Introduction
Sepsis is a severe systemic inflammatory response
associated with infectious disease, which remains a leading cause
of mortality in clinical settings (1). Sepsis usually occurs following a
severe infection, and may be triggered by numerous pathogenic
microorganisms that release various toxins into the bloodstream and
tissues (2). It has been suggested
that sepsis is caused by an exaggerated inflammatory response,
which may develop into septic shock and multiple organ dysfunction
syndrome (3). During a severe
infection, intestinal barrier dysfunction may lead to ischemia and
hypoxia, both of which have an important role in the development of
sepsis (4). In addition, emerging
evidence suggests that the intestinal tract is not only a target of
the inflammatory response, but may also be the starting point for
multiple organ dysfunction (5).
Bacterial endotoxins induce intestinal barrier dysfunction by
regulating the expression levels of various mediators (6).
Toll-like receptor 4 (TLR4) is a member of the
sensor protein family, which has been shown to have an important
role in the pathogenesis of inflammatory diseases, including
sepsis-associated mortality (7).
The activation of TLR4 during sepsis may be associated with the
stimulation of the lipopolysaccharide (LPS) endotoxin (8). The development of a TLR4 inhibitor
that prevents endotoxin and TLR4 interaction may provide a
promising therapeutic target for the treatment of sepsis (9).
Rhein (Fig. 1) is a
lipophilic anthraquinone isolated from Rheum rhabarbarum
(rhubarb) that exhibits a protective effect on platelet-activated
and factor-mediated intestinal damage in rat colon (10). The mechanism underlying the
protective activity of rhein may be associated with the generation
of nitric oxide, ion secretions and chemotaxis, and with the
apoptosis of human-derived Caco-2 colorectal cancer cells (11). Notably, rhein has been shown to
successfully repair damaged intestinal tight junctions and protect
the intestinal barrier, by enhancing the expression of zona
occludens-1 and occludin (12).
Furthermore, rhein significantly increased the expression of
interleukin (IL)-10, and decreased the expression of tumor necrosis
factor (TNF)-α (13). However, the
mechanism underlying the protective effects of rhein on LPS-induced
intestinal barrier dysfunction remains to be elucidated.
The aim of the present study was to investigate the
innate resistance of rhein to LPS-induced intestinal toxicity
during sepsis in a murine model. Furthermore, the inhibition of
rhein in the TLR4 NF-κB signaling pathway was investigated.
Materials and methods
Chemicals and reagents
LPS (L-2880:B5) was purchased from Sigma-Aldrich
(St. Louis, MO, USA). Rhein (purity >98%) was purchased from the
National Institute for the Control of Pharmaceutical and Biological
Products (Beijing, China). ELISA kits used to detect the
inflammatory cytokines IL-1β, IL-6, IL-8, IL-10, and TNF-α were
purchased from the Nanjing Jiancheng Bioengineering Institute
(Nanjing, China). The antibodies directed against IL-1β, IL-6,
IL-8, IL-10, TNF-α, TLR-4, and β-actin, as well as the
3,3′-diaminobenzidine (DAB) reagent were purchased from Wuhan
Boster Biological Technology, Ltd. (Wuhan, China). The TLR4
inhibitor TAK-242 was purchased from EMD Millipore (Billerica, MA,
USA). SDS-PAGE and polyvinylidene fluoride (PVDF) membranes were
obtained from Bio-Rad Laboratories, Inc. (Hercules, CA, USA). The
Enhanced Chemiluminescence (ECL) Advanced Western Blotting
Detection kit was purchased from GE Healthcare Life Sciences
(Chalfont, UK). All other reagents used in the present study were
purchased from Sigma-Aldrich. Anti-NF-κB p65 (phospho S536; cat.no.
ab28856) and total anti-NF-κB p65 antibody (cat. no. ab16502) were
purchased from Abcam Trading (Shanghai) Company Ltd. (Shanghai,
China).
Animal experiments
Four-week-old male BALB/c mice, weighing 18–22 g,
were provided by Shanghai Laboratory Animals Center Co., Ltd.
(Shanghai, China). The mice were maintained in controlled
conditions at 25±1°C, a relative humidity of 45% with 12 h
light/dark cycles, and were given ad libitum access to food
and water. The mice were randomly assigned to five experimental
groups, consisting of a normal control group, an LPS-stimulated
model group, a rhein group, a TLR4 inhibitor group and a rhein +
TLR4 inhibitor TAK-242 group (n=10, each group). The mice in each
group were orally administered 100 mg/kg rhein (14), and 0.3 mg/kg TAK-242 (15) for 1 h prior to being injected
intraperitoneally with 20 mg/kg LPS (16) for a further 18 h. The mice were
subsequently anesthetized with an intraperitoneal injection of 50
mg/kg pentobarbital (Merck KGaA, Darmstadt, Germany), prior to
having their blood samples taken from the retinal vein. The mice
were sacrificed under anesthesia with ketamine at 65 mg/kg and
xylazine at 6.5 mg/kg−1, i.p. (Jiancheng Bioengineering
Institute) and 1 cm from the rectum and 1 cm from the cecum were
harvested in order to evaluate the changes in intestinal injury.
