Introduction
Premature ovarian failure (POF) refers to the
presence of ovarian atrophy-associated permanent amenorrhea in
women under 40 (1,2). The levels of follicle stimulating
hormone (FSH) are significantly increased in peripheral blood while
the levels of estradiol (E2) are significantly decreased (1,3).
Histopathological examination of ovarian tissues has revealed
significant atrophy and fibrosis, marked follicular atresia and the
absence of normal follicular stages (including primordial follicle,
antral follicle, cumulus oophorus and mature egg) (3–5). The
underlying mechanisms of POF development are diverse and complex.
Currently, the pathogenesis of POF remains to be elucidated, and no
effective treatment measures exist.
Telocytes are a group of highly specialized cells in
the connective tissues of organs (6–8).
Telocytes have a small cell volume and long cytoplasmic
prolongations (typically 10–100 µm in length), referred to
as telopodes (9–11). The dichotomous branching structure
of telopodes may be observed by electron microscopy (12–14),
which reveals that telopodes consist of thick branching segments
(podoms) and thin branching segments (podomers) (15,16).
It has been demonstrated that telocytes are present in the
connective tissues of numerous organs in mammals, including
myocardial tissue (6,9,11,12),
uterus and fallopian tubes (10,11,16),
placental tissues (11), ureter
(10,11,17),
skeletal muscle (11,17,18),
lung (9,11), mammary gland (11), liver (11,19–21)
and pancreas (11,14). Although no telocyte-specific marker
has been identified to date, previous studies have reported
increased expression of cluster of differentiation (CD)34, CD117
(c-kit), platelet-derived growth factor receptor (PDGFR)α, PDGFRβ
and vimentin (6–22). Previous studies have demonstrated
that telocytes have the following biological functions: i)
Telocytes may guide the migration of cardiac stromal cells from the
endothelium to mesothelium and contribute to vascular regeneration
of myocardial tissues following myocardial infarction (6,8,11,12);
ii) telocytes form a complex network structure in the lung via
their telopodes that support the bronchial lumens of lobules, thus
preventing obstruction of lobules and small blood vessels with weak
vascular walls during the respiratory process (9,12);
iii) telocytes affect antigen presentation, immune activation and
immune tolerance, via the release of microvesicles during synapses
with immune cells (8); and iv) as
'stem cell helper cells', telocytes facilitate repair and
regeneration at injured sites through cross-linking to form stem
cell niches (11,12,16,20).
However, whether telocytes are present in the mammalian ovary and
contribute to the regulation of ovarian function remains to be
determined (11). In the present
study, electron microscopy combined with immunofluorescence
analysis and flow cytometry was performed to investigate the
hypothesis that telocytes are present in ovarian tissues in mice.
In addition, telocytes were investigated as a potential novel
marker of POF in mice induced by cyclophosphamide.
Materials and methods
Establishment of a POF mouse model
POF was induced in mice according to a previously
described method (3–5). Briefly, 8-week-old female C57BL/6
mice (n=40) were purchased from the Experimental Animal Center of
Shanghai University of Traditional Chinese Medicine (Shanghai,
China). Mice were housed in a temperature-controlled environment
under standard light-dark cycles with ad libitum access to
food and water as previously described (3). Mice were randomly divided into two
groups (20 mice/group). Mice in the POF group were injected
intraperitoneally with 70 mg/kg cyclophosphamide (Sigma-Aldrich,
St. Louis, MO, USA), followed by intraperitoneal injections of 30
mg/kg cyclophosphamide every three days for three weeks, to
establish the POF mouse model. Mice in the control group were
injected intraperitoneally with an equal volume of normal saline
every three days for three weeks. All mice were sacrificed by
cervical dislocation at three weeks and the ovaries were harvested
for subsequent analysis. The study was approved by the Ethics
Committee of Shanghai Geriatric Institute of Chinese Medicine
(Shanghai, China; ref. SHAGESYDW2015026). All experiments conformed
to standards set by the Laboratory Animal Regulation of the State
Scientific and Technological Commission (www.slarc.org.cn).
Hematoxylin and eosin (H&E)
staining
H&E staining was performed according to a
previously described method (3).
Briefly, fresh tissues were immersed in 4% paraformaldehyde
(Sigma-Aldrich) for 30 min at room temperature to allow fixation.
Tissues were dehydrated using an ethanol gradient, embedded in
paraffin, sectioned (thickness, 6 µm) and deparaffinized in
xylene (Sigma-Aldrich). Tissue sections were stained with H&E
(Sigma-Aldrich), cleared in xylene and mounted on slides using
neutral balsam (Sigma-Aldrich). The number of atretic follicles was
counted per microscope field, in three non-overlapping fields of
ovary sections from each mouse.
Immunofluorescence staining
Immunofluorescence staining was performed as
previously described (4). Briefly,
fresh tissues were fixed and sectioned as for H&E staining.
