho-p38 (Thr180/Tyr182), TLR2 and
GAPDH were purchased from Cell Signaling Technology (Danvers, MA,
USA).
Bacterial strain and P19 isolation
The M. tuberculosis strain H37Rv was obtained
from the American Type Culture Collection (ATCC 5618; Manassas, VA,
USA) in lyophilized form, was reconstituted and used as previously
described (20). M.
tuberculosis 19-kDa lipoprotein (P19) was purified as
previously described (21).
Briefly, cell-wall fractions were obtained by sonication of
resuspended M. tuberculosis H37Rv in ice-cold water (20 kHz,
5 cycles, 5 min each). Forty micrograms of protein were dissolved
in a reducing sample buffer that contained 0.05 mM EDTA, 0.1%
sodium dodecyl sulfate (SDS), 1% glycerol, 10% 2-mercaptoethanol,
and 0.5 mM/ml Tris-HCl (pH 6.8), were heated for 5 min at 95°C and
were subjected to 12% SDS-polyacrylamide gel electrophoresis. Next,
proteins were transferred onto a nitrocellulose membrane and
stained with Ponceau Red to identify the 19-kDa band; this band was
also identified using a rabbit monoclonal antibody IT-19 targeting
the 19-kDA M. tuberculosis antigen (TB Research Material and
Vaccine Testing Contract, Colarado State University, Colarado,
USA). Then the band was excised, solubilized in DMSO and
precipitated with a carbonate/bicarbonate sodium buffer (0.05 M, pH
9.6). The pellet was rinsed thrice with phosphate-buffered saline
(PBS, pH 7.4) and stored at −20°C. The concentration of the protein
was determined with the Bradford method (Bio-Rad, Hercules,
USA).
Cell cultures
The WBC 264-9C macrophage line (ATCC HB-8902) was
cultivated in RPMI-1640 medium supplemented with 15% FBS, 10 mM
HEPES (2-[4-(2-hydroxyethyl) piperazin-1-yl] ethanesulfonic acid),
2 mM L-glutamine, and 50 μg/ml gentamicin, at 37°C in a humidified
incubator containing 5% CO2. For P19 treatment, WBC
264-9C cells were placed in 12-well plates with glass coverslips at
a density of 5×105 cells/ml for 24 h. Cells were then
washed and incubated for an additional 18 h in the medium
supplemented with 0.1% FBS. The macrophage monolayers in the tissue
culture plates were washed with pre-warmed RPMI-1640 medium, which
was then replaced with 1 ml of RPMI-1640 medium supplemented with
10 mM HEPES and 0.4% human serum albumin. The cells in the controls
groups were cultured in the same RPMI-1640 medium and environmental
conditions mentioned above, but were treated with 10% DMSO vehicles
later.
Cell viability assays
The effect of P19 and curcumin on the viability of
macrophages were evaluated using the MTT assay. Briefly, WBC 264-9C
macrophages (1×104 cells/well) were seeded into a
96-well culture plate. Following adherence, the cells were treated
with various concentrations of P19 (5, 10 or 20 μg/ml) or curcumin
(10, 20, 40 and 80 μM), JNK inhibitor SP600125 (30 μM) or p38
inhibitor SB203580P38 (20 μM), and incubated at 37°C for various
time periods. Then the medium was removed, and cells were incubated
with 100 μl (0.5 mg/ml) of
3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl tetrazolium bromide (MTT;
Sigma-Aldrich) for 4 h at 37°C. The formazan crystals were
solubilized with DMSO, and the absorbance was measured at 490 nm
with a microplate reader (Tecan Trading AG, Männedorf,
Switzerland).
Cytokine assays
The concentrations of IL-6 and tumor necrosis
factor-α (TNF-α) in the supernatant of the cell cultures were
determined by using Invitrogen™ enzyme-linked immunosorbent assay
(ELISA) kits (Thermo Fisher Scientific) for the corresponding human
proteins following the manufacturer’s instructions.
Western blotting
Following treatment with P19 and/or curcumin,
macrophages were rinsed with pre-warmed PBS and lysed in an
ice-cold extraction buffer (50 mM Tris pH 7.5, 150 mM NaCl, 10%
glycerol, 1 mM EDTA, 1 mM EGTA, 1% NP-40, 1 mM dithiothreitol, and
a protease inhibitor cocktail). The homogenate was incubated on ice
for 20 min and centrifuged at 13,000 × g for 20 min at 4°C. Next,
the supernatant was collected, and the concentration of the protein
in the supernatant was quantified using the Bradford method. The
whole-cell lysate from the macrophages was subjected to 12%
SDS-PAGE, and subsequently blotted onto a nitrocellulose membrane.
The membrane was then incubated with the specific antibodies, using
GAPDH as an internal control. Quantification of the protein bands
was performed by densitometry using the QuantityOne software
(Bio-Rad).
