Introduction
Ischemic injury in the brain leads to neuronal cell
death, and neuronal cell death following cerebral ischemia is
caused by the interaction of complex processes, including
excitotoxicity, depolarization, inflammation and apoptosis
(1,2). Apoptosis is one of the major pathways
leading to cell death in cerebral ischemia injury (3). Cerebral ischemia-induced apoptosis in
the hippocampus causes memory impairment (4–6).
Terminal deoxynucleotidyl transferase-mediated dUTP
nick end labeling (TUNEL) staining detects DNA fragmentation, which
is one of the hallmarks of apoptosis. Two important groups of
proteins in the apoptotic cascades are members of the B-cell
lymphoma (Bcl)-2 family and classes of caspases (7). The Bcl-2 family can be classified
into two functionally distinct groups: Anti-apoptotic proteins and
pro-apoptotic proteins. Bcl-2, an anti-apoptotic protein, is known
to regulate apoptotic pathways and protect against cell death.
Bcl-2-associated X protein (Bax), a pro-apoptotic protein of the
Bcl-2 family, is expressed in high levels and selectively during
apoptosis, and promotes cell death (7–9). The
increment of the ratio of Bax to Bcl-2 represents the induction of
apoptosis in several tissues (8,9).
Caspase-3 is one of the key executors of apoptosis, and activation
of caspase-3 is implicated in apoptotic neuronal cell death in
animal models of stroke (1,10).
The adult central nervous system contains stem
cells, which are capable of generating new neurons. It has been
suggested that increased neurogenesis in the hippocampal dentate
gyrus may promote morphological and functional recovery (11,12).
5-Bromo-2′-deoxyuridine (BrdU) is an indicator of DNA synthesis. It
is incorporated into newly synthesized DNA during the S-phase of
the cell cycle, prior to cell division, and during the repair of
damaged DNA (13). The number of
BrdU-positive cells increases following cerebral ischemia, and this
increase in proliferation following brain damage, including during
stroke, has been suggested as an adaptive mechanism to compensate
for excessive apoptosis (14,15).
Ulinastatin is a urinary trypsin inhibitor,
originally extracted and purified from human urine, and is an
intrinsic serine-protease inhibitor (16). Urinary trypsin inhibitors have
anti-inflammatory activities, which suppress the infiltration of
neutrophils, and their release of elastase and chemical mediators
(17,18). Although the cytoprotective effects
of ulinastatin in ischemic injury have been observed in several
organs (19,20), the neuroprotective effects of
ulinastatin in ischemic cerebral injury, and their association with
levels of apoptotic molecules remain to be eluicidated.
In the present study, the effects of ulinastatin on
short-term memory, apoptotic neuronal cell death and cell
proliferation in the hippocampal regions were investigated
following transient global ischemia in gerbils. The gerbils were
provided with a step-down avoidance task, and tissue samples were
then obtained and analyzed using TUNEL staining, western blot
analysis for Bax and Bcl-2, immunohistochemistry for caspase-3 and
immunofluorescence for BrdU.
Materials and methods
Experimental animals
Adult male Mongolian gerbils (11–13-week-old) were
used in the present study. The experimental procedures were
performed in accordance with the animal care guidelines of the
National Institutes of Health (NIH) and the Korean Academy of
Medical Sciences (Seoul, Korea), and the study was approved by the
Kyung Hee University Institutional Animal Care and Use Committee
(Seoul, Korea). The gerbils were housed under controlled
temperature (20±2°C) and lighting (07:00 to 19:00 h) conditions,
with food and water made available ad libitum. The gerbils
were randomly divided into four groups (n=10 in each group): The
sham group, the ischemia group, the ischemia and 50,000 units
ulinastatin group, and the ischemia and 100,000 units ulinastatin
group.
Beginning 1 day after the induction of ischemia,
ulinastatin (Wakamoto Pharmaceutical Co., Tokyo, Japan) was
subcutaneously injected into the gerbils in the ulinastatin-treated
groups once a day for 7 consecutive days at a dose of either 50,000
or 100,000 U/kg. The gerbils in the sham and ischemia group were
administered with saline subcutaneously once a day for 7 days. BrdU
(50 mg/kg; Sigma-Aldrich, St. Louis, MO, USA) was administered
intraperitoneally to all the animals 1 h prior to ulinastatin
treatment, once each day for the same duration.
Induction of transient global
ischemia
To induce transient global ischemia in the gerbils,
a surgical procedure was performed, according to a previously
described experimental method (5).
In brief, the gerbils were anesthetized with 3% isoflurane (JW
Pharmaceutical Corporation, Seoul, Korea) in 20% O2-77%
N2. Following bilateral neck incisions, the common
carotid arteries were exposed and occluded with aneurysm clips
(Fine Science Tools, Foster City, CA, USA) for 5 min. The clips
were then removed to restore cerebral blood flow. Body temperatures
were maintained at 37±0.5°C, measured rectally, with a heating lamp
(Harvard Apparatus, Holliston, MA, USA) until the gerbils regained
consciousness. Following recovery, the animals were monitored for
an additional 2 h to prevent hypothermia. The gerbils in the sham
group underwent the same surgical procedures, with the exception of
the common carotid artery occlusion.
Step-down avoidance task
Latency in a step-down avoidance task was measured
to evaluate short-term memory, as previously described (21). The gerbils were trained in the
step-down avoidance task 7 days from the start of ulinastatin
treatment. The gerbil was placed on a 7×25 cm platform, which was
2.5 cm in height. The platform faced a 45×25 cm grid of parallel
stainless steel bars, 0.1 cm in caliber, spaced 1 cm apart. In the
training session, the animal received a 0.2 mA scramble foot shock
for 2 sec immediately upon stepping down. At 2 h after the training
session, the latency (sec) in each group was measured. The time
elapsed between when the gerbil was placed on the platform and when
the gerbil stepped down and placed all four paws on the grid was
defined as the latency. A latency >180 sec was counted as 180
sec.
