Introduction
Alzheimer's disease (AD) is a progressive
neurodegenerative disorder characterized by cognitive and memory
deterioration, changes in personality, behavioral disturbances and
an impaired ability to perform daily living activities (1). The number of patients with AD is
increasing every year. Therefore, an effective therapeutic drug is
still an urgent requirement for the treatment of AD (2). Extracellular senile plaque (SP)
deposition is one of the pathological markers in the brains of
patients with AD. SP originates from the abnormal extracellular
beta-amyloid (Aβ) aggregation caused by gene mutations in the Aβ
precursor protein (APP). A previous study demonstrated that Aβ
aggregation activated the N-methyl-d-aspartate (NMDA) receptor
(3,4), induced neurotoxicity and caused
neuronal apoptosis and degenerative death (5). Therefore, developing drugs that
inhibit neuronal death is important for delaying AD
progression.
Memantine is a non-competitive NMDA receptor
antagonist, which has been approved for the treatment of
moderate-to-severe AD. Numerous clinical trials reported that
memantine was able to improve attention, cognitive ability and
functional communication in patients with AD (6–11). In
vitro studies demonstrated that memantine reduced the secretion
of APP and Aβ in APP/PS1 transgenic mice, cultured cortical cells
and SK-N-SH human neuroblastoma cells (12,13).
Further studies also demonstrated that memantine promotes the
proliferation of neural progenitor cells and the production of
mature granular neurons in the adult hippocampus (14–16).
However, the effects of memantine on neuronal autophagy and
apoptosis remain to be elucidated. In addition, it remains
controversial whether autophagy promotes Aβ production
(17–20) or reduces Aβ levels (21–24).
The present study established an AD cell model
overexpressing APP with the 695-amino-acid Swedish mutation
(APP695) to determine the effects of memantine on cell autophagy
and apoptosis, and to explore the association between autophagy and
apoptosis, as well as the underlying mechanisms and signaling
pathways involved.
Materials and methods
Cell culture and establishment of an AD
cell model
The SH-SY5Y human neuroblastoma cell line was
purchased from the Shanghai Institute for Biological Sciences of
the Chinese Academy of Sciences (Shanghai, China). The cells were
cultured in Dulbecco's modified Eagle's medium (Gibco-BRL,
Invitrogen Life Technologies, Carlsbad, CA, USA) supplemented with
10% fetal bovine serum (GE Healthcare Life Sciences, Little
Chalfont, UK) and streptomycin/penicillin (100 U/ml) (China
National Medicines Corporation, Ltd., Beijing, China) at 37°C in an
atmosphere containing 5% CO2. The AD cell model was
established a previously described (25,26).
When the SH-SY5Y cells reached 70% confluence, the cells were
transfected with overexpression plasmid pcDNA3.1-APP695, containing
a Swedish mutant APP695 gene (Wanleibio, Shenyang, China) using the
Lipofectamine® 2000 reagent (Invitrogen Life
Technologies) according to the manufacturer's instructions. The
cells were separated into the following groups: A non-transfected
control group (Control), a transfection reagent-treated group
(Mock), an empty plasmid transfection group (pcDNA3.1) and a
pcDNA31-APP695-transfected group (APP695). The cells were collected
after 24 or 48 h of transfection for subsequent experiments.
Drug treatment
Memantine hydrochloride (Sigma-Aldrich, St. Louis,
MO, USA) was dissolved in double distilled (dd)H2O.
Rapamycin (RAPA; Melone Pharmaceutical Co., Ltd., Dalian, China)
was dissolved in dimethyl sulfoxide (DMSO; Sigma-Aldrich). After 24
or 48 h of transfection, the cells were treated with various
concentrations of memantine (1, 5 and 10 µM) to select an optimal
concentration for subsequent experiments. For enhanced autophagy
experiments, the cells were pre-treated with 0.2 µM RAPA for 24 h
prior to treatment with 5 µM memantine for 24 h.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
At 24 h post-transfection, the cells were harvested
and total RNA from each sample was extracted using a RNA simple
Total RNA kit (cat no. DP419; Tiangen Biotech Co., Ltd., Beijing,
China). The RNA was then re-suspended in 40 µl RNase-free
ddH2O and was reverse transcribed into cDNA using the
PrimeScript™ RT reagent kit (cat no. RR037A; Takara Bio Inc.,
Dalian, China). Fluorescence quantitative analysis was performed
with a SYBR® GREEN MasterMix (Tiangen Biotech Co., Ltd.)
using an Exicycler™ 96 quantitative fluorescence analyzer (Bioneer
Corporation, Daejeon, Korea). The thermocycling conditions were set
as follows: 95°C for 10 min; 40 cycles of 95°C for 10 sec, 60°C for
20 sec and 72°C for 30 sec; and final elongation at 4°C for 5 min.
β-actin was included in each reaction as an internal control, and
the relative gene expression levels were calculated using the
2−ΔΔCt method (27).
The primer sequences for the RT-qPCR were as follows: APP695 sense,
5′-GGACGATGAGGATGGTGATGAG-3′ and anti-sense,
5′-GGTACTGGCTGCTGTTGTAGGA-3′; β-actin sense,
5′-CTTAGTTGCGTTACACCCTTTCTTG-3′ and anti-sense,
5′-CTGTCACCTTCACCGTTCCAGTTT-3′. PCR primers were synthesized by
Sangon Biotech (Shanghai, China).
Western blot analysis
Following 48 h of transfection, total protein from
each group was extracted using NP-40 lysate (Beyotime Institute of
Biotechnology, Haimen, China) with 1% phenyl-methanesulfonyl
fluoride (Beyotime Institute of Biotechnology). The protein
concentration of each sample was determined using a Bicinchoninic
Acid Protein Assay kit (cat no. P0012S; Beyotime Institute of
Biotechnology). A total of 40 µg protein from each group was
separated by SDS-PAGE (5% for the spacer gel and 8% for the
separating gel) and transferred onto polyvinylidene fluoride
membranes (EMD Millipore, Billerica, MA, USA). The membranes were
blocked with 5% non-fat milk overnight and incubated at 4°C
overnight with the following primary antibodies: Rabbit polyclonal
anti-APP695 (1:500; cat no. BA0581; Wuhan Boster Biological
Technology, Ltd., Wuhan, China), rabbit polyclonal anti-caspase-3
(1:1,000; cat no. bs-0081R; BIOSS, Beijing, China), rabbit
polyclonal anti-light chain 3 (LC3; 1:1,000; cat no. 4108; Cell
Signaling Technology, Inc., Danvers, MA, USA), rabbit polyclonal
anti-autophagy protein 5 (ATG5; 1:200; cat no. BA3525-2; Wuhan
Boster Biological Technology, Ltd.), rabbit polyclonal
immunoglobulin (Ig)G to phosphorylated (p)-Akt (1:200; cat no.
