Introduction
Exposure to ultraviolet (UV) light, particularly
UV-B (290–320 nm), is a major etiological factor that contributes
to skin damage (1). Intense and
chronic UV exposure is associated with sunburn, immune suppression,
DNA damage, photoaging, and skin cancer (2,3). UV
can be directly absorbed by cellular macromolecules, thus leading
to photochemical modification of DNA and proteins (4). In addition, UV induces the generation
of reactive oxygen species (ROS), which overwhelm the antioxidant
defense system, resulting in oxidative stress and damage to cell
structure, DNA, proteins and lipids (5).
UV-B-induced DNA damage can trigger cell signaling
pathways to activate defense systems, leading to DNA repair and/or
apoptosis (6). Among these
cell-signaling proteins, activation of p53 is important in cells
with UV radiation-induced DNA damage (4,7). In
addition, oxidative stress is associated with UV-induced activation
of p53 (4). Pretreatment of cells
with N-acetyl-L-cysteine (NAC), an agent known to counteract
oxidative stress, attenuates the cellular p53 response to UV
(4). Mitogen-activated protein
kinases (MAPKs) are also involved in the response to UV radiation
(8); p38 acts as a
stress-activated MAPK that is preferentially activated by diverse
cellular stressors, including UV-B-induced oxidative stress
(9). It has previously been
reported that p38 mediates UV-induced phosphorylation of p53.
Pretreatment of cells with SB202190, a p38 inhibitor, may inhibit
the DNA binding activity of p53 and suppress p53-dependent
transcription (10). These
previous results indicated that the ROS-p38-p53 pathways are
involved in UV-induced cell damage.
UV-B-induced skin damage may be prevented by
avoidance of intense sun exposure, as well as topical and systemic
administration of antioxidants (11). Peony root (Radix Paeoniae Alba) is
a well-known herb, which has been used in Chinese medicine for
>1,200 years (12). Several
compounds, including paeoniflorin (PF), albiflorin, oxypaeoniflorin
and benzoylpaeoniflorin, are reported to be contained within peony
root (13). Among them, PF is
considered to be one of the major bioactive components (13). PF has been reported to exert
anti-inflammatory (12),
antioxidative (14) and antitumor
effects (15). Extracts of peony
root have previously been shown to exert protective effects against
UV-induced DNA damage and facial wrinkles in human skin (16). However, the anti-UV effects of
purified PF have yet to be elucidated.
The present study evaluated the protective effects
of PF on UV-induced skin damage in vitro, and demonstrated
that the effects were mediated via the ROS-p38-p53 pathway.
Materials and methods
Reagents and cell culture
PF (purity>99.8%; Fig. 1A) was purchased from Chengdu
Mansite Pharmaceutical Co., Ltd. (Chengdu, China). The HaCaT human
keratinocyte cells were purchased from the China Center for Type
Culture Collection (Wuhan, China), and were cultured in Dulbecco's
modified Eagle's medium (DMEM; Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 4 mM
L-glutamine, 3.7 g/l sodium bicarbonate, 4.5 g/l glucose and 10%
fetal bovine serum (Invitrogen; Thermo Fisher Scientific, Inc.).
Cells were maintained in a humidified incubator containing 5%
CO2 at 37°C. WST-8 was obtained from Dojindo Molecular
Technologies, Inc. (Kumamoto, Japan). Propidium iodide (PI) and
RNase A were purchased from Beyotime Institute of Biotechnology
(Haimen, China). Antibodies against phosphorylated (P)-p38 (cat.
no. 4511), p38 (cat. no. 8690), P-p53 (cat. no. 9284), p53 (cat.
no. 2524), caspase 3 (cat. no. 9662), cleaved caspase 3 (cat. no.
9664) and β-actin (cat. no. 8457) were obtained from Cell Signaling
Technology, Inc. (Danvers, MA, USA). NAC and SB203580 were
purchased from Sigma-Aldrich (St. Louis, MO, USA).