Statistical analysis
The data were expressed as mean ± SEM from three
independent experiments, and were analyzed using the SAS 8.2
software package (SAS Institute Inc., Cary, NC, USA). One-way
analysis of variance (ANOVA) followed by Bonferroni correction was
used to compare the means of three or more groups in the analyses
of cell survival, cytokine production and protein expression. The
phosphorylation levels of ERK, JNK and p38 were expressed as
phosphorylated protein/total protein. P<0.05 was considered to
indicate statistically significant differences.
Results
P19 and curcumin decrease the cell
viability of WBC 264-9C macrophages
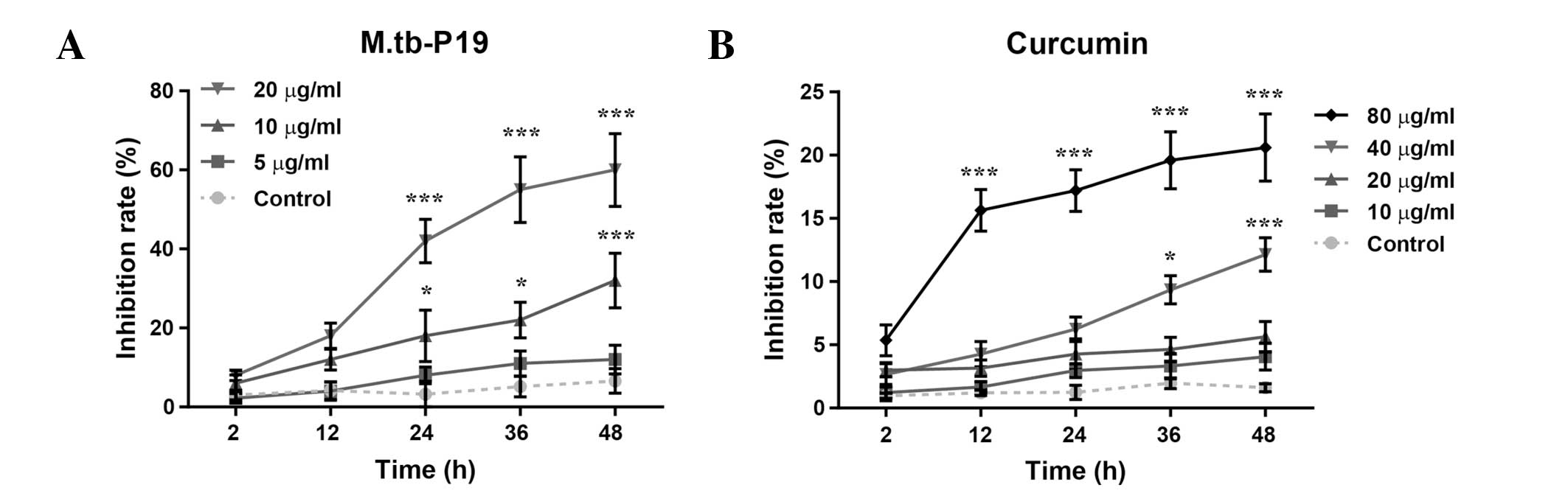
The MTT assay was used to measure the viability of
macrophages exposed to P19 or curcumin for 48 h (Fig. 1A). P19 inhibited the cell viability
of WBC 264-9C macrophages in a concentration- and time-dependent
manner (Fconcentration (3, 100) = 55.0, P<0.0001;
Ftime (4, 100) = 18.6, P<0.0001; Finteraction
(12, 100) = 4.87, P<0.0001). Post-hoc analysis revealed
that P19 significantly inhibits macrophage viability (10 μg/ml
inhibition rate, 32.4%, P<0.0001; 20 μg/ml inhibition rate,
61.5%, P<0.0001). Similarly, curcumin also inhibited macrophage
viability (Fig. 1B) in an
concentration- and time-dependent manner (Fconcentration (4,
125) = 123, P<0.0001; Ftime (4, 125) = 21.8,
P<0.0001; Finteraction (16, 125) = 4.86,
P<0.0001). In WBC 264-9C macrophages, high concentrations of
curcumin resulted in 12.1–20.6% inhibition of cell viability (40
μM, P<0.0001 and 80 μM, P<0.0001, respectively). However, low
concentrations of curcumin (10 and 20 μM) did not cause a
significant inhibition of cell viability. Therefore, 20 μg/ml P19,
and 10 and 20 μM curcumin were chosen for the following
experiments.
