Introduction
Prostate cancer, which is characterized by the
development of prostate epithelial malignant tumors, occurs solely
in men and is associated with the highest rates of morbidity and
mortality (1). Prostate cancer
poses a major public health problem worldwide (1). The incidence of prostate cancer is
particularly prevalent in older men (2). The therapeutic strategies currently
used to treat prostate cancer include watchful waiting, surgery,
radiotherapy, chemotherapy, hormone therapy and biotherapy
(3). The androgen receptor (AR)
has an important role in prostate cancer development and
progression (3).
Androgen-deprivation therapy (ADT) is an important means of
treatment for patients with prostate cancer; however, one
disadvantage is that the prostate cancer may develop resistance to
ADT over time (4). At present,
there is a dearth of effective treatment methods which are
beneficial to those patients who have developed androgen resistance
in prostate cancer. Therefore, prostate cancer therapy remains
unsatisfactory, and there is an urgent requirement to identify
novel therapeutic strategies to overcome resistance to androgens in
patients with prostate cancer.
Lysine specific demethylase 1 (LSD1) is a histone
demethylase, which exerts important roles in tumorigenesis
(5–8). LSD1 has been reported to be highly
expressed in various cancer cell types, particularly in prostate
cancer (9). Previous studies have
demonstrated that LSD1, as an AR-interacting protein, may promote
AR-dependent gene expression, which subsequently leads to the
constitutive maintenance of cancer cells via growth signals and an
enhanced risk of tumor relapse (9,10).
In addition, it has been suggested that histone modification
patterns may be used to predict the risk of prostate cancer
recurrence (11). Although LSD1
regulates the expression of a wide range of genes and is involved
in the processes of prostate cancer progression and deterioration
(9), the underlying molecular
mechanisms remain to be fully elucidated. Therefore, the inhibition
of LSD1 activity may provide a useful target for the treatment of
prostate cancer.
Cisplatin, also known as
cis-diamminedichloroplatinum or DDP, is a platinum-based
drug commonly used in the clinic as a chemotherapeutic agent. It
has numerous characteristic properties, including broad-spectrum
anticancer activity and curative effects, which render it useful
for the clinical treatment of various tumors (12). However, its use is associated with
several side effects, which serve to limit the doses that may be
administered, predominantly due to nephrotoxicity (13). Even so, it remains in use as a
standard chemotherapeutic agent for the treatment of numerous types
of cancer, including ovarian, cervical and prostate cancer
(14–17). A previous study demonstrated that
patients treated with DDP in combination with β-elemene were able
to better tolerate the chemotherapy, which afforded an improved
treatment for hormone-refractory prostate cancer (18). Therefore, how to reduce the
toxicity associated with DDP treatment is a keenly studied topic in
cancer research.
The present study aimed to provide important
insights into the effects of LSD1 knockdown and its interplay with
DDP on the proliferation, apoptosis and invasion of PC3 human
prostate cancer cells. In addition, the present study revealed
whether LSD1 knockdown could increase the sensitivity of DDP for
the treatment of prostate cancer. The results may provide important
implications for the development of novel therapeutic
strategies.
Materials and methods
Cell line and culture
The PC3 human prostate cancer cell line was
purchased from the American Type Culture Collection (Manassas, VA,
USA). The cells were grown in Gibco™ RPMI-1640 medium (Thermo
Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10%
fetal bovine serum (FBS) and 2 mM glutamine (both Sigma-Aldrich,
St. Louis, MO, USA) at 37°C in a humidified atmosphere containing
5% CO2. Plasmids encoding LSD1 small interfering (si)RNA
or mock vehicle pCMV-G&NR-U6-shRNA (GeneChem Co., Ltd.,
Shanghai, China) were transfected into the PC3 cells in 6-well
plates using a lentiviral vector (JRDUN Biotechnology, Shanghai,
China) and Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocols. Three
sequences of the LSD1 were used, as follows: Short hairpin
(sh)LSD1-1, 5′-ACGAAAGTGTCTCCGTTGA-3′; shLSD1-2,
5′-CCGACATGGCTTTCTCTTT-3′; and shLSD1-3, 5′-TCGACAGTGACCCCTTATA-3′.
The cells were split twice weekly, and cells in the logarithmic
growth phase were used for subsequent experiments.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.), and the mRNA was reverse transcribed into cDNA using the
TIANScript RT kit (Tiangen Biotech. Co. Ltd., Shanghai, China).
Subsequently, qPCR was conducted using the SYBR Green PCR Master
mix (Thermo Fisher Scientific, Inc.) and the ABI 7300 Real-Time PCR
system (Applied Biosystems; Thermo Fisher Scientific, Inc.), in
which glyceraldehyde-3-phosphate dehydrogenase (GADPH) was used as
the reference gene. The following primers were used: LSD1, forward
(F) 5′-AAGCAGGAGGACTTCAAGAC-3′, reverse (R)
5′-GCAGTGTGCGGTTTCTAATG-3′; GAPDH, F 5′-CACCCACTCCTCCACCTTTG-3′ and
R 5′-CCACCACCCTGTTGCTGTAG-3′ (Generay Biotech Co., Ltd., Shanghai,
China). The PCR cycling conditions were as follows: 95°C for 10
min, followed by 40 cycles at 95°C for 15 sec and 60°C for 1 min.
RT-qPCR data were analyzed with SDS 2.3 software. Each experiment
was repeated three times and the relative mRNA expression levels of
LSD1 were calculated using the 2−ΔΔCq method (19).
