Introduction
Atherosclerosis is a chronic inflammatory disorder
in which metabolic and immune components interact to initiate and
develop arterial lesions (1).
Macrophages are the major immune cells in arterial lesions, which
release various inflammatory cytokines and chemokines, are involved
in foam cell formation and express pattern-recognition receptors,
including, toll-like receptors (TLRs, which mediate innate and
adaptive immune responses (2).
There is substantial evidence that the stimulation of TLRs
initiates and accelerates atherosclerosis (3). Of the TLRs involved in
atherosclerosis, TLR2 and TLR4 have received the most
investigation, and 11 and 13 of these receptors have been
identified in humans and mice, respectively (4). However, the pathways that link TLRs
with cytoplasmic adaptors, including toll/interleukin-1
receptor-domain-containing adaptor-inducing interferon-β (TRIF) in
the process of atherosclerosis remain to be fully elucidated.
TLR signaling is controlled by four types of
cytoplasmic adaptors, myeloid differentiation factor 88 (MyD88),
TRIF, MyD88 adaptor-like (MAL) and TRIF-related adaptor molecule
(TRAM). During TLR4 signaling, MyD88 and MAL are recruit for the
first signaling pathway, leading to early nuclear factor-κB (NF-κB)
activation and the induction of cytokine genes (5). By contrast, TRIF and TRAM are
recruited for the second signaling pathway, which activates
interferon regulatory factor (IRF)3, late NF-κB activation and
induction of the interferon gene (6). Although TLR2 signaling also uses
MyD88 and Mal in a similar manner to TLR4, the availability of TRIF
remains to be elucidated. During TLR3 and TLR4 signaling, TRIF
initiates a signaling pathway through TNF receptor associated
factor (TRAF)3, TANK-binding kinase 1 and inhibitor of NF-κB-kinase
complex, which mediates the direct phosphorylation of IRF3 and IRF7
(7).
Several studies have focused on the role of MyD88 in
atherosclerosis. Inactivation of MyD88 leads to a reduction in
atherosclerosis mediated by reduced macrophage recruitment to the
artery wall, which is associated with reduced chemokine levels
(8). Additionally, Michelsen et
al showed that apolipoprotein
E−/−MyD88−/− mice exhibit reduced aortic
atherosclerosis and reduced macrophage accumulation (9). However, the role of TRIF in
atherosclerosis remains to be fully elucidated, although a previous
study by Lundberg et al demonstrated that the deletion of
TRIF from myeloid cells was sufficient to attenuate vessel
inflammation and protect against atherosclerosis (1). However, the role of TRIF in foam cell
formation mediated by TLR2 stimulation remains to be fully
elucidated.
Considering the significant importance of foam cell
formation in TLR2 signaling, we aimed to determine whether TRIF is
induced by TLR2 stimulation and whether TRIF is involved in foam
cell formation in macrophages and if so, to determine the molecular
mechanisms involved. Taken together, these results suggested the
importance of TRIF in TLR2 mediated foam cell formation via
inflammatory mediators, including MCP-1.
Materials and methods
Materials
Pam3CSK4 and the mouse TLR1-9
agonist kit was purchased from InvivoGen (San Diego, CA, USA).
TRIzol and small interfering (si)RNA (TRIF) were purchased from
Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Animals
A total of 20 male 6-week-old C57BL6 wild-type (WT)
mice (average weight) were purchased from Central Lab Animal (South
Korea). TLR2 deficient (average weight, C57BL6 background, 6 weeks,
male) mice were kindly provided by Dr SJ Lee (Seoul National
University, South Korea). For the experiments, a total of 20 mice
were used. The mice were housed in standard conditions (temperature
at 25±2°C, relative humidity (55±5%), 12/12 h light-dark cycle, and
free access to food and water). The mice were sacrificed by
cervical dislocation. The study was conducted in accordance with
the guidelines and protocols approved by the Institutional Animal
Care and Use Committee of Yeungnam University College of Medicine
(Daegu, South Korea, permit number: YUMC-AEC2011-007).
Cell culture
Raw 264.7 cell lines were purchased from the
American Type Culture Collection (Manassas, VA, USA) and grown in
Dulbecco's modified Eagle's medium (DMEM) supplemented with 10%
fetal bovine serum, penicillin (100 U/ml) and streptomycin (100
μg/ml; GE Healthcare Life Sciences, Logan, UT, USA) at 37°C
in a humidified atmosphere of 5% CO2 and 95%
O2. After euthanasia, the mice were sprayed with 70%
ethanol and the femurs were dissected using scissors, cutting
through the tibia below the knee joints, and through the pelvic
bone close to the hip joint. Muscles connected to the bone were
removed using clean gauze, and the femurs were placed into a
polypropylene tube containing sterile phosphate-buffered saline
(PBS) on ice. In a tissue culture hood, the bones were placed in
70% ethanol for 1 min, washed in sterile DMEM and then both
epiphyses were removed using sterile scissors and forceps. The
bones were flushed with a syringe filled with DMEM to extrude bone
marrow into a 15 ml sterile polypropylene tube. A 5 ml plastic
pipette was used to gently homogenize the bone marrow. Primary bone
marrow-derived monocytes were differentiated into bone
marrow-derived macrophages (BMDMs) by incubation in DMEM
supplemented with 10% L929 cell (ATCC, Manassas, VA,
USA)-conditioned medium, as a source of macrophage
colony-stimulating factor, for 5–7 days at 37°C in a humidified
atmosphere of 5% CO2 and 95% O2. The
macrophages were cultured in 35 mm diameter plates (or 6-well
plates) (5×105/1.5ml medium) and then treated with
Pam3CSK4 (100 ng/ml) for 2–6 h.