Low concentrations of curcumin exert a
protective effect against P19-induced inhibition of viability in
macrophages
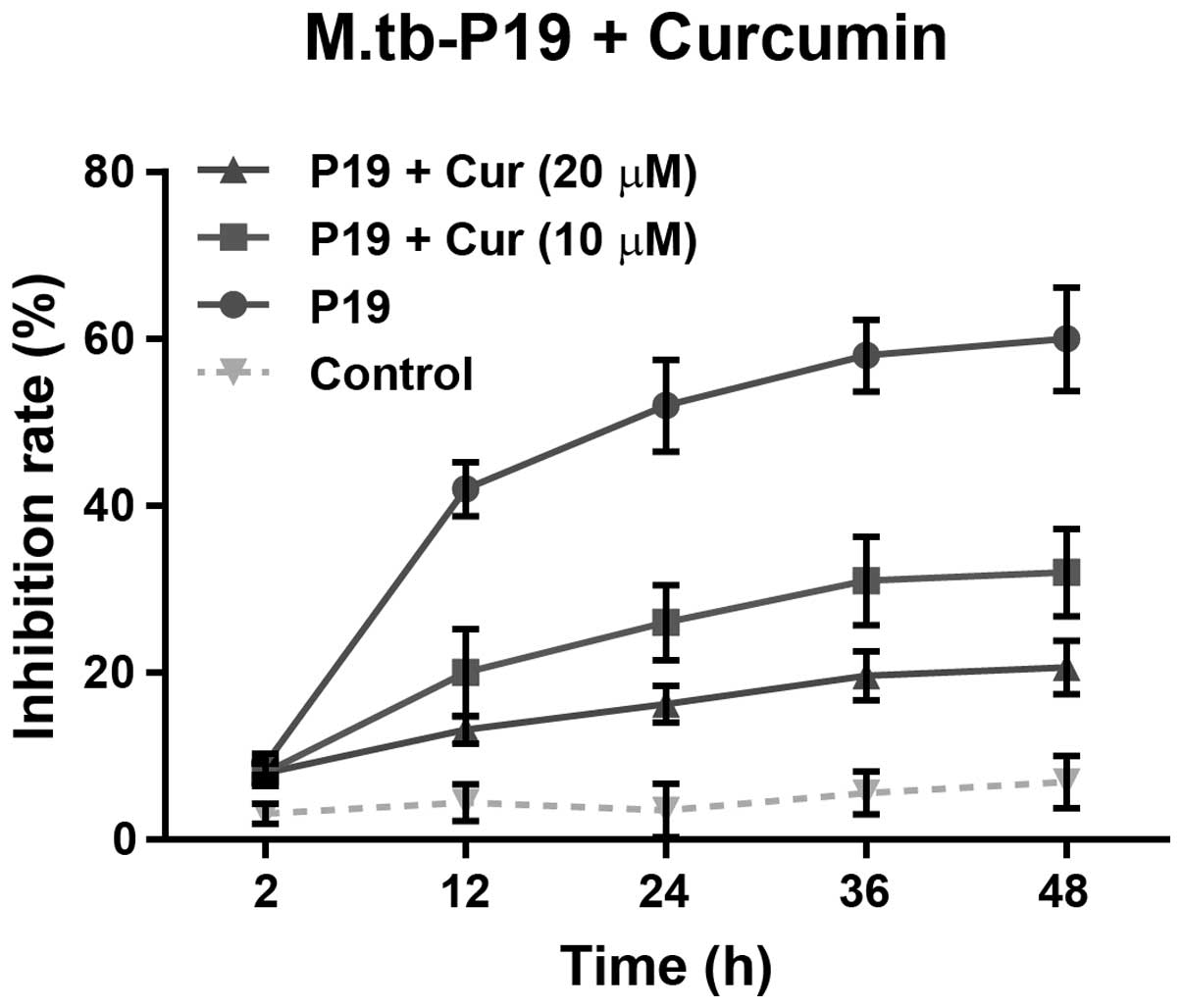
Human macrophages were exposed to vehicle (saline)
treatment as a control, or to 20 μg/ml P19 alone or combined with
10 or 20 μM curcumin for 48 h. As expected, P19 treatment alone
markedly reduced the cell viability (58.7%, P<0.0001) (Fig. 2). Notably, the combined treatment
with P19 and a low concentration of curcumin (10 μM) attenuated the
inhibition of cell viability (32.2%, P<0.0001), compared to P19
treatment alone (Fig. 2). A higher
concentration of curcumin (20 μM) further decreased (20.6%,
P<0.0001) the inhibition in cell viability induced by P19.
