Introduction
Glucose-dependent insulinotropic polypeptide (GIP)
is an incretin hormone, secreted by enteroendocrine K-cells, with
established effects on insulin secretion and pancreatic β-cell
function (1). Furthermore, GIP and
its receptor (GIPR) have been identified in several areas of the
brain, including the cortex, cerebellum and hippocampus (2–4).
However, the role of GIP within the central nervous system remains
to be elucidated. To date, GIP has been observed to exhibit a
number of neuroprotective effects, inducing progenitor cell
proliferation, improving learning and memory and enhancing synaptic
plasticity (4,5). Indeed, the genetically-induced
overexpression of GIP in mice improves their cognitive function
(6). In addition, mice with a
targeted deletion of the GIPR have been reported to have less than
half number of newly proliferating cells in the hippocampus
(4) and exhibit impaired learning
and memory, compared with the wild-type (WT) controls (7). Overall, these preliminary results
suggested that the GIPR is important in cognitive function in the
central nervous system, however, the underlying mechanisms remain
to be elucidated.
Evidence from previous studies has highlighted the
association between obesity, type 2 diabetes and cognitive
dysfunction (8–10). Previous studies in genetically
obese ob/ob mice, Zucker fa/fa rats and high-fat diet fed
obese-diabetic mice, have consistently reported impaired
performance in learning and memory tests (11–14).
Furthermore, a clear association has been demonstrated between
chronic high-fat feeding, the increased secretion of GIP and the
development of insulin resistance and obesity-associated diabetes
(15). With the incidence of human
obesity-associated diabetes expected to rise significantly, the
occurrence of cognitive deficit disorders, including Alzheimer’s
disease, is also likely to increase (16). Therefore, it is necessary to
elucidate the underlying pathological mechanisms and putative
associations between a high-fat diet, GIPR signalling and cognitive
decline.
The present study evaluated the role of GIPR
signalling and a chronic high-fat diet, alone and in combination,
on several aspects of cognitive function in mice. This included
investigating learning and memory through object recognition tests
(ORTs) and assessing the mechanisms involved by analysing the
expression levels of key hippocampal genes, which are involved in
learning and memory formation. These investigations aimed to
determine the importance of GIPR signalling and dietary composition
in controlling the fundamental aspects of memory and learning.
Materials and methods
Animals
Male C57BL/6 mice with genetic deletion of the GIPR
were used in the present study, in addition to WT controls. These
mice (32 mice in total, 10 GIPR KO and 10 WT mice on a high fat
diet, 6 GIPR KO and WT mice on a normal diet; 6–8 weeks old;
n=6-10) were derived from an in-house breeding colony, as described
previously (17,18) and were age-matched and housed in an
air-conditioned room at 22±2°C with a 12 h light/dark cycle
(08:00-20:00 h). The experimental animals had free access to
drinking water, a standard rodent diet (10% fat, 30% protein and
60% carbohydrate; percentage of total energy of 12.99 kj/g; Trouw
Nutrition, Cheshire, UK) or a high-fat diet (45% fat, 20% protein
and 35% carbohydrate; percent of total energy of 26.15 kj/g;
Special Diets Service, Essex, UK), as appropriate. Prior to
commencement of experiments, all the mice were weighed and
maintained on their respective diets for 120 days. The WT animals,
which were fed a high-fat diet exhibited significantly increased
body weight compared with the controls. All experiments were
performed according to UK Home Office Regulations (UK Animals
Scientific Procedures Act 1986), the ‘Principles of Laboratory
Animal Care’ (NIH publication No. 86–23, revised 1985) and were
approved by the University of Ulster Animal Ethics Review
Committee.
Object recognition
At the end of the dietary intervention, the groups
of mice were subjected to ORTs, as described previously (14). Briefly, the mice were initially
habituated to the exploratory arena (58 cm diameter, 38 cm high)
for 5 mins. Two identical random objects (2 marbles, 2.5 cm
diameter or 2 die, 1.2 cm side length) were subsequently placed in
the centre of the arena and, following 4 h exposure (the
acquisition phase), one of the two objects was replaced by a novel
object (a marble or dice) and the duration spent exploring each
object during a 5 min trial phase was determined using a
computerised tracking system (Tracker, Biosignal Group, New York,
USA). The recognition index (RI), which was designated as the
period of time spend exploring the novel object as opposed to the
familiar object, was calculated, as described previously(19).