Tissue sections were incubated with immunofluorescence blocking
solution (Beyotime Institute of Biotechnology, Haimen, China) for
30 min at 37°C. Subsequently, sections were washed three times in
immunofluorescence wash solution (Beyotime Institute of
Biotechnology) for 5 min at room temperature. Sections were
incubated with primary antibodies (Table I) for 45 min at 37°C. The washing
step was repeated, and sections were incubated with secondary
antibodies (Table II) for 45 min
at 37°C. Following a final repeat of the washing step, sections
were mounted using Kisser's Mounting Medium (Beyotime Institute of
Biotechnology).
 | Table IPrimary antibodies. |
Table I
Primary antibodies.
| Target | Species raised
in | Species raised
against | Application | Dilution | Catalog no. | Supplier |
|---|
| Inhibin α | Rabbit | Mouse | IHC | 1:100 | sc-30146 | SC |
| Inhibin β | Rabbit | Mouse | IHC | 1:100 | sc-50288 | SC |
| AMH | Rabbit | Mouse | IHC | 1:100 | ab103233 | AB |
| FSHR | Rabbit | Mouse | IHC | 1:100 | sc-13935 | SC |
| PDGFRα | Rabbit | Mouse | IF, FC | 1:100 | sc-431 | SC |
| PDGFRβ | Rabbit | Mouse | IF, FC | 1:100 | sc-432 | SC |
| CD34 | Goat | Mouse | IF | 1:100 | sc-7045 | SC |
| c-Kit | Rabbit | Mouse | IF, FC | 1:100 | sc-5535 | SC |
| Vimentin | Rat | Mouse | IF, FC | 1:100 | sc-5565 | SC |
| Table IISecondary antibodies. |
Table II
Secondary antibodies.
| Species raised
in | Species raised
against | Conjugated to | Application | Dilution | Catalog no. | Supplier |
|---|
| Donkey | Goat | Cy3 | IF, FC | 1:100 | ab6949 | AB |
| Goat | Rabbit | FITC | IF, FC | 1:100 | sc-2777 | SC |
| Goat | Rat | FITC | IF, FC | 1:100 | sc-2011 | SC |
| Donkey | Rabbit | HRP | IHC | 1:300 | ab205722 | AB |
Immunohistochemical staining
Immunohistochemical staining was performed as
previously described (4). Briefly,
ovarian tissue samples were washed three times with
phosphate-buffered saline (PBS; Sigma-Aldrich), fixed with 4%
paraformaldehyde (Sigma-Aldrich) for 30 min, dehydrated through a
graded series of ethanol, cleared in xylene and embedded in
paraffin. Subsequently, serial 6-µm thick sections were cut,
rinsed with 3% phosphate buffer (Sigma-Aldrich), and subjected to
heat retrieval in a microwave. The sections were then incubated
with primary antibodies (Table I)
for 45 min at 37°C. Subsequently, sections were washed three times
in immunohistochemistry wash solution (Beyotime Institute of
Biotechnology) for 5 min at room temperature. Sections were
incubated with secondary antibodies (Table II) for 45 min at 37°C. Following a
repeat of the washing step, ABC chromogenic reagent (Sigma-Aldrich)
was added to visualize bound antibody. PBS (pH 7.4) was used as a
negative control for primary antibody. Sections were mounted using
neutral balsam (Beyotime Institute of Biotechnology). Five randomly
selected fields of view (magnification, ×200; Olympus BX43; Olympus
Corporation, Tokyo, Japan) were assessed for each tissue section
and analyzed using Integrated Performance Primitives software
version 4.0 (Intel Corporation, Santa Clara, CA, USA).
Enzyme-linked immunosorbent assay
(ELISA)
Mouse blood was obtained by retro-orbital blood
collection at sacrifice (~100 µl blood from each mouse) and
centrifuged at 453 × g at 4°C for 10 min. The supernatants were
collected and these serum samples were used for ELISA. ELISAs were
performed according to the manufacturer's instructions for the
mouse E2 and FSH ELISA kits (Westang Bio-Tech Co., Ltd., Shanghai,
China). Briefly, 100 µl serum, along with mouse E2 standards
(at concentrations of 8,000, 4,000, 2,000, 1,000, 500, 250 and 125
pg/ml) and mouse FSH standards (at concentrations of 10, 5, 2.5,
1.25, 0.625, 0.312 and 0.156 ng/ml), was added to pre-coated
96-well detection plates and incubated for 60 min at 37°C. The
solution was discarded and the plates were washed three times with
wash solution. Horseradish peroxidase-conjugated detection
antibodies were added and incubated for 60 min at 37°C.
Subsequently, stop solution was added and the optical density of
each well in the 96-well plate was measured at a wavelength of 450
nm.
Flow cytometry
Cells were isolated and flow cytometry was performed
according to a previously described method (5,10).
Briefly, the murine ovaries were minced into small pieces (~1
mm3) and washed three times with PBS at 4°C. The samples
were digested using 10 mg/ml collagenase type I (Sigma-Aldrich) and
0.1% hyaluronidase (Sigma-Aldrich) in PBS for 5 min on an orbital
shaker at 37°C, and filtered through 40-µm diameter cell
strainers. These granulosa cells from ovaries were fixed in 4%
paraformaldehyde for 30 min at room temperature. Following
centrifugation at 453 × g for 5 min at 37°C, the supernatant was
discarded and the cell precipitate was resuspended in blocking
solution (Beyotime Institute of Biotechnology) for 30 min at 37°C.
Cells were centrifuged, the blocking solution was discarded, and
primary antibodies (Table I) were
added and incubated for 45 min at 37°C. Following centrifugation,
the antibodies were discarded and wash solution (Beyotime Institute
of Biotechnology) was added to the cells for 5 min at room
temperature. Cells were then centrifuged, and secondary antibodies
(Table II) were added and
incubated for 30 min at 37°C. Following centrifugation, the
antibodies were discarded and wash solution (Beyotime Institute of
Biotechnology) was added to the cells for 5 min at room
temperature. Positive cells were detected by flow cytometry (Quanta
SC; Beckman Coulter, Inc., Brea, CA, USA).
