Introduction
Primary graft dysfunction due to
ischemia/reperfusion injury is a major cause of cardiac dysfunction
and high mortality (1,2). Mitochondrial dysfunction serves a
crucial role in the pathogenesis of several cardiac diseases
including ischemia/reperfusion injury. In response to stress,
mitochondria are able to rapidly shut off the energy supply,
produce vast amounts of toxic oxygen species and release a mixture
of death-inducing proteins into the cellular milieu (3). Mitochondrial uncoupling proteins
(UCPs), which promote proton leak across the inner mitochondrial
membrane, have emerged as essential regulators of mitochondrial
membrane potential, respiratory activity and reactive oxygen
species (ROS) generation. UCP2 is able to reduce mitochondrial ROS
generation and thereby ameliorate myocardial function by promoting
mild uncoupling (4). UCPs
contribute to the elevation of cardiomyocyte tolerance against
hypoxia and re-oxygenation, and promote cellular resistance to
oxidative stress (4). UCP2 levels
have been reported to be upregulated in parallel with infarct size
reduction in preconditioned hearts (5). UCP2 gene expression is increased in
the left ventricle in response to chronic hypobaric hypoxia
(6). In the cytoprotective
hierarchy, UCP2 appears to serve a role in modulating
ischemia/anoxia tolerance in heart-derived cells. Therefore,
mitochondrial UCPs are necessary components of ischemia tolerance
and function as components of the cellular antioxidant defense
program.
The pathophysiological role of UCP2 on influencing
post-operative outcomes following heart transplantation remains to
be fully elucidated. Considering its pivotal role in the
pathogenesis of the organs post-ischemia/reperfusion injury,
further investigation on cardiac UCP2 activity and regulation will
facilitate the development of novel and more efficient organ
preservation. In the present study, the role of UCP2 following
hypothermic preservation in rat hearts was investigated using the
Langendorff perfusion system. Due to the fact that SIRT1 is an
important regulators of UCP2 expression in numerous physiological
and pathophysiological conditions, the role of SIRT1 in the
alterations of UCP2 expression was additionally investigated in the
current study.
Materials and methods
Animals
Male Sprague-Dawley rats (n=88; age, 8–10 weeks;
weight, 250–300 g) were purchased from the Experimental Animal
Center of Zhejiang University (Hangzhou, China) and cared for in
compliance with the Guide for the Care and Use of Laboratory
Animals published by the National Institutes of Health (7). Rats were housed at five rats per cage
in a controlled environment at 21–23 °C, with 40–60% humidity and
12/12-h light/dark cycle. The rats were allowed free access to
water and food. All experimental protocols were approved by the
Ethics Committee on Animal Experimentation of Zhejiang
University.
Reagents
Genipin, resveratrol, EX-527, ethylene
glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid (EGTA)
and monoclonal mouse β-actin antibody (cat. no. A5316) were
purchased from Sigma-Aldrich (St. Louis, MO, USA). The rabbit
polyclonal UCP2 antibody (cat. no. 11081-1-AP) was purchased from
Proteintech Group, Inc. (Chicago, IL, USA). Rabbit polyclonal SIRT1
(cat. no. sc-15404) and mouse monoclonal prohibitin (cat. no.
sc-377037) antibodies were purchased from Santa Cruz Biotechnology,
Inc. (Dallas, TX, USA). Krebs-Henseleit (KH) solution (pH 7.4)
consisted of: NaCl 118.0 mmol/l, KCl 4.7 mmol/l,
K2PO4 1.2 mmol/l, MgSO4 1.2
mmol/l, NaHCO3 25.0 mmol/l, CaCl2 1.25 mmol/l
and glucose 10.0 mmol/l. Celsior solution (pH 7.4) consisted of:
NaOH 100 mmol/l, KCl 15 mmol/l, MgCl2 13 mmol/l,
CaCl2 0.25 mmol/l, mannitol 60 mmol/l, lactobionate 80
mmol/l, histidine 30 mmol/l, glutamate 20 mmol/l.
In vitro hypothermic heart preservation
model
Male Sprague-Dawley rats were anesthetized with 60
mg/kg sodium pentobarbital (intraperitoneal). The hearts were
rapidly excised and washed in ice-cold KH solution, and mounted for
retrograde perfusion using a modified Langendorff method (8). Following equilibration perfusion with
KH solution (37°C) for 30 min, the left ventricular end-diastolic
pressure (LVEDP), left ventricular developed pressure (LVDP),
maximal systolic velocity of left ventricular pressure
(+dP/dtmax), maximal diastolic velocity of left
ventricular pressure (−dP/dtmax) and heart rate were
recorded as the basal value. Ice-cold Celsior solution was perfused
into the aorta to induce cardiac arrest, then the rat hearts were
subjected to 3–12 h of preservation in ice-cold Celsior solution
followed by reperfusion with KH solution (37°C) for 60 min.
Coronary flow was recorded during the equilibration and
reperfusion.
Animal grouping
Rat hearts were divided into the following groups
(n=8): i) Control group, not preserved in Celsior solution; ii)
Celsior group, preserved in Celsior solution for 3–12 h; iii)
genipin group, preserved in Celsior solution containing genipin (10
µmol/l) for 9 h; iv) resveratrol group, stored in Celsior
solution containing resveratrol (10, 20 and 40 µM) for 9 h;
v) resveratrol + EX-527 group, stored in Celsior solution
containing resveratrol (40 µM) and EX-527 (10 µmol/l)
for 9 h; and vi) EX-527 group, stored in Celsior solution
containing EX-527 (10 µmol/l) for 9 h.
Isolation of mitochondria
Cardiac mitochondria were prepared from rat hearts
as previously described (9).
Briefly, rat ventricular myocardium was excised and homogenized in
an ice-cold isolation buffer with EGTA (1 mmol/l). The homogenates
were centrifuged at 1,000 × g for 10 min at 4°C, then the
supernatants were collected and centrifuged at 10,000 × g for 20
min at 4°C. The resulting pellet was washed by resuspension in
isolation buffer without EGTA and centrifuged again at 10,000 × g
for 20 min at 4°C. The final pellet resuspended in the isolation
buffer was used for further assay. Mitochondrial purity was
assessed by western blot analysis with the mitochondrial marker
prohibitin and the cytosolic marker β-actin.