Tissue preparation
The gerbils were sacrificed immediately following
the determinination of latency. The animals were anesthetized using
Zoletil 50® (10 mg/kg intraperitoneally; Vibac
Laboratories, Carros, France), transcardially perfused with 50 mM
phosphate-buffered saline (PBS), and fixed with a freshly prepared
solution consisting of 4% paraformaldehyde (Junsei Chemical Co.,
Ltd., Tokyo, Japan) in 100 mM phosphate buffer (pH 7.4; Thermo
Fisher Scientific, Waltham, MA, USA). The brains were dissected and
post-fixed in the same fixative overnight, and then transferred
into a 30% sucrose solution (Sigma-Aldrich) for cryoprotection.
Coronal sections of 40 µm thickness were made using a
freezing microtome (CM 1510-3; Leica, Nussloch, Germany).
TUNEL staining
For visualization of DNA fragmentation, a marker of
apoptotic cell death, TUNEL staining was performed using an In Situ
Cell Death Detection kit® (Roche, Mannheim, Germany),
according to the manufacturer’s instructions (5). To begin the procedure, 10 sections
were post-fixed in ethanol-acetic acid (2:1; 500 µl/section;
Sigma-Aldrich) at −20°C, and rinsed with PBS. The sections were
then incubated with proteinase K (100 µg/ml; 5
µl/section; Sigma-Aldrich), rinsed, incubated with 3%
H2O2 (500 µl/section; Sigma-Aldrich)
at room temperature, permeabilized with 0.5% Triton X-100 (500
µl/section; Sigma-Aldrich) at −20°C, rinsed again, and then
incubated in the TUNEL reaction mixture (6 µl/section) at
37°C. The sections were then rinsed and were visualized using
Converter-POD with 0.02% 3,3′-diaminobenzidine (DAB; 100
µl/section; Sigma-Aldrich) at room temperature. Mayer’s
hematoxylin (100 µl/section; DAKO, Glostrup, Denmark) was
used for counter-staining at room temperature, and the sections
were finally mounted onto gelatin-coated slides
(Marienfeld-Superior, Lauda-Königshofen, Germany). The slides were
air dried overnight at room temperature, and coverslips were
mounted using Permount® (Thermo Fisher Scientific).
Caspase-3 immunohistochemistry
For visualization of the expression of caspase-3,
caspase-3 immunohistochemistry was performed, according to the
method described previously (5). A
total of 10 sections were removed from each brain and incubated
overnight at 4°C with mouse polyclonal anti-caspase-3 antibody
(1:500; cat. no. sc-7272; Santa Cruz Biotechnology, Inc., Santa
Cruz, CA, USA) and washed three times with PBS. The sections were
then incubated for another 1 h at room temperature with
biotinylated horse anti-mouse secondary antibody (1:100; cat. no.
BA2000; Vector Laboratories, Burlingame, CA, USA), and washed three
times with PBS. The bound secondary antibody was then amplified
using a Vector Elite ABC kit® (Vector Laboratories). The
antibody-biotin-avidin-peroxidase complexes were visualized using
0.02% DAB, and the sections were finally mounted onto
gelatin-coated slides. The slides were air dried overnight at room
temperature, and coverslips were mounted using Permount®
(Thermo Fisher Scientific).
BrdU immunofluorescence
For the detection of newly-generated cells in the
dentate gyrus, BrdU-specific immunofluorescence was performed, as
previously described (5). The
sections were first permeabilized by incubation with 0.5% Triton
X-100 in PBS (1 ml/well) for 20 min at room temperature, and were
then pretreated with 50% formamide-2X standard saline citrate (1
ml/well) at 65°C for 2 h, denatured in 2N HCl (2 ml/well) at 37°C
for 30 min, and rinsed twice in 100 mM sodium borate (pH 8.5; 1
ml/well). For double labeling of BrdU and neuronal nuclear antigen
(NeuN), the sections were incubated overnight with rat anti-BrdU
antibody (1:300, Abcam, Biomeda, CA, USA) and mouse anti-NeuN
antibody (1:500, Chemicon International, Temecula, CA, USA) at 4°C
overnight following DNA denaturation. The sections were then
incubated for 2 h with cy3-conjugated anti-rat secondary antibody
for BrdU (1:200, Jackson ImmunoResearch Laboratories, West Grove,
PA, USA) and fluorescein isothiocyanate-conjugated anti-mouse
secondary antibody for NeuN (1:200; Jackson ImmunoResearch
Laboratories). The sections were then mounted on gelatin-coated
glass slides, and the coverslips were mounted using fluorescent
mounting medium (DakoCytomation, Carpinteria, CA, USA). Images of
the fluorescent staining were captured using an epifluorescent
microscope (Nikon Eclipse 50i; Nikon Inc., Melville, NY, USA).
Western blot analysis
Western blot analysis was performed to detect the
protein expression levels of Bax and Bcl-2 in the hippocampus, as
previously described (10). The
hippocampal tissues were collected and immediately frozen at −70°C.