The length of the colons were measured and recorded. The present
study was approved by the Institutional Animal Care and Use
Committee of Zhengzhou People's Hospital, (Zhengzhou, China).
Intestinal mucosal pathology
The extent of intestinal mucosal injury was graded
following inspection under a microscope (Olympus IX71; Olympus,
Tokyo, Japan), according to the standard classification described
by Chiu et al (17).
Intestinal injury was graded as follows: 0, normal mucosal villi;
1, the gap in the epithelium increased, and capillary congestion at
the tip of villi was observed; 2, the gap in the epithelium was
expanded with moderate separation of the epithelium and lamina
propria; 3, severe separation of the epithelium and lamina propria
with damage to the villus tip; 4, villi were damaged and the
capillaries exposed in the lamina propria, an increase in the
cellular components of the lamina propria was also observed; and 5,
the intrinsic layer was damaged with incomplete bleeding and
ulcers.
Immunohistochemistry
Sections of 4 μm formalin-fixed,
paraffin-embedded colon tissue was cut for immunohistochemistry.
Following deparaffinization in xylene, the tissue sections were
hydrated using graded ethanol (100, 95 and 70%). The sections were
subsequently washed three times in deionized H2O for 1
min at room temperature, prior to being incubated in 3% hydrogen
peroxide for 10 min, in order to block the endogenous peroxidase
activity. Each section was then sequentially washed with
phosphate-buffered saline (PBS) for 5 min prior to being blocked
with 5% horse serum (Gibco-Brl, Gaithersburg, MD, USA). The
sections were then incubated overnight at 4°C with 1:100 dilution
of primary rabbit anti-TLR4 antibody (cat. no. sc-16240; Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA). Following 3–5 min washes
with PBS, the sections were treated with biotin-conjugated goat
anti-rabbit secondary antibody for 2 h at 37°C. The sections were
then treated with an avidin biotin enzyme reagent (Wuhan Boster
Biological Technology, Ltd.) for 30 min, prior to being washed
three times with PBS. The sections were subsequently stained with
DAB and DAB enhancer, prior to staining with hematoxylin for 5 sec.
Dehydration was performed using ethanol and xylene. Finally, the
sections were covered with a glass coverslip and observed under an
Olympus IX71 microscope.
Western blot analysis
The colon tissue samples were harvested from the
BALB/c mice of each group, prior to being frozen and stored at
−80°C until further use. The colon tissue samples were homogenized
in ice-cold radioimmunoprecipitation lysing buffer (Sigma-Aldrich)
and were centrifuged at 12,000 × g for 15 min at 4°C. The
supernatant was subsequently collected in order to examine protein
expression levels using western blot analysis. Protein
concentrations were determined using a bicinchoninic acid protein
assay reagent kit (Pierce Biotechnology, Inc., Rockford, IL USA).
Equal amounts of supernatant protein (40 μg) were separated
by 10% SDS-PAGE, and transferred to PVDF membranes. The membranes
were blocked with 5% skimmed dry milk in tris-buffered saline with
0.05% Tween® 20 for 1 h, and were then co-incubated
overnight at 4°C with the following primary antibodies IL-1β
(1:1,000; cat. no. sc-7884), IL-6 (1:1,000; cat. no. sc-1265), IL-8
(1:1,000; cat. no. sc-8427), IL-10 (1:1,000; cat. no. sc-1783),
TNF-α (1:500; cat. no. sc-65440), TLR-4 (1:500; cat. no. BA1717),
Phospho-NF-kB p65 (1:500), total NF-kB p65 (1:500) and β-actin
(1:400; cat. no. BM0005). The membranes were subsequently washed
with PBS supplemented with 0.05% Tween® 20 and incubated
for 2 h at room temperature with horseradish peroxidase-bound
secondary antibodies (1:1,000; Santa Cruz Biotechnology, Inc.).
β-actin was used as a loading control. Finally, the proteins were
visualized using ECL reagents, and the intensity of the
immunoreactive bands was quantified and normalized to the
respective β-actin content using Image Pro Plus (IPP) software
(Media Cybernetics, Inc., Rockville, MD, USA).
ELISA for inflammatory cytokines
Following sacrifice, blood samples of the mice were
collected in heparinized tubes and centrifuged at 10,000 × g for 10
min in order to obtain the plasma. The colon tissue samples of the
mice in each group were subsequently harvested, weighed, and
homogenized in iced saline. The samples were then centrifuged at
10,000 × g for 10 min in order to obtain the supernatant. The
levels of inflammatory cytokines IL-1β, IL-6, IL-8, IL-10, and
TNF-α present in the supernatants were assayed by ELISA, according
to the manufacturer's instructions. The optical densities of the
samples were detected using a microplate reader (ELx800; BioTek
Instruments, Inc., Winooski, VT, USA) at a wavelength of 450
nm.
Statistical analysis
The data from at least three experiments were
presented as the mean ± standard deviation. The statistical
significance of the differences between the experimental groups was
analyzed using SPSS 16.0 (SPSS Inc., Chicago, IL, USA). One-way
analysis of variance and post hoc Tukey tests were used to analyze
the data. For all experiments, P<0.05 was considered to indicate
a statistically significant difference.
