Introduction
Malignant glioma, which is the most common type of
primary brain tumor, is highly aggressive and metastatic. Survival
ranges between 12 and 40 months from initial diagnosis, and between
6 and 18 months following recurrence, despite treatment with
standard therapies, including surgery, chemotherapy and
radiotherapy (1–3). Therefore, it is necessary to develop
novel strategies for the treatment of glioma. Mitochondria are
unique cellular organelles, the primary role of which is to
generate adenosine triphosphate (ATP) through oxidative
phosphorylation, in order to ensure survival (4). Previous studies have reported that
mitochondria have a central role in regulating proliferation,
apoptosis and autophagy in cancer cells (5–7).
Therefore, disturbing mitochondrial metabolism or interfering with
mitochondrial membrane permeabilization may be considered a
promising therapeutic approach to cancer.
Autophagy is a lysosomal degradation pathway by
which cells consume intracellular materials, including damaged
organelles, proteins or hazardous substances, in order to maintain
cellular homeostasis (8).
Autophagy is upregulated in response to cellular stress, including
starvation, mitochondrial stress, endoplasmic reticulum stress and
pathogenic infection (9). The
emerging role of autophagy in cancer is considered to be a
double-edged sword. Autophagy has been reported to render cancer
cells able to tolerate therapy-induced stress; however, autophagy
can also digest organelles and limit tumorigenesis (10). Therefore, modulating the autophagic
pathway may provide novel approaches to cancer therapy and
prevention.
Manganese (Mn) is an essential mineral that can
influence intracellular and extracellular metabolism associated
with the mitochondria (11).
Apoptosis is activated by Mn compounds, which induce the rupture of
DNA and the release of cytochrome c from the mitochondrial
intermembrane space to the cytosol (12). In addition, several enzymes have
been identified, which naturally contain Mn, including pyruvate
carboxylase, arginase and Mn superoxide dismutase (13,14).
The main purpose of Mn is as a coactivator of superoxide dismutase
in mitochondria (15). It has been
reported that Mn (II) ions are predominantly transported by
divalent metal transporter 1 (DMT-1) and the
transferrin-transferrin receptor (Tf-TfR) system, which is highly
expressed in various tumors (16).
It has previously been demonstrated that Mn (II) ions can transport
across the rat blood-brain barrier, through saturable and
Tf-dependent transport mechanisms, to the brain (17–19).
Based on the aforementioned findings, the present study
hypothesized that Mn-containing compounds may cross the blood-brain
barrier and selectively kill glioma cells via DMT-1 or TfR.
The present study indicated that the complex [(Adpa)
Mn(Cl)(H2O)], designated as Adpa-Mn, which is an
inorganic compound comprised of Adpa [bis(2-pyridylmethyl)
amino-2-propionic acid] and Mn, is a promising novel anti-glioma
agent with potent selective activity in vitro. Furthermore,
it was verified that Adpa-Mn induces apoptotic cell death and
cytoprotective autophagy by triggering mitochondrial
dysfunction.
Materials and methods
Materials
The compound Adpa-Mn was synthesized by Professor
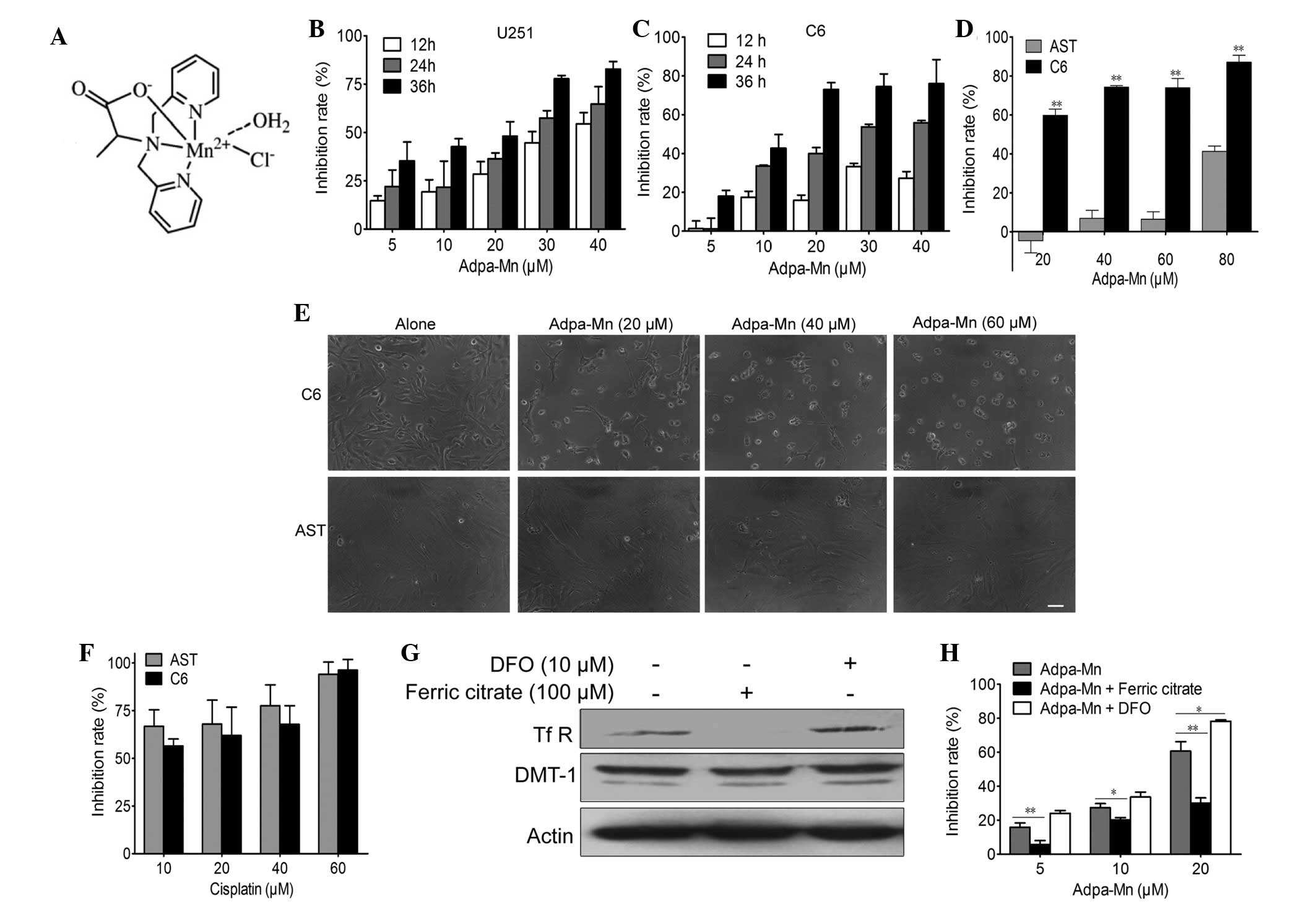
Chen Qiuyun (Fig. 1A) as
previously described (20).