sc-135650; Santa Cruz Biotechnology Inc., Dallas, TX, USA), rabbit
polyclonal IgG to B-cell lymphoma 2 (Bcl-2; 1:200; cat no. sc-492;
Santa Cruz Biotechnology, Inc.); mouse monoclonal IgG1 to
phosphoinositide 3-kinase p85α (PI3K; 1:200; cat no. sc-1637; Santa
Cruz Biotechnology, Inc.), rabbit polyclonal IgG to p-mammalian
target of rapamycin (mTOR; 1:100; cat no. sc-8319; Santa Cruz
Biotechnology, Inc.), rabbit polyclonal anti-Beclin-1 (1:5,000; cat
no. ab55878; Abcam, Cambridge, MA, USA) and rabbit monoclonal to
p-mitogen-activated protein kinase 8 (JNK1; 1:100; cat no.
EPR17557; Abcam). Membranes were then washed four times for 5 min
with Tris-buffered saline containing Tween 20, followed by
incubation with the corresponding secondary antibodies, goat
anti-rabbit immunoglobulin (Ig)G-HRP and goat anti-mouse IgG-HRP
(1:5,000; cat. nos. A0208 and A0216, respectively; Beyotime
Institute of Biotechnology), for 90 min at room temperature. For
visualization, the membranes were treated with an enhanced
chemiluminescence solution (ECL, Qihai Biotec, Shanghai, China),
exposed to X-ray film and visualized by autoradiography. The
optical density (OD) values of the target protein bands were
analyzed using Gel-Pro-Analyzer software 4.0 (Media Cybernetics,
Rockville, MD, USA) and normalized to β-actin (1:5,000;
HRP-conjugated monoclonal mouse anti-β-actin; cat. no. KC-5A08;
KangChen Bio-tech Inc., Shanghai, China).
Detection of cell viability using an MTT
assay
Following transfection for 48 h, cells were seeded
onto 96-well plates at 1×103 cells/well and treated with
various concentrations of memantine (1, 5 and 10 µM); each
experiment was performed with five replicates. After 24 and 48 h,
MTT solution (final concentration, 0.2 mg/ml; Sigma-Aldrich) was
added and incubated with the cells for 5 h. The supernatant was
carefully discarded and 200 µl DMSO (Sigma-Aldrich) was added to
fully dissolve the crystals. OD values were measured at 490 nm
using a microplate reader (ELX800; BioTek Instruments, Inc.,
Winooski, VT, USA).
Observation of autophagosomes using
acridine orange
Following 24 h transfection, the cells were seeded
in 12-well plates at a density of 1×105 cells/well.
Following culturing in the presence of the corresponding drugs, the
cells from each group were fixed in 4% paraformaldehyde (China
National Medicines Corporation, Ltd.) for 15 min and washed four
times with phosphate-buffered saline (PBS). Acridine orange
(Beijing Solarbio Science & Technology Co., Ltd., Beijing,
China) solution (0.01%) was added onto the cells in order to
completely cover them; the cells were then incubated at room
temperature for 5 min. The stained cells were then mounted with an
anti-fluorescein quencher (Beijing Solarbio Science &
Technology Co., Ltd.) and were observed under a laser scanning
confocal microscope (FV1000S-SIM/IX81; Olympus Corp., Tokyo, Japan)
and images were captured.
Detection of apoptosis using flow
cytometry
Following 24 h of transfection, the cells were
seeded into six-well plates at 1×105 cells/well. The
cells from each group were collected following drug treatment and
washed twice with PBS. The cells were then re-suspended in 500 µl
binding buffer and mixed sequentially with 5 µl Annexin
V-allophycocyanin (APC) and 5 µl 7-aminoactinomycin D (7-AAD) using
an Annexin V-APC/7-AAD Apoptosis Detection kit (cat no. KGA1025;
Nanjing KeyGen Biotech Co., Ltd., Nanjing, China), according to the
manufacturer's instructions. The cells were subsequently incubated
at room temperature in the dark for 15 min, and apoptotic rates
were detected using a FACSCalibur flow cytometer (BD Biosciences,
Franklin Lakes, USA). The data were analyzed using CellQuest Pro
3.3 software (BD Biosciences).
Immunofluorescence double staining
Following 48 h of transfection, the cells were
plated onto coverslips in six-well plates. Following cellular
treatment with each group of drugs, the cells on the coverslips
from each group were fixed in 4% paraformaldehyde for 15 min and
permeabilized in 0.1% Triton X-100 (Amresco, Solon, OH, USA) for 30
min. The cells were incubated with anti-Bcl-2 (1:200) and Beclin-1
(1:100) primary antibodies at 4°C overnight. The cells were then
incubated with cyanine 3-labeled goat anti-mouse IgG (H+L) and
fluorescein isothiocyanate-labeled goat anti-rabbit IgG (H+L)
secondary antibodies (cat nos. A0521 and A0562, respectively;
Beyotime Institute of Biotechnology) at a dilution of 1:100 for 90
min at room temperature. After staining with DAPI (Biosharp, Anhui,
China), the coverslips were mounted inversely on slides with an
anti-fluorescein quencher and were observed under a laser-scanning
confocal microscope (Olympus Corporation) and the images were
captured.
Statistical analysis
All values are expressed as the mean ± standard
deviation. The data were analyzed using Graphpad Prism 5.0
(GraphPad Software, Inc., La Jolla, CA, USA). Comparisons between
two groups were performed using a one-way analysis of variance,
whereas the comparisons among multiple groups were performed with
Bonferroni's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Establishment and identification of an AD
cell model overexpressing APP695 Swedish mutant gene
To establish an AD cell model, a pcDNA3.1-APP695
plasmid overexpressing the APP695 Swedish mutant gene was
transfected into SH-SY5Y cells, and the expression levels of APP695
in the cells were detected using RT-qPCR and western blot analyses.