Curcumin attenuates P19-induced cytokine
production in macrophages
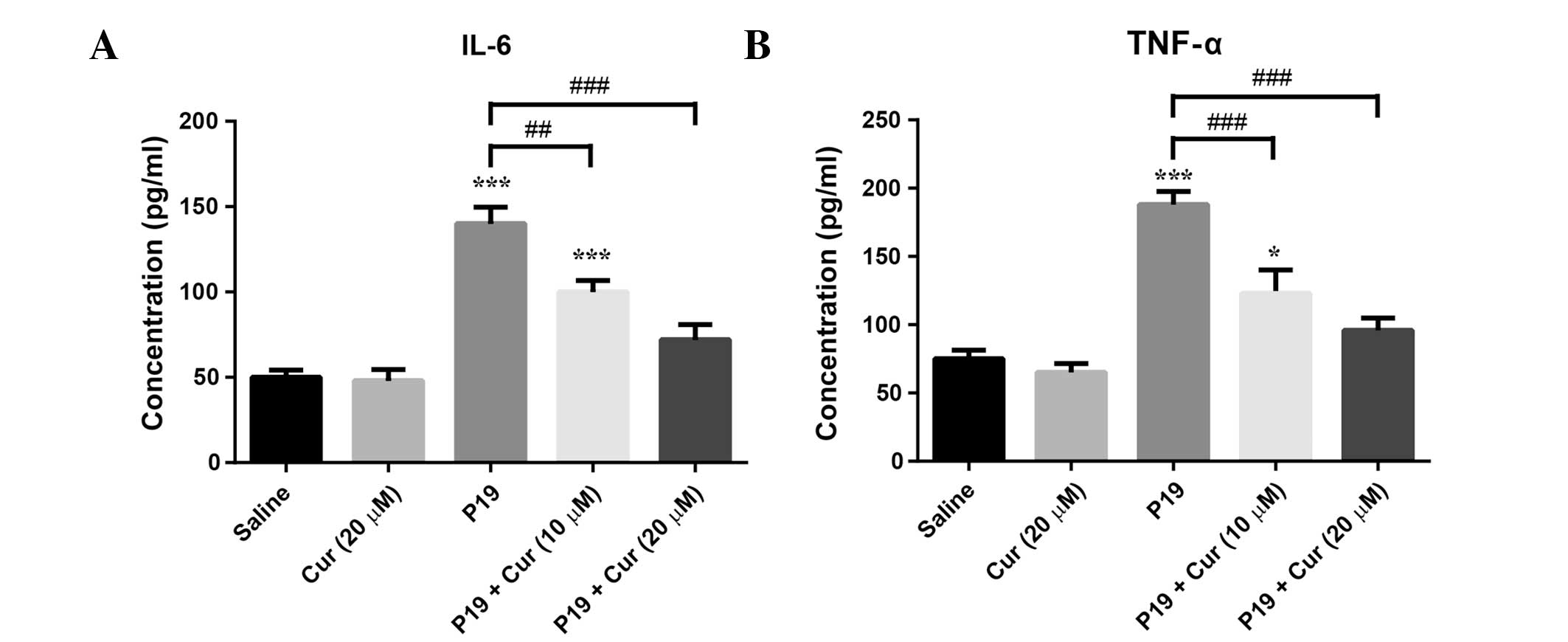
ELISA was used to measure the IL-6 and TNF-α levels
in the macrophages treated with 20 μg/ml P19 alone or combined with
10 or 20 μM curcumin for 48 h (Fig.
3). The IL-6 and TNF-α levels were not significantly affected
by treatment with 20 μM curcumin alone. As expected, IL-6 and TNF-α
levels were significantly elevated upon P19 treatment
(P<0.0001), whereas the combination of 10 μM curcumin and P19
significantly decreased the P19-induced IL-6 and TNF-α production
(IL-6, P<0.01; TNF-α, P<0.0001). Strikingly, treatment with
20 μM curcumin completely offset the P19-induced cytokine
production.
Curcumin attenuates P19-induced
phosphorylation of JNK and p38
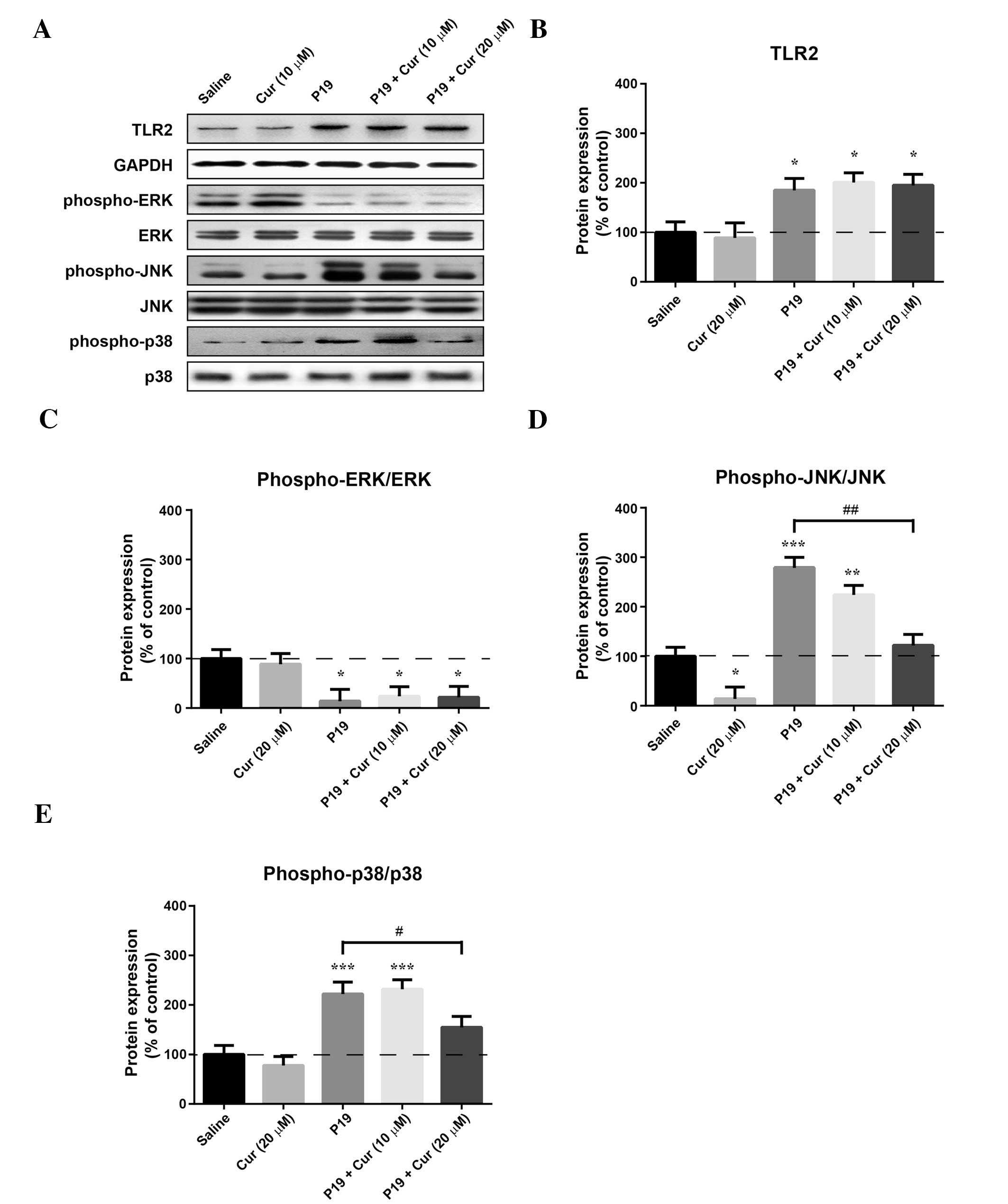
To investigate the mechanism underlying the
protective effects of curcumin against the P19-induced inflammatory
responses, the expression of TLR2 and the phosphorylation of ERK,
JNK and p38 were measured by western blot analysis (Fig. 4A). The expression of TLR2 did not
significantly change following treatment with 20 μM curcumin
(Fig. 4B), whereas treatment with
20 μg/ml P19 significantly increased the TLR2 level (P<0.05).
The P19-induced increase in the TLR2 level was not affected by
cotreatment with 10 or 20 μM curcumin.
As shown in Fig. 4,
P19 decreased the phospho-ERK level (P<0.05), and increased the
phospho-JNK (P<0.0001) and the phospho-p38 (P<0.0001) levels.
Although treatment with 20 μM curcumin alone had no significant
effect on the p38 basal level, its phosphorylation was
significantly decreased relative to treatment with P19 upon
treatment with 20 μM curcumin (P<0.05). Furthermore, the
phosphorylation of JNK was decreased by both curcumin treatment
alone (P<0.05) and when curcumin was combined with P19.
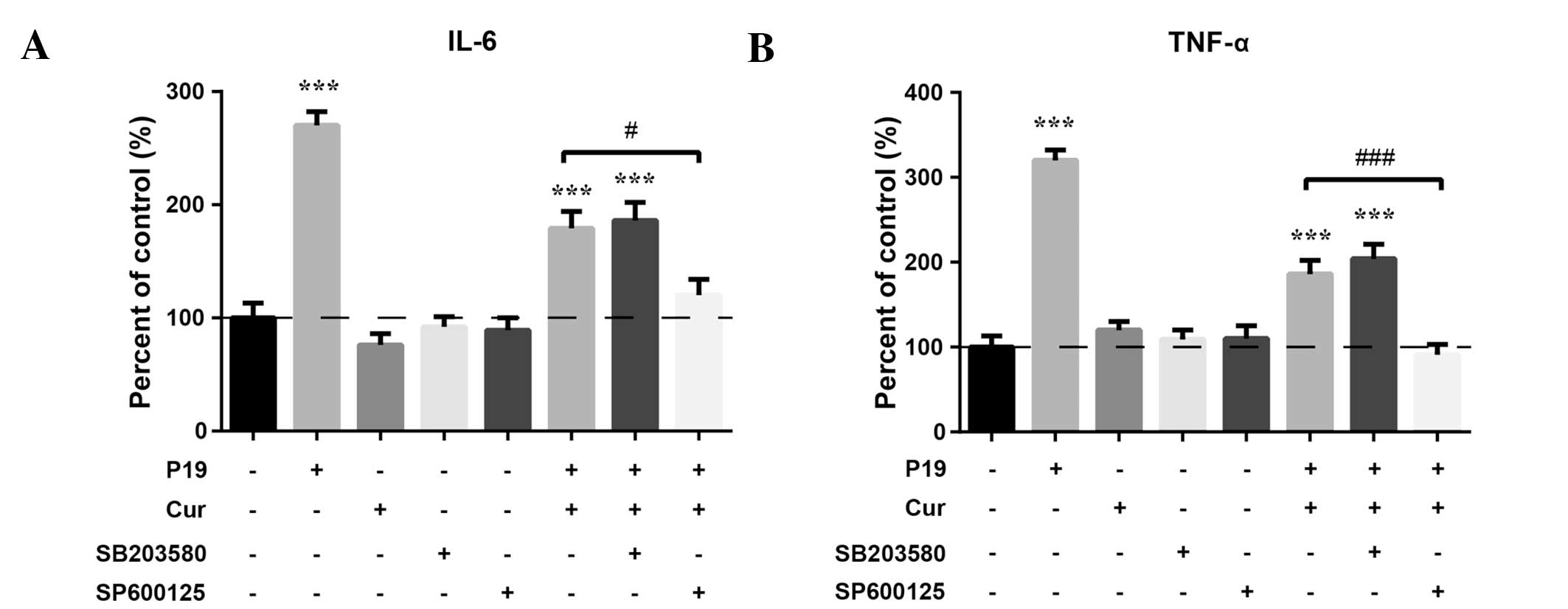
Pharmacological inhibition of JNK, but