The hippocampal tissue samples were homogenized on ice, and lysed
in lysis buffer containing 50 mM HEPES (pH 7.5), 150 mM NaCl, 10%
glycerol, 1% Triton X-100, 1 mM phenylmethylsulfonyl fluoride, 1 mM
ethylene glycol tetraacetic acid, 1.5 mM
MgCl2·6H2O, 1 mM sodium orthovanadate, and
100 mM sodium fluoride (Sigma-Aldrich). The protein content was
measured using a DC Universal Protein Assay kit (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Equal quantities of total
protein (30 µg) from each group were separated on 12%
SDS-polyacrylamide gels and transferred onto a nitrocellulose
membrane (GE Healthcare Life Sciences, Piscataway, NJ, USA). Mouse
actin antibody (1:500; Santa Cruz Biotechnology, Inc.), mouse Bax
antibody (cat. no. SC7480; 1:1000; Santa Cruz Biotechnology, Inc.),
and mouse Bcl-2 antibody (cat. no. SC7382; 1:1000; Santa Cruz
Biotechnology, Inc.) were used as the primary antibodies and
incubated overnight at 4°C. Horseradish peroxidase-conjugated
anti-mouse antibody for Bax and Bcl-2 (cat. no. RPN4201; 1:2,000;
Amersham Pharmacia Biotechnology GmbH, Freiburg, Germany) were used
as the secondary antibodies and incubated for 1 h at room
temperature. Experiments were performed under normal laboratory
conditions and at room temperature, with the exception of the
treatment of the transfer membranes. The transfer membranes were
treated at 4°C with a cold pack and pre-chilled transfer buffer (25
mM Tris, 192 mM glycine, pH 8.3). Band detection was performed
using an enhanced chemiluminescence (ECL) detection kit (cat. no.
SC2048; Western Blotting Luminol Reagent; Santa Cruz Biotechnology,
Inc.), at a wavelength of 428 nm.
Data analysis
The numbers of TUNEL-positive and caspase-3-positive
cells in the hippocampal CA1 region, and the number of
BrdU-positive cells in the hippocampal dentate gyrus were counted
using an Image-Pro® Plus computer-assisted image
analysis system (Media Cyberbetics Inc., Silver Spring, MD, USA)
attached to a light microscope (BX-51; Olympus Corporation, Tokyo,
Japan) under x10 magnification. To compare the relative expression
of proteins, the band intensities were quantified densitometrically
using Molecular Analyst™ version 1.4.1 (Bio-Rad Laboratories,
Inc.).
Statistical analysis was performed using one-way
analysis of variance followed by Duncanm’s post-hoc test.
Statistical analyses were conducted using SPSS version 21.0 (IBM,
Armonk, NY, USA). The results are expressed as the mean ± standard
error of the mean. P<0.05 was considered to indicate a
statistically significant difference.
Results
Effect of ulinastatin on latency in the
step-down avoidance task
The latencies observed in the step-down avoidance
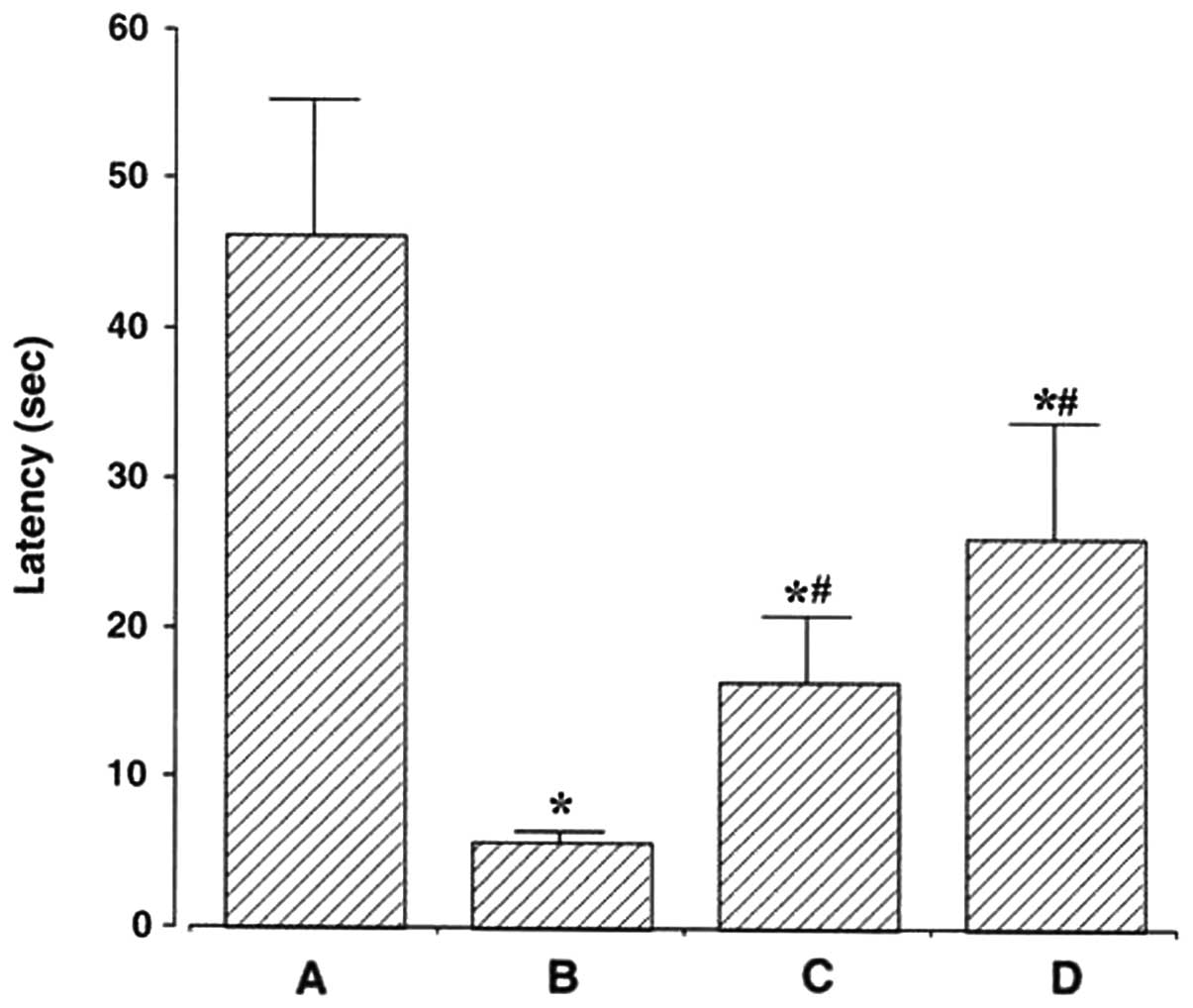
task are presented in Fig. 1.