Electroporation
The Raw 264.7 cells were cultured in 100 mm diameter
dishes at 1×106 and grown overnight prior to
electroporation using Amaxa Cel Line Nuclofector Kit V (VCA-1003,
Lonza, Switzerland). Cell were harvested and a defined number of
cells washed with PBS (2×106−4×106 cells per
electroporation). The cells were pelleted by centrifugation (100 ×
g for 2 min) and resuspended in 100 μl of the
electroporation solution provided with the kit. A total of 8
μl of the siRNA (150 pM) was added to the cell suspension
and gently mixed. The cell-siRNA mix was transferred into the Amaxa
electroporation cuvette, placed into the Nuclofector and
electroporated as described by the manufacturer's instruction.
Subsequently, 500 μl warm media was added to the cuvette,
and the sample transfered to the plate and incubated at 37°C.
Reverse transcription-polymerase chain
reaction (RT-PCR) and RT-quantitative PCR (RT-qPCR)
Total RNA was extracted from the cells using TRIzol
reagent. First-strand cDNA was synthesized from 1 μg total
RNA by employing random primers, oligo-dT and reverse transcriptase
(Promega Corporation, Madison, WI, USA). The thermal cycling
condition were as follows: 95°C for 5 min, 95°C for 1 min, 63°C for
1 min, and 72°C for 1 min, for 26–33 cycles, using a Bio-Rad C1000™
Thermal Cycler (Bio-Rad Laboratories, Inc., Hercules, CA, USA). A
total of 10 μl of the final amplification product were
electrophoresed on a 2% agarose gel containing SYBR®
Safe DNA gel stain (Thermo Fisher Scientific, Inc.). The expression
levels of TLR2, TRIF, MCP-1, LOX-1 and TF were normalized against
the β-actin control and visualized using the Fuji Intelligent Dark
Box LAS-3000 Image Reader (FujiFilm, Tokyo, Japan). Densitometric
analysis was carried out using LAS-3000 Fujifilm Image Reader and
Multi Gauge 3.0 software. The following primers were used for the
RT-qPCR: TRIF, forward 5′-GTATGGGCCCTCTGACTGAT-3′ and reverse
5′-ATAGGTGTG GTCTTCCCTGC-3′; MCP-1, forward
5′-AGAGAGCCAGACGGGAGGAA-3′ and reverse 5′-GTCACACTGGTCACTCCTAC-3′;
TF, forward 5′-CACTCATCATTGTGGGAGCAGTG-3′ and reverse
5′-CGCGACGGGGTGTTCTT-3′); Lox-1, orward
5′-AGGTCCTTGTCCACAAGACTGG-3′ and reverse
5′-ACGCCCCTGGTCTTAAAGAATTG-3′); and β-actin, forward
5′-TCCTTCGTTGCCGGTCCACA-3′ and reverse
5′-CGTCTCCGGACTCCATCACA-3′.
siRNA
Stealth control siRNA and gene-specific siRNA
against the following target gene were designed using Block-IT
Stealth RNAi designer (Invitrogen; Thermo Fisher Scientific, Inc.):
TRIF, 5′-GGACAUACGUUACACUCCACCAACA-3′.
Oil-red O and hematoxylin staining for
foam cell formation
For lipid uptake analysis, the macrophages were
cultured in 6-well plates (5×105/well) and then treated
with Pam3CSK4 (100 ng/ml) and low density
lipoprotein LDL (50 μg/ml) for 24 h at 37°C in a humidified
atmosphere of 5% CO2 and 95% O2. Prior to
Oil-red O staining, the media were removed from the wells using a
Pasteur pipette or by gently inverting the plate over a waste
container, and then gently rinsing with phosphate-buffered saline.
Subsequently, 10% formalin was added to each well for 1 h to fix
the cells, following which each well was rinsed with distilled
H2O. The wells were then rinsed with 60% isopropanol for
5 min, dried and stained with Oil-red O (Sigma-Aldrich, St. Louis,
MO, USA) and hematoxylin. Intracellular lipid droplets were
detected by light microscopy using a DIAPHOT 300 light microscope
(Nikon Corporation, Tokyo, Japan). Images were captured with an
AxioCam ICc I digital camera system (Carl Zeiss, Oberkochen,
Germany).