Measurement of reactive oxygen
species
ROS production in the cardiac mitochondria was
determined using 2,7-dichlorodihydro fluorescent diacetate
(DCFH-DA; Beyotime Institute of Biotechnology, Haimen, China)
(10). Briefly, mitochondria (10
µg) were suspended in
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid buffer and were
incubated with 10 µM DCFH-DA at 37°C for 20 min in the
darkness. The fluorescence intensity was measured using a
microplate reader (Infinite M200; Tecan Group, Ltd., Männedorf,
Switzerland), at excitation and emission wavelengths of 485 nm and
530 nm, respectively.
Myocardial adenosine triphosphate (ATP)
content measurement
Myocardial ATP content was measured using the ATP
assay kit (Beyotime Institute of Biotechnology) based on the
luciferin-luciferase reaction. Bioluminescence intensity was
determined using a microplate reader (Infinite M200). The protein
concentration of each sample was determined using the Bicinchoninic
Acid Protein Assay kit (Beyotime Institute of Biotechnology). Total
ATP levels were expressed as nmol/mg protein.
Western blotting
Left ventricular myocardium was homogenized in
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology). Proteins were separated by 10% sodium dodecyl
sulphate-polyacrylamide gel electrophoresis and then blotted onto a
nitrocellulose membrane (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA). For the detection of proteins, the
membrane was incubated with the UCP2 (1:1,000), SIRT1 (1:200) or
β-actin (1:5,000) primary antibodies overnight at 4°C, and then
incubated with goat anti-mouse (sc-2005) and goat anti-rabbit
(sc-2004) horseradish peroxidase-conjugated secondary antibodies
(1:2,000; Santa Cruz Biotechnology, Inc.) for 1 h at room
temperature. The proteins were visualized using an enhanced
chemiluminescence kit (Beyotime Institute of Biotechnology). The
band density was determined using Quantity One software (version
4.62; Bio-Rad Laboratories, Inc., Hercules, CA, USA) and normalized
to that of β-actin.
Statistical analysis
Data were expressed as the mean ± standard error and
analyzed using a one-way analysis of variance with Newman-Keuls'
post hoc test as required. P<0.05 was considered to indicate a
statistically significant difference.
Results
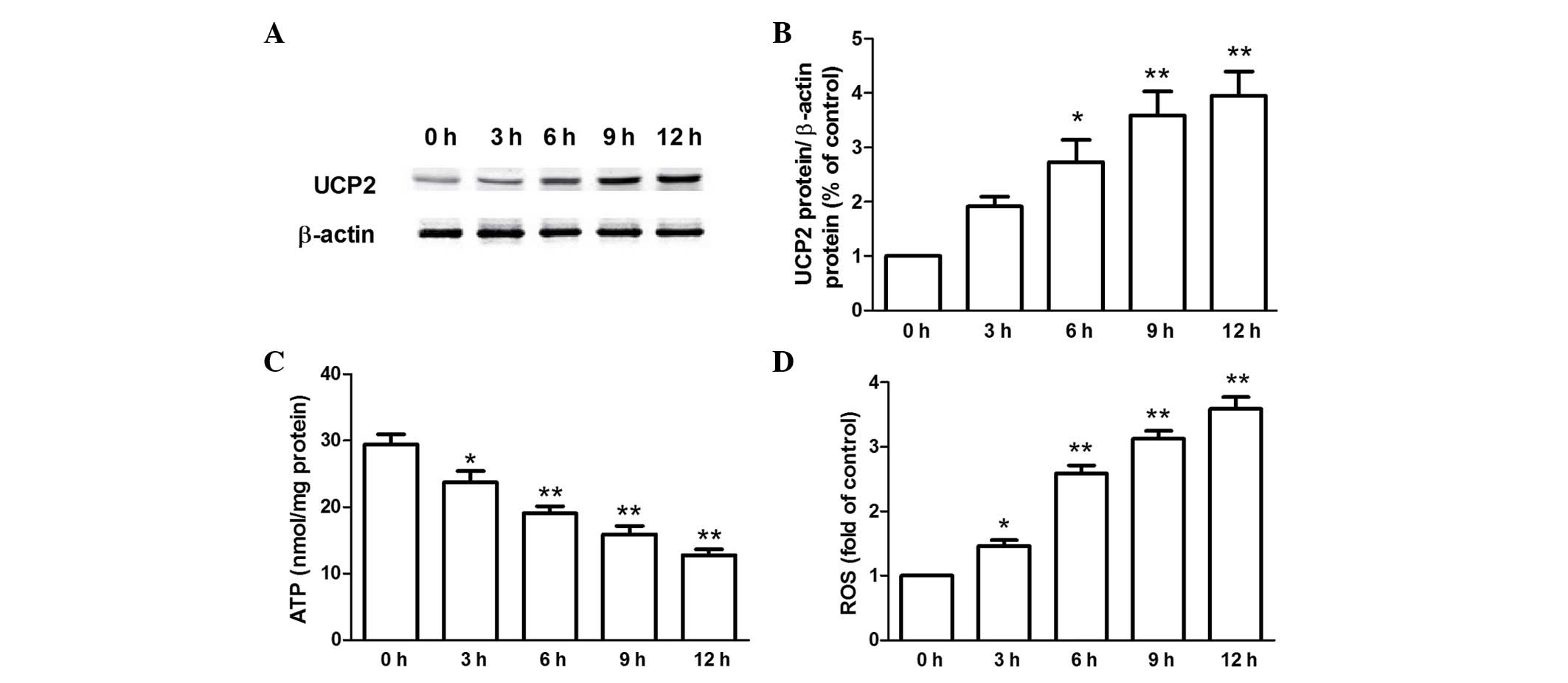
Alterations in UCP2 protein levels in rat
hearts following hypothermic preservation
Subsequent to preservation in ice-cold Celsior
solution for 3–12 h, the UCP2 protein expression levels in
hypothermic preserved rat hearts were increased in a time-dependent
manner (Fig. 1). The LVEDP was
significantly increased, the LVDP, ±dP/dtmax, heart rate
and coronary flow were significantly reduced in rat hearts
undergoing 9 h of hypothermic preservation followed by 60 min of
reperfusion (P<0.01; Fig.