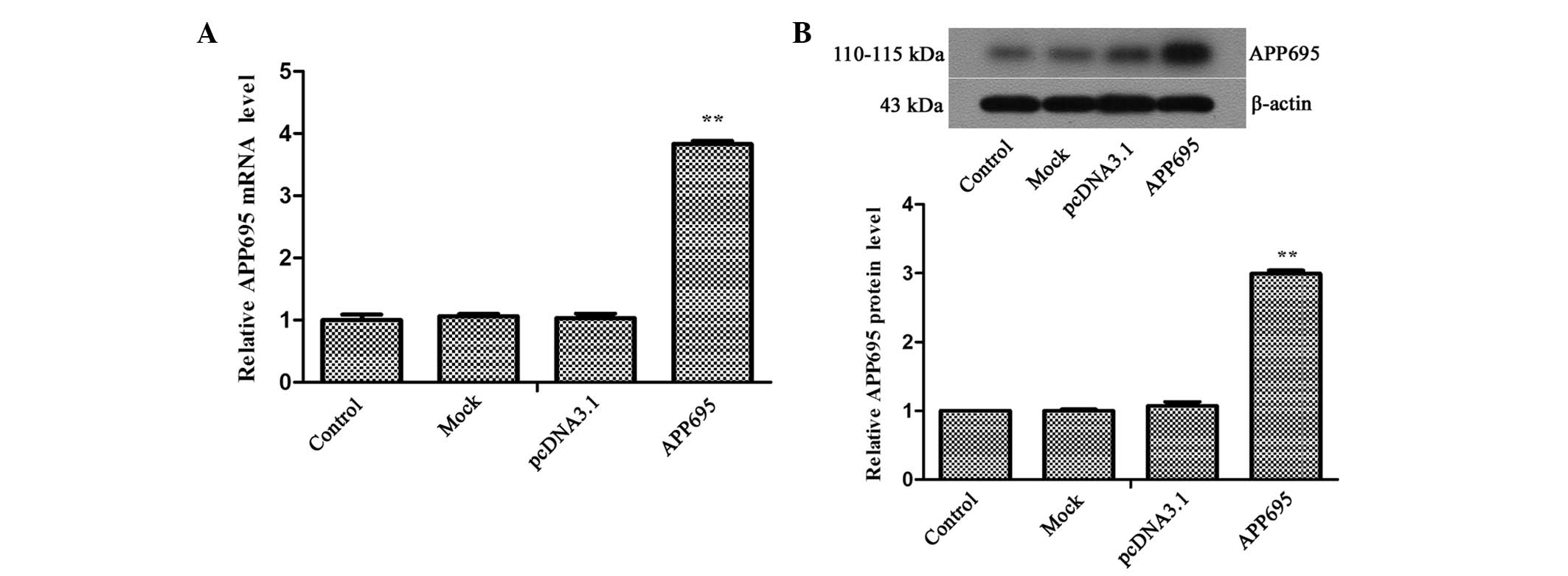
The mRNA and protein expression levels of APP695 in the APP695
transfection group were 3.83-fold (Fig. 1A; P<0.01) and 2.99-fold
(Fig. 1B; P<0.01) higher,
respectively, as compared with those in the pcDNA3.1 group. These
results suggested that the AD cell model was successfully
established.
Memantine enhances neuronal cell
viability and inhibits autophagy in a cell model of AD
The effects of various concentrations of memantine
on neuronal cell viability in the AD model were detected using an
MTT assay. The results indicated that cell viability in the APP695
group at 24 (Fig. 2A; P<0.05)
and 48 h (P<0.01) were significantly decreased, as compared with
those in the pcDNA3.1 group. However, memantine reversed the
inhibition of cell proliferation caused by APP695 overexpression.
Cell viability was significantly increased following cellular
treatment with three different doses of memantine for 24 (5 µM,
P<0.01; 1 µM and 10 µM, P<0.05) and 48 h (5 µM and 10 µM,
P<0.01), as compared with that in the APP695 group. However, no
statistically significant difference was observed between the 5
µM-treated group and the 10 µM-treated group. Therefore, 5 µM
memantine was the optimal treatment concentration, and was used in
the subsequent experiments.
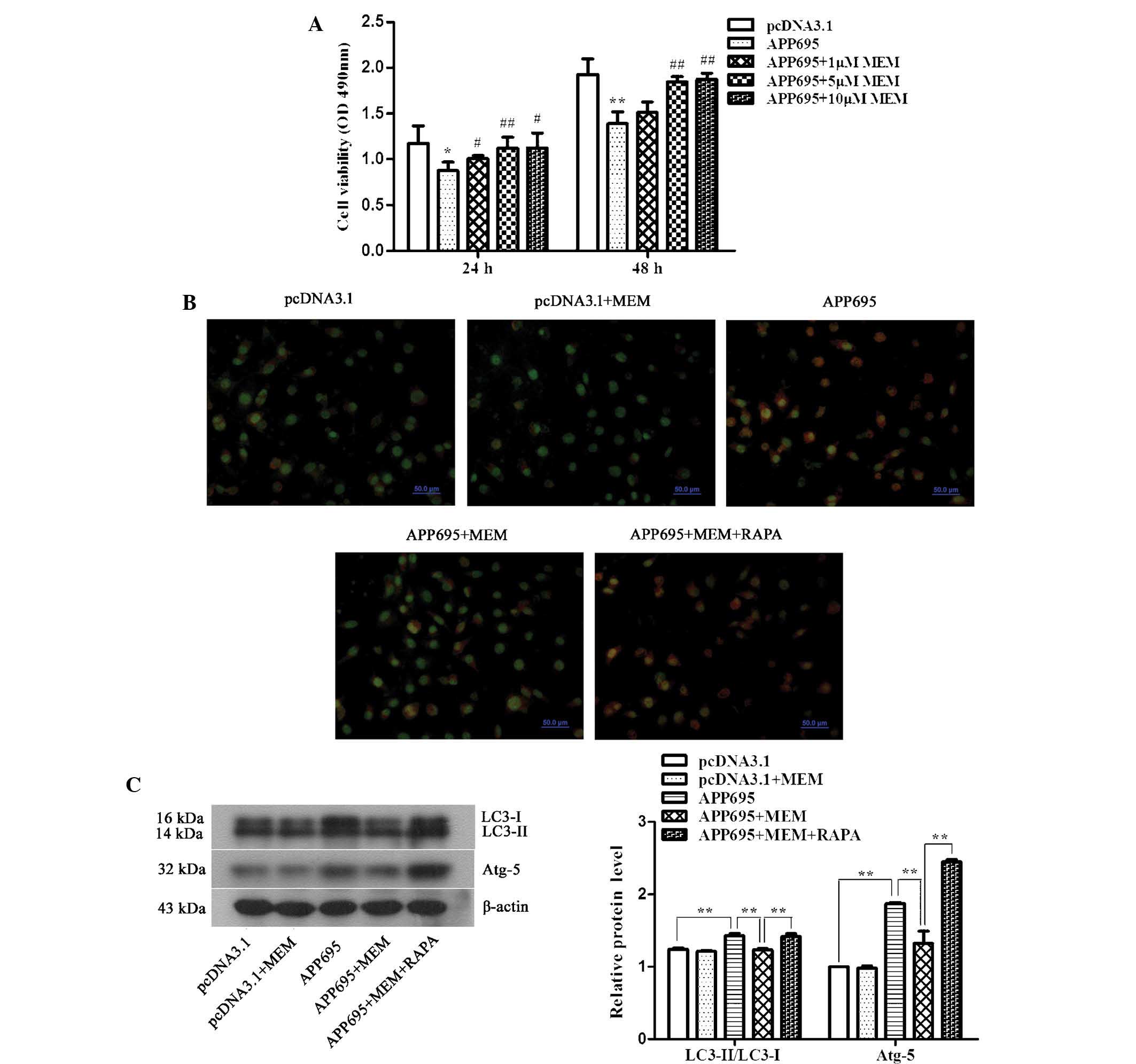 | Figure 2Increase in neuronal cell viability
and inhibition of cell autophagy in the Alzheimer's disease cell
model following treatment with memantine. (A) Detection of cell
viability using an MTT assay. Cells were inoculated onto 96-well
plates and treated with 1, 5 or 10 µM memantine. The absorbance
values (OD, 490 nm) of the cells from each group were measured at
24 and 48 h. *P<0.05 and **P<0.01, vs.
the pcDNA3.1 group; #P<0.05 and
##P<0.01, vs. the APP695 group. (B) Detection of
autophagosomes using acridine orange staining. The cells were
inoculated onto coverslips in 12-well plates. Following treatment
with the indicated drugs for 24 h, the cells were stained with
0.01% acridine orange. The results were observed under a laser
scanning confocal microscope (magnification, ×400; scale bar, 50
µm), and the most representative images are shown. Normal cells