Enzyme-linked immunosorbent assay
(EIA)
The protein levels of MCP-1 in the cell culture
supernatant were measured by EIA using a specific sandwich enzyme
immunoassay (mouse CCL2/JE/MCP-1 DuoSet ELISA kit; DY479, R&D
Systems, Inc., Minneapolis, MN, USA). Briefly, the cell culture
supernatant was placed in a 96-well microtiter plate, which was
coated with murine polyclonal antibody against mouse MCP-1.
Following incubation at room temperature for 2 h and careful
washes, HRP-conjugated polyclonal antibody against MCP-1 was added.
Following incubation for 2 h at room temperature and repeated
washes, color reagents were added. The optical density of each well
was then measured at 450 nm using an EL800 universal microplate
reader (Bio-Tek instruments, Inc., Winooski, VT, USA) using a 550
nm reference wavelength.
Statistical analysis
Results are expressed as the mean ± standard
deviation of a minimum of 3 independent assays. Statistical
significance was calculated by analysis of variance using GraphPad
Prism software, version 5.01 (GraphPad. Inc., La Jolla, CA, USA).
Groups were compared with two-way analysis of variance with a
Bonferroni post-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
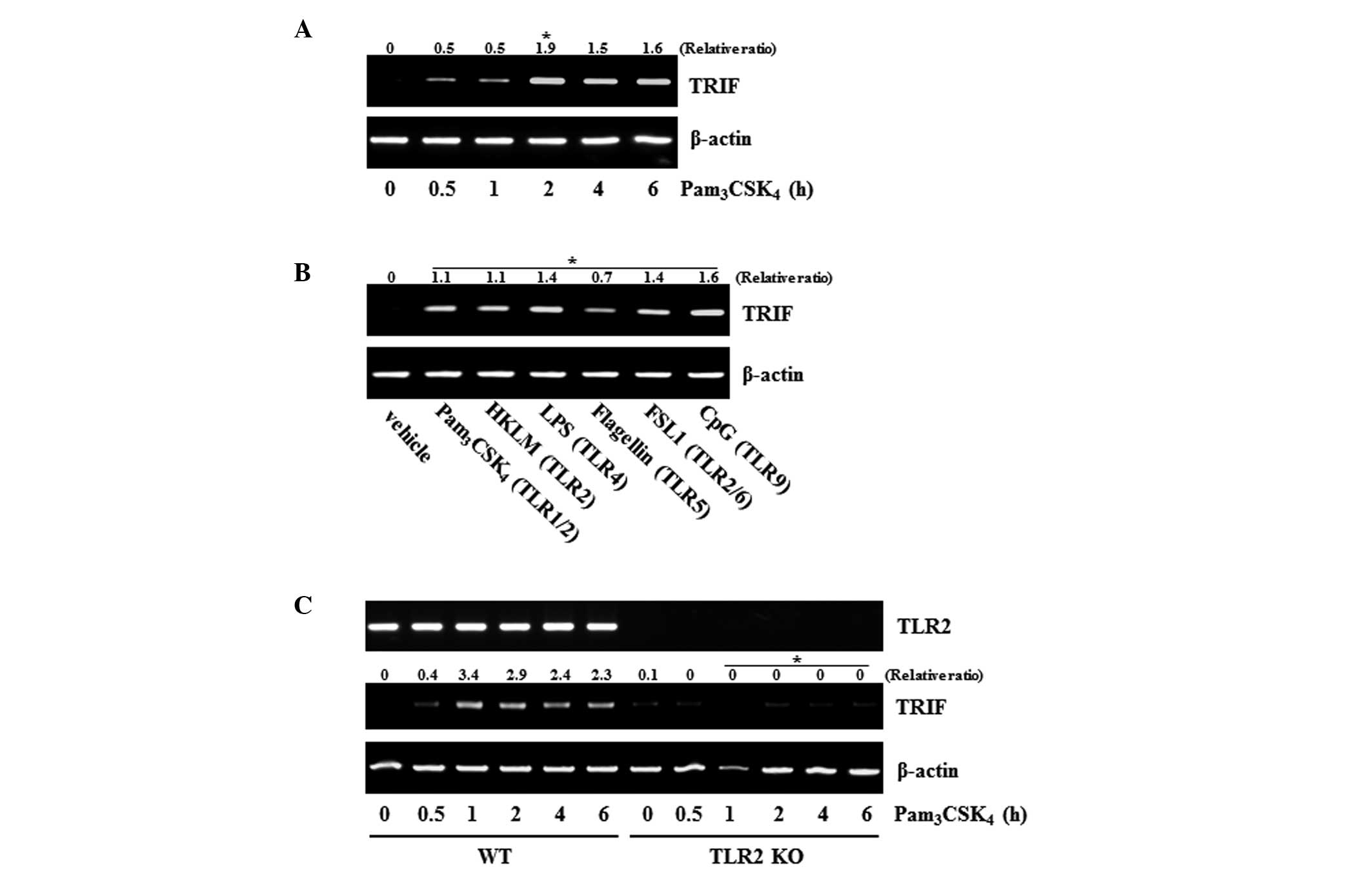
Gene expression of TRIF is induced by TLR
activation
MyD88 and TRIF are well known adaptor proteins
involved in TLR4 signaling, however, only MyD88 is involved in TLR2
signaling (10). The present study
investigated whether the TLR2 agonist,
Pam3CSK4, can induce the mRNA expression of
TRIF in the Raw 264.7 macrophage cell line. The results revealed
that Pam3CSK4 induced the mRNA expression of
TRIF in a time-dependent manner (Fig.