Preparation of samples and observation
using electron microscopy
Samples were fixed and embedded according to a
previous study (23) and the
protocol provided by KingMed Center for Clinical Laboratory Co.,
Ltd. (Guangzhou, China). Tissue samples were fixed in 1%
glutaraldehyde (Sigma-Aldrich) for 4 h followed by 1% osmium
tetroxide (Sigma-Aldrich) for 1 h. Samples were then dehydrated in
acetone and embedded in Eponate 12 resin (Ted Pella, Inc., Redding,
CA, USA). Samples were cut into ultrathin sections (thickness, 70
nm), placed on copper grids and stained using 1% uranyl acetate and
1% lead citrate (Sigma-Aldrich). The sections were observed and
images captured using a JEM-1230 transmission electron microscope
(JEOL, Ltd., Tokyo, Japan).
Statistical analysis
Statistical analyses were performed in GraphPad
Prism software version 5.00 (GraphPad Software, Inc., La Jolla, CA,
USA). The data are presented as the mean ± standard error where
applicable; differences were evaluated using Student's
t-test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Cyclophosphamide induces ovarian damage
and abnormal hormone secretion in mice
Histological analysis with H&E staining revealed
that the number of atretic follicles in the ovaries of
cyclophosphamide-treated mice was significantly increased compared
with the control group (P=0.018; Fig.
1A). In addition, the texture of ovarian tissues was dense in
control mice. Various stages of follicles were observed in ovaries,
including primordial follicles (Fig.
1B; pink arrows), antral follicles (Fig. 1B; red arrows), cumulus oophorus
(Fig. 1B; blue arrows) and mature
eggs (Fig. 1B; black arrows).
Following treatment with cyclophosphamide for three weeks, ovarian
tissues demonstrated significant atrophy, the ovarian stroma had
empty space, and apoptosis and necrosis in ovarian granulosa cells
was significant (Fig. 1B; yellow
arrows). In addition, it was difficult to identify the various
stages of follicles and mature eggs, and atretic follicles and
luteal tissue atrophy were abundant (Fig. 1B). Immunohistochemistry results
(Fig. 1C) indicated that the
expression of the ovarian granulosa cell markers anti-Müllerian
hormone (AMH) and inhibinα/β in the ovaries of POF mice was reduced
compared with control mice; however, no difference was observed in
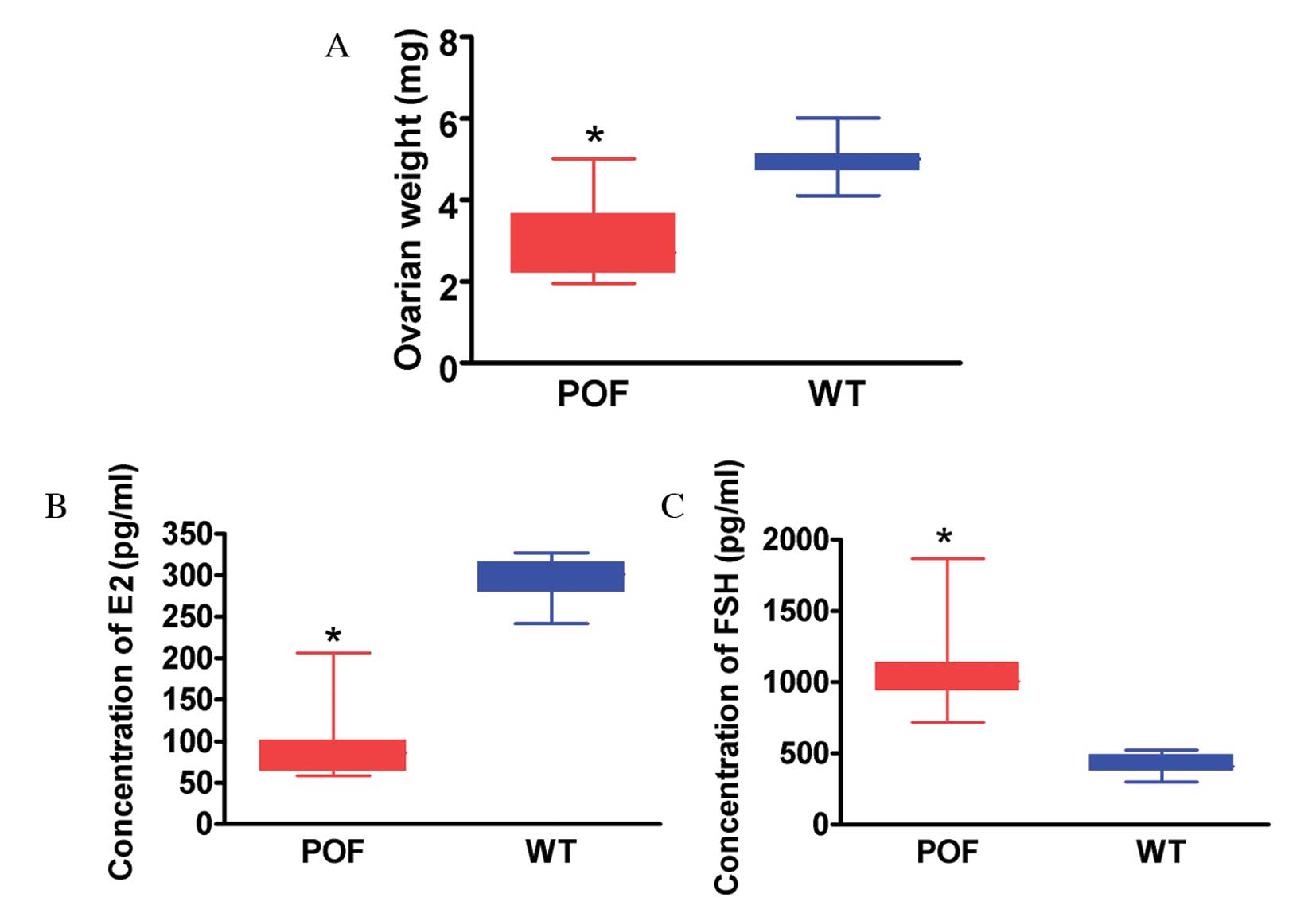
FSH receptor (FSHR) staining between the two groups. The weight of
ovarian tissues from cyclophosphamide-treated mice was
significantly reduced compared with that of control mice (P=0.022;
Fig. 2A). ELISA results
demonstrated that the peripheral blood E2 level in mice treated
with cyclophosphamide was significantly reduced compared with the
control group (P=0.015; Fig. 2B),
and the FSH level was significantly increased compared with the
control group (P=0.027; Fig. 2C).
