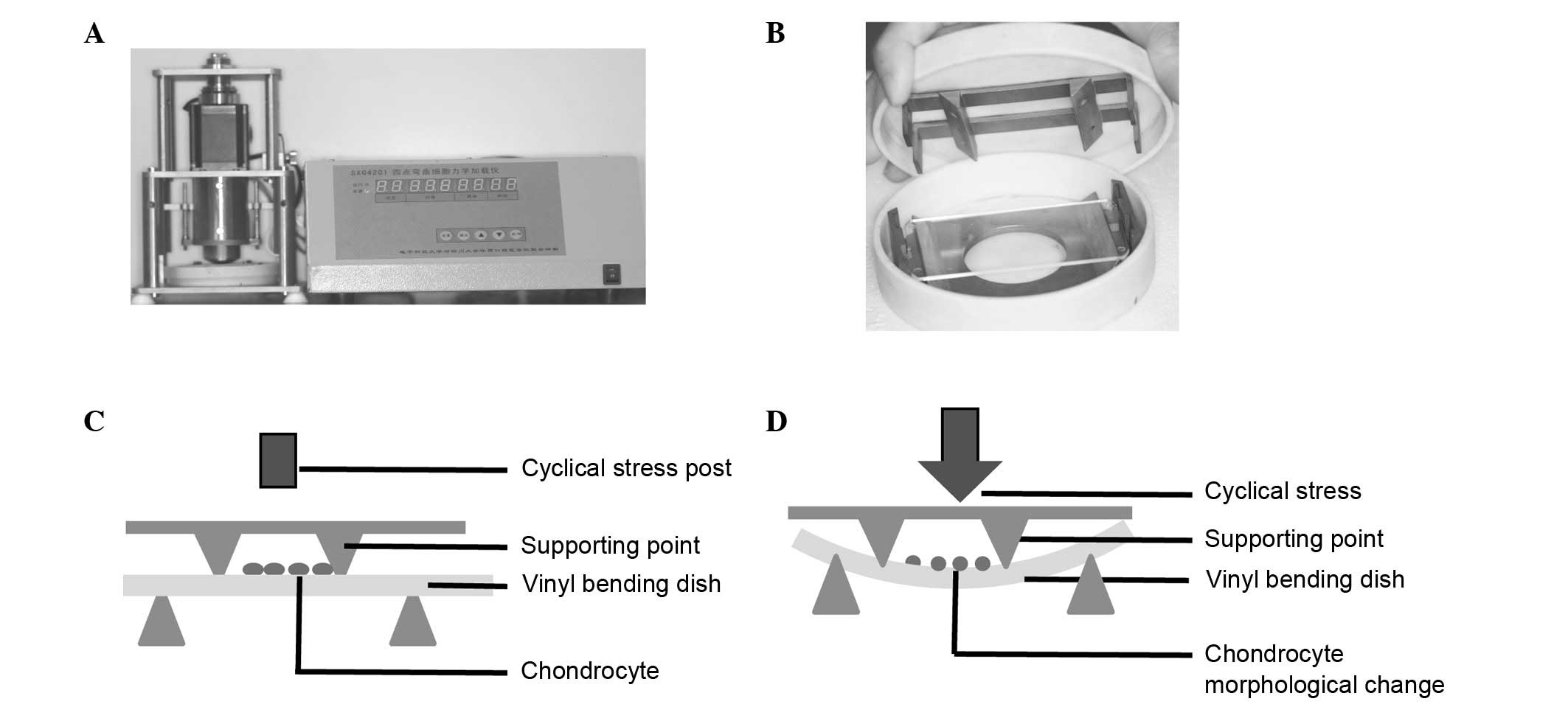
Introduction
Cells sense external stimuli through the
cytoskeleton, facilitated by actomyosin (1,2). In
conjunction with myosin, the cytoskeleton can exert traction on the
surrounding matrix of cells or on adjacent cells, contributing to
major alterations in cell morphology. In addition to sensing
mechanical forces, the cytoskeleton can transform mechanical forces
into biochemical signals. Due to the contractibility of the actin
filaments, proteins associated with the cytoskeleton may be
activated. Myosin II is the main non-muscle class of myosin,
consisting of two sets of myosin heavy chains (200 kDa) and two
sets of myosin light chain (MLC; 16–20 kDa). MLC phosphorylation
serves an important role in various cellular functions, including
contraction and motility. MLC phosphorylation can be activated by
MLC kinase (MLCK) (3).