2A–F). Celsior solution supplemented with the UCP2 inhibitor
genipin reversed the hypothermic preservation-induced cardiac
dysfunction (P<0.01; Fig.
2A–F). The decline in ATP production induced by 9 h of
preservation was also prevented by the supplement of genipin
(P<0.01; Fig. 2G). However,
genipin had no effect on the hypothermic preservation-induced
increase in mitochondrial ROS levels (Fig. 2H).
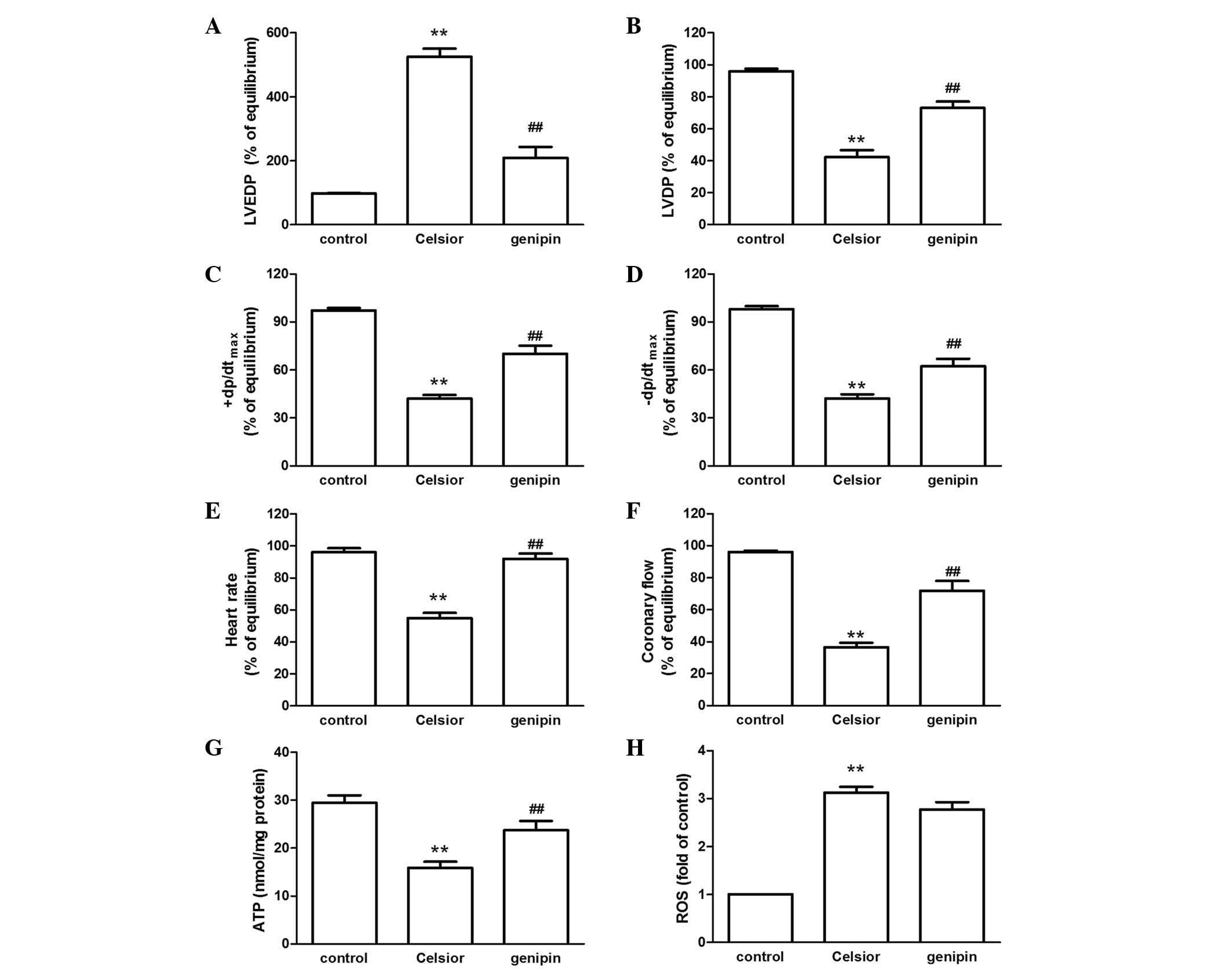 | Figure 2Effect of the UCP2 inhibitor genipin
on (A) LVEDP, (B) LVDP, (C) +dP/dtmax, (D)
−dp/dtmax, (E) heart rate, (F) coronary flow, (G) ATP
production, and (H) ROS level in rat hearts undergoing 9 h of
hypothermic preservation followed by 60 min of reperfusion. Data
are expressed as the mean ± standard error, n=8.