Histological and hormone level detection results therefore
suggested that cyclophosphamide induced ovarian granulosa cell
injury and apoptosis as well as the development of POF.
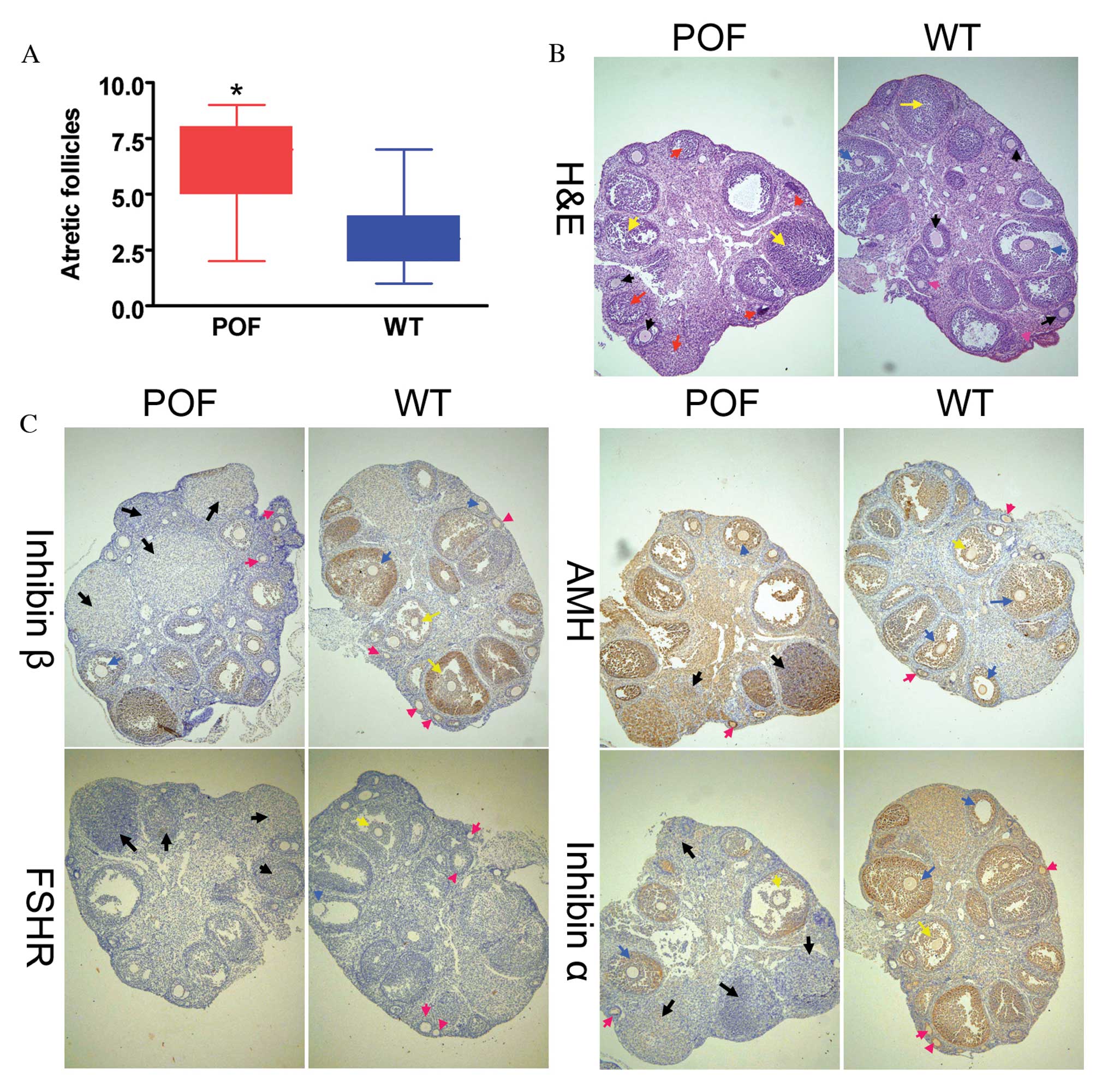 | Figure 1Histological analysis of ovaries. (A)
The number of atretic follicles in the ovarian tissues of POF mice
was significantly increased compared with control mice.