Cell Counting kit 8 (CCK-8) assay
The viability of the PC3 cells was measured using a
CCK-8 kit (Boster Biological Technology, Ltd., Wuhan, China). The
cells were seeded into a 96-well microplate at a density of
5×103/well and incubated for 24 h. The peripheral wells
of the microplate contained phosphate-buffered saline (PBS) only.
The cells were divided into four groups: Mock vehicle group, LSD1
siRNA group, mock vehicle + DDP group or LSD1 siRNA + DDP group,
according to the experimental design. The cells were treated with 5
µg/ml DDP (Beyotime Institute of Biotechnology, Haimen,
China), after which the cells were incubated for a further 24 h
prior to the assay. A total of 10 µl CCK8 solution was added
to each well containing PC3 cells, and the cells were incubated for
0, 24, 48 or 72 h. Finally, the absorbance of each well was
measured at 490 nm using a Gemini XPS microplate reader (Molecular
Devices, LLC, Sunnyvale, CA, USA).
Flow cytometric analysis
The PC3 cells (5×105 cells/ml) were
inoculated into 6-well plates. Each group comprised three double
wells on the plate. Following a 24 h incubation, the groups were
generated by addition of the appropriate reagents to the cells, and
the cells were incubated for a further 24 h. The flow cytometric
analysis was performed according to the protocol of the Annexin
V-Fluorescein Isothiocyanate Apoptosis Detection kit (Abcam,
Cambridge, UK). The apoptotic rates were analyzed immediately using
a FACSCalibur™ flow cytometer (BD Biosciences, San Jose, CA,
USA).
Cell invasive capability measured using a
Transwell assay
Following the removal of the culture medium from
each group, the cells were digested with trypsin and diluted to
1×105/ml in serum-free Dulbecco's Modified Eagle's
Medium containing 1% FBS (GE Healthcare Life Sciences, Logan, UT,
USA). A total of 800 µl culture medium containing 10% FBS
was added to the coated lower chambers of a Transwell system.
Matrigel (BD Biosciences) was coated onto the upper chambers of the
Transwell system, after which 200 µl cell suspension was
added to the upper chambers at a density of 5×104/well.
The plate was cultured for 24 h, after which the cells on the upper
layer were removed. The cells that had migrated to the lower layer
were washed with PBS, fixed with methanol and stained with 1%
crystal violet. The invasive cells were counted in five fields for
each sample under an inverted microscope (BX51; Olympus
Corporation, Tokyo, Japan), and the results were averaged. The
experiments were repeated three times.
Western blot analysis
Total protein was extracted from the cell samples
using radioimmunoprecipitation assay lysis buffer (Beyotime
Institute of Biotechnology), and was quantified using the
Bicinchoninic Acid Protein Assay kit (Thermo Fisher Scientific,
Inc.). Subsequently, equal volumes of protein (30 µg) were
separated by 12% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis followed by immunoblotting onto a nitrocellulose
membrane (EMD Millipore, Billerica, MA, USA) using an
electrophoretic transfer cell (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The membranes were blocked with 5% skimmed
milk, followed by incubation overnight at 4°C with the following
primary antibodies: Monoclonal anti-E-cadherin (#14472; 1:1,000),
polyclonal anti-cyclo-oxygenase-2 (COX-2; #4842; 1:1,000),
monoclonal anti-Smad2/3 (#8685; 1:1,000), monoclonal
anti-phosphorylated (p)-Smad2/3 (#8828; 1:1,000), monoclonal
anti-cyclin D (#2978S; 1:1,000), polyclonal anti-p-Akt (#9271;
1:1,000), polyclonal anti-Akt (#9272; 1:1,000), monoclonal
anti-extracellular signal-regulated kinase (ERK; #4695; 1:1,000),
monoclonal anti-p-ERK (#4376; 1:1,000) and monoclonal anti-GAPDH
(#5174; 1:1,500) from Cell Signaling Technology, Inc. (Danvers, MA,
USA), and polyclonal anti-transforming growth factor-β1 (TGF-β1;
ab92486; 1:800) and anti-vascular endothelial growth factor (VEGF;
ab46154; 1:1,000) from Abcam. Subsequently, the blots were washed
three times with PBS and incubated with horseradish
peroxidase-conjugated goat anti-mouse (A0216; 1:1,000) or goat
anti-rabbit (A0208; 1:1,000; both Beyotime Institute of
Biotechnology) secondary antibodies for 1 h at room temperature.
The bands were detected by reaction with enhanced chemiluminescence
detection system reagents (EMD Millipore) and exposure to X-ray
film (Kodak, Rochester, NY, USA), which was subsequently developed
and used to capture photographic images. GADPH was used to
normalize the protein expression. Band intensities were analyzed
using ImageJ 1.49 software (https://imagej.nih.gov/ij/).