not p38, enhances the protective effect of curcumin against
P19-induced inflammatory responses
To determine the role of JNK and p38 phosphorylation
in P19-induced inflammatory responses, we pre-treated the
macrophages with 20 μM SP600125 (the JNK-specific inhibitor) or 30
μM SB203580 (the p38-specific inhibitor) for 1 h prior to treatment
with 20 μM curcumin and 20 μg/ml P19. Cytokine production was
determined by ELISA. Fig. 5 shows
that treatment with the JNK inhibitor significantly decreased IL-6
(P<0.05) and TNF-α (P<0.01) levels, compared to the
combination of curcumin and P19. IL-6 and TNF-α levels were
slightly reduced after treatment with the p38 inhibitor, but the
difference was not significant. These results demonstrated that
activation of JNK, but not p38, may play a role in the protective
effect of curcumin against P19-induced inflammatory responses.
Discussion
P19 plays an important role in the apoptosis of
macrophages during M. tuberculosis infection. Induction of
macrophage apoptosis by M. tuberculosis and thus, evasion
from host defense, has been proposed as one of the mechanisms
promoting M. tuberculosis virulence. Recently, the
contribution of P19 in human M. tuberculosis infections
received increasing attention. Elucidating the mechanism underlying
P19-induced macrophage death during M. tuberculosis
infections is important for the development of new
anti-tuberculosis strategies. Furthermore, polyphenols such as
curcumin possess numerous desirable qualities for anti-tuberculosis
therapy.
Synthetic bacterial lipoproteins have been reported
to induce macrophage apoptosis through TLR2, which implies that the
mycobacterial 19-kDa lipoprotein (P19) may be a M.
tuberculosis apoptosis-inducing factor. P19 is an abundantly
expressed, cell wall-associated and secreted glycolipoprotein, with
a highly conserved six-residue consensus sequence for lipidation