*P<0.05 vs. WT; n=10. (B) H&E staining of ovarian
tissues. Granulosa cells in the ovarian tissues of POF mice
demonstrated clear edema and apoptosis, and increased atretic
follicles. Ovarian granulosa cell injury is indicated by yellow
arrows, primordial follicles by pink arrows, antral follicles by
red arrows, mature oocytes by black arrows and cumulus oophorus by
blue arrows. (C) Immunohistochemical staining revealed that in
ovarian tissues of POF mice, the expression of the ovarian
granulosa cell markers AMH, inhibinα and inhibinβ was reduced
compared with those of control mice. No difference was observed in
the expression of FSHR between the two groups. Black arrows
indicate atretic follicles, pink arrows indicate primordial
follicles, yellow arrows indicate cumulus oophorus and blue arrows
indicate mature oocytes. Magnification, ×100. H&E, hematoxylin
and eosin; POF, premature ovarian failure; WT, wild-type; AMH,
anti-Müllerian hormone; FSHR, follicle stimulating hormone
receptor. |
Telocytes are present in normal ovarian
tissues
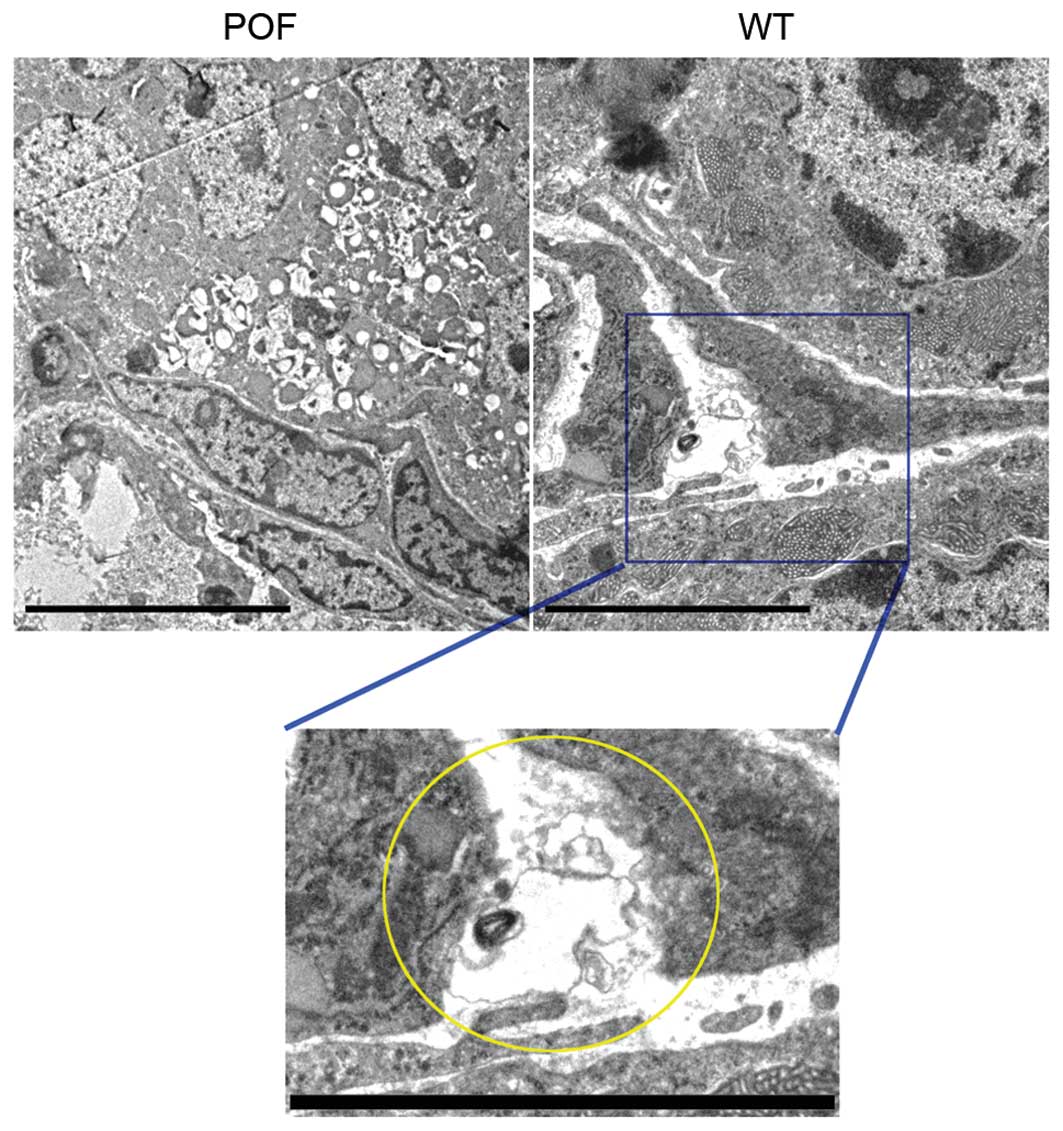
Transmission electron microscopy was performed to
confirm that telocytes were present in ovarian tissues. A unique
type of cell body was observed in the ovarian stroma in control
mice. Although the cell volume was small, it had very long
telopodes, which had a typical dichotomous branching structure and
alternating thick and thin segments (Fig. 3). However, cells with these
morphological features were not observed in ovarian tissues from
POF mice. Following a review of the literature, it was hypothesized
that these cells were telocytes.