Results
Rhein extends the colon length and
attenuates LPS-induced damage
Following stimulation with 20 mg/kg LPS, the length
of the murine colon was markedly shortened, as compared with that
of the normal mice (P<0.01). However, following treatment with
100 mg/kg rhein, the length of the colon was significantly
increased. Notably, treatment with either 0.3 mg/kg TLR4 inhibitor
TAK-242 alone or together with rhein, also increased the length of
the damaged colon (P<0.05 and P<0.01, respectively). However,
no statistically significant difference was observed between the
mice treated with rhein alone, or the mice co-treated with TAK-242
(Table I).
 | Table ILength of murine colon and level of
intestinal mucosal injury in the various mouse groups. |
Table I
Length of murine colon and level of
intestinal mucosal injury in the various mouse groups.
| Group | n | Length of colon
(cm) | Score of colon
injury |
|---|
| Normal | 10 | 10.25±0.78 | 0 |
| Model | 7 | 8.15±1.14a,b | 4.48±0.43a,b |
| Rhein (100
mg/kg) | 10 | 9.53±0.62c | 2.40±0.77d |
| Rhein (100 mg/kg) +
TAK-242 (0.3 mg/kg) | 9 | 9.64±0.89c | 1.97±0.51d |
| TAK-242 (0.3
mg/kg) | 9 | 9.43±0.93d | 2.82±0.56d |
LPS, the main endotoxin present during sepsis in the
case of severe systemic inflammatory syndrome, has been shown to
induce colon damage. As demonstrated in Table I, significant damage was observed
in the LPS-induced mice, as compared with the normal mice. However,
this damage was notably attenuated following treatment with rhein
in the presence or absence of TAK-242 (P<0.01). These results
support the hypothesis that rhein is able to attenuate LPS-induced
colon injury.
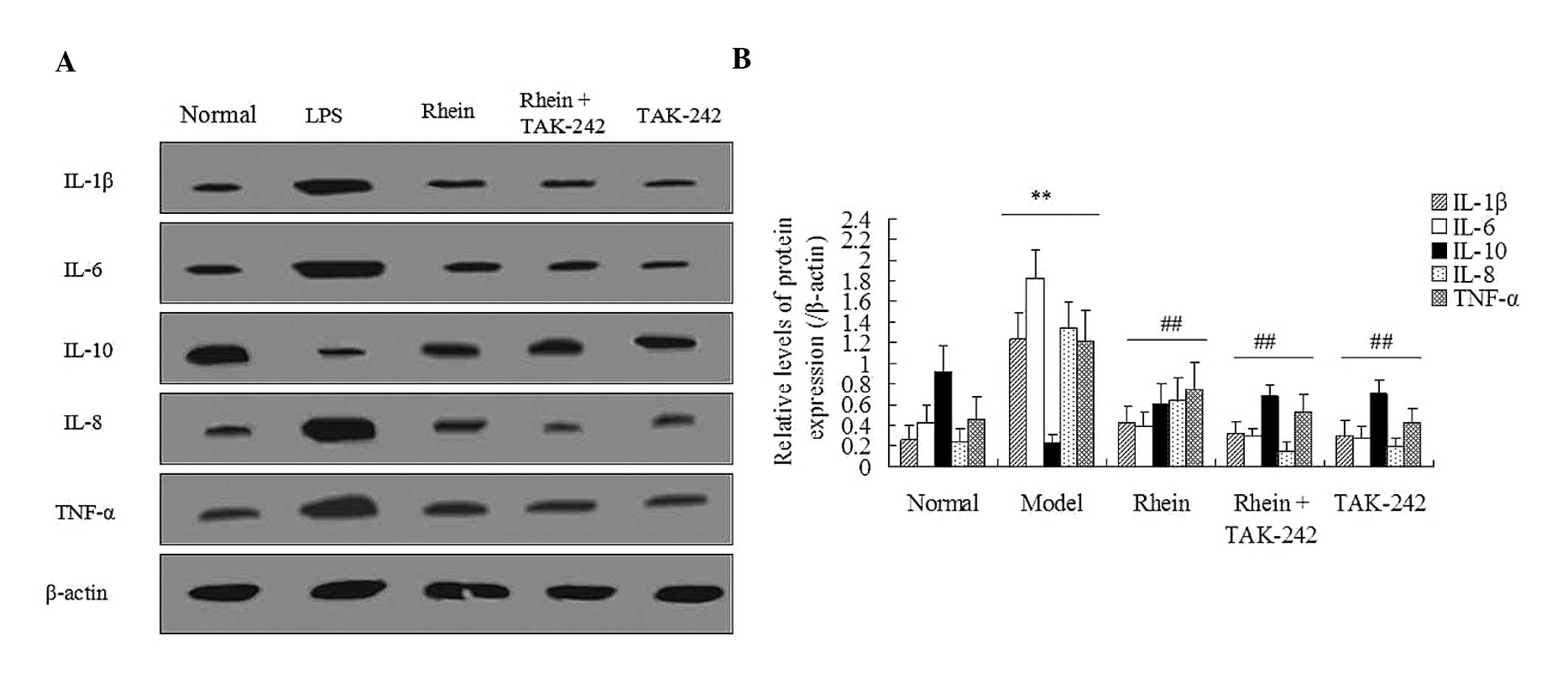
Rhein attenuates the levels of
LPS-induced inflammatory cytokines
An LPS-induced inflammatory response was associated
with damage to the mouse colon. ELISA was performed in order to
detect the expression levels of the following inflammatory
cytokines: IL-1β, IL-6, IL-8, IL-10, and TNF-α, in both the plasma
and colon tissue samples. As shown in Table II, exposure to LPS markedly
increased the concentration of the inflammatory cytokines IL-1β,
IL-6, IL-8, and TNF-α, whereas it markedly decreased the
concentration of IL-10 in the plasma and colon tissue samples, as
compared with the control (P<0.01). Notably, the rhein-treated
animals exhibited reduced expression levels of IL-1β, IL-6, IL-8,
and TNF-α, whereas they exhibited increased expression levels of
IL-10, as compared with the LPS model group (P<0.05, P<0.01,