Statistical analysis
All data are presented as the mean ± standard
deviation. The data were evaluated using the Prism 5.0 statistical
software package (GraphPad Software Inc., San Diego, CA, USA). The
two-tailed Student's t-test was used to evaluate statistical
differences between two groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
Proliferation of PC3 cells is decreased
following LSD1 knock-down and treatment with DDP
The present study hypothesized that LSD1 may have an
important role in PC3 cells, which has not been previously
investigated. Lentiviral-mediated RNA interference technology was
used to establish a stably transfected LSD1 knockdown PC3 cell
line, and the CCK-8 colorimetric assay was subsequently used to
determine cell proliferation. As shown in Fig. 1A, the mRNA expression levels of
LSD1 in the knockdown group were decreased to <60% of the
control levels (P<0.01), whereas the mRNA expression levels of
LSD1 in the mock group (i.e. control cells which were transfected
with an irrelevant interference sequence) exhibited no significant
changes. All of the groups contained similar cell numbers at the 0
h time point; however, proliferation of the LSD1 knockdown PC3
cells was significantly decreased after 24, 48 and 72 h, as
compared with the mock group (P<0.01; Fig. 1B). Furthermore, compared with the
mock group, DDP (at a concentration of 5 µg/ml) exerted a
marked inhibitory effect on PC3 cell proliferation. Notably, the
LSD1 knockdown + DDP group demonstrated a more marked inhibition on
PC3 cell proliferation, as compared with the DDP or LSD1 knockdown
groups (P<0.05). These findings indicate that the proliferative
capability of the PC3 cells was decreased following LSD1 knockdown,
thus suggesting that LSD1 may contribute to the proliferation of
PC3 cells, and that siRNA interference may result in reduced cell
growth. Furthermore, DDP had similar effects to LSD1 siRNA, and a
combination of LSD1 knockdown and DDP treatment produced a
synergistic effect.
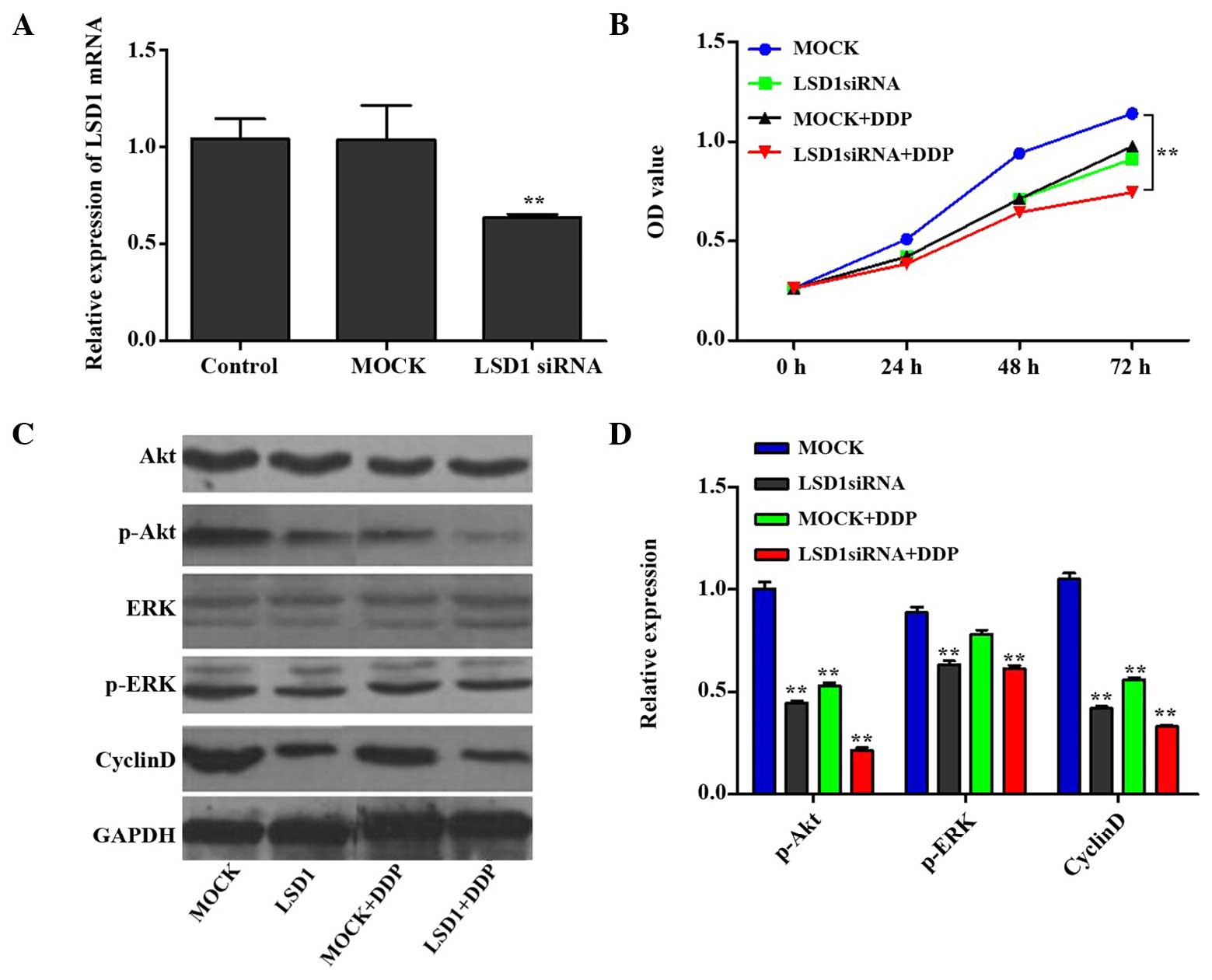 | Figure 1LSD1 RNA interference and cell growth
rate. (A) Compared with the control group, significantly decreased
mRNA expression levels of LSD1 were detected following LSD1
knockdown; however, no significant changes were observed in the
MOCK group (control cells transfected with an irrelevant
interference sequence). (B) Cell growth in each group was measured
using the Cell Counting kit-8 assay. At 0 h, all groups exhibited
very similar cell numbers, whereas at 24, 48 and 72 h, the growth
rate of the PC3 cells was significantly decreased following LSD1
knockdown, as compared with the MOCK group. Similar results were
observed in the DDP group. Notably, the combined action of LSD1
knockdown and DDP inhibited PC3 cell proliferation more markedly,
as compared with either considered in isolation. (C and D) Protein
expression levels of Akt, ERK and cyclin D are shown. Data are
presented as the mean ± standard deviation. **P<0.01
vs. the control or MOCK group. LSD1, lysine specific demethylase 1;
siRNA, small interfering RNA; DDP,
cis-diamminedichloroplatinum/cisplatin; OD, optical density;
GAPDH, glyceraldehyde-3-phosphate dehydrogenase; ERK, extracellular
signal-regulated kinase. |
LSD1 knockdown and DDP treatment inhibit
PC3 cell proliferation via regulation of the Akt and ERK signal
transduction pathways
The results of the present study revealed that cell
proliferation was inhibited when LSD1 was knocked down and the
cells were co-treated with DDP, as compared with the mock group.