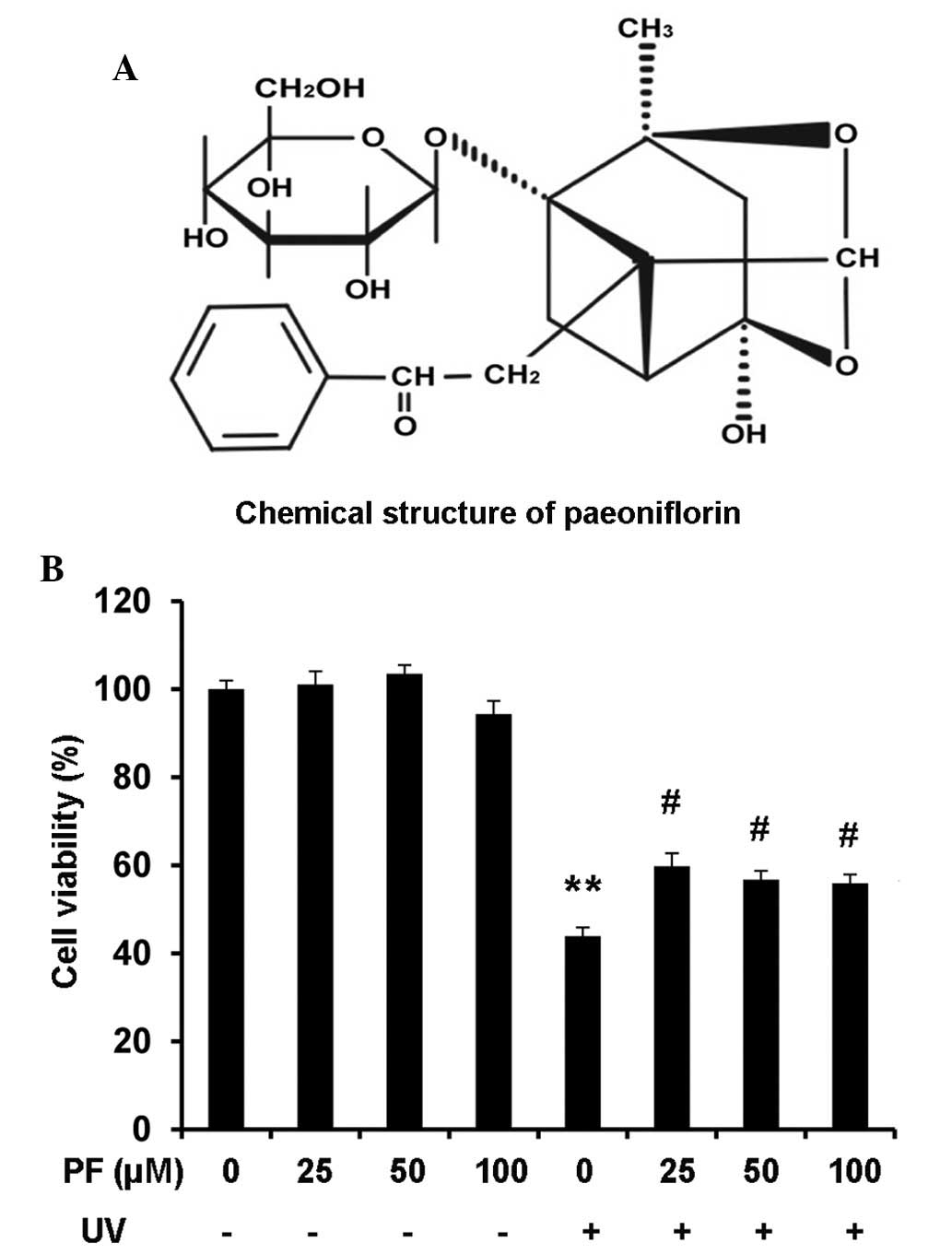 | Figure 1PF attenuated UV-B-induced decreased
cell viability in HaCaT cells. (A) Chemical structure of PF. (B)
Cell viability of HaCaT cells as measured by the WST-8 assay. Cells
were pretreated with PF (0, 25, 50, and 100 µM) for 2 h.
Following UV-B (60 mJ/cm2) radiation, the cells were
incubated for 24 h, after which, 10 µl WST-8 was added to
each well for 1 h. Optical density was measured at 450 nm using a
plate reader. Data are presented as the mean ± standard deviation.
**P<0.01, compared with the medium control group;
#P<0.01, compared with UV-B exposure alone. PF,
paeoniflorin; UV, ultraviolet. |
UV radiation
HaCaT cells were pretreated with various
concentrations of PF (0, 25, 50 and 100 µM) for 2 h in
serum-free medium. Subsequently, the cells were washed with
phosphate-buffered saline (PBS) and covered with a thin layer of
PBS. UV-B radiation was applied at a level of 60 mJ/cm2
using a Philips TL-D/08 15W weathering lamp (wavelength 290–340 nm,
peak 311 nm) (Philips, Amsterdam, The Netherlands). The radiation
intensity was monitored using a UV-B radiometer (Beijing Normal
University, Beijing, China). Following exposure to UV-B radiation,
PBS was removed and DMEM containing PF (0, 25, 50 or 100 µM)
was added to the cells, which were incubated at 37°C for a suitable
period.
Cell viability assays
PF [dissolved in dimethyl sulfoxide (DMSO)] was used
to treat the cells. The final concentration of DMSO used was
<0.1% (v/v). Cell viability was measured using the WST-8 assay
(Dojindo Molecular Technologies, Inc.), according to the
manufacturer's protocol. Briefly, HaCaT cells were seeded at a
density of 5×103 cells/well in 96-well culture plates in
DMEM, and were cultured in a humidified incubator at 37°C
overnight. The cells were pretreated with PF (0, 25, 50, and 100
µM) for 2 h. Following UV-B radiation, the cells were
incubated for a further 24 h, after which, 10 µl WST-8 was
added to each well for 1 h. Subsequently, optical density (OD) was
measured at a wavelength of 450 nm using a BIO-TEK MQX200 plate
reader (Highland Park Efficiencies, Winooski, VT, USA). The
percentage of viable cells was determined using the following
formula: Ratio (%) = [OD(PF)-OD(Blank)/OD(Control)-OD (Blank)] ×
100. Cell viability data are averages of three independent
experiments each containing six replicates.
Cell death analysis
HaCaT cells were seeded at a density of
2×105 cells/well in 6-well culture plates in DMEM, and
were cultured in a humidified incubator at 37°C for 24 h. The cells
were pretreated with 25 µM PF for 2 h. Following UV-B
radiation, the cells were incubated for a further 24 h, after
which, cells were collected and fixed in 70% ethanol for 24 h at
4°C. Subsequently, the cells were centrifuged at 300 × g for 5 min
and the cell pellet was resuspended in 400 µl PBS containing
RNase A (10 mg/ml, 50 µl) and PI (2 mg/ml, 10 µl).