respectively). Furthermore, treatment with TLR4 inhibitor TAK-242
significantly attenuated the LPS-induced inflammatory response,
increasing IL-10 expression levels and decreasing IL-1β, IL-6,
IL-8, TNF-α expression levels. These results were concordant with
those of the western blot analysis (Fig. 2). These results suggest that the
mechanism underlying the protective effects of rhein during
LPS-induced intestinal toxicity is similar to that of TAK-242.
| Table IILevel of inflammatory cytokines in the
plasma and colon tissue of LPS-induced septic mice. |
Table II
Level of inflammatory cytokines in the
plasma and colon tissue of LPS-induced septic mice.
| Cytokine (pg/ml) | Normal | Model | Rhein (100
mg/kg) | Rhein (100 mg/kg) +
TAK-242 (0.3 mg/kg) | TAK-242 (0.3
mg/kg) |
|---|
| Plasma |
| IL-1β | 38.44±5.22 | 55.41±4.17b | 45.38±2.14c | 44.66±3.07c | 46.99±4.95c |
| IL-6 | 21.45±3.73 | 38.33±4.56a | 26.82±4.26c | 24.32±3.92c | 26.18±4.01c |
| IL-8 | 44.52±5.24 | 62.49±8.74b | 51.77±4.28c | 49.51±4.54c | 49.03±2.90c |
| IL-10 | 66.54±6.83 | 37.82±8.55b | 52.54±6.77d | 55.34±7.12d | 53.11±6.03d |
| TNF-α | 7.73±1.56 | 11.95±2.12a | 8.51±1.33c | 8.22±0.94c | 8.75±1.17c |
| Colon tissue |
| IL-1β | 58.78±5.86 | 75.44±5.83a | 65.49±4.72c | 62.37±3.36d | 66.91±4.64c |
| IL-6 | 44.27±4.31 | 62.73±5.19b | 52.25±3.22c | 49.67±3.67d | 54.74±3.88c |
| IL-8 | 77.10±8.59 | 100.42±7.66b | 87.24±9.75d | 84.22±6.51d | 88.04±8.93d |
| IL-10 | 108.44±11.43 | 67.48±8.41b | 87.26±10.52d | 93.79±9.68d | 83.57±8.66d |
| TNF-α | 21.46±5.37 | 37.27±2.53a | 28.11±3.04c | 25.18±4.74c | 27.30±4.35c |
Rhein-attenuates LPS-induced TLR4
expression
In order to evaluate the effects of rhein on TLR4,
immunohistochemistry and western blot analyses were performed to
determine the expression levels of TLR4 in the colon. The results
from the immunohistochemical imaging of the sections demonstrated
the localization of TLR4 to the colon in the LPS-treated mice,
which in turn resulted in increased expression levels of TLR-4
(Fig. 3A–D). However, the
expression levels of TLR4 were significantly reduced following
treatment with rhein, both in the presence and absence of TAK-242
(P<0.01). Notably, the expression levels of TLR4 in the rhein or
TAK-242-treated mice were close to normal. In addition, there was
no statistically significant difference between the expression
levels of TLR4 in the mice co-treated with rhein and TAK-242, as
compared with the mice treated with rhein alone.