The temporomandibular joint (TMJ) is functionally
structured to withstand and adapt to mechanical loads (4–6). The
loading condition of the TMJ has been reported to be primarily
dynamic and compressive (4,7).
Comprising less than 10% of the cartilaginous tissue volume,
chondrocytes can sense and respond to their mechanical environment
(5,8). They transduce extracellular stresses
into intracellular signals, resulting in modulation of gene
expression and cellular function, for example, proliferation,
apoptosis, migration and remodeling. It was identified that loading
conditions can be optimized to promote the maintenance of normal
structure and function (9).
The mechanism of signal transduction of mandibular
condylar chondrocytes (MCCs) following application of cyclical
uniaxial compressive stress (CUCS) remains unclear. In the current
study, a computer controlled four-point bending system was used to
apply CUCS to the primary cultured chondrocytes. With this model,
it was aimed to explore whether CUCS may promote differentiation of
chondrocytes. The role of MLC-II in the mechanical stress-induced
cellular differentiation of MCCs was investigated.
Materials and methods
MCC isolation and culture
The animal experiments were approved by the
Committee on the Use of Live Animals in Teaching and Research of
Sun Yat-sen University (Guangzhou, China). Neonatal Sprague-Dawley
rats (n=75; mean weight, 15±2.3g) were purchased from the
Department of Laboratory Animal Science (Sun Yat-sen University). A
total of 5 neonatal rats were used for each experiment. The mother
rats were administered the standard pellet diet ab libitum,
with one mother and baby in each cage. The rats were maintained in
12 h light/dark cycles at a temperature of 18–24°C and a relative
humidity of 40–70%.
Mandibular condylar cartilage was isolated from
7-day-old Sprague-Dawley rats. The rats were sacrificed using 200
mg/kg sodium pentobarbital followed by cervical dislocation.
Mandibular condylar cartilage was digested with 0.25% trypsin
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) at 37°C
for 30 min. Subsequent to centrifugation for 5 min at 120 ×
g at room temperature, mandibular condylar cartilages were
digested with 0.02% type II collagenase (Invitrogen; Thermo Fisher
Scientific, Inc.) in Dulbecco's modified Eagle's medium (DMEM;
Sigma-Aldrich; Merck Millipore, Darmstadt, Germany) for 2 h at
37°C. The supernatant was collected and centrifuged for 5 min at
120 × g at room temperature. Primary condylar chondrocytes
were rinsed three times. Subsequent to counting with a
hemacytometer, cells were seeded in the culture dishes at a high
density of 1.0×106 cells/cm2 with DMEM with
10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.)
and 1% penicillin-streptomycin-amphotericin B at 37°C in a
humidified 5% CO2-containing atmosphere. Passage 3
chondrocytes were seeded on the vinyl bending dishes at a density
of 1.0×106 cells/cm2 and used in the
experiments.
Immunostaining
The chondrocytes were washed with cold
phosphate-buffered saline (PBS) three times and were fixed in 4%
formaldehyde for 15 min at room temperature. The primary
anti-rabbit collagen type II antibody (ab34712; 1:100; Abcam,
Cambridge, MA, USA) was used. A total of five images were randomly
taken in each section under a light microscope (Olympus
Corporation, Tokyo, Japan).
Alcian blue staining
Alcian blue staining was performed as described
previously (10). In brief, the
chondrocytes were fixed with paraformaldehyde (4%, 4°C) for 20 min,
and after three washes with double distilled H2O
(ddH2O), alcian blue (1%) in 3% acetic acid (pH 2.5) was used to
stain for 30 min at room temperature. Thereafter, cells were washed
with ddH2O three times and imaged by phase contrast microscopy
(Olympus Corporation).
Application of CUCS with a four-point
bending system
When the third passage of cells reached 70–80%
confluence on the vinyl bending dishes made from cell culture
dishes (Mirui; Chengdu MIRACLE Technology Co. Ltd., Sichuan,
China), the cells were starved for 12 h in serum-free medium and
then subjected to CUCS (2,000 µstrain for 2 h) using a four-point
bending system (Mirui, Sichuan, China), as described previously
(11). Briefly, the device was
designed based on beam deflection theory and the forces it supplied
were measured by cell deformation, i.e., µstrain (12). Four-point bending system was
consisting of a computer-controlled, servomotor-driven, linear
actuator assembly with an interface controller that controlled
vertical displacement and actuator ram speed (Fig. 1). Cells cultured at a static
station were considered a control.