The mixture was incubated in the dark at 37°C for 30 min and data
were acquired using a FACSCalibur flow cytometer (BD Biosciences,
San Joe, CA, USA). The cell death data were analyzed using FlowJo
software V6.0 (Tree Star, Inc., Ashland, OR, USA). The extent of
cell death was determined by evaluating the sub G1
fraction, or the percentage of cells with DNA content <2n. The
experiment was replicated three times.
Western blot analysis
HaCaT cells were seeded at a density of
5×105 cells/well in 6-well culture plates in DMEM, and
were cultured in a humidified incubator at 37°C for 24 h. The cells
were pretreated with 25 µM PF or 5 µM SB203580 for 2
h. Following UV-B radiation, the cells were incubated for a further
24 h, after which, cells were collected and resuspended in lysis
buffer [150 mmol/l NaCl, 1% NP-40, 0.5% sodium deoxycholate, 0.1%
sodium dodecyl sulfate (SDS), 50 mmol/l Tris-Cl, pH 8.0] containing
2 µg/ml aprotinin, 2 µg/ml leupeptin 40 mg/ml
phenylmethylsulfonyl fluoride and 2 mmol/l DTT. The cells were
centrifuged at 12,000 × g for 15 min, in order to remove nuclei and
cell debris. Supernatants were then immediately frozen at −80°C,
until further use. Protein concentrations were determined using the
Bradford Assay (Bio-Rad Laboratories, Inc., Hercules, CA, USA) and
30 µg cellular proteins were separated by 10%
SDS-polyacrylamide gel electrophoresis followed by electro-blotting
onto polyvinylidene difluoride membranes (45 µm; EMD
Millipore, Billerica, MA, USA). The membranes were blocked for 1 h
with 5% milk at room temperature, followed by an overnight
incubation at 4°C with the following primary antibodies: P-p38,
P-p53, p38, p53, cleaved caspase 3, caspase 3 and β-actin (all
1:1,000 dilution). Blots were washed twice with 0.1% Tween
20/Tris-buffered saline (TTBS) prior to incubation with a
horseradish peroxidase-conjugated secondary antibody (1:1,000
dilution; Cell Signaling Technology, Inc.) for 1 h at room
temperature. Blots were further washed with TTBS and were developed
by enhanced chemiluminescence using Supersignal West Femto
Chemiluminescent Substrate (Pierce; Thermo Fisher Scientific,
Inc.). Band intensities were quantified using UN-SCAN-IT Gel
Analysis software (version 6; Silk Scientific, Inc., Orem, UT,
USA). The OD for target proteins was indicated as a proportion of
β-actin OD. Western blotting was replicated three times.
Evaluation of ROS
ROS were detected using the cell-permeable
fluorescent probe 2,-7,-dichlorofluorescein-diacetate
(H2DCFDA) (Sigma-Aldrich), a non-fluorescent compound,
which is converted into highly fluorescent dichlorofluorescein
(DCF) by cellular peroxides. The cells were exposed to 25 µM
PF or 2 mM NAC for 2 h. Following radiation, the cells were
incubated for a further 6 h, after which, the cells were treated
with H2DCFDA (10 µM) in serum-free DMEM.
Following incubation at 37°C for 30 min, the cells were washed with
PBS and fluorescence was monitored by flow cytometry or
fluorescence microscopy (DM1000; Leica Camera, Wetzlar, Germany).
The mean fluorescence intensity (MFI) data were analyzed by FlowJo
software V6.0 (Tree Star, Inc.). The MFI data were replicated three
times.
Statistical analysis
All data are presented as the mean ± standard
deviation. Data analysis was performed using SPSS version 20.0 (IBM
SPSS, Amronk, NY, USA). Statistical significance was detected using
one-way analysis of variance. For comparisons between two groups,
Student's t-test was used. P<0.05 was considered to indicate a
statistically significant difference.
Results
PF attenuates UV-B-induced decreased cell
viability in HaCaT cells
Cell viability was evaluated using the WST-8 assay.
Exposure of HaCaT cells to UV-B resulted in a significant reduction
in the percentage of viable cells (45%) at 24 h, as compared with
the medium control group (P<0.01). Treatment with PF (25, 50 or
100 µM) alone did not affect cell viability; however, cells
pretreated with PF (25, 50, and 100 µM) prior to UV-B
radiation exhibited a significant increase in the percentage of
viable cells (60%). No dose-dependent effects were observed
following PF treatment (Fig.
1B).