Latency was 46.38±9.04 sec in the sham group, 5.67±0.82 sec in the
ischemia group, 16.60±4.27 sec in the ischemia and 50,000 units
ulinastatin group, and 26.00±7.78 sec in the ischemia and 100,000
units ulinastatin group. These results demonstrated that ischemic
insult caused deterioration of short-term memory, and that
ulinastatin alleviated the ischemia-induced impairment in
short-term memory.
Effect of ulinastatin on the numbers of
TUNEL-positive and caspase-3-positive cells in the hippocampal CA1
region
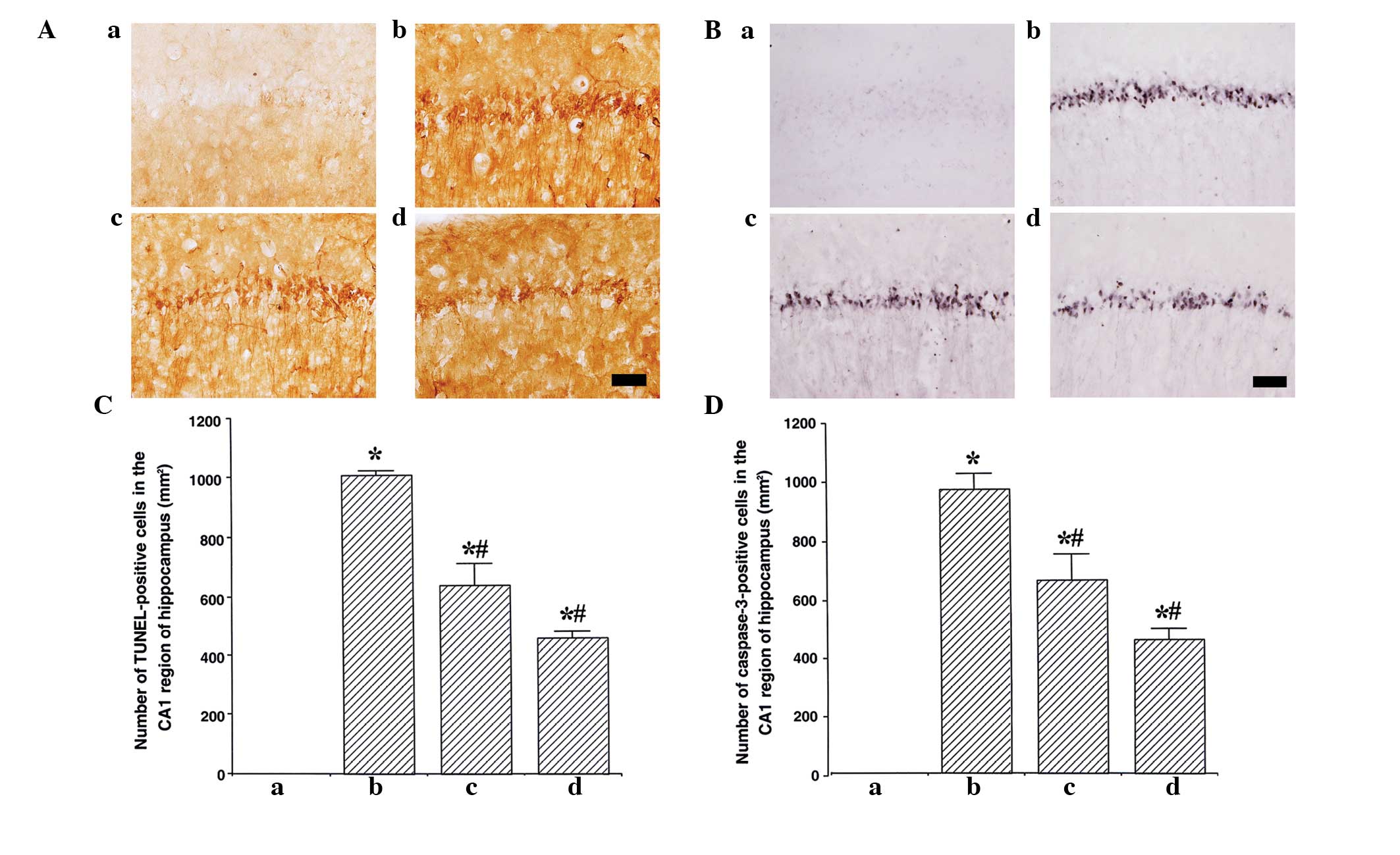
Photomicrographs of TUNEL-positive cells in the
hippocampal CA1 region are presented in Fig. 2A. The number of TUNEL-positive
cells was 0.00±0.00/mm2 in the sham group,
1,007.19±41.32/mm2 in the ischemia group,
635.66±73.51/mm2 in the ischemia and 50,000 units
ulinastatin group, 456.79±21.38/mm2 in the ischemia and
100,000 units ulinastatin group Fig.
2B). These results demonstrated that ischemic insult enhanced
DNA fragmentation in the CA1 region, and that ulinastatin treatment
suppressed the ischemia-induced DNA fragmentation.
Photomicrographs of caspase-3-positive cells in the
hippocampal CA1 region are presented in Fig. 2C. The number of caspase-3-positive
cells was 0.78±0.01/mm2 in the sham group,
964.80±52.31/mm2 in the ischemia group,
654.45±87.82/mm2 in the ischemia and 50,000 units
ulinastatin group, and 447.74±38.42/mm2 in the
ischemia-and 100,000 units ulinastatin group (Fig. 2D). These results demonstrate that
ischemic insult enhanced the expression of caspase-3 in the CA1
region, and that ulinastatin treatment suppressed the
ischemia-induced expression of caspase-3.