exhibit green fluorescence and autophagosomes exhibit orangered
fluorescence. (C) Detection of LC3-II/LC3-I and ATG5 protein
expression levels as determined by western blot analysis. β-actin
was used as an internal control. Representative results from three
experiments repeated in triplicate are shown.
**P<0.01. All of the above experiments were repeated
in triplicate and all values are expressed as the mean ± standard
deviation. Groups: pcDNA3.1, cells transfected with empty vector;
APP695, cells transfected with APP695 overexpression vector;
memantine, cells treated with 5 µM memantine for 24 h; RAPA, cells
were pre-treated with 0.2 µM RAPA for 24 h prior to treatment with
memantine. LC3, protein light chain 3; ATG-5, autophagy protein 5;
OD, optical density; APP695, β-amyloid precursor protein with
695-amino-acid mutation; RAPA, rapamycin. |
To determine the effects of memantine on the
autophagic capacity of neuronal cells in the AD model, the acridine
orange staining method was used to observe autophagosomes, and
western blotting was performed to detect the expression levels of
the autophagy-associated proteins LC3-II/LC3-I and ATG5. Acridine
orange is a weak base that has the ability to move freely across
biological membranes in an uncharged state and is characterized by
green fluorescence. Acridine orange, in its protonated form,
accumulates in acidic compartments and forms aggregates, which are
characterized by yellow-orange fluorescence. As shown in Fig. 2B, cells in the pcDNA3.1 group
exhibited low levels of green fluorescence, indicating a lack of
acidic vesicular organelles (AVOs). Conversely, APP695
over-expression induced the formation of yellow-orange fluorescent
AVOs. The LC3-II/LC3-I ratio (Fig.
2C, P<0.01) and the expression levels of ATG5 (P<0.01) in
the APP695 group were significantly increased. Following treatment
with memantine, yellow-orange fluorescence, the LC3-II/LC3-I ratio
(P<0.01) and ATG5 expression levels (P<0.01) were
significantly decreased. The RAPA pre-treatment group showed a
statistically significant increase in yellow-orange fluorescence,
the LC-3II/LC3-I ratio (P<0.01) and the ATG5 expression levels
(P<0.01).