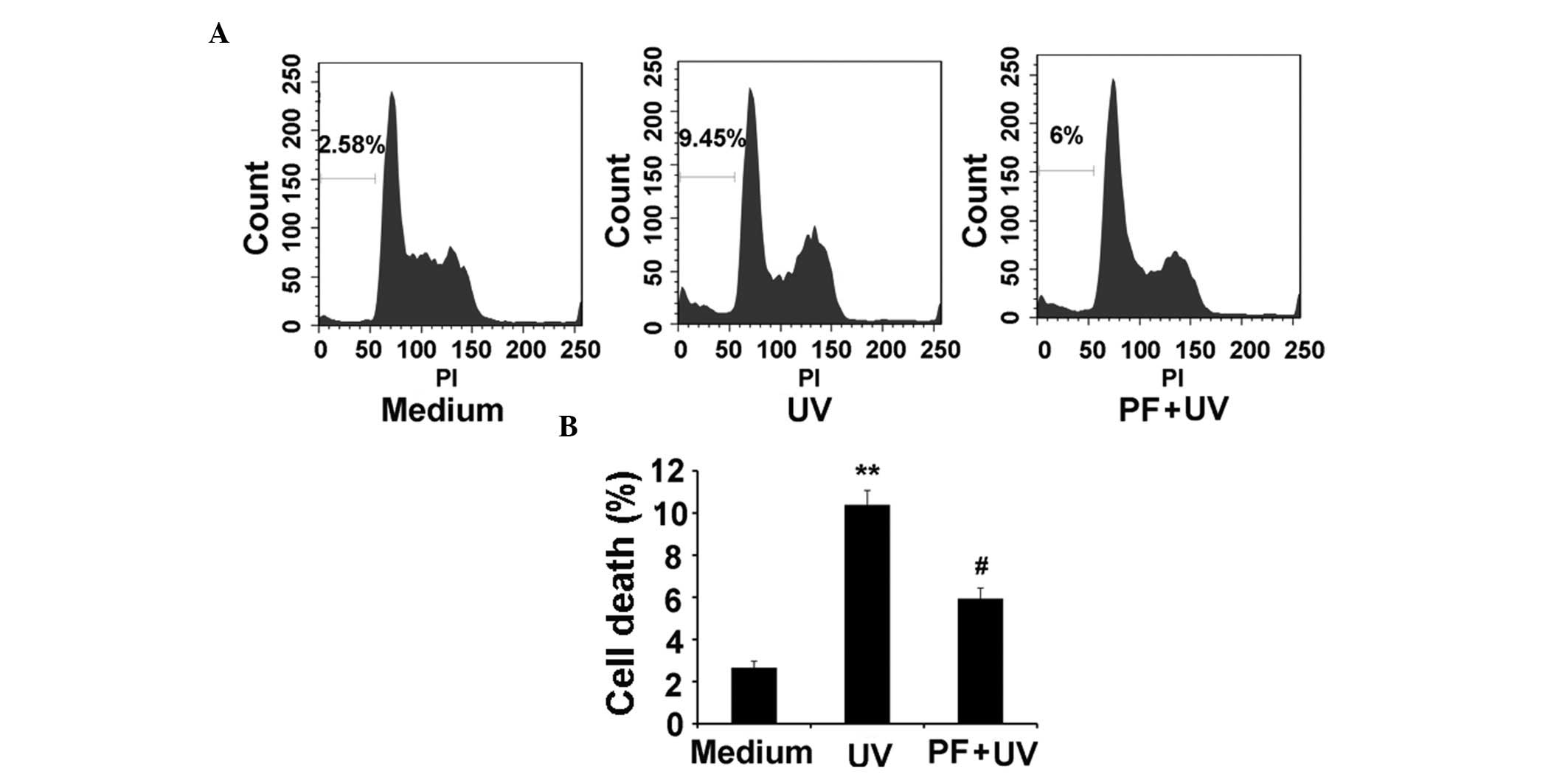
PF reduces UV-B-induced cell death in
HaCaT cells
It is well known that UV-B exposure induces
apoptosis. As demonstrated by flow cytometry, UV-B (60
mJ/cm2) radiation significantly increased cell death
(9.45%) at 24 h, as compared with in the medium control group
(2.58%), whereas PF pretreatment markedly reduced UV-B-induced cell
death (6%) (Fig. 2A and B).
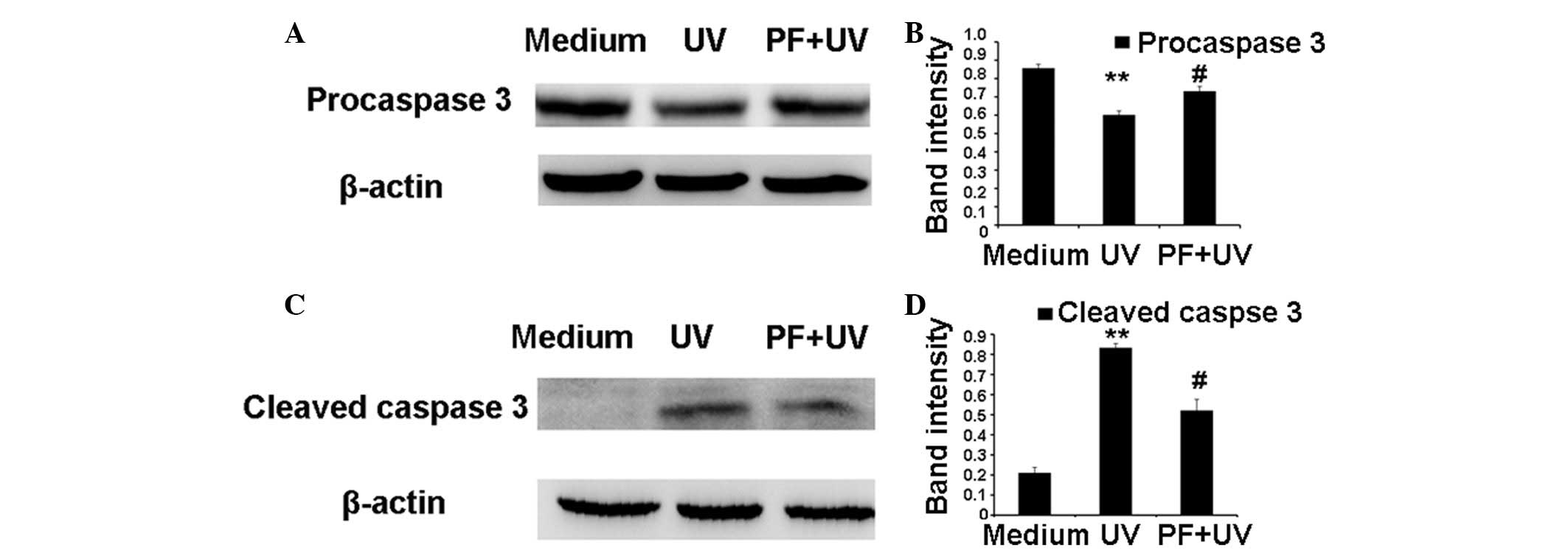
Caspase 3 is expressed in an inactive pro-form, procaspase 3. In
apoptosis, procaspase 3 is activated and generates two active
subunits, cleaved caspase 3 (17).