Since the Akt and ERK signal transduction pathways are reported to
modulate cell proliferation and tumorigenicity (20,21),
the protein expression levels of p-Akt and p-ERK were subsequently
investigated. Furthermore, cyclin D has a crucial role in the Akt
signal pathway as a regulatory protein of the cell cycle (22), and therefore the protein expression
levels of cyclin D were also investigated. The protein expression
levels of p-Akt, p-ERK and cyclin D1 were downregulated in the
treatment groups (LSD1, mock + DDP, LSD1 + DDP), as compared with
the mock group (Fig. 1C and D).
These results suggest that LSD1 RNA interference and treatment with
DDP may inhibit PC3 cell proliferation via regulation of the Akt
and ERK signal pathways.
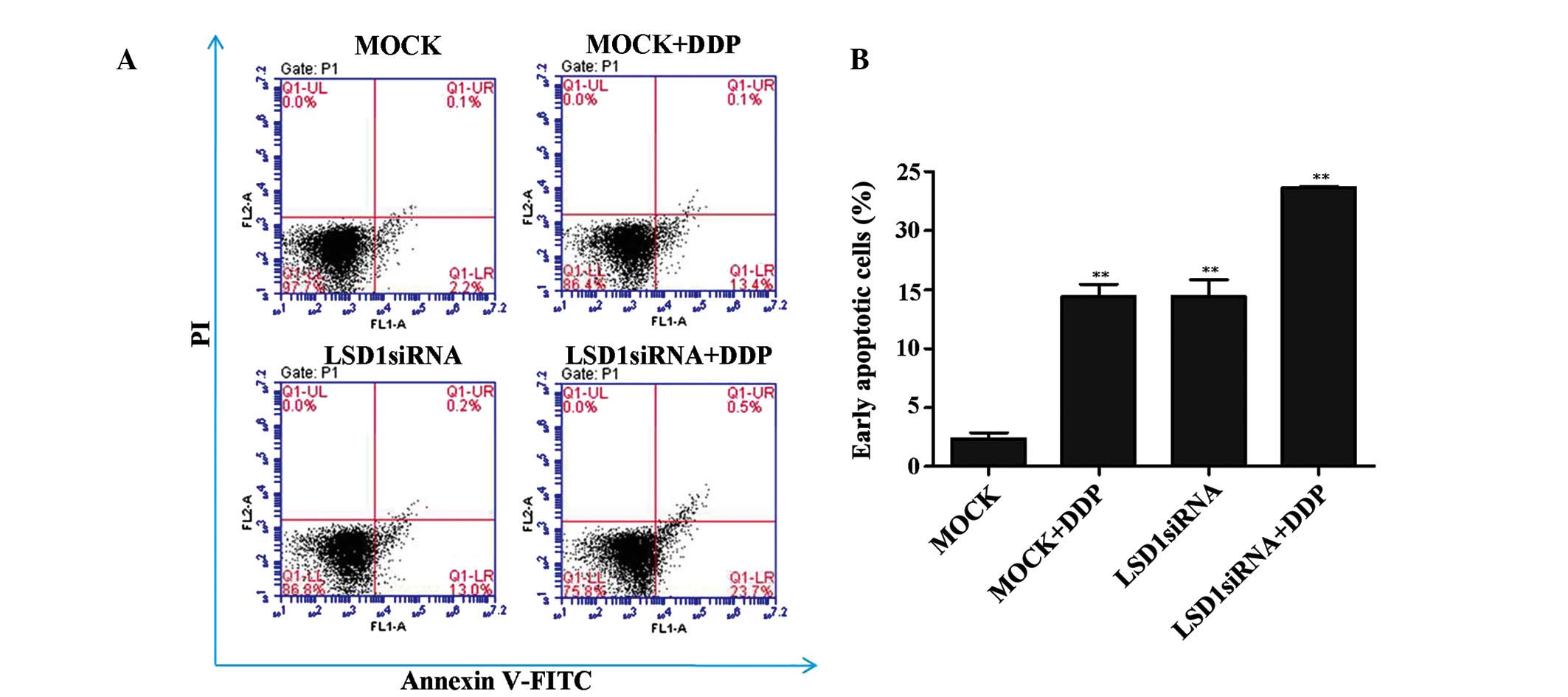
Percentage of PC3 cells in the early
apoptotic phase is increased following LSD1 knockdown and treatment
with DDP
In addition to regulating PC3 cell growth, LSD1 may
be involved in regulating cell apoptosis. As shown in Fig. 2A and B the proportion of PC3 cells
in the early apoptotic phase was significantly increased following
LSD1 knockdown, as compared with the mock group. Furthermore, it
was revealed that DDP contributes to the increased levels of
apoptosis in the PC3 cells, and that treatment with DDP exerted a
synergistic action on PC3 cell apoptosis when combined with LSD1
siRNA. These results suggest that a combination of LSD1 knockdown
and DDP treatment may contribute to the increased percentage of PC3
cells in the early apoptotic stage.