**P<0.01 vs. control group (not preserved in Celsior
solution); ##P<0.01 vs. Celsior group. UCP2,
uncoupling protein 2; LVEDP, left ventricular end-diastolic
pressure; LVDP, left ventricular developed pressure;
+dP/dtmax, maximal systolic velocity of left ventricular
pressure; −dP/dtmax, maximal diastolic velocity of left
ventricular pressure; ATP, adenosine triphosphate; ROS, reactive
oxygen species. |
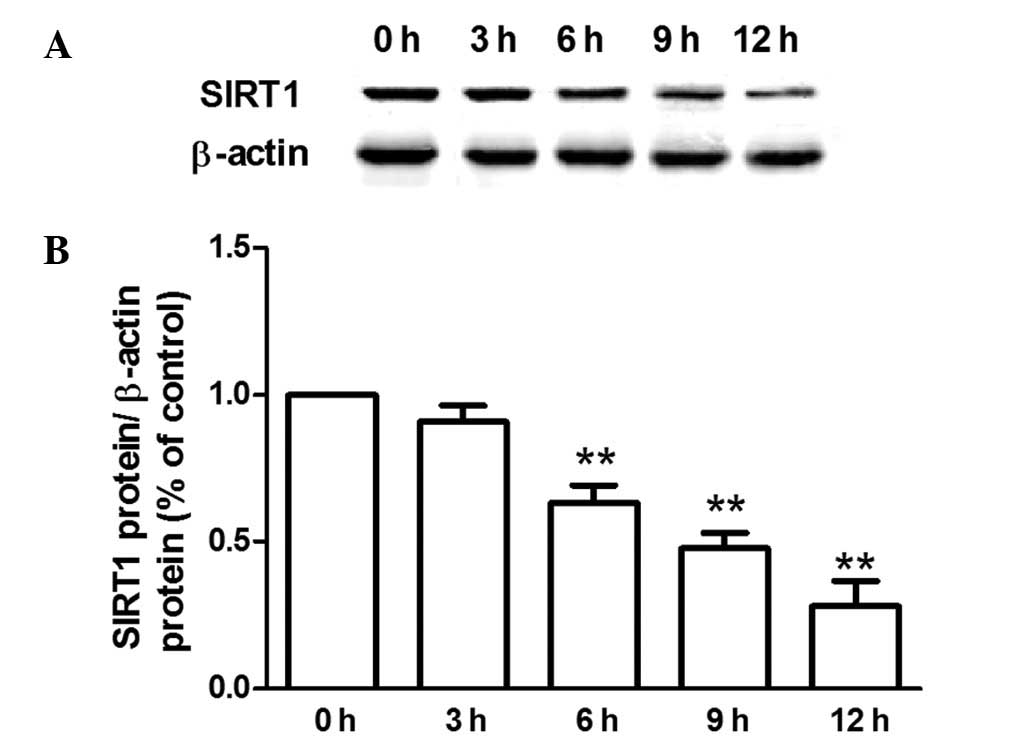
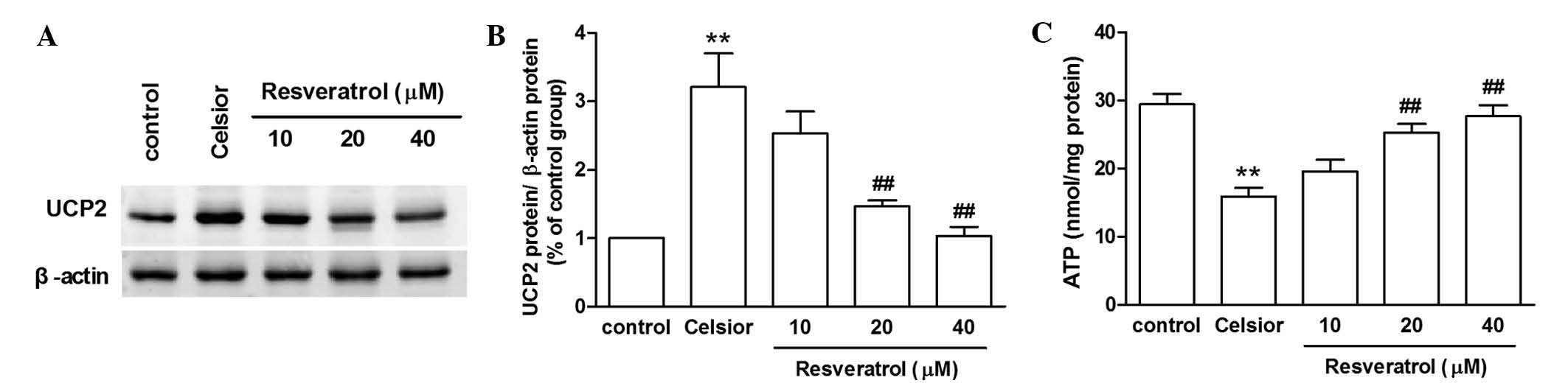
Effect of SIRT1 activation on hypothermic
preservation-induced injury of rat hearts
Compared with the control group, the SIRT1 protein
expression in rat hearts was reduced following preservation for
6–12 h (Fig. 3). The SIRT1
activator resveratrol (20 or 40 µmol/l) inhibited the UCP2
protein overexpression induced by 9 h of hypothermic preservation,
and prevented the hypothermic preservation-induced decline in ATP
production (P<0.01; Fig. 4).
Compared with 9 h of preservation rat hearts, Celsior solution
supplement with resveratrol (20 or 40 µmol/l) additionally
prevented the hypo-thermic preservation-induced increase in LVEDP,
improved the LVDP and ±dP/dtmax recovery, enhanced the
heart rate and coronary flow (Fig.
5).