As shown in Fig. 3, UV radiation
resulted in a significant decrease in the expression levels of
procaspase 3 and an increase in cleaved caspase 3 expression. PF
pretreatment partially counteracted the effects of UV-B on
procaspase 3 and cleaved caspase 3 expression.
PF inhibits the production of ROS after
UV-B radiation
UV-B exposure results in the generation of ROS,
which induce cell damage. The MFI of DCF was used to evaluate the
production of ROS. As shown in Fig.
4, ROS production was significantly increased following UV-B
radiation. Pretreatment with 25 µM PF or 2 mM NAC (a ROS
scavenger), markedly inhibited the production of ROS after UV-B
radiation.
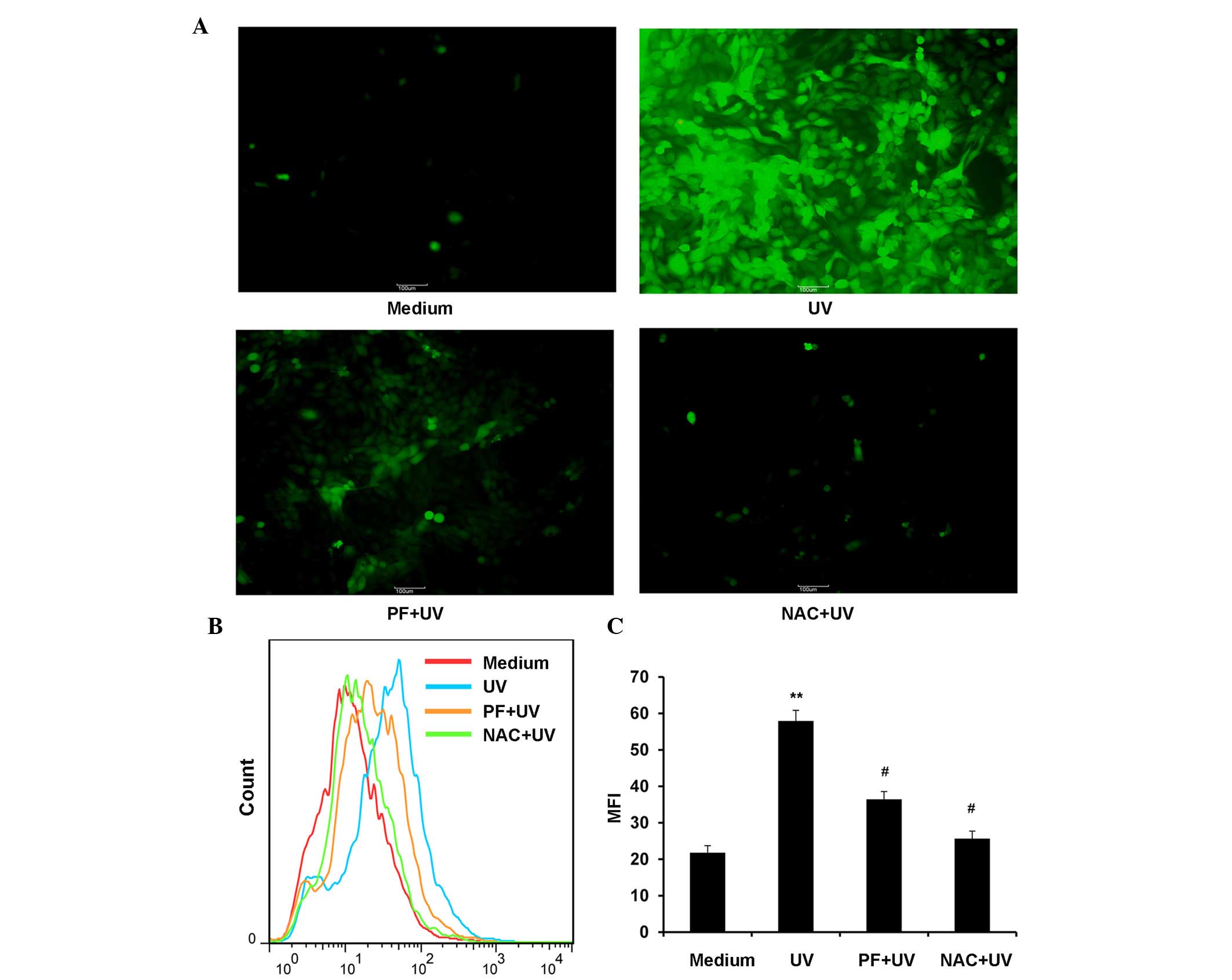 | Figure 4PF inhibited the production of ROS
after UV-B radiation. For the ROS assay, cells were exposed to 25
µM PF or 2 mM NAC for 2 h. After UV-B (60 mJ/cm2)
radiation, the cells were incubated for 6 h and were then treated
with 2,-7,-dichlorofluorescein-diacetate (10 µM). Following
incubation at 37°C for 30 min, the cells were washed with
phosphate-buffered saline and fluorescence was monitored by flow
cytometry or fluorescence microscopy. MFI data were analyzed using
FlowJo software V6.0. (A) Representative fluorescence microscopy
image of ROS (magnification ×100). (B) Representative image of ROS
MFI. (C) Statistical analysis of ROS MFI. Data are presented as the
mean ± standard deviation. **P<0.01, compared with
the medium control group, #P<0.01, compared with UV-B
exposure alone. PF, paeoniflorin; ROS, reactive oxygen species; UV,
ultraviolet; NAC, N-acetyl-L-cysteine; MFI, mean fluorescence
intensity. |
PF inhibits UV-B-induced activation of
p38 and p53 in HaCaT cells
The activation of p38 and p53 is associated with
UV-B-induced cell damage. Western blotting (Fig. 5) demonstrated that UV-B radiation
significantly increased the expression levels of P-p38 and P-p53.