Invasive capability of PC3 cells is
decreased following LSD1 knockdown and treatment with DDP via
regulation of the TGF-β1/Smad2/3 signal transduction pathway
Invasive capability is essential for the malignant
progression of tumors. As shown in Fig. 3A and B, the invasive capability of
the PC3 cells was decreased following LSD1 knockdown or treatment
with DDP, as compared with the mock group. Furthermore, a
combination of LSD1 knockdown and DDP treatment exerted a
synergistic effect on the decline in invasive ability. These
results suggest that LSD1 may have a crucial role in the invasion
of PC3 cells.
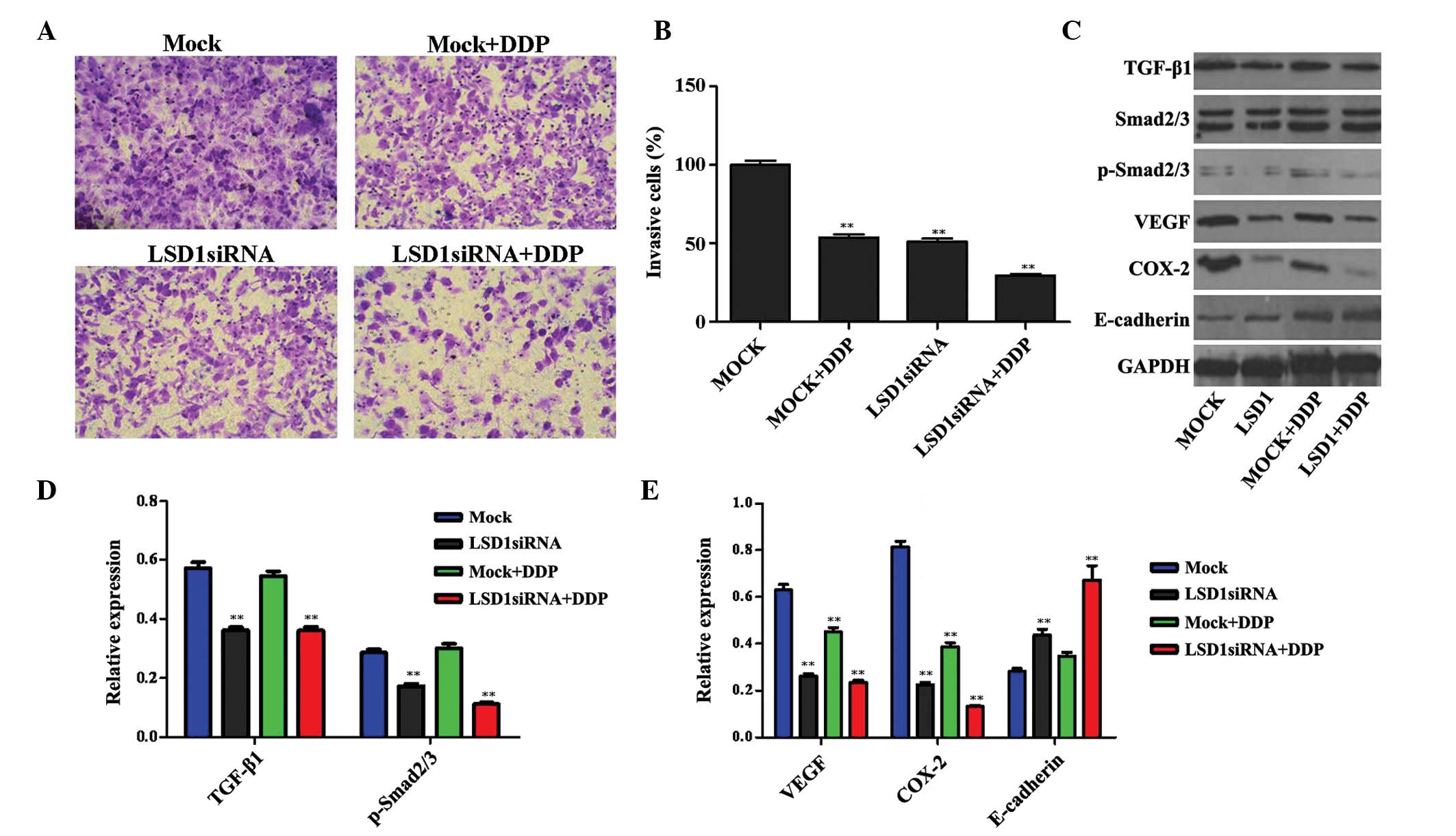 | Figure 3Changes in the invasive capability of
PC3 cells following LSD1 RNA interference were detected using the
Transwell method. (A and B) Images are shown detailing cellular
invasion of the various cell groups (magnification, ×200). Compared
with the MOCK group, the invasive capability of the PC3 cells was
decreased following LSD1 knockdown or DDP treatment. Furthermore,
LSD1 knockdown and treatment with DDP exerted a synergistic effect
on the decrease in invasive capability. (C) Expression levels of
proteins associated with the TGF-β1/Smad2/3 signaling pathway were
detected. (D and E) Quantification of the western blotting data.