Immunofluorescence analysis
Immunofluorescence staining was used to detect the
cytoskeleton. Chondrocytes were prepared on vinyl bending dishes at
a density of 5.0×104 cells/cm2. Subsequent to stress
loading, the chondrocytes were washed with cold PBS three times and
fixed in 4% formaldehyde for 15 min at room temperature, followed
by permeabilization with 0.1% Triton X-100 in PBS (Sigma-Aldrich;
Merck Millipore) for 10 min. F-actin was stained with a
Rhodamine-phalloidin mixed solution (Life Technologies; Thermo
Fisher Scientific, Inc.) for 1 h, and after three washes with PBS,
4′,6-diamidino-2-phenylindole (Sigma-Aldrich; Merck Millipore) was
used to stain the nucleolus for 10 min at room temperature. All
fluorescent staining images were taken under a confocal microscopy
(LSM 710; Zeiss, Oberkochen, Germany) using a 40X oil immersion
objective lens.
Alkaline phosphatase (ALP) activity
assay
Cellular ALP activity was determined using the
p-nitrophenyl phosphate method (LabAssay™ ALP; Wako Pure Chemical
Industries, Ltd., Osaka, Japan) according to the manufacturer's
instructions. The enzyme activity (units/mg protein) is equal to
the concentration of p-nitrophenol (nmol/ml) that is released by
the sample within 15 min, subsequent to excluding the background.
The ALP activity of each sample was normalized to the protein
concentration that was detected using a Bicinchoninic Acid (BCA)
protein assay kit (Pierce Biotechnology, Inc., Rockford, IL,
USA).
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was isolated from cultured chondrocytes
using 1 ml of TRIzol (Life Technologies; Thermo Fisher Scientific,
Inc.), after the cells were compressed or submitted to agonist
(blebbistatin) treatment. The total RNA was reverse transcribed
according to the manufacturer's protocol, with 1 µg of RNA. The RT
protocol was as follows: 65°C for 5 min, maintained on ice for 1
min at 42°C for 60 min, 70°C for 10 min and finally maintained at
4°C. Reverse transcription was completed using a Tiangen RNA PCR
kit (Tiangen Biotech Co., Ltd., Beijing, China). Transcriptional
levels of the tested genes runt-related transcription factor 2
(RUNX2) (NC_000083.6) and MLC (NC_000073.6) were quantified by
RT-qPCR with ABI Prism 7000 (Applied Biosystems; Thermo Fisher
Scientific, Inc.). PCR primers are presented in Table I, and Platinum SYBR Green qPCR
Super Mix-UDG (Life Technologies; Thermo Fisher Scientific, Inc.)
was used. RT-qPCR was conducted in 10 µl reaction mixture
containing 2 µl cDNA samples. The thermal cycling conditions were
as follows: 90°C for 15 sec, 62.5°C for 15 sec, and 72°C for 15
sec. The RT-qPCR consisted of 40 cycles, after an initial
denaturation step (94°C for 2 min). The relative quantity of the
mRNA level was obtained using the comparative quantification cycle
(Cq) method (∆∆Cq) (13), with
glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as the internal
reference.
 | Table I.Primer sequences used in reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primer sequences used in reverse
transcription-quantitative polymerase chain reaction.
| Gene | Forward primer | Reverse primer | Size (bp) | Accession
number |
|---|
| RUNX2 |
5-GCCGGGAATGATGAGAACTA-3 |
5-TGGGGAGGATTTGTGAAGAC-3 | 200 | NM 053470.2 |
| MLC |
5-TCAAGAAACAGACCCCCAAG-3 |
5-CCACGATCCTCTCAAAGAGC-3 | 171 | NM 020104.2 |
| ALP |
5-GACAAGAAGCCCTTCACAGC-3 |
5-ACTGGGCCTGGTAGTTGTTG-3 | 118 | NM_013059.1 |
| GAPDH |
5-ATGACTCTACCCACGGCAAG-3 |
5-TACTCAGCACCAGCATCACC-3 | 76 | NC 005111.3 |
Western blot analysis
After the cells were compressed or/and treated with
the agonist, protein was collected by lysis with phosphatase
inhibitor and protease inhibitor (Roche Diagnostics Corporation,
Indianapolis, IN, USA). They were quantified with the BCA protein
assay kit. Proteins (30 µg) were separated with 10% sodium dodecyl
sulfate-polyacrylamide gels, then, electrophoretically transferred
to polyvinylidene difluoride membranes (Roche Diagnostics
Corporation) for 2 h. The membranes were incubated in blocking
buffer [Tris-buffered saline (TBS), 0.1% Tween-20, 2% bovine serum
albumin (Sigma-Aldrich)] at room temperature for 1 h. Subsequent to
washing of the membranes 3 times in TBS (5 min/wash), they were
treated with phosphorylated MLC (cat. no. 3671; 1:1,000; polyclonal
rabbit anti-rat; Cell Signaling Technology, Inc., Danvers, MA,
USA), total MLC (cat. no. 3672; 1:1,000; polyclonal rabbit
anti-rat; Cell Signaling Technology, Inc.) and RUNX2 (cat. no.