Effect of ulinastatin treatment on the
protein expression of Bcl-2 and Bax in the hippocampus
To verify the effects of ulinastatin on the
expression of the apoptosis-associated proteins, the relative
expression levels of Bax and Bcl-2 in the hippocampu were
determined. When the level of Bax (24 kDa) in the sham group was
set at 1.00, the level of Bax was 4.22±0.74 in the ischemia group,
3.24±0.55 in the ischemia and 50,000 units ulinastatin group, and
2.34±0.57 in the ischemia and 100,000 units ulinastatin group.
These results revealed that ischemic insult enhanced the expression
of Bax in the hippocampus, and that ulinastatin suppressed the
ischemia-induced expression of Bax (Fig. 3B).
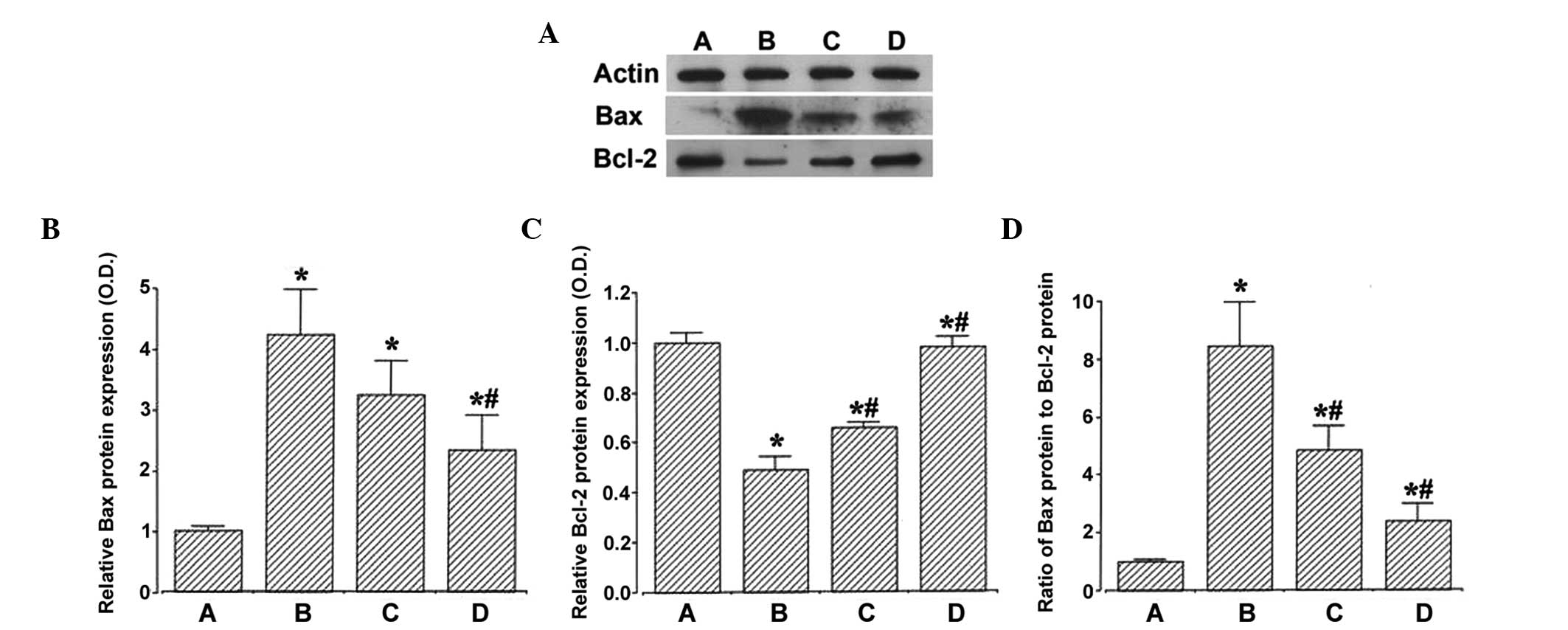 | Figure 3Effects of ulinastatin on the
expression levels of Bax and Bcl-2 in the hippocampus. (A)
Expression levels of Bax and Bcl-2, detected using western blot
analysis. (B) Relative expression of Bax. (C) Relative expression
of Bcl-2. (D) Ratio of Bax to Bcl-2. (A) Sham-operation, (B)
ischemia-induction, (C) ischemia and 50,000 units ulinastatin, (D)
ischemia and 100,000 unitsulinastatin. Data are presented as the
mean ± standard error of the mean. *P<0.05, compared with the
sham-operation group; #P<0.05, compared with the
ischemia-induction group. Blc-2, B cell lymphoma 2; Bax,
Bcl-2-associated X protein; OD, optical density. |
When the level of Bcl-2 (26–29 kDa) in the sham
group was set at 1.00, the level of Bcl-2 was 0.49±0.05 in the
ischemia group, 0.66±0.03 in the ischemia and 50,000 units
ulinastatin group, and 0.98±0.01 in the ischemia and 100,000 units
ulinastatin group. These results indicated that ischemic insult
suppressed the expression of Bcl-2 in the hippocampus, and that
ulinastatin enhanced the exprssion of Bcl-2 in the ischemic gerbils
(Fig. 3C).
Ratio of Bax to Bcl-2
When the ratio of Bax to Bcl-2 in the sham group was
set at 1.00, the ratio of Bax to Bcl-2 was 8.46±1.50 in the
ischemia group, 4.85±0.83 in the ischemia and 50,000 units
ulinastatin group, and 2.34±0.57 in the ischemia and 100,000 units
ulinastatin group. These results demonstrated that ischemic insult
enhanced the ratio of Bax to Bcl-2 in the hippocampus, and that
ulinastatin suppressed this ischemia-induced increase in the Bax to
Bcl-2 ratio (Fig. 3C).