adjacent to the hydrophobic signal peptide. Lopez et al
showed that P19 induces apoptosis in the human monocytic cell line
THP-1 and in human monocyte-derived macrophages (22). Consistently, our data showed that
P19 induces the apoptosis of macrophages in a time- and
dose-dependent manner. The apoptosis-inducing effect of P19 may be
TLR2-dependent, since pre-treatment with blocking anti-TLR2
antibody was previously shown to inhibit the P19-induced apoptosis
in THP-1 cells or TLR2-expressing Chinese hamster ovary cells
(22). Recently, Sanchez et
al revealed a novel mechanism for the M.
tuberculosis-induced death of macrophages, whereby P19 triggers
an intrinsic or mitochondrial pathway, associated with cytochrome
C and the apoptosis-inducing mitochondrial factor (11). Overall, P19 may induce macrophage
apoptosis.
Curcumin has been known to inhibit the proliferation
and/or induce cell death in a number of human cancer cell lines
(23–27). Interestingly, curcumin is
non-cytotoxic to healthy cells, and its pharmacological safety is
well demonstrated by the fact that people in certain countries have
consumed curcumin as a dietary spice for centuries in amounts
exceeding 10 mg/day without any side-effects (28). The present study provided evidence
that curcumin can induce the death of macrophages in a dose- and
time-dependent manner. Although the fact that curcumin induces
apoptosis is well established, the exact mechanism underlying this
effect is not yet fully elucidated. Potential mechanisms of
curcumin-induced apoptosis include inhibition of various protein
kinases such as PKC, JNK, and the EGF receptor kinase. Curcumin can
also activate apoptotic pathways, such as p53-mediated apoptosis,
in human basal cell carcinoma cells under stress conditions
(29). Mitochondrial dysfunction
triggered by enhanced Bax expression is also involved in
curcumin-induced apoptosis (30,31).