Conversely, PF pretreatment markedly reduced the expression levels
of P-p38 and P-p53 following UV-B-radiation. Both UV radiation and
PF pretreatment did not alter the expression levels of total p38
and p53. It has previously been reported that p38 kinase mediates
UV-induced phosphorylation of p53 protein (10). As shown in Fig. 5C and D, SB203580, a p38 inhibitor,
significantly reduced the expression levels of P-p53 after UV-B
radiation.
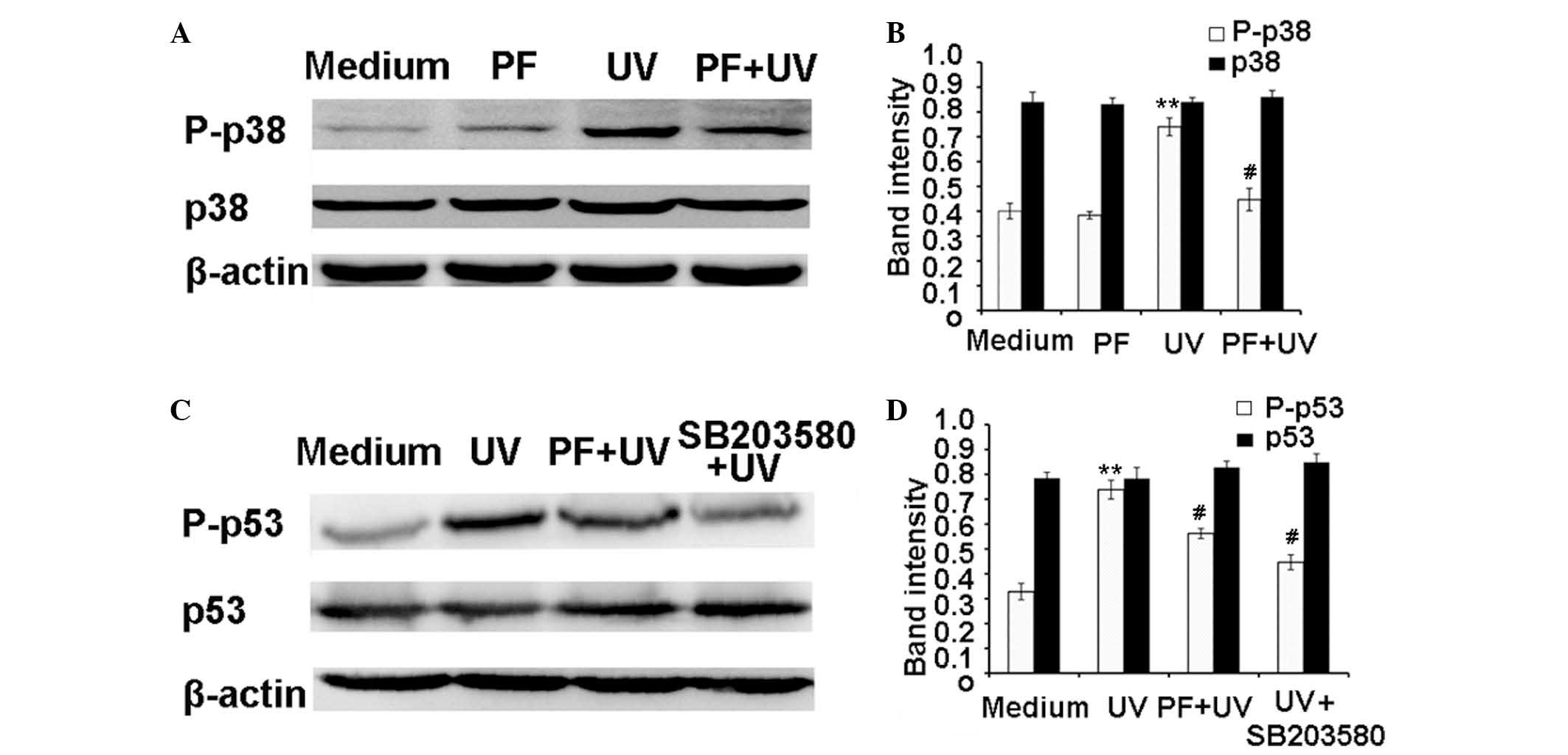 | Figure 5PF inhibited UV-B induced activation
of p38 and p53. For western blotting, the cells were pretreated
with 25 µM PF or 5 µM SB203580 for 2 h. Following
UV-B radiation, the cells were incubated for 24 h. Total protein
was extracted, and P-p38, p38, P-p53 and p53 expression levels were
detected by western blot analysis. β-actin was used as a loading
control. (A) Representative western blot of P-p38 and p38. (B)
Quantification of band intensities of P-p38 and p38. (C)
Representative western blot of P-p53 and p53. (D) Quantification of
band intensities of P-p53 and p53. Band intensities were quantified
using UN-SCAN-IT gel analysis software. Data are presented as the
mean ± standard deviation. **P<0.01, compared with
the medium control; # P<0.01, compared with UV-B
exposure alone. PF, paeoniflorin; UV, ultraviolet; P,
phosphorylated. |
Discussion
UV radiation is one of the most harmful
environmental factors that contribute to skin damage (18). UV exposure induces extensive
generation of ROS (19), which
react with DNA, proteins and fatty acids, resulting in photoaging
and skin cancer (20). One
approach to protecting human skin against UV radiation is the use
of antioxidants (21,22). In recent years, naturally occurring
herbal compounds have gained considerable attention as protective
agents for UV exposure (19,23–25).