1A). To demonstrate the general effect of TLRs on the
expression of TRIF, other types of TLR agonist were assessed. All
of the agonists examined (TLR1/2, TLR4, TLR5, TLR6/2 and TLR9)
induced the gene expression of TRIF, with effects similar to those
of the TLR2 agonist (Fig. 1B).
These results suggested that the gene expression of TRIF by TLR
agonists is a general characteristic shown in TLRs. Whether
Pam3CSK4 affects the gene expression of TRIF
was also determined in BMDMs. The gene expression of TRIF was
induced by Pam3CSK4 in the BMDMs from the WT
mice, however, its expression was not affected in the BMDMs from
the TLR2-knockout mice (TLR2 KO; Fig.
1C).
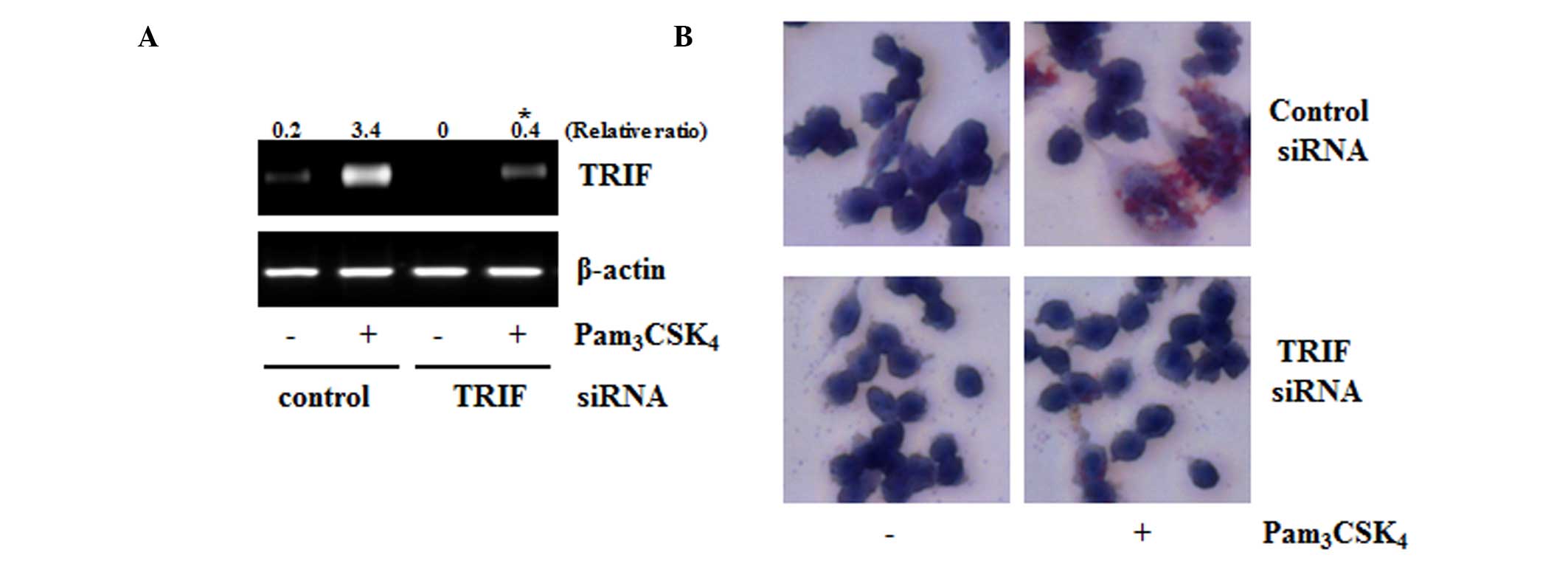
TRIF is involved in TLR2-induced foam
cell formation
Previous studies have suggested that TLRs is
involved in the pathology of atherosclerosis (11,12).
Among TLRs, TLR2 is also involved in the progression of atheroma. A
previous study reported that TLR2 is a receptor for foam cell
formation (13,14); therefore, the present study
attempted to determine the role of TRIF in TLR2-mediated foam cell
formation using the siRNA technique. Compared with the control
siRNA-transfected cells, the TRIF-siRNA transfected cells showed
decreased mRNA expression of TRIF. The TLR2-mediated foam cell
formation also decreased in the TRIF siRNA-transfected cells,
compared with the control cells (Fig.