Cisplatin was purchased from Jiangsu Hengrui Medicine Co., Ltd.
(Lianyungang, China).
3-[4,5-dimehyl-2-thiazolyl]-2,5-diphenyl-2H-tetrazolium bromide
(MTT) was purchased from Amresco, LLC (Solon, OH, USA). JC-1 for
mitochondrial membrane potential, Dulbecco's modified Eagle's
medium (DMEM) for U251 and C6, RPMI-1640 for rat astrocyte cells,
trypsin-EDTA solution for cell detachment and the Annexin
V/propidium iodide (PI) kit for apoptotic detection were purchased
from Thermo Fisher Scientific, Inc. (Waltham, MA, USA). Ferric
citrate, 4′,6-diamidino-2-phenylindole (DAPI), desferrioxamine
(DFO) and cyclosporin A (CsA) were purchased from Sigma-Aldrich
(St. Louis, MO, USA). Fetal bovine serum (FBS) was obtained from
Sijiqing Biological Engineering Materials Co., Ltd. (Hangzhou,
China). The 2′,7′-dichlorofluorescein-diacetate (DCFH-DA) kit was
purchased from Beyotime Institute of Biotechnology (Nantong,
China). Anti-microtubule-associated protein 1 light chain 3 (LC3;
cat. no. 3868; 1:1,000), anti-poly (ADP-ribose) polymerase (PARP;
cat. no. 9536; 1:1000), anti-caspase-9 (cat. no. 9508, 1:1,000),
anti-caspase-7 (cat. no. 9494, 1:1,000), anti-caspase-3 (cat. no.
9665; 1:1,000) and anti-B-cell lymphoma 2 (Bcl-2;cat. no. 15071;
1:1,000), cytochrome c (cat. no. 4280, 1:1000), cytochrome
c oxidase subunit IV (cat. no. 4850; 1:1,000) were purchased
from Cell Signaling Technology, Inc. (Danvers, MA, USA) and were
diluted with 1% bovine serum albumin (BSA). Anti-β-actin antibody
(cat. no. SC-8432; 1:200) was obtained from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA), anti-DMT-1 antibody (cat.
no. 20507-1-AP; 1:200) was purchased from Proteintech Group, Inc.
(Rosemont, IL, USA), and anti-TfR antibody (cat. no. MA5-11441;
1:200) was purchased from Thermo Fisher Scientific, Inc.
Monodansylcadaverin (MDC), 3-methyladenine (3-MA), N-acetylcysteine
(NAC), ferric citrate, DFO, CsA and chloroquine (CQ) were purchased
from Sigma-Aldrich. All other chemicals were of high purity, and
were purchased from commercial sources. The study was approved by
the ethics committee of Jiangsu University (Zhenjiang, China).
Cell culture
The U251 human glioma cell line, and the C6 rat
glioma cell line were obtained from the Cancer Cell Repository
(Shanghai Cell Bank; Shanghai, China). The cells were maintained in
DMEM supplemented with 10% (v/v) heat-inactivated FBS and
antibiotics (100 U/ml penicillin and 100 U/ml streptomycin;
Beyotime Institute of Biotechnology) at 37°C in a humidified
atmosphere containing 5% CO2. Primary astrocyte cells
isolated from three newborn Sprague-Dawley rats were cultured in
RPMI-1640 as described previously (21).
Cell viability assay
The cells were plated at a density of
~4×103 viable cells/well in 96-well plates in the
presence of 5, 10, 20, 30, or 40 µM of the compound for 12,
24, or 36 h. MTT (1 mg/ml) was added for 3 h followed by dimethyl
sulfoxide to dissolve the formazan product, and the U251 and C6
cell viabilities were measured using a 96-well plate reader
(SpectraMax 190; Molecular Devices,Sunnyvale, CA, USA) at 490
nm.
Cell apoptosis assay
The cells were plated at a density of
~1×105 viable cells/well in 6-well plates. Cells were
harvested following 5, 10, 20 µM Adpa-Mn treatments for 24
h. Cells were collected using trypsin-EDTA and incubated with
Annexin V/PI at room temperature for 15 min in the dark, then
analyzed using a FACSCalibur flow cytometer (BD Biosciences,
Franklin Lakes, NJ, USA). CellQuest (Becton Dickinson, Mountain
View, CA, USA) was applied to analyze flow cytometric results.
Annexin V+/PI− and Annexin
V+/PI+ cells were considered to be early and
late phase apoptotic cells, respectively.
Visualization of MDC-labeled
vacuoles
Autophagic vacuoles were labeled with MDC. Briefly,
U251 cells grown on coverslips were incubated with 50 µM MDC
in phosphate-buffered saline (PBS) at 37°C for 10 min. Alterations
in cellular fluorescence were observed under a Nikon eclipse Ti
fluorescence microscope (Nikon Corporation, Tokyo, Japan;
excitation, 380–420 nm; emission, 450 nm).
Western blot analysis
Proteins were extracted from the cells using lysis
buffer (30 mM Tris, pH 7.5; 150 mM sodium chloride; 1 mM
phenylmethylsulfonyl fluoride; 1 mM sodium orthovanadate; 1%
Nonidet P-40; 10% glycerol; and phosphatase and protease
inhibitors; Roche Diagnostics, Basel, Switzerland). Then, 20
µg protein was separated by 15% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis at a constant voltage
setting of 110 V for 80 min and were electrophoretically
transferred onto polyvinylidene fluoride membranes (Thermo Fisher
Scientific Inc.) at a constant current setting of 300 mA for 90
min. Membrane blocking was performed with 1% BSA for 1 h. The
membranes were probed with primary antibodies diluted with 1% BSA
overnight at 4°C, followed by an incubation with horseradish
peroxidase-conjugated goat anti-mouse (cat. no. sc-2969) and goat
anti-rabbit (cat. no. sc-2768; Santa Cruz Biotechnology, Inc.)
secondary antibodies for 2 h at room temperature. Detection was
performed using a LumiGLO Chemiluminescent Substrate system (KPL,
Gaithersburg, MD, USA).