PF is a novel natural antioxidant, which is isolated
from peony root (26,27). The present study evaluated the
protective effects of PF on UV-induced skin damage. The results
demonstrated that treatment with PF significantly increased the
percentage of viable keratinocytes following UV-B exposure. It is
well known that UV-B exposure induces apoptosis, and caspase 3 has
an important role in the execution of apop-tosis (17). Cell death analysis in the present
study revealed that PF treatment markedly reduced UV-B
radiation-induced apop-tosis in keratinocytes, which was
accompanied by increased procaspase 3 expression and decreased
cleaved caspase 3 expression, as compared with the UV-B radiation
group.
The ROS-p38-p53 pathway is involved in UV-B-induced
skin damage; oxidative stress induced by ROS can initiate MAPK
signaling by phosphorylation of MAPK proteins. p38 acts as a
stress-activated MAPK that is preferentially activated by
UV-generated ROS (9). In the
present study, treatment with PF substantially reduced the
production of ROS following exposure to UV. The UV-induced
phosphorylation of p38 was also significantly inhibited by PF.
Activation of p53 is important in cells with DNA damage caused by
UV radiation (4,7). It has previously been reported that
p38 mediates UV-induced phosphorylation of p53 (10). In the present study, treatment with
PF significantly reduced the production of ROS, and inhibited the
activation of p38 and p53 in human keratinocytes. In addition, NAC,
a ROS scavenger, and SB203580, a p38 inhibitor, significantly
inhibited UV-B-induced decreased cell viability in human
keratinocytes (data not shown). These results further demonstrate
that the ROS-p38-p53 pathway is involved in UV-induced skin
damage.
Numerous studies suggest that the activation of
nuclear factor-E2-related factor 2 (Nrf2) protects cutaneous
keratinocytes and fibroblasts against the cytotoxic effects of UV
(28–31). Nrf2 has emerged as a promising
molecular target for the pharmacological prevention of skin damage
caused by solar UV exposure. PF has been reported to protect
radiation-induced pulmonary endothelial cells injury through the
Nrf2 pathway (32). Further
studies are, therefore, required in order to investigate the role
of Nrf2 in the anti-UV effects of PF.
In conclusion, the present study reported that PF is
able to attenuate UV-B-induced cell damage in human keratinocytes.
Notably, the present study confirmed that these effects were
mediated, at least in part, via inhibition of the ROS-p38-p53
pathway. Further studies are required to investigate the in
vivo anti-UV effects of PF.
Acknowledgments
The present study was funded by grants from the
National Natural Science Foundation of China (no. 81102541),
Shanghai Municipal Commission of Health and Family Planning (grant
no. 201440336) and the Scientific Research Project supported by
Huashan Hospital, Fudan University (no. 2014QD09).
References
|
1
|
de Gruijl FR: Photocarcinogenesis: UVA vs.
UVB radiation. Skin Pharmacol Appl Skin Physiol. 15:316–320. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tyrrell RM: Ultraviolet radiation and free
radical damage to skin. Biochem Soc Symp. 61:47–53. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Johnson BE: Formation of thymine
containing dimers in skin exposed to ultraviolet radiation. Bull
Cancer. 65:283–297. 1978.PubMed/NCBI
|
|
4
|
Renzing J, Hansen S and Lane DP: Oxidative
stress is involved in the UV activation of p53. J Cell Sci.
109:1105–1112. 1996.PubMed/NCBI
|
|
5
|
Jurkiewicz BA and Buettner GR: Ultraviolet
light-induced free radical formation in skin: An electron
paramagnetic resonance study. Photochem Photobiol. 59:1–4. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Darzynkiewicz Z, Zhao H, Halicka HD, Rybak
P, Dobrucki J and Wlodkowic D: DNA damage signaling assessed in
individual cells in relation to the cell cycle phase and induction
of apoptosis. Crit Rev Clin Lab Sci. 49:199–217. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lee CH, Wu SB, Hong CH, Yu HS and Wei YH:
Molecular mechanisms of UV-induced apoptosis and its effects on
skin residential cells: The implication in UV-based phototherapy.
Int J Mol Sci. 14:6414–6435. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bae JY, Choi JS, Choi YJ, Shin SY, Kang
SW, Han SJ and Kang YH: (−)Epigallocatechin gallate hampers
collagen destruction and collagenase activation in
ultraviolet-B-irradiated human dermal fibroblasts: Involvement of
mitogen-activated protein kinase. Food Chem Toxicol. 46:1298–1307.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Johnson GL and Lapadat R:
Mitogen-activated protein kinase pathways mediated by ERK, JNK, and
p38 protein kinases. Science. 298:1911–1912. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang C, Ma WY, Maxiner A, Sun Y and Dong
Z: p38 kinase mediates UV-induced phosphorylation of p53 protein at
serine 389. J Biol Chem. 274:12229–12235. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ganceviciene R, Liakou AI, Theodoridis A,
Makrantonaki E and Z ouboulis CC: Skin anti-aging strategies.