Effect of ulinastatin on the number of
BrdU-positive cells in the hippocampal dentate gyrus
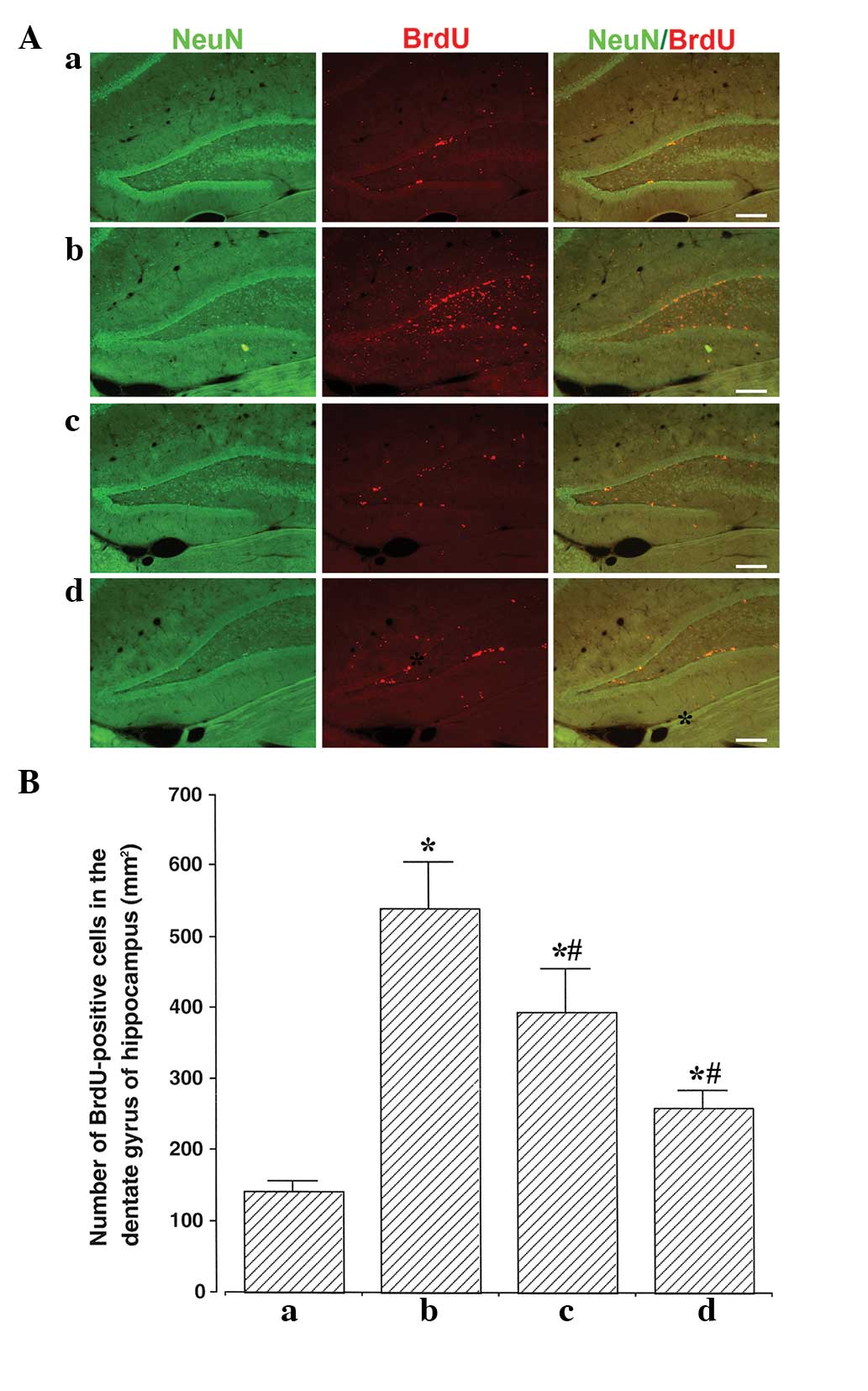
Photomicrographs of BrdU-positive cells in the
hippocampal dentate gyrus are presented in Fig. 4A. The number of BrdU-positive cells
was 141.75±14.53/mm2 in the sham group,
540.00±65.56/mm2 in the ischemia group,
393.58±61.06/mm2 in the ischemia and 50,000 units
ulinastatin group, and 260.36±22.84/mm2 in the ischemia
and 100,000 units ulinastatin group (Fig. 4B). These results revealed that
ischemic insult enhanced cell proliferation in the hippocampal
dentate gyrus, and that ulinastatin suppressed this
ischemia-induced cell proliferation.
Discussion
The deletion of the CA1 pyramidal neurons is
associated with severe impairment of hippocampal-dependent brain
functions, including spatial learning ability and short-term memory
(4,22,23).
Following transient global ischemia in gerbils, short-term memory
in the step-down avoidance task is impaired (5). In the present study, latency in the
step-down avoidance task was shortened following the induction of
transient global ischemia, however, it was improved following
treatment with ulinastatin. The results of these experiments
indicated that ulinastatin alleviated the short-term memory
impairment induced by transient global ischemia in gerbils.
Pyramidal neurons in the hippocampal CA1 region are
particularly sensitive to transient global ischemia (24). The upregulation and activation of
caspase-3 in the early stages of apoptosis is a hallmark of
ischemia (1,6). Increased numbers of TUNEL-positive
and caspase-3-positive cells in the hippocampal CA1 region is
observed following transient global ischemia in gerbils (5). In the present study, the numbers of
TUNEL-positive and caspase-3-positive cells in the hippocampal CA1
region increased following ischemic insult, indicating that
cerebral ischemia induced apoptotic cell death in the hippocampal
CA1 region. Treatment with ulinastatin suppressed the
ischemia-induced increase in DNA fragmentation and expression of
caspase-3 in the hippocampal CA1 region, indicating that
ulinastatin suppressed cerebral ischemia-induced apoptotic cell
death in this region.