Mitochondrial membrane potential
assay
Alterations to mitochondrial membrane potential were
measured using JC-1. U251 cells were washed with PBS and were
incubated with 5 µg/ml JC-1 at 37°C for 30 min. Cells were
then washed twice with PBS and were immediately assessed by
fluorescence spectrometry (Spectra Max Gemini; Molecular Devices).
A 488-nm filter was used to detect the excitation of JC-1. Emission
filters of 535 and 595 nm were used to quantify the population of
mitochondria exhibiting green (JC-1 monomers) and red (JC-1
aggregates) fluorescence. The ratio of red/green fluorescence was
used to reflect the mitochondrial membrane potential.
Determination of intracellular ATP
levels
ATP content was measured according to the
luciferin-luciferase method, which is based on the requirement of
ATP for luciferase to produce light. The cells were plated at a
density of ~1×105 viable cells/well in 6-well plates.
Cells were harvested following treatment with 20 µM Adpa-Mn
for 6, 12 and 24 h and were assayed for ATP using a chemical
luciferase ATP assay kit (Beyotime Institute of Biotechnology). The
quantity of ATP in the experimental samples was calculated from a
standard curve prepared with ATP, and was expressed as nmol/mg
protein.
Statistical analysis
Differences between groups were analyzed using a
two-tailed Student's t-test with GraphPad Prism version 5.00 for
Windows (GraphPad Software, San Diego CA, USA) and differences
among groups were analyzed using one-way analysis of variance. All
values are expressed as the mean ± standard deviation and P<0.05
was considered to indicate a statistically significant difference.
All experiments were repeated at least three times.
Results
Adpa-Mn exhibits significant and
selective anti-glioma activity
To determine whether Adpa-Mn has an inhibitory
effect on glioma cells, the U251 human glioma cell line and the C6
rat glioma cell line were subjected to an MTT assay. Treatment with
Adpa-Mn inhibited U251 and C6 cell proliferation in a dose- and
time-dependent manner (Fig. 1B and
1C).
Subsequently, it was determined whether Adpa-Mn
exhibits cancer cell selectivity. Treatment with Adpa-Mn exhibited
significant selectivity towards glioma cells (C6) over normal rat
astrocytes (Fig. 1D and E),
whereas cisplatin did not exert this selectivity (Fig. 1F). This preferential toxicity
toward cancer cells over non-cancer cells suggests the potential
use of this compound as an antitumor therapeutic option.
Due to the high expression of TfR on tumor cells,
the cancer cell selectivity of Adpa-Mn may be due to its transport
mechanism. As detected by western blotting and MTT assay, when the
expression of TfR was inhibited by ferric citrate (100 µM)
pretreatment for 24 h, the inhibitory effect of Adpa-Mn on U251
cells was significantly decreased. Conversely, when TfR expression
was upregulated following DFO (10 µM) pretreatment for 24 h,
Adpa-Mn induced U251 cell inhibition more efficiently (Fig. 1G and H).
Adpa-Mn induces apoptotic cell death
To investigate the potential anticancer mechanisms
of Adpa-Mn, U251 malignant glioma cells were treated with various
concentrations of Adpa-Mn for 24 h. Apoptotic cell death was
detected using 4′,6-diamidino-2-phenylindole, a typical nuclear
stain that can visualize nuclear condensation. The results
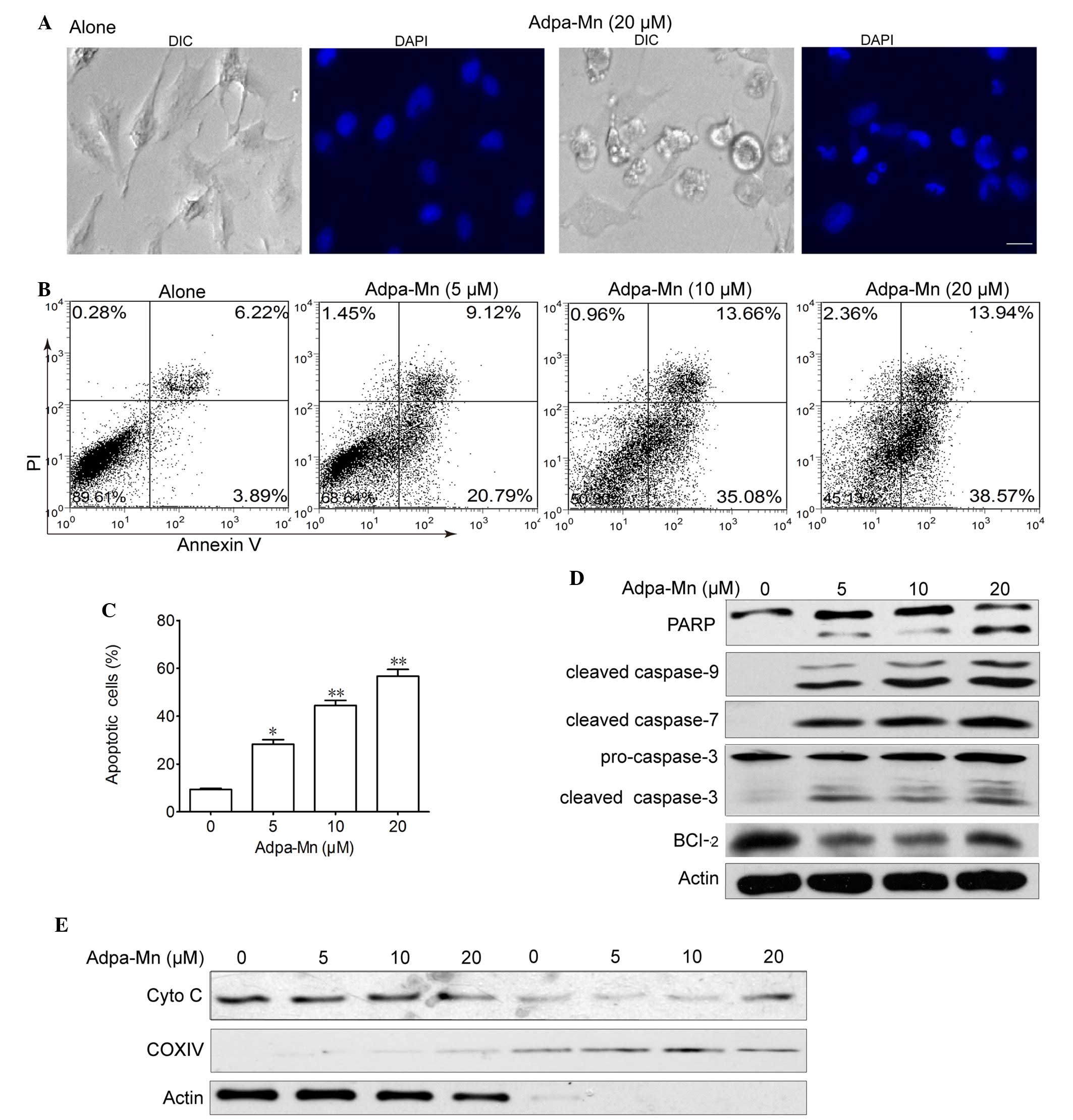
demonstrated that nuclei became condensed and fragmented, and
apoptotic bodies were detected following treatment with Adpa-Mn
(Fig. 2A). Adpa-Mn-induced
apoptosis was further detected using Annexin V/PI staining. As
presented in Fig. 2B and C,
Annexin V+ cells were abundantly increased from 10.1 to
48.7 and 52.5% following treatment with 10 and 20 µM
Adpa-Mn, respectively. To determine the involvement of the caspase
cascade in Adpa-Mn-induced apoptosis, the expression levels of
caspases were detected by western blotting. The hallmarks of
apoptosis, including PARP, caspase-9, caspase-7 and caspase-3, were
activated/cleaved in U251 cells following treatment with Adpa-Mn in
a dose-dependent manner (Fig. 2D).