2A and B). These results suggested that TRIF was an important
adaptor for TLR2-mediated foam cell formation.
Inflammatory mediators are upregulated by
TLR2 stimulation
TRIF is also likely to be important adaptor for
TLR2-mediated foam cell formation. To identify genes associated
with TRIF, the present study aimed to identify genes, which control
foam cell formation. Chemokines are involved in the pathogenesis of
atherosclerosis by promoting the directed migration of inflammatory
cells. MCP-1 is a representative chemokine involved in foam cell
formation (15). In the present
study, treatment of the cells with Pam3CSK4
markedly increased the mRNA expression of MCP-1 in a time-dependent
manner. TF is a major risk factor for atherosclerosis, and
lectin-Lox-1 is also crucial in oxidized-LDL-mediated
atherosclerosis (14,16). Therefore, the present study
measured changes in the expression levels of these two genes
following TLR2 stimulation. The expression levels of the two genes
increased significantly, in a time-dependent manner, although the
reaction time was marginally different for the expression of MCP-1
(Fig. 3A). The present study also
confirmed whether these genes were dependent on TLR2 using KO
mice-derived BMDMs. Whereas the expression levels of the genes were
induced by Pam3CSK4 in the BMDMs from the WT
mice, their expression levels remained unchanged in the BMDMs from
the TLR2-knockout mice (Fig.
3B).
 | Figure 3Pam3CSK4
induces inflammatory mediators via a TLR2-dependent pathway. (A)
Raw 264.7 cells were treated with Pam3CSK4
(100 ng/ml) for the indicated times. The mRNA expression levels of
MCP-1, TF and Lox-1 were determined using RT-PCR analysis and
normalized against the β-actin control. (B) BMDMs were isolated
from the bone marrow of WT or TLR2 KO mice and treated with
Pam3CSK4 (100 ng/ml) for the indicated times.
The expression gene expression levels of MCP-1, TF and Lox-1 genes
were determined using RT-PCR analysis. The results shown are
representative of experiments repeated in triplicate with similar
results. Values are presented as the mean, P<0.05 vs. the
control. TRIF, toll/interleukin-1 receptor-domain-containing
adaptor-inducing interferon-β; TLR2, toll-like receptor 2; MCP-1,
monocyte chemoattractant protein-1; TF, tissue factor; Lox-1,
lectin-like oxidized low-density lipoprotein receptor-1; RT-PCR,
transcription-quantitative polymerase chain reaction; WT,
wild-type; KO, knockout. |
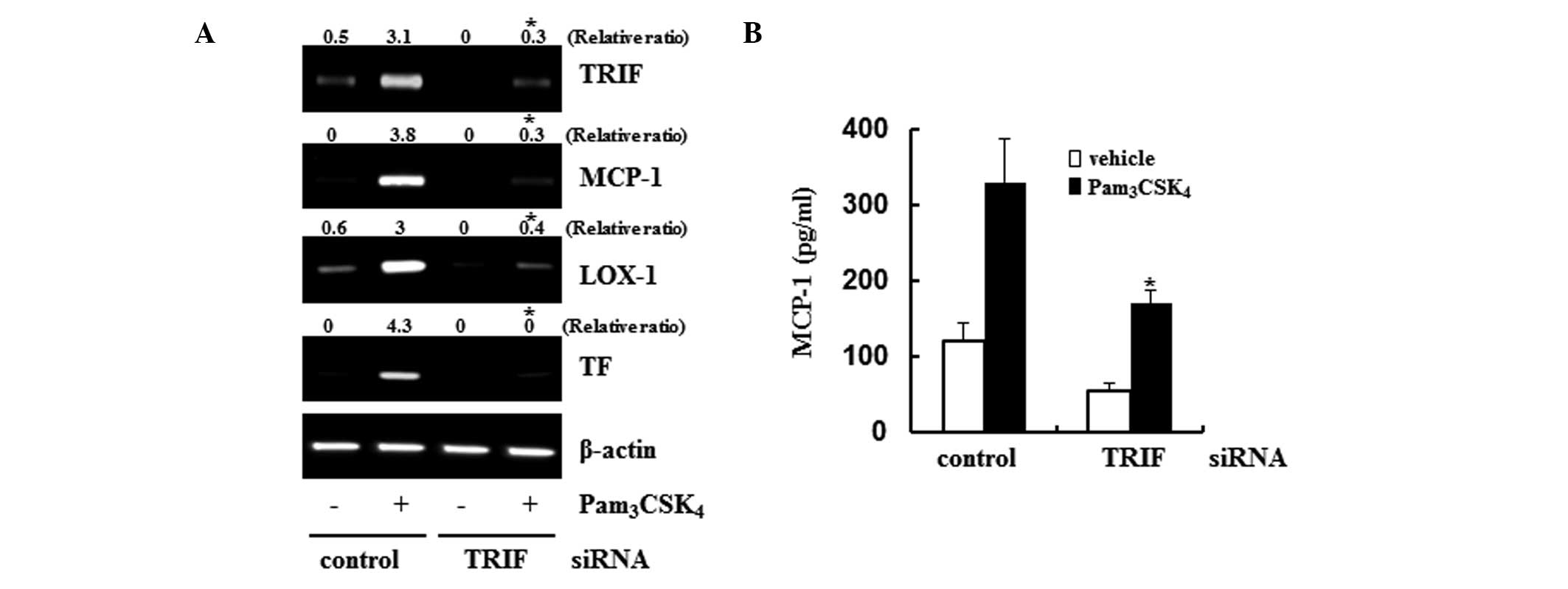
Expression levels of MCP-1, TF and Lox-1
are TRIF dependent
The present study used siRNA to confirm the role of
TRIF in the expression of MCP-1, TF and Lox-1. The TRIF siRNA
technique was found to be successful in downregulation its level of
gene expression. With the downregulation in the level of TRIF, the
gene expression levels of MCP-1, TF and Lox-1 were also markedly
attenuated on examination of the
Pam3CSK4-induced responses (Fig. 4A). To confirm the changes in MCP-1,
the present study assessed the protein levels of MCP-1 using an
EIA. Production of the MCP-1 protein increased ~3.5-fold in the
supernatants of the control siRNA-transfected cells following TLR2
stimulation. However, the protein expression of MCP-1 was reduced