Moreover, curcumin can block the nuclear factor-κB (NF-κB) cell
survival pathway (32,33). However, the mechanism underlying
curcumin-induced apoptosis has been mainly revealed from tumor cell
lines, whereas the mechanism underlying curcumin-induced apoptosis
in macrophages still remains unclear.
We found that low concentrations of curcumin (10 and
20 μM) exert a protective effect against P19-induced apoptosis in
macrophages, and attenuate P19-induced cytokine production. The
protective effect may be mediated by JNK inhibition, since JNK, but
not p38 inhibitors enhanced the protective effect of curcumin.
Although additional data are needed to clarify the protective
effect of curcumin against P19-induced apoptosis, we can conclude
that curcumin and P19 may play opposing roles in inducing
macrophage apoptosis. TLR2 receptor-induced JNK activation is one
of the candidate signaling pathways involved in P19-induced
apoptosis. In human monocytes and neutrophils, curcumin treatment
decreased the gene expression of TLR2 and impaired the
function of the protein (19). By
contrast, P19 was shown to induce TLR2 activation, followed by JNK
activation (11,34).
In summary, we demonstrated that both P19 and high
doses of curcumin can decrease the viability of human macrophages.
Low doses of curcumin reduce the P19-induced apoptosis and the
increased cytokine levels in the macrophages. The protective effect
of curcumin may be attributed to its ability to block the
TLR2-mediated JNK activation induced by P19. Our study has shed
some light on the mechanism underlying the protective effect of
curcumin against P19-induced macrophage apoptosis, and suggests
that curcumin may be used as a therapeutic agent for the treatment
of tuberculosis.
References
|
1
|
Dye C, Scheele S, Dolin P, Pathania V and
Raviglione MC: Consensus statement. Global burden of tuberculosis:
estimated incidence, prevalence, and mortality by country. WHO
Global Surveillance and Monitoring Project. JAMA. 282:677–686.
1999. View Article : Google Scholar
|
|
2
|
Bhatt K and Salgame P: Host innate immune
response to Mycobacterium tuberculosis. J Clin Immunol.
27:347–362. 2007. View Article : Google Scholar
|
|
3
|
Akira S, Uematsu S and Takeuchi O:
Pathogen recognition and innate immunity. Cell. 124:783–801. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Underhill DM, Ozinsky A, Smith KD and
Aderem A: Toll-like receptor-2 mediates mycobacteria-induced
proinflammatory signaling in macrophages. Proc Natl Acad Sci USA.
96:14459–14463. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Quesniaux V, Fremond C, Jacobs M, et al:
Toll-like receptor pathways in the immune responses to
mycobacteria. Microbes Infect. 6:946–959. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Holscher C, Reiling N, Schaible UE, et al:
Containment of aerogenic Mycobacterium tuberculosis infection in
mice does not require MyD88 adaptor function for TLR2, -4 and -9.
Eur J Immunol. 38:680–694. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cole ST, Brosch R, Parkhill J, et al:
Deciphering the biology of Mycobacterium tuberculosis from
the complete genome sequence. Nature. 393:537–544. 1998.
|
|
8
|
Boom WH, Husson RN, Young RA, David JR and
Piessens WF: In vivo and in vitro characterization of murine T-cell
clones reactive to Mycobacterium tuberculosis. Infect Immun.
55:2223–2229. 1987.PubMed/NCBI
|
|
9
|
Brightbill HD, Libraty DH, Krutzik SR, et
al: Host defense mechanisms triggered by microbial lipoproteins
through toll-like receptors. Science. 285:732–736. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Diaz-Silvestre H, Espinosa-Cueto P,
Sanchez-Gonzalez A, et al: The 19-kDa antigen of Mycobacterium
tuberculosis is a major adhesin that binds the mannose receptor
of THP-1 monocytic cells and promotes phagocytosis of mycobacteria.
Microb Pathog. 39:97–107. 2005.
|
|
11
|
Sanchez A, Espinosa P, Garcia T and
Mancilla R: The 19 kDa Mycobacterium tuberculosis lipoprotein
(LpqH) induces macrophage apoptosis through extrinsic and intrinsic
pathways: a role for the mitochondrial apoptosis-inducing factor.
Clin Dev Immunol. 2012:9505032012. View Article : Google Scholar
|
|
12
|
Kuo ML, Huang TS and Lin JK: Curcumin, an
antioxidant and anti-tumor promoter, induces apoptosis in human
leukemia cells. Biochim Biophys Acta. 1317:95–100. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Goel A, Kunnumakkara AB and Aggarwal BB:
Curcumin as ‘Curecumin’: from kitchen to clinic. Biochem Pharmacol.