Consistent with the decreased expression of Bcl-2 (Fig. 2D), which is an integral membrane
protein located mainly on the outer mitochondrial membrane that
inhibits apoptosis, cytochrome c was transferred from the
mitochondrial intermembrane space to the cytoplasm (Fig. 2E). These results suggest that
Adpa-Mn may induce significant apoptosis of U251 cells.
Adpa-Mn induces protective autophagy
Cellular features of necrosis, apoptosis and
autophagy frequently occur together in response to death signals
and toxic stress (22). The
present study determined whether Adpa-Mn was able to induce cell
death via autophagy. Autophagy is characterized by the accumulation
of vesicles and the formation of autophagosomes, which can be
detected by the presence of membrane-bound LC3-phospholipid
conjugates (23). MDC is a
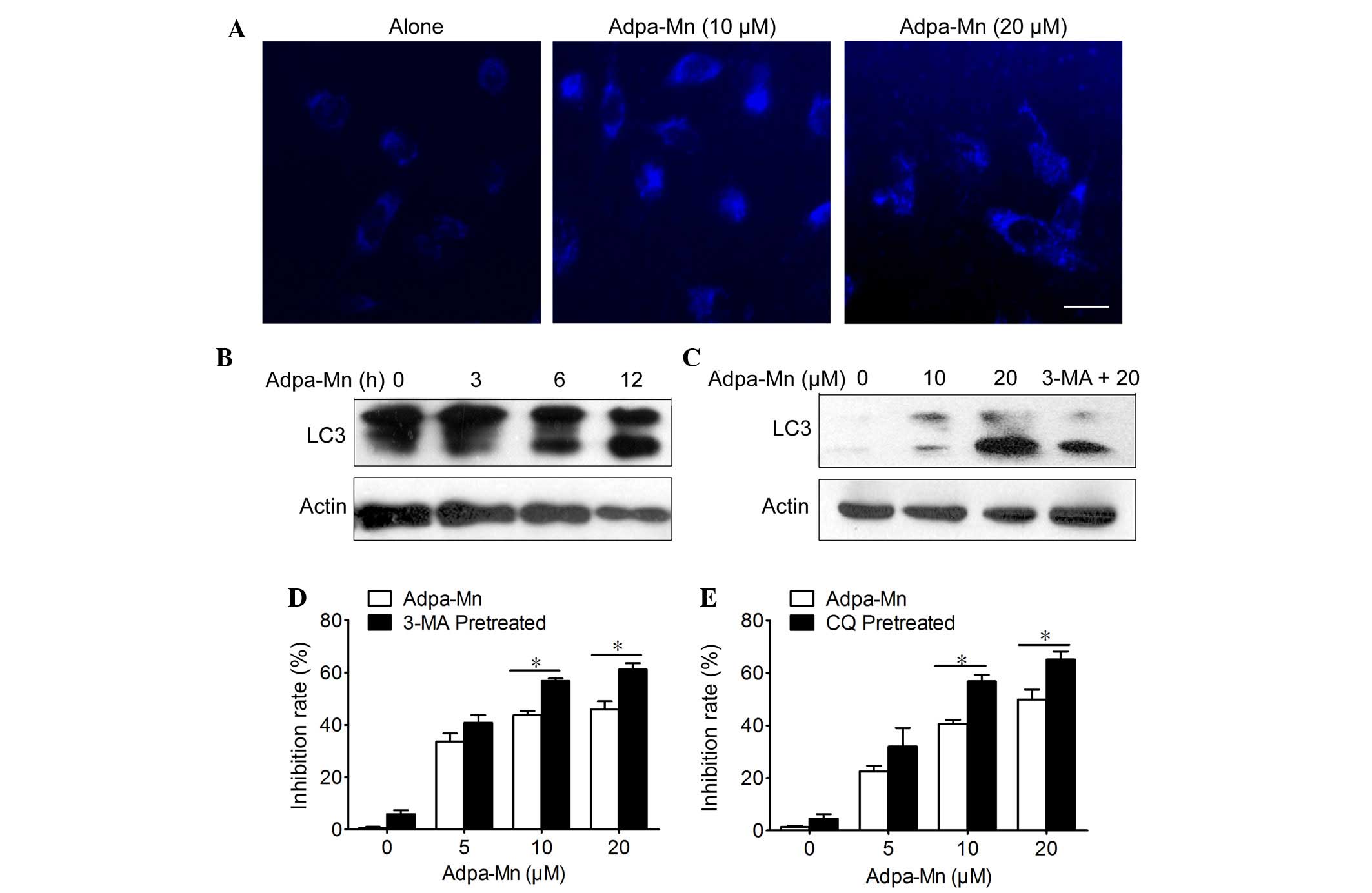
specific marker of autophagic vacuoles (Fig. 3A) and treatment with Adpa-Mn (10
and 20 µM) for 12 h triggered the accumulation of
MDC-stained acidic vesicular organelles (AVO) in U251 cells. In
addition, Adpa-Mn yielded a time- and dose-dependent increase in
the expression levels of LC3 II (the processed form of LC3;
Fig. 3B and C). In addition, the
LC3 II/LC3 I ratio was increased in a dose- and time-dependent
manner following treatment with Adpa-Mn. These data suggest that
autophagy was enhanced following treatment with Adpa-Mn. It has
previously been reported that apoptosis and autophagy are not
mutually exclusive pathways, but have been shown to act in synergy
and also to counter each other (24). Pretreatment with 3-MA, an inhibitor
of autophagy, and CQ, an inhibitor of lysosomal degradation,
decreased Adpa-Mn induced LC3 II accumulation and LC3 II/LC3 I
ratio (Fig. 3C), and accelerated
cell death (Fig. 3D and E).