Data are presented as the mean ±standard deviation.
**P<0.01, compared with the MOCK group. LSD1, lysine
specific demethylase 1; siRNA, small interfering RNA; DDP,
cis-diamminedichloroplatinum/cisplatin; VEGF, vascular endothelial
growth factor; COX-2, cyclooxygenase-2; TGF-β1, transforming growth
factor-β1; p-, phosphorylated; GAPDH, glyceraldehyde 3-phosphate
dehydrogenase. |
TGF-β1 is a cytokine peptide, which is associated
with various biological roles in cancer (23). The Smad proteins act as substrates
of the TGF-β1 receptor, and their activation by phosphorylation
propagates the TGF-β1 signal transduction. In particular, the
TGF-β1/Smad2/3 signaling pathway is involved in mechanisms
underlying the invasion and metastasis of prostate cancer cells, a
process which is also regulated by tumor angiogenesis, the host
immune system, the cells themselves and the surrounding matrix
microenvironment (24–26). Consequently, the protein expression
levels of TGF-β1, p-Smad2/3, VEGF, COX-2 and E-cadherin, all of
which are associated with the TGF-β1/Smad2/3 signal transduction
pathway, were investigated. The results revealed that the
expression levels of TGF-β1, p-Smad2/3, VEGF and COX-2 in the
DDP-treated cells were downregulated, whereas the protein
expression levels of E-cadherin were upregulated. Furthermore, the
combination of LSD1 knockdown and treatment with DDP exerted a
synergistic effect, as compared with either treatment taken in
isolation or with the mock cells (Fig.
3C–E).
Discussion
Prostate cancer is a complex disease, and numerous
controversies are associated with aspects of different treatment
strategies (27). The
identification of genetic and molecular events that may improve the
early detection of prostate cancer, or that could be used as
therapeutic targets, is a top priority in this line of study.
LSD1 is a flavin-dependent amine oxidase, which has
been reported to interact with the AR (9). Previous studies have reported that
LSD1 is overexpressed in prostate cancer (9,28).
DDP is an inorganic compound that is widely used in cancer therapy
(12). In the present study, the
antiproliferative, proapoptotic and anti-invasive effects of LSD1
knockdown and DDP treatment, either in isolation or in combination,
on PC3 prostate cancer cells were investigated. The results of the
present study may lead to an improved understanding regarding how
LSD1 knockdown and DDP treatment affect the physiological activity
of prostate cancer cells, thereby providing a novel target for
therapeutic intervention in prostate cancer.
A combination of LSD1 knockdown and DDP treatment
effectively suppressed the proliferation of PC3 cells. The Akt and
ERK signaling pathways have been reported to be important pathways
closely associated with cell proliferation (29,30).
In the present study, western blotting results suggested that the
protein expression levels of the associated proteins, p-Akt, cyclin
D1 and p-ERK, were markedly decreased in the LSD1 siRNA, DDP
treatment and LSD1 siRNA + DDP cell groups, as compared with the
mock group. These findings indicated that LSD1 knockdown and DDP
treatment may effectively inhibit the proliferation of PC3 cells
via regulation of the Akt and ERK signaling pathways. Flow
cytometry was also performed to determine the extent of apoptosis
in the prostate cancer cells. All of the treatments, i.e. LSD1
knockdown and treatment with DDP, either alone or in combination,
resulted in an increased induction of PC3 cell apoptosis.
Tumor cell invasion and metastasis are complicated
by the existence, and interplay, of several mechanisms (31). The effects of LSD1 knockdown on the
invasive capability of the PC3 prostate cancer cells were examined
in the present study. The results demonstrated that knockdown of
LSD1, in combination with DDP, suppressed the invasion of prostate
cancer cells. TGF-β is a cytokine peptide, which exerts various
biological activities. A previous study demonstrated that TGF-β1
was able to promote the invasion and metastasis of prostate tumor
cells (32). Smad proteins are the
sole substrates of TGF-β1 identified in the TGF-β1 signal
transduction pathway (33). In the
present study, the protein expression levels of TGF-β1 and
p-Smad2/3 were decreased, which implied that this signaling pathway
may be primarily responsible for the process by which LSD1
knockdown and DDP treatment inhibited the invasion of PC3 cells.
TGF-β1/Smad2/3 signaling is known to promote specific mechanisms
underlying the processes of cell invasion and metastasis in
prostate cancer, via regulation of angiogenesis, the immune defense
system, changes in the substrate microenvironment or alterations to
the cells themselves (34–36).