ab23981; 1:1,000; polyclonal rabbit anti-rat; Abcam) antibodies
overnight at 4°C. They were incubated with anti-rabbit IgG (cat.
no. 7074; 1:2,000 dilution in TBS-T; Cell Signaling Technology,
Inc. and cat. no. BA1058; 1:1,000 dilution in TBS-T; Wuhan Boster
Biological Technology, Ltd., Wuhan, China) coupled with horseradish
peroxidase for 1 h at room temperature. Protein detection was
performed and visualized using Enhanced Chemiluminescence Advance
Western Blotting Detection Reagent kit (EMD Millipore, Billerica,
MA, USA). Protein loading was estimated using GAPDH (cat. no. 3683;
1:2,500; polyclonal rabbit anti-rat; Cell Signaling Technology,
Inc.). The intensity of the protein fragments was quantified using
the ImageJ version 1.48 software (National Institutes of Health,
Bethesda, MD, USA).
Selective inhibitors of signal
transduction pathways
ML-7 (MLCK inhibitor, 10 µM; Cayman Chemical,
Michigan, MI, USA) and blebbistatin (non-muscle myosin II
inhibitor, 20 µM; Cayman Chemical) were selected to inhibit the MLC
pathways. Briefly, chondrocytes were pretreated with the inhibitors
individually for 1 h prior to the application of compression
(14,15). The inhibitor was also present
during the 2-h CUCS treatment. Subsequently, osteogenic RUNX2 was
assayed at 0 h (mRNA) and 24 h (protein). When the third passage
chondrocytes seeded on vinyl bending reached 50% confluence, the
transfection procedure followed using Lipofectamine™ RNAiMAX
(Invitrogen; Thermo Fisher Scientific, Inc.) with forward
transfection. Briefly, 7.5 µl MLC RNA interference (RNAi) duplex
(Guangzhou RiboBio Co., Ltd., Guangzhou, China) added to 250 µl
Opti-MEM reduced serum medium without serum. A total of 2.5 µl RNAi
MAX (Invitrogen; Thermo Fisher Scientific, Inc.) added to 250 µl
Opti-MEM reduced serum medium without serum. They were mixed at
room temperature for 10 min, and then transferred into the cell
medium without the antibody. At 12 h subsequent to transfection,
the culture medium was replaced with fresh DMEM with 10% FBS.
Transfection efficiency was estimated by immunofluorescence and
RT-qPCR. The RNAi-treated cells were continually cultured after 24
or 48 h and used for the experiments that followed.
Statistical analysis
All data are presented as the mean ± standard error
unless otherwise stated. Statistical analysis was conducted using
one-way analysis of variance or Student's t-test. P<0.05 was
considered to indicate a statistically significant difference. All
of the experiments were performed in triplicate.