Autophagy inhibition decreased cell viability, which indicated that
Adpa-Mn-induced autophagy exerts a protective role in U251
cells.
Adpa-Mn triggers mitochondrial
dysfunction
Since mitochondria are critical in apoptosis and
autophagy, the present study aimed to determine the effects of
Adpa-Mn on mitochondrial function. JC-1 was used to evaluate
mitochondrial membrane potential. Treatment with 5, 10 and 20
µM Adpa-Mn for 12 h led to a decrease in mitochondrial
membrane potential, as detected by enhanced green intensity and
reduced red intensity of JC-1 (Fig.
4A). Subsequently, the ratio of red and green fluorescence
intensity was determined using a fluorescent microplate reader,
which indicated a dose- and time-dependent decrease (Fig. 4B). DCFH-DA was used to determine
the effects of Adpa-Mn on intracellular ROS generation in U251
cells. As presented in Fig. 4C and
D, the fluorescence intensity of DCFH-DA per 1×104
cells was significantly elevated following treatment with Adpa-Mn
in a dose- and time-dependent manner, thus suggesting that
intracellular ROS production was increased. Furthermore, exposure
of U251 cells to Adpa-Mn resulted in ATP depletion (Fig. 4E). These results suggest that
Adpa-Mn triggers mitochondrial dysfunction in U251 cells.
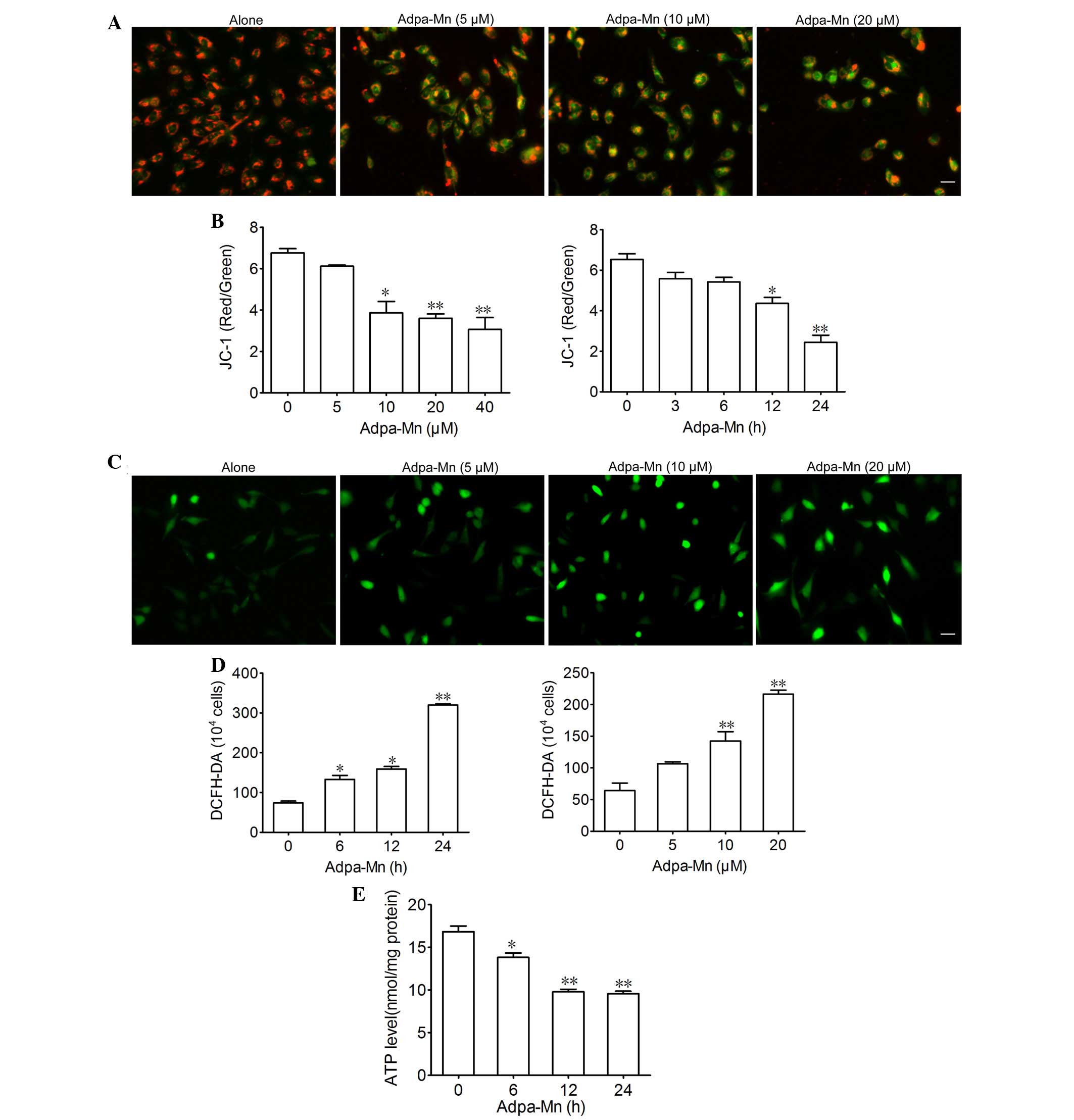 | Figure 4Adpa-manganese (Mn) treatment induced
mitochondrial dysfunction. (A and B) U251 cells were exposed to
Adpa-Mn (20 µM for 3, 6, 12 and 24 h; or 5, 10, 20 and 40
µM for 12 h) and mitochondrial membrane potential was
determined by JC-1 staining and fluorescence microscopy (scale bar,
10 µm), and was quantified by fluorescence spectrometry
(red/green fluorescence ratio). (C and D) U251 cells were exposed
to Adpa-Mn (20 µM for 6, 12 and 24 h; or 5, 10 and 20
µM for 12 h) and reactive oxygen species generation was
detected by 2′,7′-dichlorofluorescein-diacetate (DCFH-DA) staining
(scale bar, 10 µm). (E) U251 cells were exposed to 20
µM Adpa-Mn for 6, 12 and 24 h, and adenosine triphosphate
(ATP) levels were measured using the luciferin-luciferase assay.