The formation of blood vessels is crucial in tumor
growth, invasion and metastasis (37). The present study revealed that
VEGF, as one of the most important promoters of angiogenesis
(38), was downregulated in the
LSD1 siRNA group. In addition, epithelial-mesenchymal transition is
considered to be a key step in the process of tumor metastasis. For
example, E-cadherin is as a Ca2+-dependent glycoprotein,
which performs an essential role in cell invasion and migration
(39,40). The results of the present study
implied that LSD1 knockdown and DDP treatment inhibited the
invasion of the PC3 cells by upregulating the expression levels of
E-cadherin. COX-2, which is a rate-limiting enzyme in the
prostaglandin biosynthesis pathway, is able to promote tumor
angiogenesis (41). The western
blotting results of the present study demonstrated that the protein
expression levels of COX-2 were decreased in DDP-treated LSD1
knockdown cells, with a similar result obtained for VEGF. These
results may verify the hypothesis that LSD1 knockdown inhibits the
invasion of PC3 cells by regulating the TGF-β1/Smad2/3 signal
pathway, and a combination of LSD1 siRNA and DDP treatment may lead
to more pronounced effects.
In conclusion, the present study aimed to identify
the proapoptotic, antiproliferative and anti-invasive effects of
LSD1 knockdown, in combination with DDP treatment, on PC3 human
prostate cancer cells, and to offer an explanation for the
underlying mechanism. The results of the present study may provide
novel insights into the molecular mechanism underlying the
progression and pathogenesis of prostate cancer, and may be useful
for the optimization of therapeutic interventions for the treatment
of this disease.
References
|
1
|
Center MM, Jemal A, Lortet-Tieulent J,
Ward E, Ferlay J, Brawley O and Bray F: International variation in
prostate cancer incidence and mortality rates. Eur Urol.
61:1079–1092. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bechis SK, Carroll PR and Cooperberg MR:
Impact of age at diagnosis on prostate cancer treatment and
survival. J Clin Oncol. 29:235–241. 2011. View Article : Google Scholar :
|
|
3
|
Chen CD, Welsbie DS, Tran C, Baek SH, Chen
R, Vessella R, Rosenfeld MG and Sawyers CL: Molecular determinants
of resistance to antiandrogen therapy. Nat Med. 10:33–39. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shahinian VB, Kuo YF, Freeman JL and
Goodwin JS: Risk of fracture after androgen deprivation for
prostate cancer. N Engl J Med. 352:154–164. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lv T, Yuan D, Miao X, Lv Y, Zhan P, Shen X
and Song Y: Overexpression of LSD1 promotes proliferation,
migration and invasion in non-small cell lung cancer. PloS One.
7:e350652012. View Article : Google Scholar
|
|
6
|
Zhao ZK, Dong P, Gu J, Chen L, Zhuang M,
Lu WJ, Wang DR and Liu YB: Overexpression of LSD1 in hepatocellular
carcinoma: A latent target for the diagnosis and therapy of
hepatoma. Tumor Biol. 34:173–180. 2013. View Article : Google Scholar
|
|
7
|
Qin Y, Zhu W, Xu W, Zhang B, Shi S, Ji S,
Liu J, Long J, Liu C, Liu L, et al: LSD1 sustains pancreatic cancer
growth via maintaining HIF1α-dependent glycolytic process. Cancer
Lett. 347:225–232. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kauffman EC, Robinson BD, Downes MJ,
Powell LG, Lee MM, Scherr DS, Gudas LJ and Mongan NP: Role of
androgen receptor and associated lysine-demethylase coregulators,
LSD1 and JMJD2A, in localized and advanced human bladder cancer.
Mol Carcinog. 50:931–944. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Metzger E, Wissmann M, Yin N, Müller JM,
Schneider R, Peters AH, Günther T, Buettner R and Schüle R: LSD1
demethylates repressive histone marks to promote
androgen-receptor-dependent transcription. Nature. 437:436–439.
2005.PubMed/NCBI
|
|
10
|
Kahl P, Gullotti L, Heukamp LC, Wolf S,
Friedrichs N, Vorreuther R, Solleder G, Bastian PJ, Ellinger J,
Metzger E, et al: Androgen receptor coactivators lysine-specific
histone demethylase 1 and four and a half LIM domain protein 2
predict risk of prostate cancer recurrence. Cancer Res.
66:11341–11347. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Seligson DB, Horvath S, Shi T, Yu H, Tze
S, Grunstein M and Kurdistani SK: Global histone modification
patterns predict risk of prostate cancer recurrence. Nature.
435:1262–1266. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Natile G and Coluccia M: Current status of
trans-platinum compounds in cancer therapy. Coord Chem Rev.
216:383–410. 2001. View Article : Google Scholar
|
|
13
|
Miller RP, Tadagavadi RK, Ramesh G and
Reeves WB: Mechanisms of Cisplatin nephrotoxicity. Toxins (Basel).
2:2490–2518. 2010. View Article : Google Scholar
|
|
14
|
Florea AM and Büsselberg D: Cisplatin as
an anti-tumor drug: Cellular mechanisms of activity, drug
resistance and induced side effects. Cancers (Basel). 3:1351–1371.
2011. View Article : Google Scholar
|
|
15
|
Gumulec J, Balvan J, Sztalmachova M,
Raudenska M, Dvorakova V, Knopfova L, Polanska H, Hudcova K,
Ruttkay-Nedecky B, Babula P, et al: Cisplatin-resistant prostate
cancer model: Differences in antioxidant system, apoptosis and cell
cycle. Int J Oncol. 44:923–933. 2014.
|
|
16
|
Rose PG, Sill MW, McMeekin DS, Ahmed A,
Salani R, Yamada SD, Wolfson AH, Fusco N and Fracasso PM: A phase I
study of concurrent weekly topotecan and cisplatin chemotherapy
with whole pelvic radiation therapy in locally advanced cervical
cancer: A gynecologic oncology group study. Gynecol Oncol.