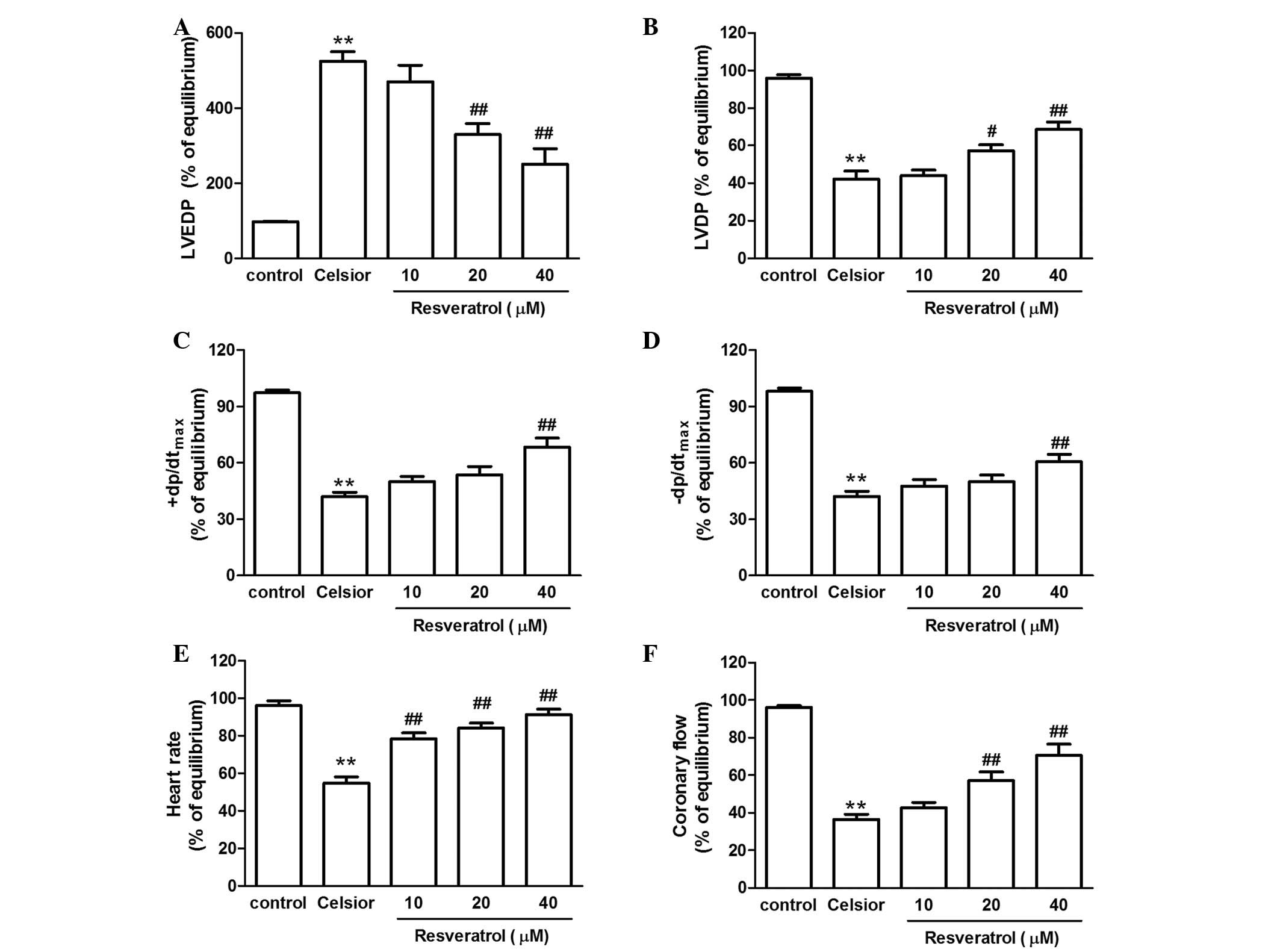 | Figure 5Effect of resveratrol on (A) LVEDP,
(B) LVDP, (C) +dP/dtmax, (D) −dp/dtmax, (E)
heart rate and (F) coronary flow in rat hearts undergoing 9 h of
hypothermic preservation followed by 60 min of reperfusion. Data
are expressed as the mean ± standard error, n=8.
**P<0.01 vs. control group; #P<0.05,
##P<0.01 vs. Celsior group. LVEDP, left ventricular
end-diastolic pressure; LVDP, left ventricular developed pressure;
+dP/dtmax, maximal systolic velocity of left ventricular
pressure; −dP/dtmax, maximal diastolic velocity of left
ventricular pressure. |
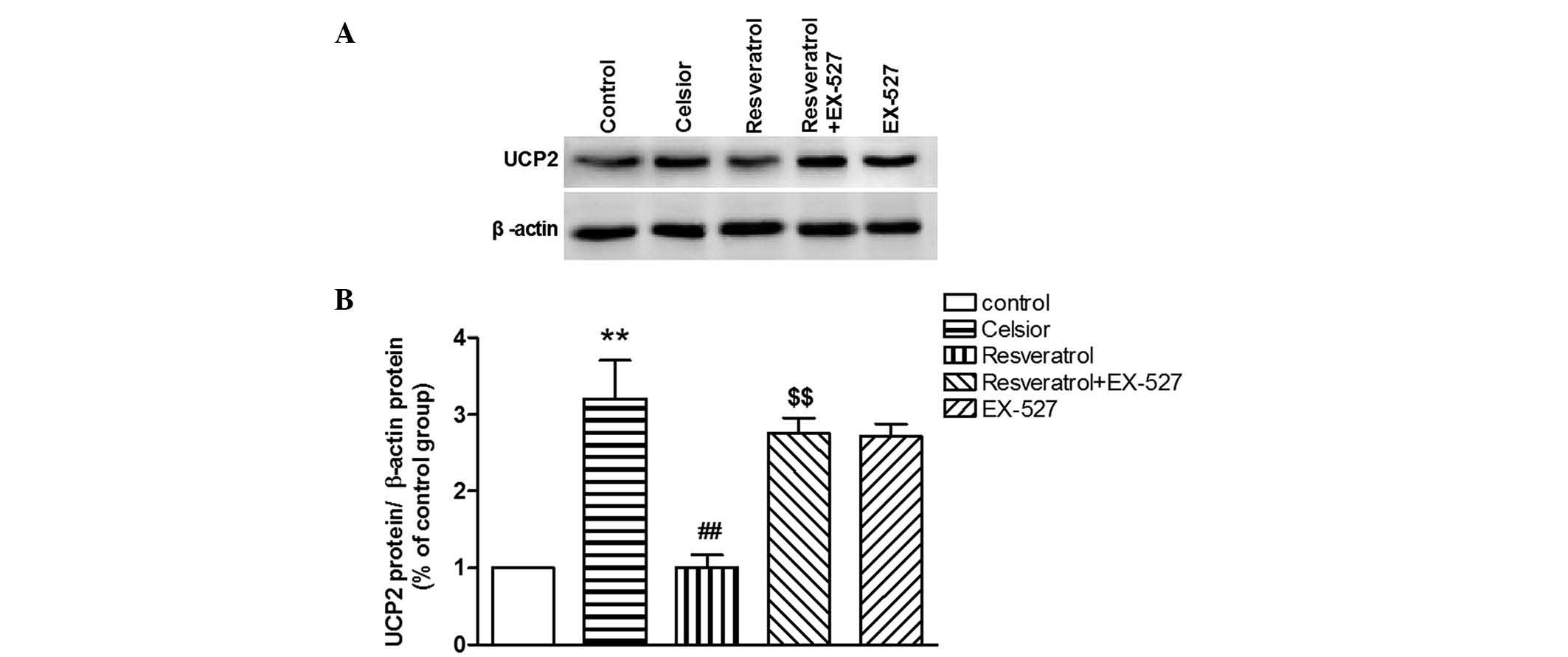
Effect of SIRT1 inhibitor on
resveratrol-induced improvement of cardiac function in hypothermic
preserved rat hearts
Compared with the Celsior group, the SIRT1 inhibitor
EX-527 did not significantly effect the UCP2 protein expression and
cardiac function in hypothermic preserved rat hearts. However,
EX-527 was able to prevent the resveratrol-induced inhibition of
UCP2 overexpression (P<0.01; Fig.