Dermatoendocrinol. 4:308–319. 2012. View Article : Google Scholar
|
|
12
|
He DY and Dai SM: Anti-inflammatory and
immunomodulatory effects of Paeonia lactiflora pall, a traditional
Chinese herbal medicine. Front Pharmacol. 2:102011. View Article : Google Scholar
|
|
13
|
Kim SH, Lee MK, Lee KY, Sung SH, Kim J and
Kim YC: Chemical constituents isolated from Paeonia lactiflora
roots and their neuroprotective activity against oxidative stress
in vitro. J Enzyme Inhib Med Chem. 24:1138–1140. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wankun X, Wenzhen Y, Min Z, Weiyan Z, Huan
C, Wei D, Lvzhen H, Xu Y and Xiaoxin L: Protective effect of
paeoniflorin against oxidative stress in human retinal pigment
epithelium in vitro. Mol Vis. 17:3512–3522. 2011.
|
|
15
|
Wang H, Zhou H, Wang CX, Li YS, Xie HY,
Luo JD and Zhou Y: Paeoniflorin inhibits growth of human colorectal
carcinoma HT 29 cells in vitro and in vivo. Food Chem Toxicol.
50:1560–1567. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee S, Lim JM, Jin MH, Park HK, Lee EJ,
Kang S, Kim YS and Cho WG: Partially purified paeoniflorin exerts
protective effects on UV-induced DNA damage and reduces facial
wrinkles in human skin. J Cosmet Sci. 57:57–64. 2006.PubMed/NCBI
|
|
17
|
Bratton SB, MacFarlane M, Cain K and Cohen
GM: Protein complexes activate distinct caspase cascades in death
receptor and stress-induced apoptosis. Exp Cell Res. 256:27–33.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kligman LH: Prevention and repair of
photoaging: Sunscreens and retinoids. Cutis. 43:458–465.
1989.PubMed/NCBI
|
|
19
|
Erden Inal M, Kahraman A and Köken T:
Beneficial effects of quercetin on oxidative stress induced by
ultraviolet A. Clin Exp Dermatol. 26:536–539. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Goihman-Yahr M: Skin aging and photoaging:
An outlook. Clin Dermatol. 14:153–160. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tebbe B: Relevance of oral supplementation
with antioxidants for prevention and treatment of skin disorders.
Skin Pharmacol Appl Skin Physiol. 14:296–302. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Afaq F and Mukhtar H: Photochemoprevention
by botanical antioxidants. Skin Pharmacol Appl Skin Physiol.
15:297–306. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Katiyar SK and Elmets CA: Green tea
polyphenolic antioxidants and skin photoprotection (Review). Int J
Oncol. 18:1307–1313. 2001.PubMed/NCBI
|
|
24
|
Widyarini S, Spinks N, Husband AJ and
Reeve VE: Isoflavonoid compounds from red clover (Trifolium
pratense) protect from inflammation and immune suppression induced
by UV radiation. Photochem Photobiol. 74:465–470. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Svobodová A, Psotová J and Walterová D:
Natural phenolics in the prevention of UV-induced skin damage. A
review. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub.
147:137–145. 2003. View Article : Google Scholar
|
|
26
|
Chen T, Guo ZP, Jiao XY, Zhang YH, Li JY
and Liu HJ: Protective effects of peoniflorin against hydrogen
peroxide-induced oxidative stress in human umbilical vein
endothelial cells. Can J Physiol Pharmacol. 89:445–453. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhong SZ, ma SP and Hong ZY: Peoniflorin
activates Nrf2/ARE pathway to alleviate the Abeta (1–42)-induced
hippocampal neuron injury in rats. Yao Xue Xue Bao. 48:1353–1357.
2013.In Chinese. PubMed/NCBI
|
|
28
|
Saw CL, Huang MT, Liu Y, Khor TO, Conney
AH and Kong AN: Impact of Nrf2 on UVB-induced skin
inflammation/photo-protection and photoprotective effect of
sulforaphane. Mol Carcinog. 50:479–486. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Schafer M and Werner S: Nrf2 - A regulator
of keratinocyte redox signaling. Free Radic Biol Med. 88:243–252.
2015. View Article : Google Scholar
|
|
30
|
Dinkova-Kostova AT, Jenkins SN, Fahey JW,
Ye L, Wehage SL, Liby KT, Stephenson KK and Wade KL: Protection
against UV-light-induced skin carcinogenesis in SKH-1 high-risk
mice by sulforaphane-containing broccoli sprout extracts. Cancer
Lett. 240:243–252. 2006. View Article : Google Scholar
|
|
31
|
Wondrak GT, Cabello CM, Villeneuve NF,
Zhang S, Ley S, Li Y, Sun Z and Zhang DD: Cinnamoyl-based
Nrf2-activators targeting human skin cell photo-oxidative stress.
Free Radic Biol Med. 45:385–395. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yu J, Zhu X, Qi X, Che J and Cao B:
Paeoniflorin protects human EA. hy926 endothelial cells against
gamma-radiation induced oxidative injury by activating the
NF-E2-related factor 2/heme oxygenase-1 pathway. Toxicol Lett.
218:224–234. 2013. View Article : Google Scholar : PubMed/NCBI
|