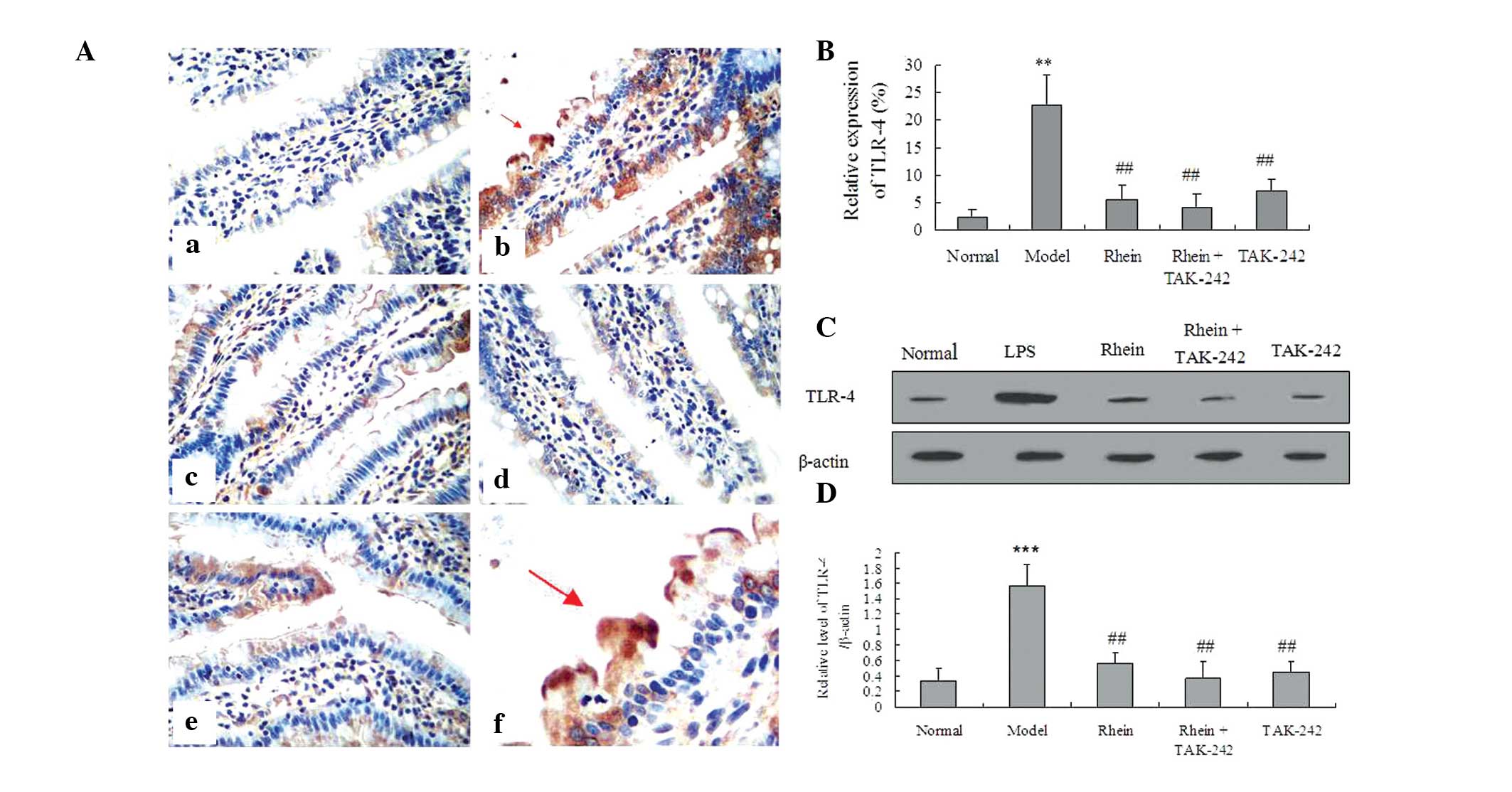 | Figure 3Effects of rhein on TLR4 expression in
colon tissue. (A and B) Immunohistochemistry of TLR4 expression;
(a) normal, (b) 20 mg/kg LPS, (c) 20 mg/kg LPS + 100 mg/kg rhein,
(d) 20 mg/kg LPS + 100 mg/kg rhein + 0.3 mg/kg TAK-242, (e) 20
mg/kg LPS + 0.3 mg/kg TAK-242, (f) enlargement of (A)b image. (C
and D) Western blotting of TLR4 expression. The data are presented
as the mean ± standard deviation (n=3). **P<0.01,
***P<0.001, vs. the normal control group;
##P<0.01, vs. the model group (magnification, ×400).