Data are presented as the mean ± standard deviation of three
independent experiments. *P<0.05,
**P<0.01 vs. control (0 µM/0 h Adpa-Mn). |
Adpa-Mn induces cell death dependent on
mitochondrial dysfunction
Due to the finding that Adpa-Mn triggers
mitochondrial membrane potential breakdown and ROS generation, the
present study aimed to elucidate the association between ROS,
mitochondria and Adpa-Mn-induced cell death (Fig. 5). Pretreatment with CsA (2
µM), an inhibitor of the mitochondrial permeability
transition pore (MPTP) and NAC, a ROS scavenger, reduced
Adpa-Mn-induced ROS production (Fig.
5A), whereas only CsA pretreatment was able to reverse
Adpa-Mn-induced membrane potential collapse (Fig. 5B). Furthermore, MTT assay and
Annexin V/PI staining indicated that cell death or apoptosis
triggered by Adpa-Mn was significantly reduced by CsA pretreatment,
but was aggravated to some extent by NAC pretreatment (Fig. 5C and E). In addition, CsA and NAC
were able to reduce LC3 II expression (Fig. 5D). These findings indicate that
Adpa-Mn may disrupt the mitochondrial membrane to induce apoptosis
and protective autophagy in U251 cells.
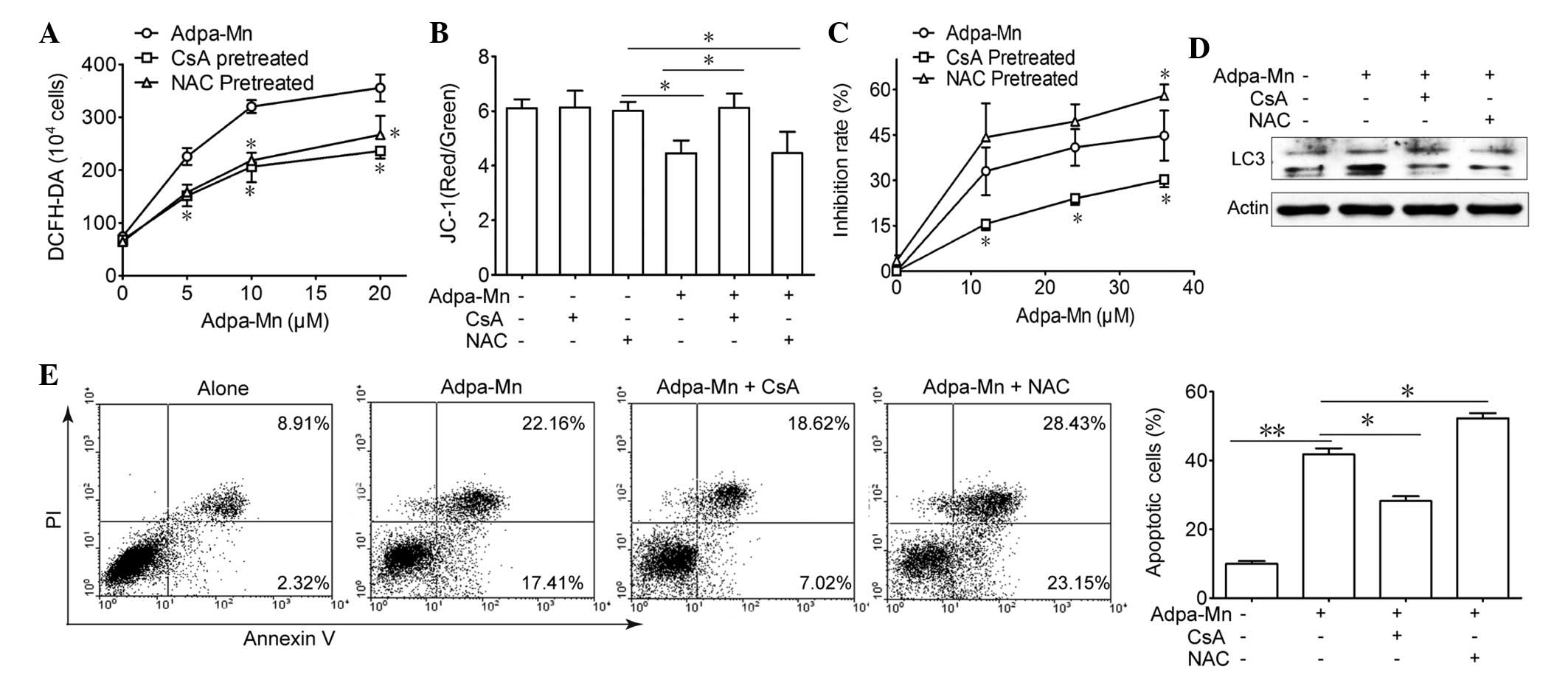 | Figure 5Cell death induced by Adpa-manganese
(Mn) treatment was dependent on mitochondrial dysfunction. (A) U251
cells were exposed to 5, 10, 15 and 20 µM Adpa-Mn for 24 h
with or without N-acetylcysteine (NAC; 5 mM) or cyclosporin A (CsA;
2 µM) pretreatment (2 h). Reactive oxygen species (ROS)
levels were examined by 2′,7′-dichlorofluorescein-diacetate
(DCFH-DA) staining. (B) U251 cells were exposed to 20 µM
Adpa-Mn for 12 h with or without NAC (5 mM) or CsA (2 µM)
pretreatment (2 h), and mitochondrial membrane potential was
evaluated by JC-1 staining. (C) U251 cells were exposed to 10, 20,
30 and 40 µM Adpa-Mn for 24 h with or without NAC (5 mM) or
CsA (2 µM) pretreatment (2 h), and cell viability was
examined by 3-[4,5-dimehyl-2-thiazolyl]-2,5-diphenyl-2H-tetrazolium
bro mide assay. (D) U251 cells were exposed to 20 µM Adpa-Mn
for 12 h with or without NAC (5 mM) or CsA (2 µM) (2 h), and
microtubule-associated protein 1 light chain 3 (LC3) expression was
evaluated by western blotting. (E) U251 cells were exposed to 20
µM Adpa-Mn for 24 h with or without NAC (5 mM) or CsA (2
µM) pretreatment (2 h), and apoptosis was evaluated by
Annexin V/propidium iodide (PI) staining. Data are presented as the
mean ± standard deviation of three independent experiments.