Cyclophosphamide induces a reduction in
telocyte number and marker expression levels
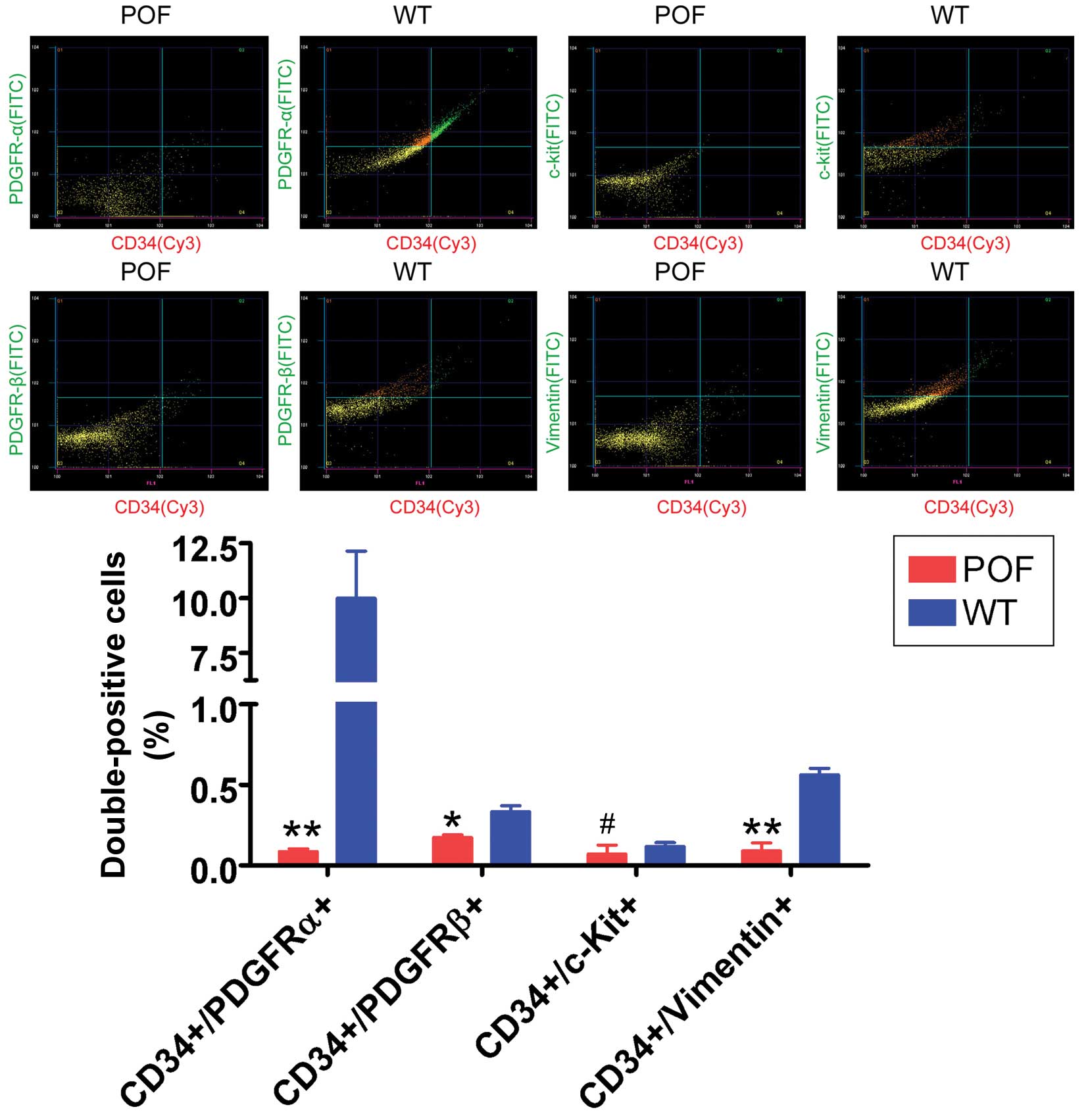
Flow cytometry detection of telocyte markers
revealed that the number of CD34/PDGFRα, CD34/PDGFRβ and
CD34/vimentin double-positive cells in the ovaries of POF mice was
significantly reduced compared with control mice (P=0.031; Fig. 4), whereas the number of CD34/c-kit
double-positive cells was not significantly different between the
two groups (P=0.061; Fig. 4). In
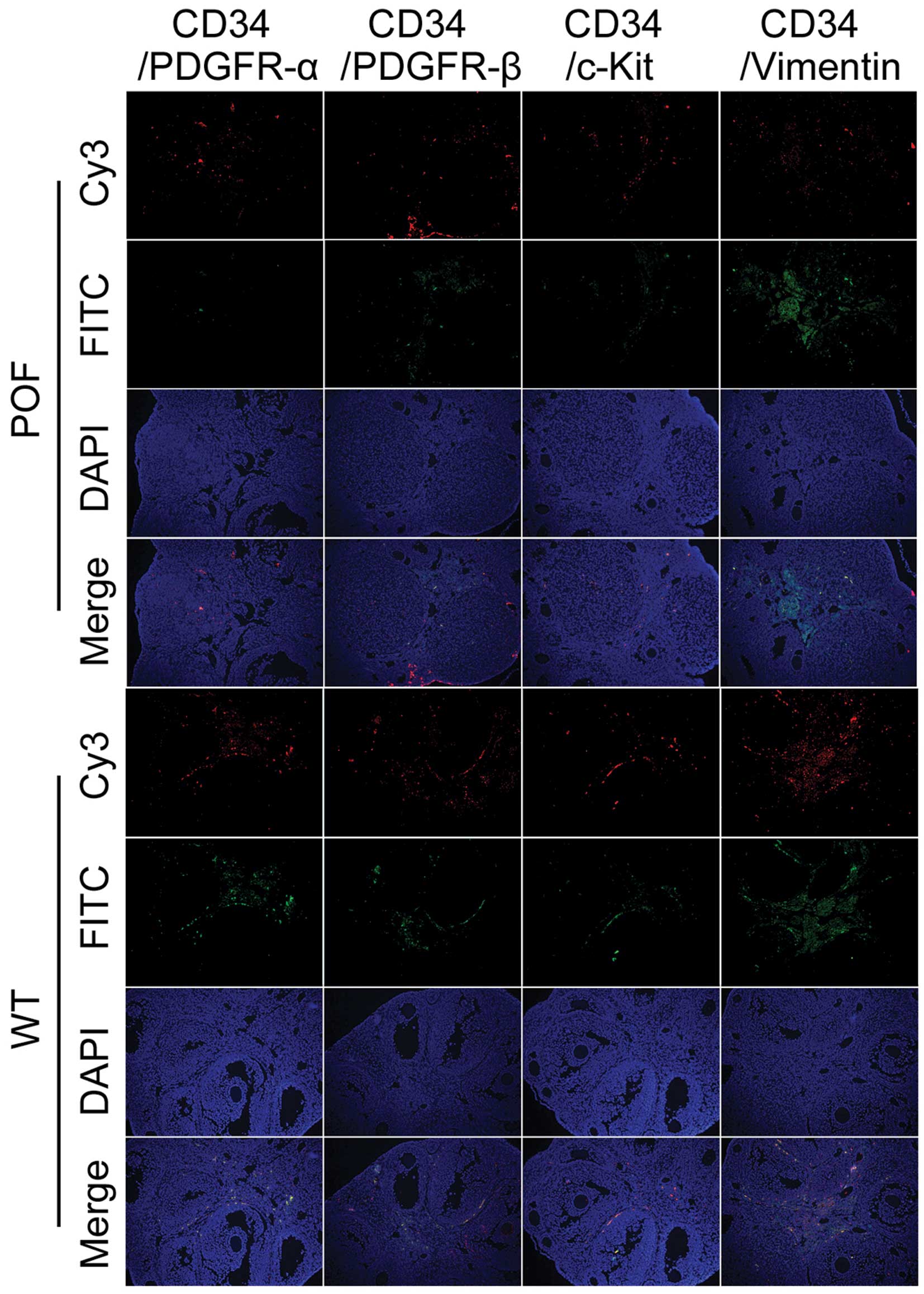
addition, immunofluorescence staining results indicated that the
number of CD34/PDGFRα, CD34/PDGFRβ and CD34/vimentin
double-positive cells in the ovarian tissues of POF mice was
decreased compared with control mice (Fig. 5). These results suggested that the
number of telocytes significantly decreased in ovarian tissues of
mice following cyclophosphamide treatment.
Discussion
POF is associated with complex risk factors and high
morbidity (2). Typically, studies
have focused on the effects of risk factors, including genetic
variations, immunity and inflammation, environmental hormones and
contamination, personal stress, and endocrine disorders (1,2).
During the development of POF, lesions are most prominent in egg
and ovarian granulosa cells (4,5,24).
Ovarian granulosa cells, similar to Sertoli cells of the male
reproduction system, support female reproductive cells i.e. mature
egg cells (3), surrounding these
cells during follicle development to form a unique
microenvironment. Ovarian granulosa cells secrete hormones and
cytokines for egg maturation, including estrogen, FSH,
anti-Müllerian antibody, inhibinα/β and growth hormone, to support
the development, maturation and ovulation of follicles (3). Ovarian granulosa cell apoptosis
reduces the follicular reserve capacity of the ovary, thus causing
the development of POF (1,5). In the ovaries of POF patients,
ovarian granulosa cells typically exhibit apoptosis and edema
(4). The primary function of the
ovary is to provide a location for egg maturation. Therefore, in
the ovaries of POF patients, the number of atretic follicles is
significantly increased and the number of healthy follicles is
significantly decreased; in addition, the number of follicles at
various stages is significantly decreased (2). As well as egg and ovarian granulosa
cells, other types of cell, including ovarian stromal cells,
ovarian epithelial cells and ovarian microvascular endothelial