Hippocampal gene expression
Animals were sacrificed by lethal inhalation of
CO2 followed by cervical dislocation. Hippocampal tissue
was excised at the end of the dietary intervention period by
careful surgical excision, snap frozen and processed for gene
expression by reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) following total RNA extraction. The total RNA
was extracted and purified using Tripure Isolation reagent (Roche
Diagnostics, West Sussex, UK). The concentration and purity of the
extracted RNA was determined using a nanophotometer
(NanoPhotometer™ Pearl; Implen, Munich, Germany) at an absorbance
of 260 nm. The cDNA was synthesised (final concentration, 1.5
μg) using a Transcriptor First Strand cDNA Synthesis kit
(Roche Diagnostics) according to the manufacturer’s instructions.
Gene expression analysis was performed using a Roche RealTime ready
qPCR assay, LightCycler 480 Probes Master and a hot start reaction
mix (Roche Diagnostics). The following RT-qPCR target specific
primers were used: Glucagon-like peptide-1 receptor (GLP-1R);
mammalian target of rapamycin (mTOR); neurotrophic tyrosine kinase
receptor type 2 (NTRK2); sirtuin 1 (SIRT1); synaptophysin (SYP) and
vascular endothelial growth factor (VEGF). The gene expression was
normalised to the expression of hypoxanthine guanine phosphoribosyl
transferase. Briefly, the specific primer (10 pmol in 20 μl
reaction volume, 0.5 pmol/μl) and cDNA (4 0 ng/μl.)
were added to each well (containing fluorescein-labelled short
hydrolysis probes and PCR-grade water) to a final reaction volume
of 10 μl. The PCR conditions were 95°C for 10 min, followed
by cDNA amplification for 45 cycles with 95°C denaturation for 10
sec, 60°C annealing for 30 sec and 72°C elongation for 1 sec,
followed by 30 sec cooling at 40°C. The relative quantification was
calculated using the 2−ΔΔCT method, to determine the
differences in gene expression levels between the samples (20).
Statistical analysis
One-way analysis of variance, followed by