125:158–162. 2012. View Article : Google Scholar
|
|
17
|
Stordal B, Hamon M, McEneaney V, Roche S,
Gillet JP, O'Leary JJ, Gottesman M and Clynes M: Resistance to
paclitaxel in a cisplatin-resistant ovarian cancer cell line is
mediated by P-glycoprotein. PloS One. 7:e407172012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li QQ, Wang G, Reed E, Huang L and Cuff
CF: Evaluation of cisplatin in combination with β-elemene as a
regimen for prostate cancer chemotherapy. Basic Clin Pharmacol
Toxicol. 107:868–876. 2010.PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
20
|
Jin F, Irshad S, Yu W, Belakavadi M,
Chekmareva M, Ittmann MM, Abate-Shen C and Fondell JD: ERK and AKT
signaling drive MED1 overexpression in prostate cancer in
association with elevated proliferation and tumorigenicity. Mol
Cancer Res. 11:736–747. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rick FG, Schally AV, Szalontay L, Block
NL, Szepeshazi K, Nadji M, Zarandi M, Hohla F, Buchholz S and Seitz
S: Antagonists of growth hormone-releasing hormone inhibit growth
of androgen-independent prostate cancer through inactivation of ERK
and Akt kinases. Proc Natl Acad Sci USA. 109:1655–1660. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Harashima N, Inao T, Imamura R, Okano S,
Suda T and Harada M: Roles of the PI3K/Akt pathway and autophagy in
TLR3 signaling-induced apoptosis and growth arrest of human
prostate cancer cells. Cancer Immunol Immunother. 61:667–676. 2012.
View Article : Google Scholar
|
|
23
|
Leivonen SK and Kähäri VM: Transforming
growth factor-beta signaling in cancer invasion and metastasis. Int
J Cancer. 121:2119–2124. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nicholson B and Theodorescu D:
Angiogenesis and prostate cancer tumor growth. J Cell Biochem.
91:125–150. 2004. View Article : Google Scholar
|
|
25
|
Wikström P, Damber J and Bergh A: Role of
transforming growth factor-beta1 in prostate cancer. Microsc Res
Tech. 52:411–419. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Denmeade SR and Isaacs JT: A history of
prostate cancer treatment. Nat Rev Cancer. 2:389–396. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kashyap V, Ahmad S, Nilsson EM, Helczynski
L, Kenna S, Persson JL, Gudas LJ and Mongan NP: The lysine specific
demethylase-1 (LSD1/KDM1A) regulates VEGF-A expression in prostate
cancer. Mol Oncol. 7:555–566. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
De Luca A, Maiello MR, D'Alessio A,
Pergameno M and Normanno N: The RAS/RAF/MEK/ERK and the PI3K/AKT
signalling pathways: Role in cancer pathogenesis and implications
for therapeutic approaches. Expert Opin Ther Targets. 16(Suppl 2):
S17–S27. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kinkade CW, Castillo-Martin M, Puzio-Kuter
A, Yan J, Foster TH, Gao H, Sun Y, Ouyang X, Gerald WL,
Cordon-Cardo C and Abate-Shen C: Targeting AKT/mTOR and ERK MAPK
signaling inhibits hormone-refractory prostate cancer in a
preclinical mouse model. J Clin Invest. 118:3051–3064.
2008.PubMed/NCBI
|
|
31
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Danielpour D: Functions and regulation of
transforming growth factor-beta (TGF-beta) in the prostate. Eur J
Cancer. 41:846–857. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
ten Dijke P and Hill CS: New insights into
TGF-beta-Smad signalling. Trends Biochem Sci. 29:265–273. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tuxhorn JA, McAlhany SJ, Yang F, Dang TD
and Rowley DR: Inhibition of transforming growth factor-beta
activity decreases angiogenesis in a human prostate cancer-reactive
stroma xenograft model. Cancer Res. 62:6021–6025. 2002.PubMed/NCBI
|
|
35
|
Donkor MK, Sarkar A, Savage PA, Franklin
RA, Johnson LK, Jungbluth AA, Allison JP and Li MO: T cell
surveillance of oncogene-induced prostate cancer is impeded by T
cell-derived TGF-β1 cytokine. Immunity. 35:123–134. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Micalizzi DS, Farabaugh SM and Ford HL:
Epithelial-mesenchymal transition in cancer: Parallels between
normal development and tumor progression. J Mammary Gland Biol
Neoplasia. 15:117–134. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kirsch M, Schackert G and Black PM:
Angiogenesis, metastasis, and endogenous inhibition. J Neurooncol.
50:173–180. 2000. View Article : Google Scholar
|
|
38
|
Ferrara N, Gerber HP and LeCouter J: The
biology of VEGF and its receptors. Nat Med. 9:669–676. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pàmies P: E-cadherin-guided migration. Nat
Mater. 13:6642014. View
Article : Google Scholar
|
|
40
|
Canel M, Serrels A, Frame MC and Brunton
VG: E-cadherin-integrin crosstalk in cancer invasion and
metastasis. J Cell Sci. 126:393–401. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Muraki C, Ohga N, Hida Y, Nishihara H,
Kato Y, Tsuchiya K, Matsuda K, Totsuka Y, Shindoh M and Hida K:
Cyclooxygenase-2 inhibition causes antiangiogenic effects on tumor
endothelial and vascular progenitor cells. Int J Cancer. 130:59–70.
2012. View Article : Google Scholar
|