cells are affected by POF; however, there have been limited studies
on their functions.
A literature search revealed reports of a unique
cell type present in the respiratory, cardiovascular and digestive
systems; these cells were telocytes (6–22).
Telocytes are predominantly localized in the connective tissues of
organs. Telocyte features include a small cell volume, very long
cytoplasmic prolongations (telopodes) and a typical dichotomous
branching structure of these telopodes (6–22).
Telopodes comprise a complex network structure, which supports the
lumen of lobular bronchioles and blood vessels to prevent
obstruction. In addition, as 'stem cell helper cells', telocytes
form stem cell niches through cross-linking to facilitate the
repair and regeneration of injured sites (6–22).
However, in mammals, not every organ contains telocytes and it has
not been reported whether they are present in ovaries.
As telocytes function to maintain the
microenvironment, it was hypothesized that ovaries may contain
telocytes. Therefore, transmission electron microscopy was
performed to analyze the ultra-microstructure of ovarian tissues
from control and POF mice. Very long cells were observed in the
stroma of healthy mouse ovaries. The somatic synapses of these
cells revealed a triangular shape and very long and thin
prolongations with a typical dichotomous branching structure,
suggesting that these cells may be ovarian tissue telocytes.
However, cells with the above morphological features were not
observed in the ovaries of POF mice. Subsequently, the expression
levels of the telocyte-specific markers CD34, CD117, PDGFRα/β and
vimentin were evaluated in ovarian tissues. The results of flow
cytometry and immunofluorescence suggested that the number of
CD34/PDGFRα, CD34/PDGFRβ and CD34/vimentin double-positive cells in
ovarian cells of wild-type mice was significantly increased
compared with POF mice. In addition, immunohistochemical staining
and flow cytometry experiments suggested that various telocyte
markers were expressed in the ovarian tissues of control mice.
However, when mice were treated with cyclophosphamide, the
expression of telocytes and their markers decreased. Based on the
results of the present study, telocytes in ovarian tissues may have
an important function in maintaining the ovarian microenvironment.
The ovarian tissue injury induced by cyclophosphamide was not
confined to ovarian granulosa cells and eggs, as ovarian telocytes
were also affected.