Results
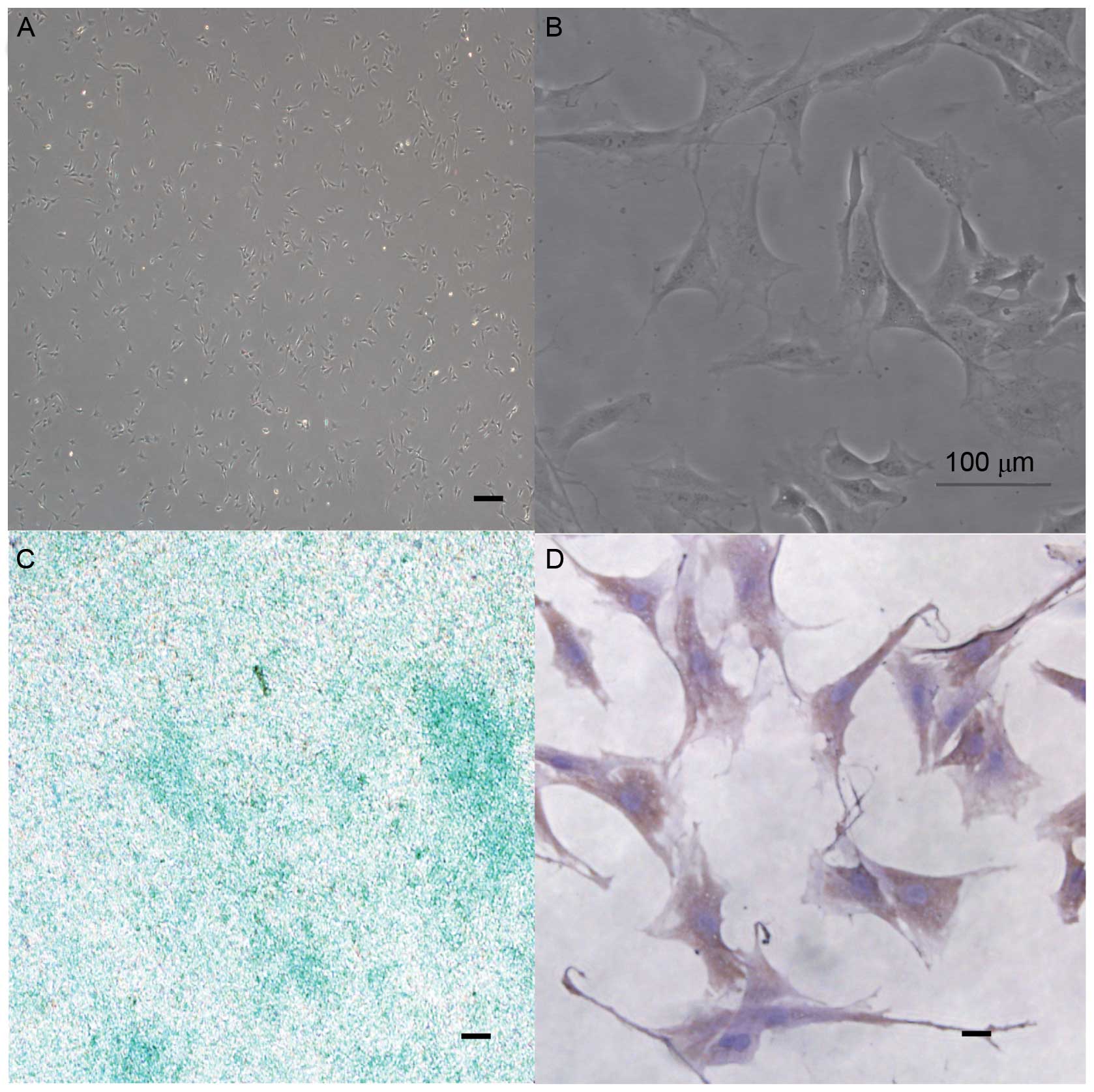
MCCs morphology observed and
identified
Polygonal-shaped, primary MCCs were observed in
cultures at passage 3 (Fig. 2A and
B). Induction of glycosaminoglycan (GAG) synthesis stained
positively with alcian blue (Fig.
2C) and collagen type II stained positive on vinyl bending
dishes (Fig. 2D). These
observations indicated that the isolated cells were MCCs.