*P<0.05 and **P<0.01 vs. Adpa-Mn. |
Discussion
The success of cisplatin in the treatment of
patients with cancer has resulted in the hypothesis that other
metal complexes may be considered as potential drugs in future
chemotherapy regimens. The present study aimed to verify whether
the designed Mn-compound, Adpa-Mn, may be used as an anti-glioma
lead compound. In the present study, Adpa-Mn was demonstrated to be
active against glioma cells, whereas it had little effect on normal
astrocytes. In addition, Adpa-Mn time and dose-dependently induced
mitochondrial dysfunction, as determined by mitochondrial collapse
and ROS generation, finally resulting in apoptotic cell death.
Apoptosis is a type of cell death that is
characterized by nuclear condensation and fragmentation, and
apoptotic body emergence without plasma membrane breakdown. During
apoptosis, numerous death signals converge on mitochondria,
particularly the release of cytochrome c as a consequence of
the increased permeability of the outer mitochondrial membrane,
which subsequently activates downstream caspase signaling (25). In the present study, Adpa-Mn was
revealed to induce apoptosis mediated by the mitochondrial pathway
(Fig. 2).
Autophagy is a dynamic process, which is often
termed autophagic flux, whereby autophagosomes are formed in
response to stimuli, engulf the cellular content and damaged
organelles, and fuse with lysosomes; the contents of the
autophagosome are subsequently degraded (26). Autophagy has previously been
referred to as a physiological process that has a protective role
in cells that encounter environmental stress, including starvation
and pathogenic infection (27).
Excess autophagy can also act as a pro-death mechanism; therefore,
it has been classified as type II programmed cell death or
autophagic cell death (28). In
U251 cells, the following characteristics of autophagy were
detected following Adpa-Mn treatment: Formation of AVOs and
elevated ratio of LC3-II to LC3-I. When combined with the autophagy
inhibitors, CQ and 3-MA, cell viability was markedly inhibited
(Fig. 3). These results suggested
that autophagy may have a protective role during Adpa-Mn treatment,
which is similar to what has been reported for other chemicals
(29,30).
ROS are predominantly generated from mitochondria
and have a central role in cell death processes, including
apoptosis and autophagy (31,32).
Under excessive oxidative stress, the accumulation of ROS reaches a
threshold level that triggers opening of the MPTP or oxidation of
the mitochondrial outer membrane, which in turn leads to the
simultaneous collapse of mitochondrial membrane potential and a
transient increase in ROS generation by the respiratory chain that
transforms O2 into ATP (33,34).
Adpa-Mn was able to induce mitochondrial dysfunction, including
mitochondrial membrane potential collapse, ROS accumulation and ATP
depletion. Conversely, pretreatment with NAC and CSA significantly
inhibited Adpa-Mn-induced ROS accumulation. In addition, NAC and
CSA hampered Adpa-Mn-induced autophagy, whereas CsA decreased, but
NAC aggravated, apoptosis (Fig.
5). CsA was also able to prevent Adpa-Mn-induced mitochondrial
membrane potential collapse, whereas NAC could not. These data
suggested that Adpa-Mn treatment destroyed mitochondria, thus
leading to apoptosis of U251 cells; however, ROS that originated
from the damaged mitochondria triggered protective autophagy.
In our previous study, the Adpa-Mn complex exhibited
high toxicity on cancer cell lines, but showed weak DNA binding and
cleavage activity (20). In
addition, Adpa-Mn was shown to induce apoptotic cell death of HeLa
cells (35). Conversely, whereas
Adpa-Mn-induced autophagy in HeLa cells acted as a form of cell
death, it was protective in U251 cells. These findings suggested
that there may be different mechanisms underlying autophagy
regulation in HeLa and U251 cells, which require further
investigation.
In conclusion, the present study demonstrated that
Adpa-Mn exhibited selective inhibition on glioma cell proliferation
coupled with mitochondria-mediated apoptosis and increased
autophagy. In addition, Adpa-Mn-induced autophagy exerted
protective effects on glioma cells. Therefore, Adpa-Mn alone, or
combined with autophagy inhibitors, may be considered a novel
option for the treatment of glioma.
Acknowledgments
The authors of the present would you like to thank
the following: The National Natural Science Foundation of China
(grant no. 81402938); the Natural Science Foundation of Jiangsu
Province (grant nos. BK2012710 and 20140575); and the Grant of
Jiangsu University (grant no. 13JDG064).
References
|
1
|
Walker MD, Green SB, Byar DP, Alexander E
Jr, Batzdorf U, Brooks WH, Hunt WE, MacCarty CS, Mahaley MS Jr,
Mealey J Jr, et al: Randomized comparisons of radiotherapy and
nitrosoureas for the treatment of malignant glioma after surgery. N
Engl J Med. 303:1323–1329. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Subach BR, Witham TF, Kondziolka D,
Lunsford LD, Bozik M and Schiff D: Morbidity and survival after 1,
3-bis (2-chloroethyl)-1-nitrosourea wafer implantation for
recurrent glioblastoma: A retrospective case-matched cohort series.