In conclusion, mouse ovarian tissues appear to
contain telocytes, which highly express CD34, PDGFRα/β and
vimentin. The numbers of these cells were significantly reduced
following cyclophosphamide treatment. Therefore, telocytes may
serve as a potential novel marker of POF induced by
cyclophosphamide. Future studies are required to investigate the
mechanism underlying the effect of POF on telocytes.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81273794 and
81202811), the China Postdoctoral Science Foundation (grant nos.
2014M550250 and 2015T80455) and the Shanghai Natural Science
Foundation (grant no. 16ZR1434000).
References
|
1
|
Beck-Peccoz P and Persani L: Premature
ovarian failure. Orphanet J Rare Dis. 1:92006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Vujović S, Ivović M, Tancić-Gajić M,
Marina L, Barać M, Arizanović Z, Nenezić A, Ivanisević M, Micić J,
Sajić S and Micić D: Premature ovarian failure. Srp Arh Celok Lek.
140:806–811. 2012. View Article : Google Scholar
|
|
3
|
Liu T, Huang Y, Zhang J, Qin W, Chi H,
Chen J, Yu Z and Chen C: Transplantation of human menstrual blood
stem cells to treat premature ovarian failure in mouse model. Stem
Cells Dev. 23:1548–1557. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu T, Qin W, Huang Y, Zhao Y and Wang J:
Induction of estrogen-sensitive epithelial cells derived from
human-induced pluripotent stem cells to repair ovarian function in
a chemotherapy-induced mouse model of premature ovarian failure.
DNA Cell Biol. 32:685–698. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu T, Huang Y, Guo L, Cheng W and Zou G:
CD44+/CD105+ human amniotic fluid mesenchymal stem cells survive
and proliferate in the ovary long-term in a mouse model of
chemotherapy-induced premature ovarian failure. Int J Med Sci.
9:592–602. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li H, Lu S, Liu H, Ge J and Zhang H:
Scanning electron microscope evidence of telocytes in vasculature.
J Cell Mol Med. 18:1486–1489. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hatta K, Huang ML, Weisel RD and Li RK:
Culture of rat endometrial telocytes. J Cell Mol Med. 16:1392–1396.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bei Y, Zhou Q, Fu S, Lv D, Chen P, Chen Y,
Wang F and Xiao J: Cardiac telocytes and fibroblasts in primary
culture: Different morphologies and immunophenotypes. PLoS One.
10:e01159912015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zheng Y, Cretoiu D, Yan G, Cretoiu SM,
Popescu LM, Fang H and Wang X: Protein profiling of human lung
telocytes and microvascular endothelial cells using iTRAQ
quantitative proteomics. J Cell Mol Med. 18:1035–1059. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li L, Lin M, Wang R, Zhang C, Qi G, Xu M,
Rong R and Zhu T: Renal telocytes contribute to the repair of
ischemically injured renal tubules. J Cell Mol Med. 18:1144–1156.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cretoiu SM and Popescu LM: Telocytes
revisited. Biomol Concepts. 5:353–369. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Popescu LM, Curici A, Wang E, Zhang H, Hu
S and Gherghiceanu M: Telocytes and putative stem cells in ageing
human heart. J Cell Mol Med. 19:31–45. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang XJ, Yang J, Liu Z, Yang G and Shen
ZJ: Telocytes damage in endometriosis-affected rat oviduct and
potential impact on fertility. J Cell Mol Med. 19:452–462. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vannucchi MG, Traini C, Manetti M,
Ibba-Manneschi L and Faussone-Pellegrini MS: Telocytes express
PDGFRα in the human gastrointestinal tract. J Cell Mol Med.
17:1099–1108. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cismasiu VB and Popescu LM: Telocytes
transfer extracellular vesicles loaded with microRNAs to stem
cells. J Cell Mol Med. 19:351–358. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Roatesi I, Radu BM, Cretoiu D and Cretoiu
SM: Uterine telocytes: A review of current knowledge. Biol Reprod.
93:102015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Vannucchi MG, Traini C, Guasti D, Del
Popolo G and Faussone-Pellegrini MS: Telocytes subtypes in human
urinary bladder. J Cell Mol Med. 18:2000–2008. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li H, Zhang H, Yang L, Lu S and Ge J:
Telocytes in mice bone marrow: Electron microscope evidence. J Cell
Mol Med. 18:975–978. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fu S, Wang F, Cao Y, Huang Q, Xiao J, Yang
C and Popescu LM: Telocytes in human liver fibrosis. J Cell Mol
Med. 19:676–683. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang F, Song Y, Bei Y, Zhao Y, Xiao J and
Yang C: Telocytes in liver regeneration: Possible roles. J Cell Mol
Med. 18:1720–1726. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li J, Shen F, Guan C, Wang W, Sun X, Fu X,
Huang M, Jin J and Huang Z: Activation of Nrf2 protects against
triptolide-induced hepatotoxicity. PLoS One. 9:e1006852014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Milia AF, Ruffo M, Manetti M, Rosa I,
Conte D, Fazi M, Messerini L and Ibba-Manneschi L: Telocytes in
Crohn's disease. J Cell Mol Med. 17:1525–1536. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Du X, Wang H, Xu F, Huang Y, Liu Z and Liu
T: Enterovirus 71 induces apoptosis of SHSY5Y human neuroblastoma
cells through stimulation of endogenous microRNA let-7b expression.
Mol Med Rep. 12:953–959. 2015.PubMed/NCBI
|
|
24
|
Hartmann BW, Kirchengast S, Albrecht A,
Huber JC and Söregi G: Effect of hormone replacement therapy on
growth hormone stimulation in women with premature ovarian failure.
Fertil Steril. 68:103–107. 1997. View Article : Google Scholar : PubMed/NCBI
|