The increased expression of Bcl-2 inhibits apoptosis
(25), while the overexpression of
Bax promotes apoptosis (26). It
has been suggested that the balance between Bax and Bcl-2 is
imporant in the determination of cell death or survival (27). The induction of intracranial
hemorrhage suppresses the expression of Bcl-2 and increases the
expression of Bax, resulting in an increase of the Bax to Bcl-2
ratio (10). In the present study,
the expression of Bax increased, the expression of Bcl-2 decreased,
and the ratio of Bax to Bcl-2 increased following ischemic insult.
These results indicated that cerebral ischemia accelerated
apoptotic cell death by inhibiting anti-apoptotic molecules and
activating pro-apoptotic molecules in the hippocampus. By contrast,
treatment with ulinastatin suppressed the expression of Bax and
enhanced the expression of Bcl-2 in the hippocampus, resulting in a
decrease in the Bax to Bcl-2 ratio. These results indicated that
ulinastatin enhanced anti-apoptotic molecules and suppressed
pro-apoptotic molecules in the hippocampus.
Increasing neurogenesis in the hippocampal dentate
gyrus improves learning ability and memory function (8,11).
However, pathological conditions, including brain inflammation, are
associated with an increase in neurogenesis without memory
enhancement (21,28). In the present study, cell
proliferation in the hippocampal dentate gyrus was significantly
increased following ischemic attack, indicating that ischemia
induced excessive apoptotic neuronal cell death in the hippocampus.
Treatment with ulinastatin alleviated the ischemia-induced cell
proliferation in the hippocampal dentate gyrus, suggesting that
ulinastatin exerted a suppressive effect on the ischemia-induced
apoptosis in the hippocampus.
Urinary trypsin inhibitors have been used to treat
pancreatitis, septic shock and hemorrhagic shock (29–31).
Chen et al (32) reported
that the protective effects of ulinastatin may be associated with
the upregulation of Bcl-2, an inhibitor of cell apoptosis, in a
hemorrhagic shock animal model. Sung et al (33) reported that ulinastatin exerts
analgesic and anti-inflammatory effects by suppressing
cyclooxygenase-2 and inducible nitric oxide synthase through the
downregulation of nuclear factor κ-B activity in BV2 mouse
microglial cells.
In the present study, ulinastatin inhibited
ischemia-induced apoptosis in the hippocampus by suppressing
apoptosis-associated molecules, thus ameliorating ischemia-induced
short-term memory impairment. These results suggested the potential
use of ulinastatin as a therapeutic agent for patients with
cerebral stroke.
Acknowledgments
This study was supported by the Program of Kyung Hee
University for the Young Researcher of Medical Science in 2007 (no.
KHU-20071478).
References
|
1
|
Benchoua A, Guégan C, Couriaud C, Hosseini
H, Sampaïo N, Morin D and Onténiente B: Specific caspase pathways
are activated in the two stages of cerebral infarction. J Neurosci.
21:7127–7134. 2001.PubMed/NCBI
|
|
2
|
Dirnagl U, Iadecola C and Moskowitz MA:
Pathobiology of ischaemic stroke: an integrated view. Trends
Neurosci. 22:391–397. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mattson MP, Duan W, Pedersen WA and
Culmsee C: Neurodegenerative disorders and ischemic brain diseases.
Apoptosis. 6:69–81. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hartman RE, Lee JM, Zipfel GJ and Wozniak
DF: Characterizing learning deficits and hippocampal neuron loss
following transient global cerebral ischemia in rats. Brain Res.
1043:48–56. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ko IG, Shin MS, Kim BK, Kim SE, Sung YH,
Kim TS, Shin MC, Cho HJ, Kim SC, Kim SH, Kim KH, Shin DH and Kim
CJ: Tadalafil improves short-term memory by suppressing
ischemia-induced apoptosis of hippocampal neuronal cells in
gerbils. Pharmacol Biochem Behav. 91:629–635. 2009. View Article : Google Scholar
|
|
6
|
Sim YJ, Kim SS, Kim JY, Shin MS and Kim
CJ: Treadmill exercise improves short-term memory by suppressing
ischemia-induced apoptosis of neuronal cells in gerbils. Neurosci
Lett. 372:256–261. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Korsmeyer SJ: BCL-2 gene family and the
regulation of programmed cell death. Cancer Res. 59:S1693–S1700.
1999.
|
|
8
|
Kim SE, Ko IG, Kim BK, Shin MS, Cho S, Kim
CJ, Kim SH, Baek SS, Lee EK and Jee YS: Treadmill exercise prevents
aging-induced failure of memory through an increase in neurogenesis
and suppression of apoptosis in rat hippocampus. Exp Gerontol.
45:357–365. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Oltvai ZN, Milliman CL and Korsmeyer SJ:
Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that
accelerates programmed cell death. Cell. 74:609–619. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hwang L, Choi IY, Kim SE, Ko IG, Shin MS,
Kim CJ, Kim SH, Jin JJ, Chung JY and Yi JW: Dexmedetomidine
ameliorates intracerebral hemorrhage-induced memory impairment by
inhibiting apoptosis and enhancing brain-derived neurotrophic
factor expression in the rat hippocampus. Int J Mol Med.