Red arrows indicate positive expression. LPS, lipopolysaccharide;
TLR, toll-like receptor. |
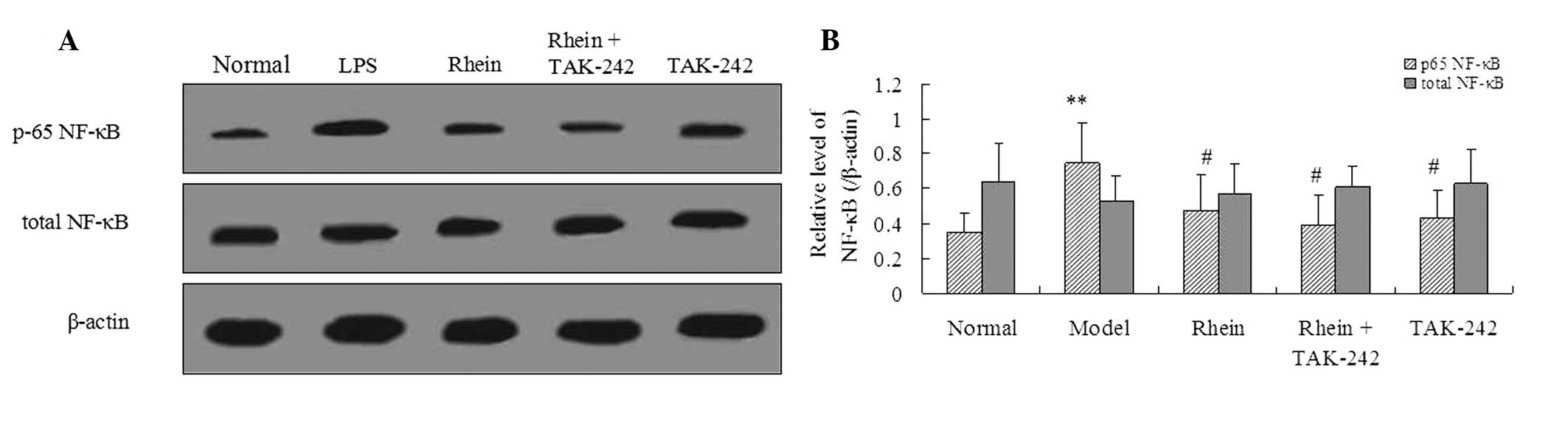
Rhein reduces NF-κB phosphorylation via
TLR4
To further investigate the effects of the rhein
signaling pathway on LPS-induced intestinal toxicity during sepsis,
the expression levels of both phosphorylated and total NF-κB were
analyzed. Following treatment with LPS, the expression levels of
the phosphorylated NF-κB were notably increased, as compared with
the control. This upregulation was reduced following treatment with
100 mg/kg rhein (P<0.05). However, the expression levels of
total NF-κB were not significantly different in any of the four
groups, as compared with the control group. Notably, LPS-mediated
NF-κB phosphorylation was inhibited by treatment with TAK-242
(Fig. 4).
Discussion
Sepsis is regarded as one of the leading causes of
mortality in critically ill patients exhibiting a systemic
inflammatory response caused by infection. The onset of sepsis
usually occurs following an excessive or uncontrolled inflammatory
response (18). The causal
relationship between intestinal damage and sepsis indicates that
the intestinal tract is not only a target organ, but also the
starting point of sepsis (5,19).
The LPS endotoxin is one of the most notable components of the
bacterial cell wall, and is released into the host during bacterial
growth, reproduction, lysis, or cell death (3). In addition, LPS has been suggested as
a major causative factor for the onset of sepsis and septic shock
caused by gram-negative bacilli (20). The accumulation of LPS activates an
inflammatory response that leads to damage of the intestinal
mucosal barrier, which in turn causes an increase in intestinal
mucosa permeability that promotes endotoxin translocation. The
present study demonstrated the protective effect of rhein, a
natural constituent of Rheum rhabarbarum, on LPS-induced
intestinal damage, and determined that rhein acted via the TLR4
NF-κB signaling pathway.
The TLR4 receptor detects foreign antigens and
initiates the innate immune response. The ability of TLR4 to
rapidly detect pathogens is key to the inflammatory response
initiated in patients with severe sepsis (21). The LPS-induced overexpression of
TLR4 is associated with changes in the levels of inflammatory
mediators. Accumulating evidence has suggested that LPS-induced
overactivation of the TLR4 signaling pathway leads to an increase
in the production of pro-inflammatory mediators, with fatal
consequences to the host (21–23).
The results of the present study demonstrated that exposure to 20
mg/kg LPS increased the expression levels of TLR4 in mice colon
tissue. These results support the hypothesis that LPS activates the
TLR4 signaling pathway, which in turn regulates the inflammatory
response (23). Conversely, the
expression levels of TLR4 in the colon of LPS-treated mice were
inhibited by rhein and TAK-242. Notably, the action of rhein on
LPS-induced colon injury was similar to the action of TAK-242. Gao
et al (24) previously
demonstrated that rhein exhibited both pro- and anti-inflammatory
activity on LPS-activated macrophages, by inhibiting the IκB
kinase. However, little research has investigated the target of
rhein in vivo. The results of the present study provide a
novel insight into the protective effects of rhein on LPS-induced
colon injury in mice.
LPS-induced inflammatory responses may impair
intestinal integrity (25). During
the initiation of the inflammatory septic process, changes occur in
the expression levels of numerous inflammatory mediators, including
an increase in the expression levels of pro-inflammatory cytokines
IL-8, IL-6 and IL-1β, and anti-inflammatory cytokine IL-10
(26). Sepsis and endotoxemia have
been associated with increased production of IL-1β, IL-6, IL-8, and
TNF-α in the plasma and colon tissue (26). The results of the present study
showed that the presence of LPS resulted in increased expression of
IL-1β, IL-6, IL-8, TNF-α in the plasma and colon tissue. Wang et
al (27) previously
demonstrated that both TNF-α and IL-1β may be involved in the
regulation of gastrointestinal IL-6 production during endotoxemia
in mice.
IL-10 is an early mediator and pleiotropic cytokine
of the inflammatory signaling cascade that becomes elevated within
a few hours of injury (28).
Numerous studies have demonstrated that the anti-inflammatory
cytokine IL-10 acts as a key mediator for the maintenance of gut
homeostasis, by inhibiting selective elements within the
inflammatory signaling cascade (28,29).