6), and abolish the resveratrol-induced protection against
cardiac dysfunction in the hypothermic preserved rat heart
(P<0.01; Fig. 7).
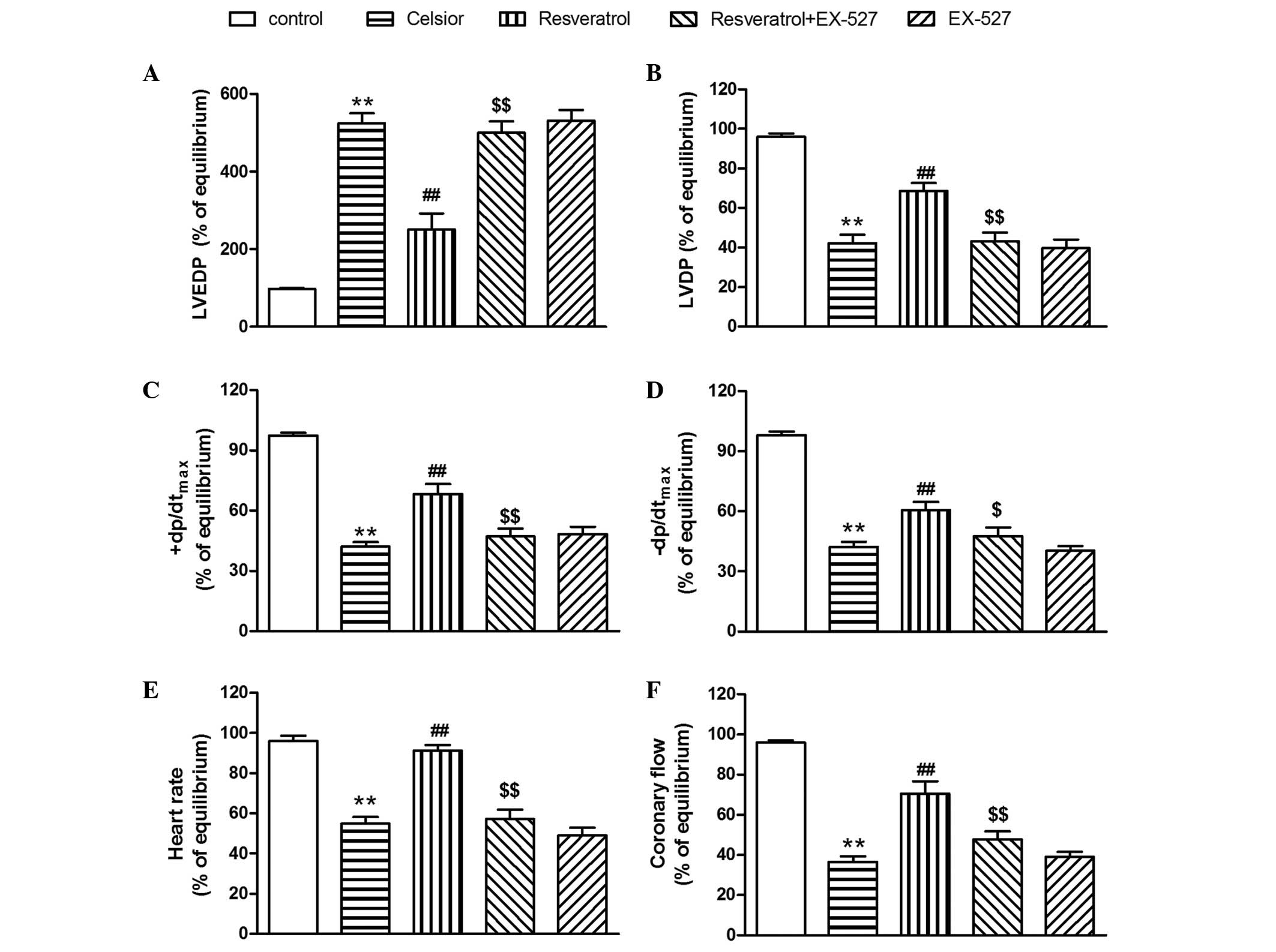 | Figure 7SIRT1 inhibitor EX-527 abolishes the
resveratrol-induced improvement of (A) LVEDP, (B) LVDP, (C)
+dP/dtmax, (D) −dp/dtmax, (E) heart rate and
(F) coronary flow in hypothermic preserved hearts. Data are
expressed as the mean ± standard error, n=8. **P<0.01
vs. control group (not preserved in Celsior solution);
##P<0.01 vs. Celsior group. $P<0.05,
$$P<0.01 vs. resveratrol (40 µM) group. SIRT1,
silent mating type information regulation 2 homolog 1; LVEDP, left
ventricular end-diastolic pressure; LVDP, left ventricular
developed pressure; +dP/dtmax, maximal systolic velocity
of left ventricular pressure; −dP/dtmax, maximal
diastolic velocity of left ventricular pressure. |
Discussion
Successful organ preservation is imperative to
reduce the ischemia-reperfusion injury in clinical heart
transplantation. Cardiac mitochondrial dysfunction is deemed as one
of the key challenges limiting heart preservation. UCP2, as an
inner mitochondrial membrane proton carrier that uncouples ATP
synthesis, is able to facilitate proton leak into the mitochondrial
matrix to promote partial uncoupled respiration; however if in
excess, will facilitate mitochondrial and cellular damage. In the
present study, the UCP2 protein expression in rat hearts was
observed to increase in a time-dependent manner following cold
preservation. The UCP2 inhibitor genipin inhibited the hypothermic
preservation-induced cardiac dysfunction, prevented decline in ATP
production, indicating that overexpression of UCP2 serves a
negative role in myocardial ischemic injury.