Neurosurgery. 45:17–23. 1999. View Article : Google Scholar
|
|
3
|
Chamberlain MC and Kormanik P: Salvage
chemotherapy with taxol for recurrent anaplastic astrocytomas. J
Neurooncol. 43:71–78. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Attardi G and Schatz G: Biogenesis of
mitochondria. Annu Rev Cell Biol. 4:289–333. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bender A, Opel D, Naumann I, Kappler R,
Friedman L, von Schweinitz D, Debatin KM and Fulda S: PI3K
inhibitors prime neuroblastoma cells for chemotherapy by shifting
the balance towards pro-apoptotic Bcl-2 proteins and enhanced
mitochondrial apoptosis. Oncogene. 30:494–503. 2011. View Article : Google Scholar
|
|
6
|
Susin SA, Lorenzo HK, Zamzami N, Marzo I,
Snow BE, Brothers GM, Mangion J, Jacotot E, Costantini P, Loeffler
M, et al: Molecular characterization of mitochondrial
apoptosis-inducing factor. Nature. 397:441–446. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nakahira K, Haspel JA, Rathinam VA, Lee
SJ, Dolinay T, Lam HC, Englert JA, Rabinovitch M, Cernadas M, Kim
HP, et al: Autophagy proteins regulate innate immune responses by
inhibiting the release of mitochondrial DNA mediated by the NALP3
inflammasome. Nat Immunol. 12:222–230. 2011. View Article : Google Scholar
|
|
8
|
Mathew R and White E: Autophagy in
tumorigenesis and energy metabolism: Friend by day, foe by night.
Curr Opin Genet Dev. 21:113–119. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
He C and Klionsky DJ: Regulation
mechanisms and signaling pathways of autophagy. Annu Rev Genet.
43:67–93. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kimura T, Takabatake Y, Takahashi A and
Isaka Y: Chloroquine in cancer therapy: A double-edged sword of
autophagy. Cancer Rese. 73:3–7. 2013. View Article : Google Scholar
|
|
11
|
Williams R: Free manganese (II) and iron
(II) cations can act as intracellular cell controls. FEBS Lett.
140:3–10. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ansari KI, Kasiri S, Grant JD and Mandal
SS: Apoptosis and anti-tumour activities of manganese(III)-salen
and -salphen complexes. Dalton Trans. 8525–8531. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kono Y and Fridovich I: Isolation and
characterization of the pseudocatalase of Lactobacillus plantarum.
J Biol Chem. 258:6015–6019. 1983.PubMed/NCBI
|
|
14
|
Law NA, Caudle T and Pecoraro VL:
Manganese redox enzymes and model systems: Properties, structures
and reactivity. Adv Inorg Chem. 46:305–440. 1998. View Article : Google Scholar
|
|
15
|
Luk E, Carroll M, Baker M and Culotta VC:
Manganese activation of superoxide dismutase 2 in Saccharomyces
cerevisiae requires MTM1, a member of the mitochondrial carrier
family. Proc Natl Acad Sci USA. 100:10353–10357. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Calzolari A, Oliviero I, Deaglio S,
Mariani G, Biffoni M, Sposi NM, Malavasi F, Peschle C and Testa U:
Transferrin receptor 2 is frequently expressed in human cancer cell
lines. Blood Cells Mol Dis. 39:82–91. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Aschner M and Aschner JL: Manganese
transport across the blood-brain barrier: Relationship to iron
homeostasis. Brain Res Bull. 24:857–860. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Aschner M and Gannon M: Manganese (Mn)
transport across the rat blood-brain barrier: Saturable and
transferrin-dependent transport mechanisms. Brain Res Bull.
33:345–349. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li SJ, Jiang L, Fu X, Huang S, Huang YN,
Li XR, Chen JW, Li Y, Luo HL, Wang F, et al: Pallidal index as
biomarker of manganese brain accumulation and associated with
manganese levels in blood: A meta-analysis. PloS One. 9:e939002014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen QY, Huang J, Li JF and Gao J:
Synthesis, interaction with mitochondrial and cancer cells of a
dinuclear manganese(II) complex:
Mn2(Adpa)2Cl4. Chinese J Inorg
Chem. 24:1789–1793. 2008.In Chinese.
|
|
21
|
McCarthy KD and de Vellis J: Preparation
of separate astroglial and oligodendroglial cell cultures from rat
cerebral tissue. J Cell Biol. 85:890–902. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lemasters JJ, Qian T, He L, Kim JS, Elmore
SP, Cascio WE and Brenner DA: Role of mitochondrial inner membrane
permeabilization in necrotic cell death, apoptosis, and autophagy.
Antioxid Redox Signal. 4:769–781. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tanida I, Minematsu-Ikeguchi N, Ueno T and
Kominami E: Lysosomal turnover, but not a cellular level, of
endogenous LC3 is a marker for autophagy. Autophagy. 1:84–91. 2005.
View Article : Google Scholar
|
|
24
|
Eisenberg-Lerner A, Bialik S, Simon HU and
Kimchi A: Life and death partners: Apoptosis, autophagy and the
cross-talk between them. Cell Death Differ. 16:966–975. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Desagher S and Martinou JC: Mitochondria
as the central control point of apoptosis. Trends Cell Biol.
10:369–377. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Thorburn A: Autophagy and its effects:
Making sense of double-edged swords. PLoS Biol. 12:e10019672014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Klionsky DJ and Emr SD: Autophagy as a
regulated pathway of cellular degradation. Science. 290:1717–1721.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Galluzzi L, Maiuri MC, Vitale I, Zischka
H, Castedo M, Zitvogel L and Kroemer G: Cell death modalities:
Classification and pathophysiological implications. Cell Death
Differ. 14:1237–1243. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Han J, Hou W, Goldstein LA, Lu C, Stolz
DB, Yin XM and Rabinowich H: Involvement of protective autophagy in
TRAIL resistance of apoptosis-defective tumor cells. J Biol Chem.
283:19665–19677. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li J, Hou N, Faried A, Tsutsumi S,
Takeuchi T and Kuwano H: Inhibition of autophagy by 3-MA enhances
the effect of 5-FU-induced apoptosis in colon cancer cells. Ann
Surg Oncol. 16:761–771. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Scherz-Shouval R, Shvets E, Fass E, Shorer
H, Gil L and Elazar Z: Reactive oxygen species are essential for
autophagy and specifically regulate the activity of Atg4. EMBO J.
26:1749–1760. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Scherz-Shouval R and Elazar Z: ROS,
mitochondria and the regulation of autophagy. Trends Cell Biol.
17:422–427. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Garlid KD and Beavis AD: Evidence for the
existence of an inner membrane anion channel in mitochondria.
Biochim Biophys Acta. 853:187–204. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zorov DB, Juhaszova M and Sollott SJ:
Mitochondrial ROS-induced ROS release: An update and review.
Biochim Biophys Acta. 1757:509–517. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu J, Guo W, Li J, Li X, Geng J, Chen Q
and Gao J: Tumor-targeting novel manganese complex induces
ROS-mediated apoptotic and autophagic cancer cell death. Int J Mol
Med. 35:607–616. 2015.PubMed/NCBI
|