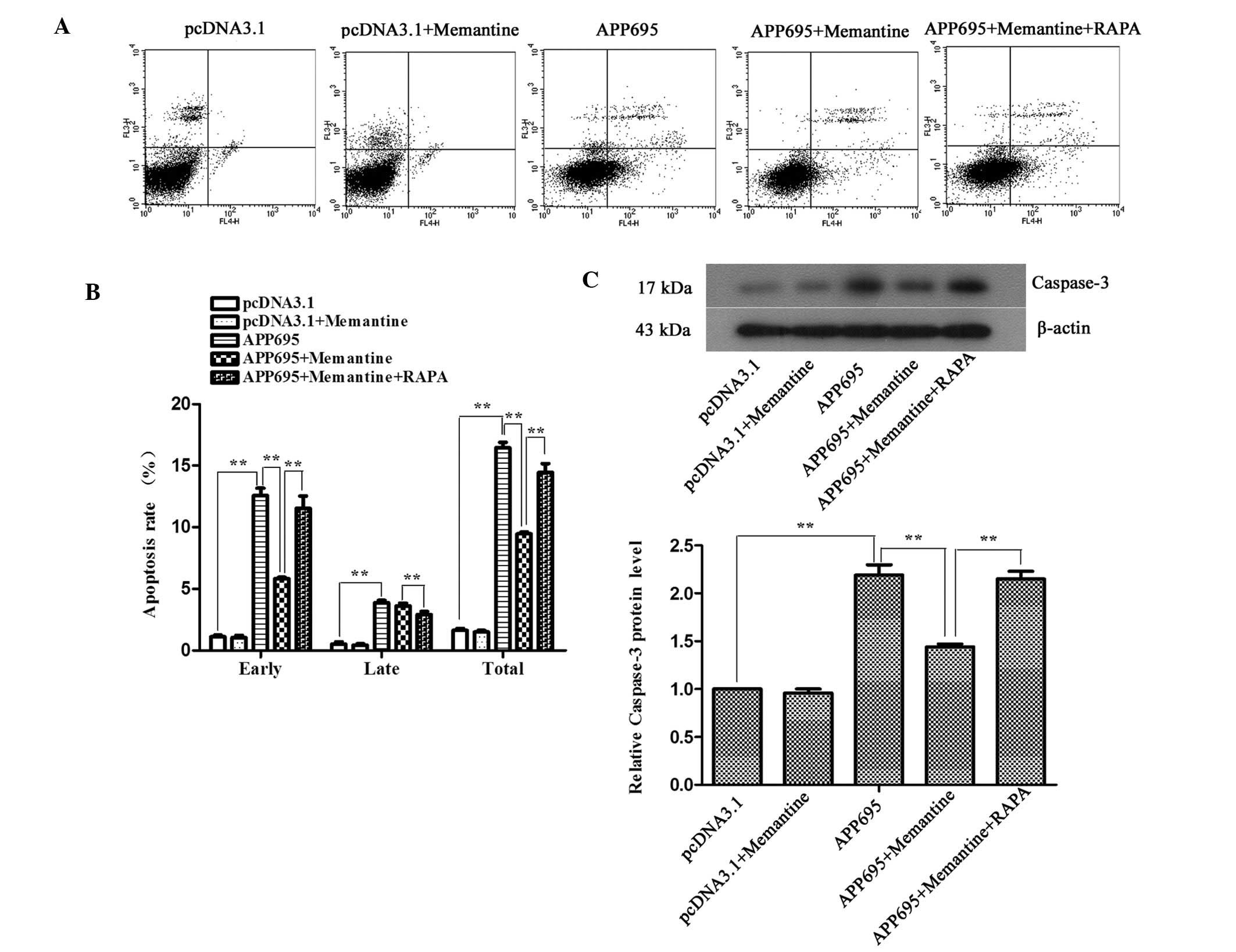
Memantine inhibits neuronal apoptosis in
a cell model of AD and RAPA-induced autophagy promotes cell
apoptosis
To detect the effects of memantine on neuronal
apoptosis in the AD model and to clarify the association between
autophagy and apoptosis, the apoptotic rates of the cells in each
group were determined using flow cytometry following Annexin
V-APC/7-AAD double staining. The expression levels of the
apoptosis-associated protein caspase-3 were detected by western
blot analysis. The apoptotic rates of the cells in the APP695 group
were significantly increased, as compared with those in the
pcDNA3.1 group (Fig. 3A and B;
P<0.01); the expression levels of caspase-3 also increased by
2.19-fold (Fig. 3C; P<0.01).
Memantine treatment significantly decreased the apoptotic rate
(P<0.01) and caspase-3 expression compared with those in the
APP695 group (P<0.01), indicating that memantine was able to
effectively inhibit neuronal apoptosis induced by APP695
overexpression. When the cells were pre-treated with RAPA,
autophagy induced by RAPA significantly increased the apoptosis
rates and promoted neuronal apoptosis (P<0.01).
Memantine regulates autophagic signaling
pathways in neuronal cells
The mechanisms underlying the inhibition of
autophagy by memantine were determined by detecting the activation
of the two mTOR-dependent and -independent signaling pathways.
Western blot analysis demonstrated that the expression levels of
p-PI3K, p-Akt and p-mTOR in the APP695 group were significantly
downregulated, as compared with those in the pcDNA3.1 group
(Fig. 4A; P<0.01). Following
treatment of the cells in the APP695 group with memantine, the
expression levels of the three signaling molecules significantly
increased (P<0.01), suggesting that APP695 overexpression
inhibited the PI3K/Akt/mTOR signaling pathway in neuronal cells and
to activate autophagy, whereas memantine reversed the inhibitory
state of the PI3K/Akt/mTOR signaling pathway, suppressing
autophagy. In the mTOR-independent signaling pathway, p-JNK1 and
Beclin-1 expression levels were significantly upregulated in the
APP695 group, as compared with those in the pcDNA3.1 group
(Fig. 4B; P<0.01), whereas the
expression levels of Bcl-2 expression were significantly
downregulated (P<0.01), which was attenuated by treatment with
memantine. Immunofluorescence staining was performed to further
investigate the expression of the Bcl-2/Beclin-1 complex. The
results indicated that the localizations of the Bcl-2 and Beclin-1
proteins in the cells were matched. The Bcl-2/Beclin-1 complex in
the pcDNA3.1 group and the pcDNA3.1 + Memantine group exhibited
orange-yellow fluorescence in the cytoplasm. In the APP695 group,
Bcl-2 was phosphorylated, and its expression levels were decreased;
therefore, the cytoplasm emitted emerald green fluorescence. In the
APP695 + Memantine group, the expression of the Bcl-2/Beclin-1
complex was increased and the color of the fluorescence in the
cytoplasm had returned to orange-yellow, similar to that in the
empty vector-transfected cells. These results indicated that
memantine activated the Bcl-2/Beclin-1 complex to inhibit autophagy
in the AD cell model via the suppression of JNK1 phosphorylation.
In conclusion, the above results demonstrated that the
mTOR-dependent as well as-independent pathways are involved in the
inhibition of cell autophagy by memantine.