Newman-Keuls post-hoc test were used for statistical analysis using
Prism 5 software (Graph-Pad Prism®, San Diego, CA, USA).
The data are expressed as the mean ± standard error of the mean.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effects of GIPR KO and high-fat diet
alone, and in combination, on object recognition
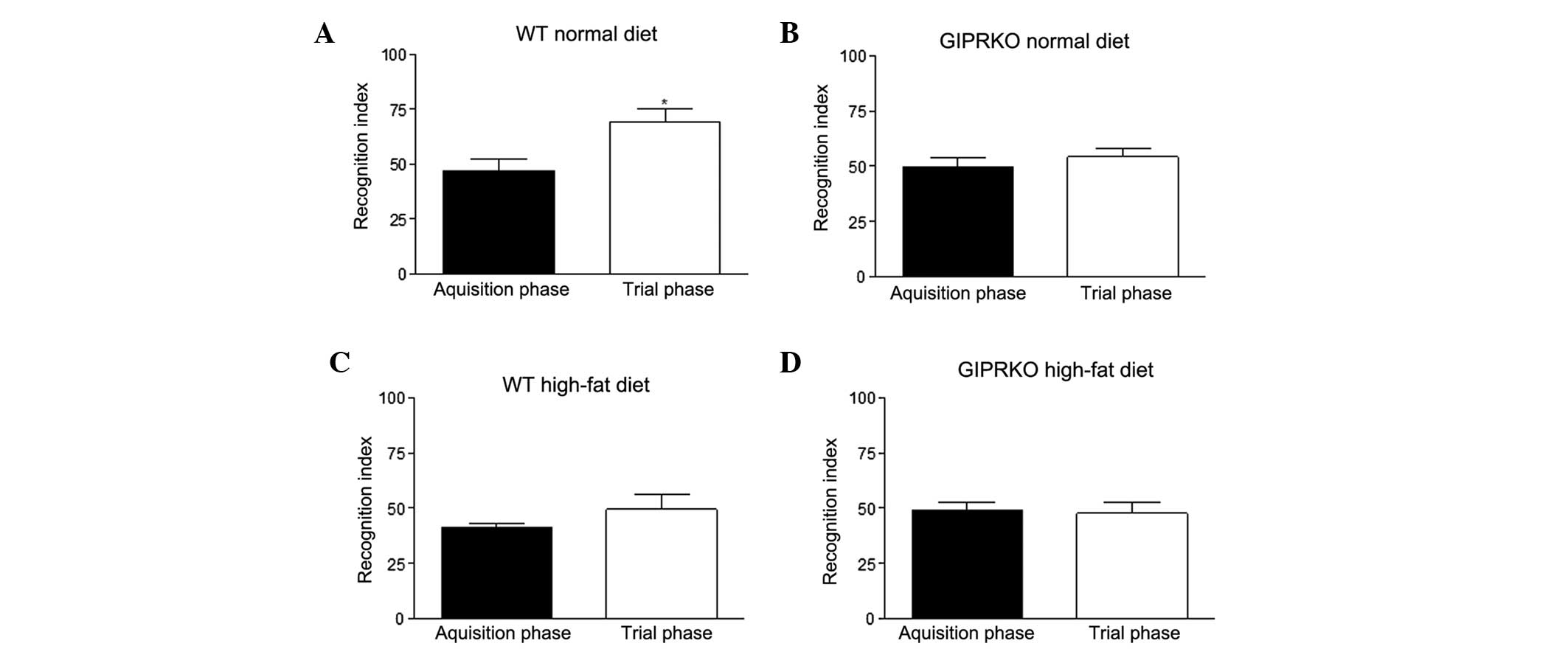
In the ORT, no difference or bias was observed in
the RI between any of the groups of mice during the acquisition
period. During the trial phase, the WT mice fed a normal diet had
significantly (P<0.05) increased RI compared with the
acquisition phase (Fig. 1A),
indicating a preference to examine the novel object. By contrast,
the WT mice fed a high-fat diet and the GIPR KO mice fed either the
standard maintenance or the high-fat diet failed to discriminate
between the novel and the familiar object during the trial phase
(Fig. 1B–D).
Effects of GIPR KO and high-fat feeding
alone, and in combination, on hippocampal gene expression
levels
The assessment of the hippocampal gene expression
levels revealed that a high-fat diet significantly decreased the
expression levels of mTOR, NTRK2 and SYP (P<0.001, P<0.05 and
P<0.05; respectively) in the WT mice (Fig. 2A–C). Notably, the GIPR KO mice fed
a standard rodent diet also exhibited significantly (P<0.05)
decreased hippocampal expression of mTOR compared with the WT mice
(Fig. 2A). Similarly, the GIPR KO
mice fed a high-fat diet exhibited significantly (P<0.05)
decreased expression of mTOR, however, this was higher (P<0.001)
compared with the WT mice fed a high-fat diet (Fig. 2A). In addition, the GIPR KO mice
fed a high-fat diet also exhibited decreased hippocampal gene
expression of NTRK2 compared with the WT controls (Fig. 2B). The expression levels of
hippocampal GLP-1R, SIRT1 and VEGF were not significantly different
in any of the groups of mice (Fig. 2D
and E).
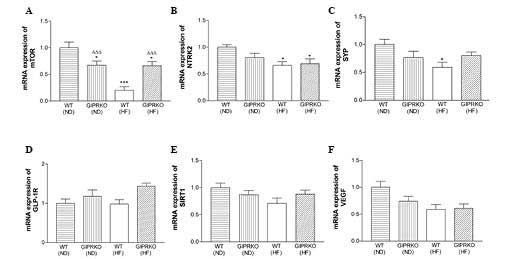 | Figure 2Effects of GIPR KO alone or in
combination with a high-fat diet on hippocampal gene expression.