to almost basal levels in the TRIF siRNA-transfected cells
(Fig. 4B). These results suggested
that TRIF was an important adaptor protein, which controlled foam
cell formation via the production of MCP-1.
Discussion
The present study initially found that
Pam3CSK4 treatment increased the gene
expression of TRIF in macrophages, suggesting that TRIF contributed
to TLR2 signaling. TLR2 has previously been shown to be regulated
via a MyD88-dependent signaling pathway only. However, the present
study found that the TRIF adaptor protein also appeared to be a
regulator of TLR2-mediated foam cell formation. In addition, as
TRIF controls the expression of MCP-1, TF and Lox-1, which are
representative regulators of foam cell formation, the present study
hypothesized that TRIF was also an important adaptor protein in
TLR2-mediated foam cell formation, together with MyD88.
TLR signaling is divided into an MyD88-dependent
pathway and an MyD88-independent signaling pathway (17). The MyD88-dependent signaling
pathway is used by all TLRs, with the exception of TLR3. Signaling
via the MyD88-dependent pathways leads to the activation of
mitogen-activated protein kinase and the inhibitor of NF-κB (IκB)
kinase complex, resulting in activation of activator protein (AP)-1
and NF-κB, respectively (18,19).
By contrast, TLR3 is only present in the MyD88-independent pathway.
TRIF is the predominant adaptor protein in the MyD88-independent
pathway, and can associate with TRAF6 to activate AP-1 and NF-κB,
as in the MyD88-dependent pathway. TRIF can also interact with
TRAF3 and phosphoinositide 3-kinase, resulting in the activation of
interferon regulatory factor (IRF)3 and IRF2, respectively
(20,21). TLR4 uses MyD88 and TRIF as adaptor
proteins (22,23).
Generally, TRIF is constantly expressed in
macrophages to convey signals via the interaction with downstream
proteins, inducing various functions through the activation of
specific transcription factors. However, the results of the present
study showed that TLR2 stimulation elevated the gene expression of
TRIF itself. To date, few mechanisms have been shown to increase
the gene expression levels of TRIF. In the present study, the gene
expression levels of TRIF were upregulated by all the TLR agonists
assessed, indicating that this may be a general observation in TLR
signaling. Due to the TRIF antibody quality, the present study was
only able to verify the increase in gene levels.
Pam3CSK4 treatment increased the expression
of TRIF, along with other inflammatory mediators, whereas the
downregulation of TRIF by siRNA decreased foam cell formation and
inflammatory mediators. Therefore, the changes in the levels of
TRIF may be an essential factor stimulating foam cell formation. In
addition, the downregulation of TRIF decreased the expression
levels of MCP-1, TF and Lox-1, which regulate foam cell formation.
These three genes are known to be important mediators involved in
atherosclerosis through foam cell formation (14–16).
MCP-1, which is also referred to as CCL2, is a small
cytokine, which belongs to the CC chemokine family. MCP-1 is one of
the key chemokines involved in the regulation of migration and
infiltration of monocytes/macrophages (24,25).