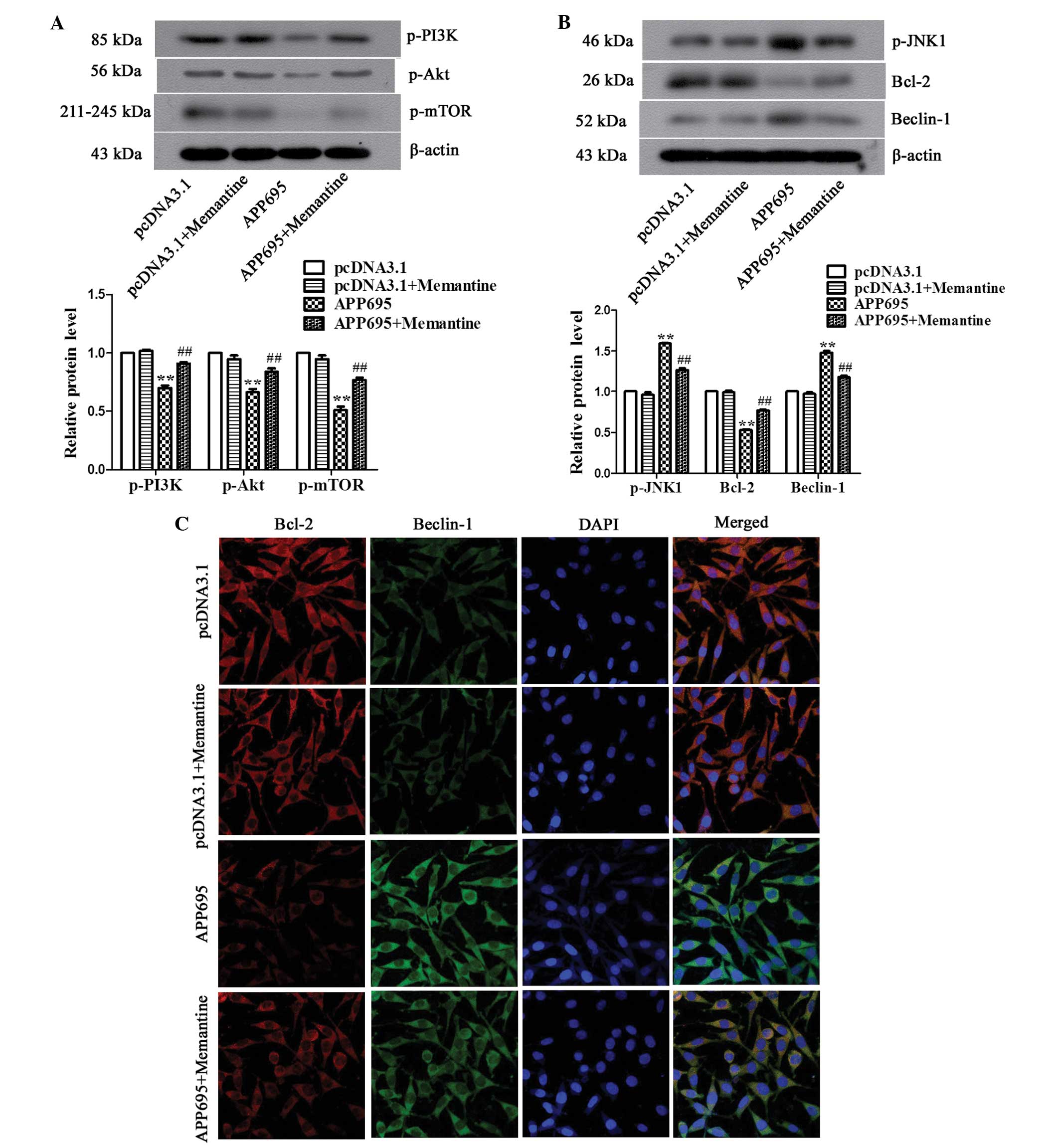 | Figure 4Regulation of autophagy-associated
signaling pathways in neuronal cells by memantine. (A) Detection of
the expression levels of PI3K, Akt and mTOR in the mTOR-dependent
signaling pathway, as determined by western blot analysis. (B)
Detection of the expression of p-JNK1, Bcl-2 and Beclin-1 in the
mTOR-independent pathway, as determined by western blot analysis.
(C) Observation of the expression of the Bcl-1/Beclin-1 complex
using immunofluorescence. Bcl-2 stained with Cy3-labeled goat
anti-mouse IgG (red), Beclin-1 stained with FITC-labeled goat
anti-rabbit IgG (green) and cell nuclei stained with DAPI (blue) in
the cells were observed under a laser scanning confocal microscope
(magnification, ×600). Representative data/images of three
independent experiments are shown. Values are expressed as the mean
± standard deviation. **P<0.01, vs. the pcDNA3.1
group; ##P<0.01 vs. the APP695 group. Groups:
pcDNA3.1, cells transfected with empty vector; APP695, cells
transfected with APP695 overexpression vector; memantine, cells
treated with 5 µM memantine for 24 h. P, phosphorylated; PI3K,
phosphoinositide 3-kinase; mTOR, mammalian target of rapamycin;
JNK1, mitogen-activated protein kinase 8; Bcl-2, B cell lymphoma 2;
Cy3, cyanine 3; IgG, immunoglobulin G, FITC, fluorescein
isothiocyanate. |
Discussion
Blocking of NMDA receptors by memantine is an
effective method for the treatment of AD; however, its effects on
neuronal autophagy and apoptosis in AD have remained to be
elucidated. The present study established an AD cell model that
overexpressed APP695. The results indicated that memantine
increased cell viability in the AD model. Memantine exerted its
anti-autophagic and anti-apoptotic functions via the mTOR-dependent
and mTOR-independent signaling pathways. These results
preliminarily explored the mechanisms underlying the
anti-autophagic effects of memantine. In addition, pre-treatment of
the AD cell model with the autophagy enhancer RAPA demonstrated
that autophagy promoted cell apoptosis. These results provided
experimental evidence regarding the controversial issue of the
association between autophagy and apoptosis.
An increase in autophagosome formation (28) and activation of caspases was
detected (29) in the neurons of
AD patients. Numerous studies have reported that Aβ accumulation
induced NMDA receptor overactivation (30,31),
whereas NMDA activation promoted neuronal autophagy or apoptosis.
Gao et al (32) treated rat
cardiomyocytes with the NMDA receptor antagonist MK-801, thereby
demonstrating that apoptosis in cardiomyocytes was significantly
improved following treatment, and that the expression levels of the
apoptosis-associated proteins Bcl-2-associated X protein and
caspase-3 decreased significantly. Therefore, these results led to
the speculation that memantine, which is an NMDA receptor
antagonist, may inhibit neuronal autophagy and apoptosis. The
present study established an AD cell model by upregulating APP695
expression and demonstrated that memantine increased the viability
of SH-SY5Y cells overexpressing APP695. In addition, 5 µM memantine
significantly inhibited cell autophagy and apoptosis. However,
these inhibitory effects were reversed by pre-incubation with
RAPA.