The mRNA expression levels of (A) mTOR, (B) NTRK2, (C) SYP, (D)
GLP-1R, (E) SITR1 and (F) VEGF were examined following 120 days of
dietary intervention. The data are expressed as the mean ± standard
error of the mean for six mice (*P<0.05 and ***P<0.001,
compared with WT mice fed a normal diet; ΔΔΔP<0.001,
compared with WT mice fed a high-fat diet). WT, wild-type; GIPR,
glucose-dependent insulinotropic polypeptide receptor; KO,
knockout; ND, normal diet; HD, high-fat diet; mTOR, mammalian
target of rapamycin; NTRK2, neurotrophic tyrosine kinase receptor
type 2; SYP, synaptophysin; GLP-1R, glucagon-like peptide 1
receptor; SITR1, sirtuin 1; VEGF, vascular endothelial growth
factor. |
Discussion
Chemical and genetic reduction of GIPR signalling,
or chronic consumption of a high-fat diet, have been demonstrated
independently to impair learning and memory ability (4,7,12,13).
However, the precise mechanisms and pathologies underlying these
phenomena remain to be elucidated. The present study investigated
the effects of genetic GIPR KO alone, and in combination with a
chronic high-fat diet, on recognition memory and the expression
levels of a panel of hippocampal genes, which are involved in
maintaining normal cognitive function.
The ORT is a popular and informative tool used for
assessing recognition memory in rodents (21). In the present study, WT mice
maintained on a standard rodent maintenance diet exhibited normal
recognition memory, preferring to investigate the novel object
during ORTs. In accordance with previous findings, the GIPR KO mice
or mice fed a chronic high-fat diet demonstrated significantly
impaired recognition memory (7,13).
Therefore, it was not unexpected that the GIPR KO mice fed a
high-fat diet also exhibited markedly reduced memory function.
However, the absence of additive detrimental effects of GIPR
annulment combined with a high-fat diet on recognition memory,
suggested that similar pathways may be involved in mediating this
action, although this requires further investigation.
Normal hippocampal function is particularly
important for the preservation of recognition memory (22). Experimental evidence suggested that
the deleterious effect of chronic high-fat feeding on learning and
memory is associated with altered hippocampal gene expression
levels (23). Furthermore, the
GIPR KO mice exhibited significant reductions in progenitor cell
proliferation in the dentate gyrus of the hippocampus (7). Notably, GIPR KO mice and mice fed a
chronic high-fat diet have previously been demonstrated to inhibit
long term potentiation (LTP) in the hippocampus, the major cellular
mechanism underlying learning and memory (7,13).
Therefore, to extend this previous investigation, the present study
investigated the effects of GIPR KO alone, and in combination with
a high-fat diet, on the expression levels of hippocampal genes,
which are known to be involved in cognitive function.
In the present study, the hippocampal expression of
mTOR, a protein kinase with an established role in the maintenance
of LTP (24), was significantly
reduced by GIPR KO and a high-fat diet. Therefore, the expression
of mTOR and subsequent activity clearly have major implications on
the cognitive ability of each of these groups of mice. Notably,
GIPR KO partially reversed the severe detrimental effect of a
high-fat diet on the expression of mTOR, although the expression
levels remained reduced compared with the WT controls. This may
appear marginally contradictory, however, it may be linked to the
opportunity for lifelong adaptive mechanisms in genetic KO animals
(17). GLP-1 action, also
associated with positive effects on cognition (13), has previously been observed to be
upregulated in GIPR KO mice (17).
In the present study, increased hippocampal expression of GLP-1R
was observed in each group of GIPR KO mice, however was not
statistically significant. Similarly, the hippocampal expression of
SYP was significantly reduced by a chronic high-fat diet, however,
concurrent GIPR KO restored the expression of SYP almost to normal
levels. SIRT1 has previously been revealed as an important factor
for normal cognitive function and object recognition (23,25),
however, the hippocampal expression of SIRT1 was unaffected by GIPR
KO or a high-fat diet in the present study.