31:1047–1056. 2013.PubMed/NCBI
|
|
11
|
Gould E, Tanapat P, Hastings NB and Shors
TJ: Neurogenesis in adulthood: a possible role in learning. Trends
Cogn Sci. 3:186–192. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schmidt W and Reymann KG: Proliferating
cells differentiate into neurons in the hippocampal CA1 region of
gerbils after global cerebral ischemia. Neurosci Lett. 334:153–156.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kempermann G, Kuhn HG and Gage FH: Genetic
influence on neurogenesis in the dentate gyrus of adult mice. Proc
Natl Acad Sci USA. 94:10409–10414. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu J, Solway K, Messing RO and Sharp FR:
Increased neurogenesis in the dentate gyrus after transient global
ischemia in gerbils. J Neurosci. 18:7768–7778. 1998.PubMed/NCBI
|
|
15
|
Takasawa K, Kitagawa K, Yagita Y, Sasaki
T, Tanaka S, Matsushita K, Ohstuki T, Miyata T, Okano H, Hori M and
Matsumoto M: Increased proliferation of neural progenitor cells but
reduced survival of newborn cells in the contralateral hippocampus
after focal cerebral ischemia in rats. J Cereb Blood Flow Metab.
22:299–307. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tanaka Y, Maehara S, Sumi H, Toki N,
Moriyama S and Sasaki K: Purification and partial characterization
of two forms of urinary trypsin inhibitor. Biochim Biophys Acta.
705:192–199. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nakatani K, Takeshita S, Tsujimoto H,
Kawamura Y and Sekine I: Inhibitory effect of serine protease
inhibitors on neutrophil-mediated endothelial cell injury. J Leukoc
Biol. 69:241–247. 2001.PubMed/NCBI
|
|
18
|
Yamaguchi Y, Ohshiro H, Nagao Y, Odawara
K, Okabe K, Hidaka H, Ishihara K, Uchino S, Furuhashi T, Yamada S,
Mori K and Ogawa M: Urinary trypsin inhibitor reduces C-X-C
chemokine production in rat liver ischemia/reperfusion. J Surg Res.
94:107–115. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cao ZL, Okazaki Y, Naito K, Ueno T,
Natsuaki M and Itoh T: Ulinastatin attenuates reperfusion injury in
the isolated blood-perfused rabbit heart. Ann Thorac Surg.
69:1121–1126. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yano T, Anraku S, Nakayama R and Ushijima
K: Neuroprotective effect of urinary trypsin inhibitor against
focal cerebral ischemia-reperfusion injury in rats. Anesthesiology.
98:465–473. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim SE, Ko IG, Park CY, Shin MS, Kim CJ
and Jee YS: Treadmill and wheel exercise alleviate
lipopolysaccharide-induced short-term memory impairment by
enhancing neuronal maturation in rats. Mol Med Rep. 7:31–36.
2013.
|
|
22
|
Bendel O, Bueters T, von Euler M, Ove
Ogren S, Sandin J and von Euler G: Reappearance of hippocampal CA1
neurons after ischemia is associated with recovery of learning and
memory. J Cereb Blood Flow Metab. 25:1586–1595. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
von Euler M, Bendel O, Bueters T, Sandin J
and von Euler G: Profound but transient deficits in learning and
memory after global ischemia using a novel water maze test. Behav
Brain Res. 166:204–210. 2006. View Article : Google Scholar
|
|
24
|
Schmidt-Kastner R and Freund TF: Selective
vulnerability of the hippocampus in brain ischemia. Neuroscience.
40:599–636. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kitagawa K, Matsumoto M, Tsujimoto Y,
Ohtsuki T, Kuwabara K, Matsushita K, Yang G, Tanabe H, Martinou JC,
Hori M and Yanagihara T: Amelioration of hippocampal neuronal
damage after global ischemia by neuronal overexpression of BCL-2 in
transgenic mice. Stroke. 29:2616–2621. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Merry DE and Korsmeyer SJ: Bcl-2 gene
family in the nervous system. Annu Rev Neurosci. 20:245–267. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Upadhyay D, Panduri V, Ghio A and Kamp DW:
Particulate matter induces alveolar epithelial cell DNA damage and
apoptosis: role of free radicals and the mitochondria. Am J Respir
Cell Mol Biol. 29:180–187. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ekdahl CT, Kokaia Z and Lindvall O: Brain
inflammation and adult neurogenesis: the dual role of microglia.
Neuroscience. 158:1021–1029. 2009. View Article : Google Scholar
|
|
29
|
Maciejewski R, Burdan F, Burski K, Madej
B, Ziemiakowicz R, Dabrowski A and Wallner G: Selected biochemical
parameters and ultrastructural picture of pancreas due to
Ulinastatin treatment of experimental acute pancreatitis. Exp
Toxicol Pathol. 56:305–311. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Inoue K, Takano H, Shimada A, Yanagisawa
R, Sakurai M, Yoshino S, Sato H and Yoshikawa T: Urinary trypsin
inhibitor protects against systemic inflammation induced by
lipopolysaccharide. Mol Pharmacol. 67:673–680. 2005. View Article : Google Scholar
|
|
31
|
Masuda T, Sato K, Noda C, Ikeda KM,
Matsunaga A, Ogura MN, Shimizu K, Nagasawa H, Matsuyama N and Izumi
T: Protective effect of urinary trypsin inhibitor on myocardial
mitochondria during hemorrhagic shock and reperfusion. Crit Care
Med. 31:1987–1992. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen CC, Liu ZM, Wang HH, He W, Wang Y and
Wu WD: Effect of ulinastatin on renal ischemia-reperfusion injury
in rats. Acta Pharmacol Sin. 25:1334–1340. 2004.PubMed/NCBI
|
|
33
|
Sung YH, Shin MS, Ko IG, Kim SE, Kim CJ,
Ahn HJ, Yoon HW and Lee BJ: Ulinastatin suppresses
lipopolysaccharide-induced prostaglandin E2 synthesis and nitric
oxide production through the downregulation of nuclear factor-κB in
BV2 mouse microglial cells. Int J Mol Med. 13:1030–1036. 2013.
|