In AD research, the association between neuronal
autophagy and apoptosis is controversial. Nilsson et al
(19) reported that APP and its
hydrolytic enzymes were not degraded by autophagosomes following
the conversion of autophagosomes, thereby leading to the production
of large amounts of Aβ (17,19,20)
and an increase in cell apoptosis. Spilman et al (21) reported that enhancement of
autophagy reduced the levels of Aβ, thereby inhibiting apoptosis
(21–24). In the present study, AD model cells were
pre-treated with the autophagy enhancer RAPA and were compared with
cells in other treatment groups. The results demonstrated that the
enhancement of neuronal autophagy significantly increased the
apoptotic rates of the cells, which was concordant with the results
of Nilsson et al (19).
mTOR is an important regulator of autophagy, and the
mTOR-dependent signaling pathway is regulated by PI3K/Akt/mTOR.
Inhibition of this signaling pathway may activate cell autophagy
(33–36). In the mTOR-independent signaling pathway, Bcl-2 is
phosphorylated via the protein kinase JNK1 to separate Bcl-2 and
Beclin-1; autophagy is then activated by Beclin-1 (37). The present study demonstrated that
memantine was able to significantly upregulate the expression of
the signaling factors PI3K, Akt and mTOR, to activate the
mTOR-dependent autophagy signaling pathway, and to reduce the
autophagic capacity of neuronal cells. In addition, the present
study also detected that memantine inhibited the phosphorylation of
the Bcl-2/Beclin-1 complex by JNK1 to prevent the activation of
cell autophagy by Beclin-1. These results suggested that the two
signaling pathways were involved in the inhibition of cell
autophagy by memantine.
In conclusion, the results of the present study
demonstrated that memantine exerted significant anti-autophagic and
anti-apoptotic effects in an APP695-overexpressing cell model of
AD, the mechanism of which involved the mTOR-dependent as well
as-independent autophagic signaling pathways. The present study
supported the clinical use of memantine for the treatment of AD.
Furthermore, the present study confirmed that autophagy in the AD
cell model was able to promote cell apoptosis, thus providing
experimental evidence regarding the association between autophagy
and apoptosis.
References
|
1
|
Ballard C, Gauthier S, Corbett A, Brayne
C, Aarsland D and Jones E: Alzheimer's disease. Lancet.
377:1019–1031. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Huang Y and Mucke L: Alzheimer mechanisms
and therapeutic strategies. Cell. 148:1204–1222. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
De Felice FG, Velasco PT, Lambert MP,
Viola K, Fernandez SJ, Ferreira ST and Klein WL: Abeta oligomers
induce neuronal oxidative stress through an N- methyl-d- aspartate receptor- dependent
mechanism that is blocked by the Alzheimer drug memantine. J Biol
Chem. 282:11590–11601. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Texidó L, Martín-Satué M, Alberdi E,
Solsona C and Matute C: Amyloid β peptide oligomers directly
activate NMDA receptors. Cell Calcium. 49:184–190. 2011. View Article : Google Scholar
|
|
5
|
Zheng L, Kågedal K, Dehvari N, Benedikz E,
Cowburn R, Marcusson J and Terman A: Oxidative stress induces
macroautophagy of amyloid beta- protein and ensuing apoptosis. Free
Radic Biol Med. 46:422–429. 2009. View Article : Google Scholar
|
|
6
|
Lorenzi M, Beltramello A, Mercuri NB,
Mercuri NB, Canu E, Zoccatelli G, Pizzini FB, Alessandrini F,
Cotelli M and Rosini S: Effect of memantine on resting state
default mode network activity in Alzheimer's disease. Drugs Aging.
28:205–217. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Saxton J, Hofbauer RK, Woodward M,
Gilchrist NL, Potocnik F, Hsu HA, Miller ML, Pejović V, Graham SM
and Perhach JL: Memantine and functional communication in
Alzheimer's disease: Results of a 12- week, international,
randomized clinical trial. J Alzheimers Dis. 28:109–118. 2012.
|
|
8
|
Schulz JB, Rainer M, Klünemann HH, Kurz A,
Wolf S, Sternberg K and Tennigkeit F: Sustained effects of once -
daily memantine treatment on cognition and functional communication
skills in patients with moderate to severe Alzheimer's disease:
Results of a 16- week open- label trial. J Alzheimers Dis.
25:463–475. 2011.
|
|
9
|
Liu MY, Wang S, Yao WF, Zhang ZJ, Zhong X,
Sha L, He M, Zheng ZH and Wei MJ: Memantine improves spatial
learning and memory impairments by regulating NGF signaling in
APP/PS1 transgenic mice. Neuroscience. 273:141–151. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Danysz W and Parsons CG: The NMDA receptor
antagonist memantine as a symptomatological and neuroprotective
treatment for Alzheimer's disease: Preclinical evidence. Int J
Geriatr Psychiatry (Suppl 1). 18:S23–S32. 2003. View Article : Google Scholar
|
|
11
|
Peskind ER, Potkin SG, Pomara N, Ott BR,
Graham SM, Olin JT and McDonald S: Memantine treatment in mild to
moderate Alzheimer disease: A 24- week randomized, controlled
trial. Am J Geriatr Psychiatry. 14:704–715. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ray B, Banerjee PK, Greig NH and Lahiri
DK: Memantine treatment decreases levels of secreted Alzheimer's
amyloid precursor protein (APP) and amyloid beta (A beta) peptide
in the human neuroblastoma cells. Neurosci Lett. 470:1–5. 2010.
View Article : Google Scholar
|
|
13
|
Alley GM, Bailey JA, Chen D, Ray B, Puli
LK, Tanila H, Banerjee PK and Lahiri DK: Memantine lowers amyloid-
beta peptide levels in neuronal cultures and in APP/PS1 transgenic
mice. J Neurosci Res. 88:143–154. 2010. View Article : Google Scholar
|
|
14
|
Maekawa M, Namba T, Suzuki E, Yuasa S,
Kohsaka S and Uchino S: NMDA receptor antagonist memantine promotes
cell proliferation and production of mature granule neurons in the
adult hippocampus. Neurosci Res. 63:259–266. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Namba T, Maekawa M, Yuasa S, Kohsaka S and
Uchino S: The Alzheimer's disease drug memantine increases the
number of radial glia-like progenitor cells in adult hippocampus.