NTRK2, a gene encoding for the tyrosine kinase
receptor, TrkB, on neuronal cells, is important in hippocampal
synaptic plasticity and neurogenesis (26). The present study revealed that the
chronic consumption of a high-fat diet by the WT or GIPR KO mice
significantly reduced the hippocampal expression of NTRK2.
Therefore, a potential mechanism for the reported impairment of
neurogenesis by high-fat feeding (27) may be associated with a reduced
expression of NTRK2, which may ultimately decrease the activity of
its ligand, brain-derived neurotrophic factor (BDNF). BDNF has well
recognised beneficial actions on the growth and differentiation of
neurons and synapses (28).
Notably, the expression of VEGF, which is important in hippocampal
neurogenesis (29,30), was not significantly reduced in all
GIPR KO and high-fat fed mice. Taken together, GIPR KO deletion and
a chronic high-fat diet may impair hippocampal neurogenesis via
complementary pathways.
In conclusion, GIPR KO and chronic consumption of a
high-fat diet exhibited negative effects on hippocampal-dependent
recognition memory. The pathways involved in the maintenance of
hippocampal synaptic plasticity, including SYP, NTRK2 and
particularly mTOR, appeared to be key in this respect. In addition,
the results suggested that a deficit in hippocampal neurogenesis
may also be important in the observed reduction in recognition
memory. These findings highlight the importance of GIPR signalling
and dietary content for the maintenance of normal cognitive
function.
Acknowledgments
The present study was supported by the Department of
Education and Learning, Northern Ireland, and a grant from the
European Foundation for the Study of Diabetes/GlaxoSmithKline. The
GIPR KO mice were derived from breeding pairs, which were kindly
supplied by Professor B Thorens (Lausanne, Switzerland).
References
|
1
|
Gault VA, O’Harte FP and Flatt PR:
Glucose-dependent insulinotropic polypeptide (GIP): anti-diabetic
and anti-obesity potential? Neuropeptides. 37:253–263. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Usdin TB, Mezey E, Button DC, Brownstein
MJ and Bonner TI: Gastric inhibitory polypeptide receptor, a member
of the secretin-vasoactive intestinal peptide receptor family, is
widely distributed in peripheral organs and the brain.
Endocrinology. 133:2861–2870. 1993.PubMed/NCBI
|
|
3
|
Nyberg J, Anderson MF, Meister B, et al:
Glucose-dependent insulinotropic polypeptide is expressed in adult
hippocampus and induces progenitor cell proliferation. J Neurosci.
25:1816–1825. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nyberg J, Jacobsson C, Anderson MF and
Eriksson PS: Immunohistochemical distribution of glucose-dependent
insulinotropic polypeptide in the adult rat brain. J Neurosci Res.
85:2099–2119. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Faivre E, Hamilton A and Hölscher C:
Effects of acute and chronic administration of GIP analogues on
cognition, synaptic plasticity and neurogenesis in mice. Eur J
Pharmacol. 674:294–306. 2012. View Article : Google Scholar
|
|
6
|
Ding KH, Zhong Q, Xie D, et al: Effects of
glucose-dependent insulinotropic peptide on behavior. Peptides.
27:2750–2755. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Faivre E, Gault VA, Thorens B and Hölscher
C: Glucose-dependent insulinotropic polypeptide receptor knockout
mice are impaired in learning, synaptic plasticity and
neurogenesis. J Neurophysiol. 105:1574–1580. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cukierman T, Gerstein HC and Williamson
JD: Cognitive decline and dementia in diabetes-systematic overview
of prospective observational studies. Diabetologia. 48:2460–2469.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Luchsinger J, Reitz C, Patel B, et al:
Relation of diabetes to mild cognitive impairment. Arch Neurol.
64:570–575. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Strachan MWJ, Reynolds RM, Frier BM,
Mitchell RJ and Price JF: The relationship between type 2 diabetes
and dementia. Br Med Bull. 88:131–146. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Greenwood CE and Winocur G: Glucose
treatment reduces memory deficits in young adult rats fed high-fat
diets. Neurobiol Learn Mem. 75:179–189. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Biessels GJ and Gispen WH: The impact of
diabetes on cognition: what can be learned from rodent models?