In immune modulator-impaired IL-10(−/−) mice,
LPS was shown to promote and exacerbate intestinal inflammation
(30). In addition, a previous
study reported that TLR-4(lps-lps-) mice exhibited
markedly increased mRNA expression levels of IL-10 (31). In the present study, the expression
levels of IL-10 in the colon tissue and plasma of mice exposed to
LPS were significantly increased by rhein treatment in the presence
or absence of TAK-242. These results suggested that rhein was able
to attenuate LPS-induced intestinal inflammation, and emphasize the
activation of IL-10 in the pathogenesis of sepsis and
endotoxemia.
The NF-κB transcription factor mediates the
inflammatory response of sepsis and endotoxemia (32). LPS-induced NF-κB activation
associated with TLR4 caused colon shortening and increased
expression levels of IL-1β, IL-6, and TNF-α in mice (33). In the present study, a marked
increase in NF-κB phosphorylation occurred following LPS treatment.
However, treatment with rhein significantly inhibited LPS-induced
NF-κB phosphorylation. The inhibition of TLR4 by TAK-242 also
attenuated increased NF-κB phosphorylation. No statistically
significant difference was detected in the total NF-κB levels
between the various groups. The results of the present study
demonstrated that the increase in LPS-induced NF-κB phosphorylation
was markedly inhibited by treatment with rhein and TLR4 inhibitor
TAK-242.
In conclusion, the present study demonstrated that
rhein significantly attenuated the LPS-induced inflammatory
response involved in intestinal injury during sepsis. This
protection may be associated with the LPS/TLR4/NF-κB signaling
pathway. The results of the present study provide a novel insight
into the mechanism underlying the protective effects of rhein on
LPS-induced intestinal injury.
Acknowledgments
The present study was supported by a grant from the
National Natural Science Foundation of China (grant no.
81202917).
References
|
1
|
Rajkumari N, Mathur P, Sharma S, Gupta B,
Bhoi S and Misra MC: Procalcitonin as a predictor of sepsis and
outcome in severe trauma patients: A prospective study. J Lab
Physicians. 5:100–108. 2013. View Article : Google Scholar
|
|
2
|
Duran-Bedolla J, Montes de Oca-Sandoval
MA, Saldaña-Navor V, Villalobos-Silva JA, Rodriguez MC and
Rivas-Arancibia S: Sepsis, mitochondrial failure and multiple organ
dysfunction. Clin Invest Med. 37:E58–E69. 2014.PubMed/NCBI
|
|
3
|
Hatakeyama N and Matsuda N: Mechanisms of
inflammatory response and organ dysfunction: Organ-protective
strategy by anesthetics. Curr Pharm Des. Feb 3–2014.Epub ahead of
print. View Article : Google Scholar
|
|
4
|
Luo HM, Du MH, Lin ZL, et al: Valproic
acid treatment inhibits hypoxia-inducible factor 1α accumulation
and protects against burn-induced gut barrier dysfunction in a
rodent model. PLoS One. 8:e775232013. View Article : Google Scholar
|
|
5
|
Fink MP: Intestinal epithelial
hyperpermeability: Update on the pathogenesis of gut mucosal
barrier dysfunction in critical illness. Curr Opin Crit Care.
9:143–151. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chatterjee N, Das S, Bose D, Banerjee S,
Jha T and Saha KD: Leishmanial lipid suppresses the bacterial
endotoxin-induced inflammatory response with attenuation of tissue
injury in sepsis. J Leukoc Biol. 96:325–336. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fatemi K, Radvar M, Rezaee A, et al:
Comparison of relative TLR-2 and TLR-4 expression level of disease
and healthy gingival tissue of smoking and non-smoking patients and
periodontally healthy control patients. Aust Dent J. 58:315–320.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Musie E, Moore CC, Martin EN and Scheld
WM: Toll-like receptor 4 stimulation prior to or after
Streptococcus pneumoniae induced sepsis improves survival and is
dependent on T-cells. PLoS One. 9:e860152014. View Article : Google Scholar
|
|
9
|
Yang JC, Wu SC, Rau CS, et al: Inhibition
of the phosphoinositide 3-kinase pathway decreases innate
resistance to lipopolysaccharide toxicity in TLR4 deficient mice. J
Biomed Sci. 21:202014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mascolo N, Autore G, Izzo AA, Biondi A and
Capasso F: Effects of senna and its active compounds rhein and
rhein-anthrone on PAF formation by rat colon. J Pharm Pharmacol.
44:693–695. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Raimondi F, Santoro P, Maiuri L, et al:
Reactive nitrogen species modulate the effects of rhein, an active
component of senna laxatives, on human epithelium in vitro. J
Pediatr Gastroenterol Nutr. 34:529–534. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Peng SN, Zeng HH, Fu AX, Chen XW and Zhu
QX: Effects of rhein on intestinal epithelial tight junction in IgA
nephropathy. World J Gastroenterol. 19:4137–4145. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang Q, Ma YM, Wang ZT and Wang CH:
Differences in pharmacokinetics and anti-inflammatory effects
between decoction and maceration of Sanhuang Xiexin Tang in rats
and mice. Planta Med. 79:1666–1673. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guo MZ, Li XS, Xu HR, Mei ZC, Shen W and
Ye XF: Rhein inhibits liver fibrosis induced by carbon
tetrachloride in rats. Acta Pharmacol Sin. 23:739–744.