TF, which is the key initiator of the coagulation cascade, binds
factor VIIa, resulting in activation of factor IX and factor X,
ultimately leading to fibrin formation. TF is involved in the
pathogenesis of atherosclerosis by promoting thrombus formation
(26). Oxidized LDL is also
crucial in the initiation and progression of atherosclerosis
through a variety of mechanisms, including promoting foam cell
generation and activating inflammatory processes (27). Lox-1, a type II membrane protein
with a typical C-type lectin structure, has been identified as the
predominant receptor for oxidized-LDL (28). Reported data have revealed that
Lox-1 is important in atherosclerosis (16). On examination of these three
factors, which are essential in atherosclerosis, their gene
expression levels were reduced by TRIF knockdown, suggesting the
presence of a direct association between TRIF and foam cell
formation. Therefore, when the expression of TRIF is increased by
TLR2 stimulation, inflammatory mediators, including MCP-1, produce
and eventually promote the conversion of macrophages into foam
cells. Therefore, the results of the present study showed that the
reduction in expression levels of the above genes by TRIF knockdown
was directly associated with foam cell formation.
The production of these three genes regulating foam
cell formation is known to be regulated by the MyD88-dependent
pathway in TLR signaling. However, the results of the present study
showed that these three genes were regulated in a
TLR2/TRIF-dependent manner. Associated reports have shown that the
TLR4/TRIF pathway, in addition to the TLR4/MyD88 pathway, is also
an important process (29). It was
also previously reported that TRIF knockdown in differentiated
neuronal cells decreases the expression of Lox-1 (30). TRIF-dependent pathways may be
associated with TLR2 signaling, as it is with TLR-4 signaling.
However, the role of the TRIF-dependent pathway in TLR2 signaling
has received less investigation.
The results of the present study, which investigated
macrophages from mice with TRIF knockdown, provided evidence
supporting the connection between TRIF and TLR2 signaling in foam
cell formation. The results of the present study indicated that
Pam3CSK4 stimulated macrophage foam cell
formation, and induced the expression levels of MCP-1, TF and Lox-1
in the context of an immune response to TLR2 via a TRIF-dependent
pathway. In this regard, identification of TRIF as an important
regulator of TLR2 signaling in macrophages may represent a possible
therapeutic strategy for regulating inflammation and
atherosclerosis.
Abbreviations:
|
TLR
|
toll-like receptor
|
|
MyD88
|
myeloid differentiation factor 88
|
|
NF-κB
|
nuclear factor-κB
|
|
IRF
|
interferon regulatory factor
|
|
MCP-1
|
monocyte chemoattractant protein-1
|
|
TRAF
|
tumor necrosis factor
receptor-associated factor
|
|
BMDM
|
bone marrow-derived macrophage
|
|
siRNA
|
small interfering RNA
|
|
TF
|
tissue factor
|
|
Lox-1
|
lectin-like oxidized low-density
lipoprotein receptor-1
|
|
LDL
|
low density lipoprotein
|
Acknowledgments
This study was supported by the 2013 Yeungnam
University Research Grant (grant no. 213A061034).
References
|
1
|
Lundberg AM, Ketelhuth DF, Johansson ME,
Gerdes N, Liu S, Yamamoto M, Akira S and Hansson GK: Toll-like
receptor 3 and 4 signalling through the TRIF and TRAM adaptors in
haematopoietic cells promotes atherosclerosis. Cardiovasc Res.
99:364–373. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Galkina E and Ley K: Immune and
inflammatory mechanisms of atherosclerosis (*). Annu Rev Immunol.
27:165–197. 2009. View Article : Google Scholar
|
|
3
|
Erridge C: The roles of toll-like
receptors in atherosclerosis. J Innate Immun. 1:340–349. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Akira S, Uematsu S and Takeuchi O:
Pathogen recognition and innate immunity. Cell. 124:783–801. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Verstrepen L, Bekaert T, Chau TL,
Tavernier J, Chariot A and Beyaert R: TLR-4, IL-1R and TNF-R
signaling to NF-kappaB: Variations on a common theme. Cell Mol Life
Sci. 65:2964–2978. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mogensen TH: Pathogen recognition and
inflammatory signaling in innate immune defenses. Clin Microbiol
Rev. 22:240–273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ahmed S, Maratha A, Butt AQ, Shevlin E and
Miggin SM: TRIF-mediated TLR3 and TLR4 signaling is negatively
regulated by ADAM15. J Immunol. 190:2217–2228. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Falck-Hansen M, Kassiteridi C and Monaco
C: Toll-like receptors in atherosclerosis. Int J Mol Sci.
14:14008–14023. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Michelsen KS, Wong MH, Shah PK, Zhang W,
Yano J, Doherty TM, Akira S, Rajavashisth TB and Arditi M: Lack of
toll-like receptor 4 or myeloid differentiation factor 88 reduces
atherosclerosis and alters plaque phenotype in mice deficient in
apolipoprotein E. Proc Natl Acad Sci USA. 101:10679–10684. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Newton K and Dixit VM: Signaling in innate
immunity and inflammation. Cold Spring Harb Perspect Biol.