Neurobiol Aging. 26(Suppl 1): 36–41. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gault VA, Porter WD, Flatt PR and Hölscher
C: Actions of exendin-4 therapy on cognitive function and
hippocampal synaptic plasticity in mice fed a high-fat diet. Int J
Obes. 34:1341–1344. 2010. View Article : Google Scholar
|
|
14
|
Porter WD, Flatt PR, Hölscher C and Gault
VA: Liraglutide improves hippocampal synaptic plasticity associated
with increased expression of Mash1 in ob/ob mice. Int J Obes.
37:678–684. 2013. View Article : Google Scholar
|
|
15
|
Irwin N and Flatt PR: Evidence for
beneficial effects of compromised gastric inhibitory polypeptide
action in obesity-related diabetes and possible therapeutic
implications. Diabetologia. 52:1724–1731. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tang J, Pei Y and Zhou G: When aging-onset
diabetes is coming across with Alzheimer disease: comparable
pathogenesis and therapy. Exp Gerontol. 48:744–750. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Preitner F, Ibberson M, Franklin I, et al:
Thorens, Gluco-incretins control insulin secretion at multiple
levels as revealed in mice lacking GLP-1 and GIP receptors. J Clin
Invest. 113:635–645. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mieczkowska A, Irwin N, Flatt PR, Chappard
D and Mabilleau G: Glucose-dependent insulinotropic polypeptide
(GIP) receptor deletion leads to reduced bone strength and quality.
Bone. 56:337–342. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dere E, Huston JP and De Souza Silva MA:
The pharmacology, neuroanatomy and neurogenetics of one-trial
object recognition in rodents. Neurosci Biobehav Rev. 31:673–704.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-Delta Delta CT Method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
21
|
Aggleton JP: One trial object recognition
by rats. Q J Exp Psychol. 37:279–294. 1985. View Article : Google Scholar
|
|
22
|
Broadbent NJ, Squire LR and Clark RE:
Spatial memory, recognition memory and the hippocampus. Learn Mem.
17:5–11. 2010. View Article : Google Scholar :
|
|
23
|
Heyward FD, Walton RG, Carle MS, et al:
Adult mice maintained on a high-fat diet exhibit object location
memory deficits and reduced hippocampal SIRT1 gene expression.
Neurobiol Learn Mem. 98:25–32. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hoeffer CA and Klann E: mTOR signaling: at
the crossroads of plasticity, memory and disease. Trends Neurosci.
33:67–75. 2010. View Article : Google Scholar
|
|
25
|
Michán S, Li Y, Chou MM, et al: SIRT1 is
essential for normal cognitive function and synaptic plasticity. J
Neurosci. 30:9695–9707. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gruart A, Sciar retta C,
Valenzuela-Harrington M, Delgado-García JM and Minichiello L:
Mutation at thr TrkB PLC{gamma}-docking site affects hippocampal
LTP and associative learning in conscious mice. Learn Mem.
14:54–62. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Park HR, Park M, Choi J, Park KY, Chung HY
and Lee J: A high-fat diet impairs neurogenesis: involvement of
lipid peroxidation and brain-derived neurotrophic factor. Neurosci
Lett. 4:235–239. 2010. View Article : Google Scholar
|
|
28
|
Acheson A, Conover JC, Fandl JP, et al: A
BDNF autocrine loop in adult sensory neurons prevents cell death.
Nature. 374:450–453. 1994. View
Article : Google Scholar
|
|
29
|
Cao L, Jiao X, Zuzga DS, et al: VEGF links
hippocampal activity with neurogenesis, learning and memory. Nat
Genet. 36:827–835. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Plaschke K, Staub J, Ernst E and Marti HH:
VEGF overexpression improves mice cognitive abilities after
unilateral common carotid artery occlusion. Exp Neurol.
214:285–292. 2008. View Article : Google Scholar : PubMed/NCBI
|