Glia. 57:1082–1090. 2009. View Article : Google Scholar
|
|
16
|
Namba T, Yabe T, Gonda Y, et al: Pigment
epithelium-derived factor upregulation induced by memantine, an
N-methyl-d-aspartate receptor antagonist, is involved in increased
proliferation of hippocampal progenitor cells. Neuroscience.
167:372–383. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ghavami S, Shojaei S, Yeganeh B, Ande SR,
Jangamreddy JR, Mehrpour M, Christoffersson J, Chaabane W, Moghadam
AR and Kashani HH: Autophagy and apoptosis dysfunction in
neuro-degenerative disorders. Prog Neurobiol. 112:24–49. 2014.
View Article : Google Scholar
|
|
18
|
Nixon RA: Autophagy, amyloidogenesis and
Alzheimer disease. J Cell Sci. 120:4081–4091. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nilsson P, Loganathan K, Sekiguchi M,
Matsuba Y, Hui K, Tsubuki S, Tanaka M, Iwata N, Saito T and Saido
TC: Aβ secretion and plaque formation depend on autophagy. Cell
Reports. 5:61–69. 2013. View Article : Google Scholar
|
|
20
|
Yu WH, Cuervo AM, Kumar A, Peterhoff CM,
Schmidt SD, Lee JH, Mohan PS, Mercken M, Farmery MR and Tjernberg
LO: Macroautophagy– a novel Beta- amyloid peptide- generating
pathway activated in Alzheimer's disease. J Cell Biol. 171:87–98.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Spilman P, Podlutskaya N, Hart MJ, Debnath
J, Gorostiza O, Bredesen D, Richardson A, Strong R and Galvan V:
Inhibition of mTOR by rapamycin abolishes cognitive deficits and
reduces amyloid- beta levels in a mouse model of Alzheimer's
disease. PLoS One. 5:e99792010. View Article : Google Scholar
|
|
22
|
Caccamo A, Majumder S, Richardson A,
Strong R and Oddo S: Molecular interplay between mammalian target
of rapamycin (mTOR), amyloid- beta, and Tau: Effects on cognitive
impairments. J Biol Chem. 285:13107–13120. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tan CC, Yu JT, Tan MS, Jiang T, Zhu XC and
Tan L: Autophagy in aging and neurodegenerative diseases:
Implications for pathogenesis and therapy. Neurobiol Aging.
35:941–957. 2014. View Article : Google Scholar
|
|
24
|
Zhu XC, Yu JT, Jiang T and Tan L:
Autophagy modulation for Alzheimer's disease therapy. Mol
Neurobiol. 48. pp. 702–714. 2013, View Article : Google Scholar
|
|
25
|
Ye X, Tai W, Bao X, Chen X and Zhang D:
FLZ inhibited γ- secretase selectively and decreased Aβ
mitochondrial production in APP- SH- SY5Y cells. Naunyn
Schmiedebergs Arch Pharmacol. 387:75–85. 2014. View Article : Google Scholar
|
|
26
|
Shen YE, Wang Y, Yu GC, Liu C, Zhang ZY
and Zhang LM: Effects of edaravone on amyloid-β precursor protein
processing in SY5Y- APP695 cells. Neurotox Res. 24:139–147. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang L, Zhang D, Yu Y, Guan H, Qiao C and
Shang T: RNA interference- mediated silencing of laminin receptor 1
(LR1) suppresses migration and invasion and downregulates matrix
metalloproteinase (MMP)- 2 and MMP- 9 in trophoblast cells:
Implication in the pathogenesis of preeclampsia. J Mol Histol.
44:661–668. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nixon RA, Wegiel J, Kumar A, Yu WH,
Peterhoff C, Cataldo A and Cuervo AM: Extensive involvement of
autophagy in Alzheimer disease: An immunoelectron microscopy study.
J Neuropathol Exp Neurol. 64:113–122. 2005.PubMed/NCBI
|
|
29
|
Louneva N, Cohen JW, Han LY, albot K,
Wilson RS, Bennett DA, Trojanowski JQ and Arnold SE: Caspase- 3 is
enriched in post-synaptic densities and increased in Alzheimer's
disease. Am J Pathol. 173:1488–1495. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kaufman AM, Milnerwood AJ, Sepers MD,
Coquinco A, She K, Wang L, Lee H, Craig AM, Cynader M and Raymond
LA: Opposing roles of synaptic and extrasynaptic NMDA receptor
signaling in cocultured striatal and cortical neurons. J Neurosci.
32:3992–4003. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Caballero B and Coto- Montes A: An insight
into the role of autophagy in cell responses in the aging and
neurodegenerative brain. Histol Histopathol. 27:263–275.
2012.PubMed/NCBI
|
|
32
|
Gao X, Xu X, Pang J, Zhang C, Ding JM,
Peng X, Liu Y and Cao JM: NMDA receptor activation induces
mitochondrial dysfunction, oxidative stress and apoptosis in
cultured neonatal rat cardiomyocytes. Physiol Res. 56:559–569.
2007.
|
|
33
|
Han W, Pan H, Chen Y, Sun J, Wang Y, Li J,
Ge W, Feng L, Lin X, Wang X, et al: EGFR tyrosine kinase inhibitors
activate autophagy as a cytoprotective response in human lung
cancer cells. PLoS One. 6:e186912011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu J, Hu XJ, Jin B, Qu XJ, Hou KZ and Liu
YP: β- Elemene induces apoptosis as well as protective autophagy in
human non- small- cell lung cancer A549 cells. J Pharm Pharmacol.
64:146–153. 2012. View Article : Google Scholar
|
|
35
|
Wang P, Guo QS, Wang ZW and Qian HX: HBx
induces HepG- 2 cells autophagy through PI3K/Akt- mTOR pathway. Mol
Cell Biochem. 372:161–168. 2013. View Article : Google Scholar
|
|
36
|
Yang YH, Chen K, Li B, Chen JW, Zheng XF,
Wang YR, Jiang SD and Jiang LS: Estradiol inhibits osteoblast
apoptosis via promotion of autophagy through the ER- ERK- mTOR
pathway. Apoptosis. 18:1363–1375. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Russell RC, Tian Y, Yuan H, Park HW, Chang
YY, Kim J, Kim H, Neufeld TP, Dillin A and Guan KL: ULK1 induces
autophagy by phosphorylating Beclin- 1 and activating VPS34 lipid
kinase. Nat Cell Biol. 15:741–750. 2013. View Article : Google Scholar : PubMed/NCBI
|