4:a0060492012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Curtiss LK and Tobias PS: Emerging role of
toll-like receptors in atherosclerosis. J Lipid Res. 50(Suppl):
S340–S345. 2009. View Article : Google Scholar :
|
|
12
|
Mann DL: The emerging role of innate
immunity in the heart and vascular system: For whom the cell tolls.
Circ Res. 108:1133–1145. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Keyel PA, Tkacheva OA, Larregina AT and
Salter RD: Coordinate stimulation of macrophages by microparticles
and TLR ligands induces foam cell formation. J Immunol.
189:4621–4629. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Park DW, Lyu JH, Kim JS, Chin H, Bae YS
and Baek SH: Role of JAK2-STAT3 in TLR2-mediated tissue factor
expression. J Cell Biochem. 114:1315–1321. 2013. View Article : Google Scholar
|
|
15
|
Park DW, Baek K, Kim JR, Lee JJ, Ryu SH,
Chin BR and Baek SH: Resveratrol inhibits foam cell formation via
NADPH oxidase 1-mediated reactive oxygen species and monocyte
chemotactic protein-1. Exp Mol Med. 41:171–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee JG, Lim EJ, Park DW, Lee SH, Kim JR
and Baek SH: A combination of Lox-1 and Nox1 regulates
TLR9-mediated foam cell formation. Cell Signal. 20:2266–2275. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mitchell D, Yong M, Schroder W, Black M,
Tirrell M and Olive C: Dual stimulation of MyD88-dependent
toll-like receptors induces synergistically enhanced production of
inflammatory cytokines in murine bone marrow-derived dendritic
cells. J Infect Dis. 202:318–329. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dauphinee SM and Karsan A:
Lipopolysaccharide signaling in endothelial cells. Lab Invest.
86:9–22. 2006. View Article : Google Scholar
|
|
19
|
Oeckinghaus A, Hayden MS and Ghosh S:
Crosstalk in NF-κB signaling pathways. Nat Immunol. 12:695–708.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Honda K and Taniguchi T: IRFs: Master
regulators of signalling by toll-like receptors and cytosolic
pattern-recognition receptors. Nat Rev Immunol. 6:644–658. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Randall RE and Goodbourn S: Interferons
and viruses: An interplay between induction, signalling, antiviral
responses and virus countermeasures. J Gen Virol. 89:1–47. 2008.
View Article : Google Scholar
|
|
22
|
Guijarro-Muñoz I, Compte M,
Álvarez-Cienfuegos A, Álvarez-Vallina L and Sanz L:
Lipopolysaccharide activates toll-like receptor 4 (TLR4)-mediated
NF-κB signaling pathway and proinflammatory response in human
pericytes. J Biol Chem. 289:2457–2468. 2014. View Article : Google Scholar
|
|
23
|
Weighardt H, Jusek G, Mages J, Lang R,
Hoebe K, Beutler B and Holzmann B: Identification of a TLR4- and
TRIF-dependent activation program of dendritic cells. Eur J
Immunol. 34:558–564. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Deshmane SL, Kremlev S, Amini S and Sawaya
BE: Monocyte chemoattractant protein-1 (MCP-1): An overview. J
Interferon Cytokine Res. 29:313–326. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kundu S, Roome T, Bhattacharjee A,
Carnevale KA, Yakubenko VP, Zhang R, Hwang SH, Hammock BD and
Cathcart MK: Metabolic products of soluble epoxide hydrolase are
essential for monocyte chemotaxis to MCP-1 in vitro and in vivo. J
Lipid Res. 54:436–447. 2013. View Article : Google Scholar :
|
|
26
|
Steffel J, Luscher TF and Tanner FC:
Tissue factor in cardiovascular diseases: Molecular mechanisms and
clinical implications. Circulation. 113:722–731. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Reiss AB and Cronstein BN: Regulation of
foam cells by adenosine. Arterioscler Thromb Vasc Biol. 32:879–886.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Murphy JE, Vohra RS, Dunn S, Holloway ZG,
Monaco AP, Homer-Vanniasinkam S, Walker JH and Ponnambalam S:
Oxidised LDL internalisation by the LOX-1 scavenger receptor is
dependent on a novel cytoplasmic motif and is regulated by
dynamin-2. J Cell Sci. 121:2136–2147. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xie H, Zhou H, Wang H, Chen D, Xia L, Wang
T and Yan J: Anti-β(2)GPI/β(2)GPI induced TF and TNF-α expression
in monocytes involving both TLR4/MyD88 and TLR4/TRIF signaling
pathways. Mol Immunol. 53:246–254. 2013. View Article : Google Scholar
|
|
30
|
Ding Z, Liu S, Wang X, Khaidakov M, Dai Y,
Deng X, Fan Y, Xiang D and Mehta JL: Lectin-like ox-LDL receptor-1
(LOX-1)-toll-like receptor 4 (TLR4) interaction and autophagy in
CATH.a differentiated cells exposed to angiotensin II. Mol
Neurobiol. 51:623–632. 2015. View Article : Google Scholar
|