2002.PubMed/NCBI
|
|
15
|
Ni JQ, Ouyang Q, Lin L, et al: Role of
toll-like receptor 4 on lupus lung injury and atherosclerosis in
LPS-challenge ApoE−/−mice. Clin Dev Immunol.
2013:4768562013. View Article : Google Scholar
|
|
16
|
Kim WH, Song HO, Jin CM, et al: The
methanol extract of Azadirachta indica A. juss leaf protects mice
against lethal endotoxemia and sepsis. Biomol Ther (Seoul).
20:96–103. 2012. View Article : Google Scholar
|
|
17
|
Chiu CJ, McArdle AH, Brown R, Scott HJ and
Gurd FN: Intestinal mucosal lesion in low-flow states. I. A
morphological, hemodynamic, and metabolic reappraisal. Arch Surg.
101:478–483. 1970. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cordioli RL, Cordioli E, Negrini R and
Silva E: Sepsis and pregnancy: Do we know how to treat this
situation? Rev Bras Ter Intensiva. 25:334–344. 2013. View Article : Google Scholar
|
|
19
|
Van Leeuwen PA, Boermeester MA, Houdijk
AP, et al: Clinical significance of translocation. Gut. 35(Suppl
1): 28–S34. 1994. View Article : Google Scholar
|
|
20
|
Cross AS, Sidberry H and Sadoff JC: The
human antibody response during natural bacteremic infection with
gram-negative bacilli against lipopolysaccharide core determinants.
J Infect Dis. 160:225–236. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Piazza O, Pulcrano G, Fiori PL, et al:
Toll-like receptor kinetics in septic shock patients: A preliminary
study. Int J Immunopathol Pharmacol. 25:425–433. 2012.PubMed/NCBI
|
|
22
|
Drago-Serrano ME, de la Garza-Amaya M,
Luna JS and Campos-Rodríguez R: Lactoferrin-lipopolysaccharide
(LPS) binding as key to antibacterial and antiendotoxic effects.
Int Immunopharmacol. 12:1–9. 2012. View Article : Google Scholar
|
|
23
|
Verstak B, Stack J, Ve T, et al: The TLR
signaling adaptor TRAM interacts with TRAF6 to mediate activation
of the inflammatory response by TLR4. J Leukoc Biol. 96:427–436.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gao Y, Chen X, Fang L, et al: Rhein exerts
pro- and anti-inflammatory actions by targeting IKKβ inhibition in
LPS-activated macrophages. Free Radic Biol Med. 72:104–112. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sautner T, Wessely C, Riegler M, et al:
Early effects of catecholamine therapy on mucosal integrity,
intestinal blood flow, and oxygen metabolism in porcine endotoxin
shock. Ann Surg. 228:239–248. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Redondo AC, Ceccon ME, Silveira-Lessa AL,
et al: TLR-2 and TLR-4 expression in monocytes of newborns with
late-onset sepsis. J Pediatr (Rio J). 90:472–478. 2014. View Article : Google Scholar
|
|
27
|
Wang Q, Wang JJ, Boyce S, Fischer JE and
Hasselgren PO: Endotoxemia and IL-1 beta stimulate mucosal IL-6
production in different parts of the gastrointestinal tract. J Surg
Res. 76:27–31. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ueda Y, Kayama H, Jeon SG, et al:
Commensal microbiota induce LPS hyporesponsiveness in colonic
macrophages via the production of IL-10. Int Immunol. 22:953–962.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Paul G, Khare V and Gasche C: Inflamed gut
mucosa: Downstream of interleukin-10. Eur J Clin Invest. 42:95–109.
2012. View Article : Google Scholar
|
|
30
|
Im E, Riegler FM, Pothoulakis C and Rhee
SH: Elevated lipopolysaccharide in the colon evokes intestinal
inflammation, aggravated in immune modulator-impaired mice. Am J
Physiol Gastrointest Liver Physiol. 303:G490–G497. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chung YW, Choi JH, Oh TY, Eun CS and Han
DS: Lactobacillus casei prevents the development of dextran
sulphate sodium-induced colitis in Toll-like receptor 4 mutant
mice. Clin Exp Immunol. 151:182–189. 2008. View Article : Google Scholar
|
|
32
|
Teo JD, Macary PA and Tan KS: Pleiotropic
effects of Blastocystis spp. Subtypes 4 and 7 on ligand-specific
toll-like receptor signaling and NF-κB activation in a human
monocyte cell line. PLoS One. 9:e890362014. View Article : Google Scholar
|
|
33
|
Lee JH, Lee B, Lee HS, et al:
Lactobacillus suntoryeus inhibits pro-inflammatory cytokine
expression and TLR-4-linked NF-kappaB activation in experimental
colitis. Int J Colorectal Dis. 24:231–237. 2009. View Article : Google Scholar
|