The exact role of upregulation of UCP2 in rat heart
mitochondria remains to be determined. UCP2 has been suggested to
protect cardiomyocytes against oxidative stress by dissipating the
mitochondrial proton gradient and mitochondrial membrane potential,
thereby reducing mitochondrial ROS generation. McLeod et al
(5) reported that preconditioning
in rat cardiac derived myoblasts is abolished following UCP2
depletion by RNA-interference. However, in apparent conflict with
its uncoupling role, UCP2 has additionally been hypothesized to be
essential for mitochondrial Ca2+ uptake, which has a
protective action by stimulating mitochondrial ATP production. UCP2
overexpression attenuates Ca2+ overload and the
production of ROS in mitochondria (11). UCP2 additionally increases
sensitivity of adult rat cardiac myocytes to hypoxia-reoxygenation
via ATP depletion and acidosis (12). In the present study, the UCP2
inhibitor genipin inhibited the hypothermic preservation-induced
cardiac dysfunction, prevented decline in ATP production induced by
9 h of preservation, however had no effect on the hypothermic
preservation-induced increase in mitochondrial ROS level,
suggesting that elevated UCP2 in the heart during hypothermic
preservation UCP2 may result in mitochondrial damage and
consequently the attenuation of ATP production. As reported,
diminution of ATP synthesis is able to trigger the closing of the
mitochondrial ATP-sensitive potassium channel, which is aim in
cellular protection. In previous studies, Celsior solution
supplemented with diazoxide significantly enhanced the LVDP
recovery rate and reduced the apoptotic index (9).
There are various mechanisms whereby mitochondrial
UCPs can be evoked in the context of cardiac ischemia and
reperfusion (13). ROS (14), beta-adrenergic stimulation
(15), free fatty acids (16) and SIRT1 have been reported as
regulators of UCP2 expression. SIRT1, an nicotine adenine
dinucleotide(+)-dependent deacetylase, is a regulator responsible
for various biological effects, predominantly in metabolism and
aging (17–21). SIRT1 is essential for the
maintenance of cardiac mitochondrial integrity and normal postnatal
myocardium development (22). In
hearts from SIRT1-deficient mice, morphological and functional
mitochondrial abnormalities were observed. The expression of SIRT1
is downregulated in advanced heart failure samples compared with
healthy control cardiomyocytes (23). SIRT1-dependent lysine deacetylation
occurs during ischemic preconditioning and may serve a role in
cardio-protective signaling (24).
Downstream targets of SIRT1 include peroxisome
proliferator-activated receptor (PPAR)-γ, PPARγ coactivator-1α and
UCP2 (25). Resveratrol is a
natural polyphenolic compound that has cardioprotective, anticancer
and anti-inflammatory properties (26,27),
through a SIRT1-dependent (28) or
independent pathway (29). It was
identified that the SIRT1 protein expression in rat hearts reduced
following hypothermic preservation, while Celsior solution
supplemented with the SIRT1 activator resveratrol inhibited the
UCP2 protein overexpression, prevented the decline in ATP
production, improving cardiac function. The SIRT1 inhibitor EX-527
abolished the resveratrol-induced inhibition of UCP2 overexpression
and cardiac protection in the hypothermic preserved rat heart.
Thus, it was suggested that SIRT1 is required for cardioprotection
against ischemia-reperfusion injury, and reduction of SIRT1 is
associated with overexpression of UCP2. In the kidney and brain,
SIRT1 is suggested to regulate the mitochondrial UCP2 via the
peroxisome proliferator-activated receptor-γ coactivator-1α/PPARα
signaling pathway (30,31).
In conclusion, these results suggest that
downregulation of UCP2 expression in the hypothermic preserved rat
heart in part initiated the protection mechanism via the SIRT1
pathway.
Acknowledgments
The current study was supported by the National
Natural Science Foundation of China (grant no. 81270178), the
Science and Technology Department of Zhejiang Province (grant no.
2015C37129), and Jiaxing Science and Technology Project (grant no.
2012AY1075-5).
References
|
1
|
Shah KB and Parameshwar J: Advances in
heart transplantation: The year in review. J Heart Lung Transplant.
30:241–246. 2011. View Article : Google Scholar
|
|
2
|
Cannon RM, Hughes MG, Jones CM, Eng M and
Marvin MR: A review of the United States experience with combined
heart-liver transplantation. Transpl Int. 25:1223–1228. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Baines CP: The cardiac mitochondrion:
Nexus of stress. Annu Rev Physiol. 72:61–80. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Alán L, Smolková K, Kronusová E, Santorová
J and Jezek P: Absolute levels of transcripts for mitochondrial
uncoupling proteins UCP2, UCP3, UCP4 and UCP5 show different
patterns in rat and mice tissues. J Bioenerg Biomembr. 41:71–78.
2009. View Article : Google Scholar
|
|
5
|
McLeod CJ, Aziz A, Hoyt RF Jr, McCoy JP Jr
and Sack MN: Uncoupling proteins 2 and 3 function in concert to
augment tolerance to cardiac ischemia. J Biol Chem.
280:33470–33476. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zungu M, Alcolea MP, Garcia-Palmer FJ,
Young ME and Essop MF: Genomic modulation of mitochondrial
respiratory genes in the hypertrophied heart reflects adaptive
changes in mitochondrial and contractile function. Am J Physiol
Heart Circ Physiol. 293:H2819–H2825. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals: Guide for the Care and Use of Laboratory Animals. 8th
edition. National Academies Press; Washington, D.C: 2011
|
|
8
|
Yan ZK, Hu ZB, Pan XH, Chen YY, Zhang XM
and Shen YL: Diazoxide supplemented Celsior solution improves
hypothermic heart preservation effect in rat through activation of
mitochondrial ATP-sensitive potassium channel. Pharm Biol.