75:787–809. 2008.
|
|
14
|
Marathe SA, Dasgupta I, Gnanadhas DP and
Chakravortty D: Multifaceted roles of curcumin: two sides of a
coin! Expert Opin Biol Ther. 11:1485–1499. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bukhari SN, Franzblau SG, Jantan I and
Jasamai M: Current prospects of synthetic curcumin analogs and
chalcone derivatives against Mycobacterium tuberculosis. Med
Chem. 9:897–903. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kawamori T, Lubet R, Steele VE, et al:
Chemopreventive effect of curcumin, a naturally occurring
anti-inflammatory agent, during the promotion/progression stages of
colon cancer. Cancer Res. 59:597–601. 1999.PubMed/NCBI
|
|
17
|
Lu YP, Chang RL, Lou YR, et al: Effect of
curcumin on 12-O-tetradecanoylphorbol-13-acetate- and ultraviolet B
light-induced expression of c-Jun and c-Fos in JB6 cells and in
mouse epidermis. Carcinogenesis. 15:2363–2370. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Azuine MA and Bhide SV: Chemopreventive
effect of turmeric against stomach and skin tumors induced by
chemical carcinogens in Swiss mice. Nutr Cancer. 17:77–83. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shuto T, Ono T, Ohira Y, et al: Curcumin
decreases toll-like receptor-2 gene expression and function in
human monocytes and neutrophils. Biochem Biophys Res Commun.
398:647–652. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schlesinger LS, Bellinger-kawahara CG,
Payne NR and Horwitz MA: Phagocytosis of Mycobacterium
tuberculosis is mediated by human monocyte complement receptors
and complement component-C3. J Immunol. 144:2771–2780.
1990.PubMed/NCBI
|
|
21
|
Sanchez A, Espinosa P, Garcia T and
Mancilla R: The 19 kDa Mycobacterium tuberculosis lipoprotein
(LpqH) induces macrophage apoptosis through extrinsic and intrinsic
pathways: a role for the mitochondrial apoptosis-inducing factor.
Clin Dev Immunol. 2012:9505032012. View Article : Google Scholar
|
|
22
|
Lopez M, Sly LM, Luu Y, Young D, Cooper H
and Reiner NE: The 19-kDa Mycobacterium tuberculosis protein
induces macrophage apoptosis through toll-like receptor-2. J
Immunol. 170:2409–2416. 2003.
|
|
23
|
Thangapazham RL, Sharma A and Maheshwari
RK: Multiple molecular targets in cancer chemoprevention by
curcumin. AAPS J. 8:E443–E449. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Singh RP and Agarwal R: Mechanisms of
action of novel agents for prostate cancer chemoprevention. Endocr
Relat Cancer. 13:751–778. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Salehi P, Makhoul G, Roy R, Malhotra M,
Mood ZA and Daniel SJ: Curcumin loaded NIPAAM/VP/PEG-A
nanoparticles: physicochemical and chemopreventive properties. J
Biomater Sci Polym Ed. 24:574–588. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lopez-Lazaro M: Anticancer and
carcinogenic properties of curcumin: considerations for its
clinical development as a cancer chemopreventive and
chemotherapeutic agent. Mol Nutr Food Res. 52(Suppl 1): S103–S127.
2008.
|
|
27
|
Campbell FC and Collett GP:
Chemopreventive properties of curcumin. Future Oncol. 1:405–414.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ammon HP and Wahl MA: Pharmacology of
Curcuma longa. Planta Med. 57:1–7. 1991.
|
|
29
|
Jee SH, Shen SC, Tseng CR, Chiu HC and Kuo
ML: Curcumin induces a p53-dependent apoptosis in human basal cell
carcinoma cells. J Invest Dermatol. 111:656–661. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Uddin S, Hussain AR, Manogaran PS, et al:
Curcumin suppresses growth and induces apoptosis in primary
effusion lymphoma. Oncogene. 24:7022–7030. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sen S, Sharma H and Singh N: Curcumin
enhances Vinorelbine mediated apoptosis in NSCLC cells by the
mitochondrial pathway. Biochem Biophys Res Commun. 331:1245–1252.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Singh S and Aggarwal BB: Activation of
transcription factor NF-kappa B is suppressed by curcumin
(diferuloylmethane) [corrected]. J Biol Chem. 270:24995–25000.
1995.
|
|
33
|
Bush JA, Cheung KJ Jr and Li G: Curcumin
induces apoptosis in human melanoma cells through a Fas
receptor/caspase-8 pathway independent of p53. Exp Cell Res.
271:305–314. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Watanabe I, Ichiki M, Shiratsuchi A and
Nakanishi Y: TLR2-mediated survival of Staphylococcus aureus
in macrophages: a novel bacterial strategy against host innate
immunity. J Immunol. 178:4917–4925. 2007.PubMed/NCBI
|