47:1060–1066. 2009. View Article : Google Scholar
|
|
9
|
Yang F, Chen WL, Zheng MZ, Yu GW, Xu HJ,
Shen YL and Chen YY: Heat shock protein 90 mediates anti-apoptotic
effect of diazoxide by preventing the cleavage of Bid in
hypothermic preservation rat hearts. J Heart Lung Transplant.
30:928–934. 2011.PubMed/NCBI
|
|
10
|
Zhou HY, Zhang LN, Zheng MZ, Wang LL, Chen
YY and Shen YL: Improved myocardial function with supplement of
levosimendan to celsior solution. J Cardiovasc Pharmacol.
64:256–265. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Teshima Y, Akao M, Jones SP and Marbán E:
Uncoupling protein-2 overexpression inhibits mitochondrial death
pathway in cardiomyocytes. Circ Res. 93:192–200. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bodyak N, Rigor DL, Chen YS, Han Y,
Bisping E, Pu WT and Kang PM: Uncoupling protein 2 modulates cell
viability in adult rat cardiomyocytes. Am J Physiol Heart Circ
Physiol. 293:H829–H835. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Toda C and Diano S: Mitochondrial UCP2 in
the central regulation of metabolism. Best Pract Res Clin
Endocrinol Metab. 28:757–764. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Brand MD, Affourtit C, Esteves TC, Green
K, Lambert AJ, Miwa S, Pakay JL and Parker N: Mitochondrial
superoxide: Production, biological effects and activation of
uncoupling proteins. Free Radic Biol Med. 37:755–767. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ishizawa M, Mizushige K, Noma T, Namba T,
Guo P, Murakami K, Tsuji T, Miyatake A, Ohmori K and Kohno M: An
antioxidant treatment potentially protects myocardial energy
metabolism by regulating uncoupling protein 2 expression in a
chronic beta-adrenergic stimulation rat model. Life Sci.
78:2974–2982. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Murray AJ, Panagia M, Hauton D, Gibbons GF
and Clarke K: Plasma free fatty acids and peroxisome
proliferator-activated receptor alpha in the control of myocardial
uncoupling protein levels. Diabetes. 54:3496–3502. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yacoub R, Lee K and He JC: The Role of
SIRT1 in Diabetic Kidney Disease. Front Endocrinol (Lausanne).
5:1662014.
|
|
18
|
Rehan L, Laszki-Szcząchor K,
Sobieszczańska M and Polak-Jonkisz D: SIRT1 and NAD as regulators
of ageing. Life Sci. 105:1–6. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Philp A and Schenk S: Unraveling the
complexities of SIRT1-mediated mitochondrial regulation in skeletal
muscle. Exerc Sport Sci Rev. 41:174–181. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kitada M and Koya D: SIRT1 in type 2
diabetes: Mechanisms and therapeutic potential. Diabetes Metab J.
37:315–325. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sundaresan NR, Pillai VB and Gupta MP:
Emerging roles of SIRT1 deacetylase in regulating cardiomyocyte
survival and hypertrophy. J Mol Cell Cardiol. 51:614–618. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Planavila A, Dominguez E, Navarro M,
Vinciguerra M, Iglesias R, Giralt M, Lope-Piedrafita S, Ruberte J
and Villarroya F: Dilated cardiomyopathy and mitochondrial
dysfunction in Sirt1-deficient mice: A role for Sirt1-Mef2 in adult
heart. J Mol Cell Cardiol. 53:521–531. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lu TM, Tsai JY, Chen YC, Huang CY, Hsu HL,
Weng CF, Shih CC and Hsu CP: Downregulation of Sirt1 as aging
change in advanced heart failure. J Biomed Sci. 21:572014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nadtochiy SM, Yao H, McBurney MW, Gu W,
Guarente L, Rahman I and Brookes PS: SIRT1-mediated acute
cardioprotection. Am J Physiol Heart Circ Physiol. 301:H1506–H1512.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chaudhary N and Pfluger PT: Metabolic
benefits from Sirt1 and Sirt1 activators. Curr Opin Clin Nutr Metab
Care. 12:431–437. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pangeni R, Sahni JK, Ali J, Sharma S and
Baboota S: Resveratrol: Review on therapeutic potential and recent
advances in drug delivery. Expert Opin Drug Deliv. 11:1285–1298.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Granzotto A and Zatta P: Resveratrol and
Alzheimer's disease: Message in a bottle on red wine and cognition.
Front Aging Neurosci. 6:952014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zong Y, Sun L, Liu B, Deng YS, Zhan D,
Chen YL, He Y, Liu J, Zhang ZJ, Sun J and Lu D: Resveratrol
inhibits LPS-induced MAPKs activation via activation of the
phosphatidylinositol 3-kinase pathway in murine RAW 264.7
macrophage cells. PLoS One. 7:e441072012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Centeno-Baez C, Dallaire P and Marette A:
Resveratrol inhibition of inducible nitric oxide synthase in
skeletal muscle involves AMPK but not SIRT1. Am J Physiol
Endocrinol Metab. 301:E922–E930. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rubattu S, Di Castro S, Cotugno M, Bianchi
F, Mattioli R, Baima S, Stanzione R, Madonna M, Bozzao C, Marchitti
S, et al: Protective effects of Brassica oleracea sprouts extract
toward renal damage in high-salt-fed SHRSP: Role of AMPK/PPARα/UCP2
axis. J Hypertens. 33:1465–1479. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang SJ, Zhao XH, Chen W, Bo N, Wang XJ,
Chi ZF and Wu W: Sirtuin 1 activation enhances the
PGC-1/mitochondrial antioxidant system pathway in status
epilepticus. Mol Med Rep. 11:521–526. 2015.
|