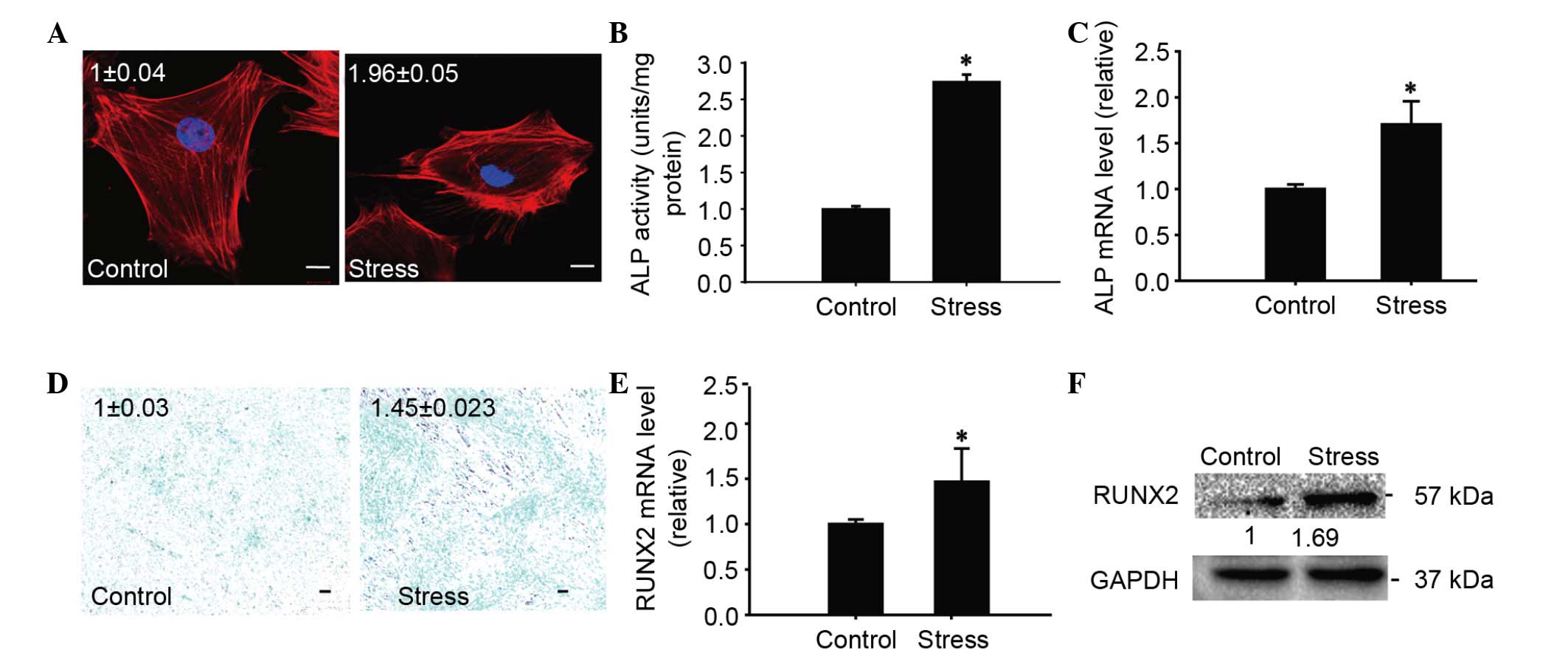
CUCS promoted chondrocyte
differentiation
To investigate the potential role of myosin under
CUCS, the effects of CUCS on the differentiation of chondrocytes
were first evaluated. CUCS treatment was able to induce early
morphological alterations in TMJ chondrocytes. Actin filaments
became thicker and the cells were paralleled with the orientation
of the compression, compared with the random distribution of the
unloaded control cells (Fig. 3A).
ALP mRNA levels were observed to be upregulated immediately, which
was consistent with the ALP activity observed at 48 h post CUCS
(Fig. 3B and C). Subsequently, the
GAG levels increased following CUCS (Fig. 3D). The mRNA levels of the
osteogenic differentiation marker RUNX2 were increased following
the CUCS treatment (P<0.01; Fig.
3E). The protein levels of RUNX2 increased at 24 h subsequent
to the CUCS treatment, in accordance with the gene expression
(P<0.05; Fig. 3F).
CUCS elevated the expression of TMJ
chondrocyte contractile markers
It was identified that subsequent to CUCS treatment,
the levels of MLC increased rapidly (P<0.05; Fig. 4A), while proportions of
phosphorylated MLC and the total MLC increased immediately
subsequent to CUCS treatment, in agreement with the gene expression
results (P<0.05; Fig. 4B). To
test the role of the MLCK in response to CUCS, chondrocytes were
treated with one of the dominant negative MLCK inhibitors (ML-7).
The phosphorylation of MLC was reduced (P<0.05) by CUCS,
compared with the non-treated control cells (Fig. 4C). To investigate the role of MLC
in the response to CUCS, chondrocytes were treated with non-muscle
myosin II inhibitors (blebbistatin). The inhibitor blebbistatin
marginally inhibited the expression of phosphorylated MLC-II
(P>0.05), while the applied stress significantly increased the
expression of phosphorylated MLC-II (P<0.05; Fig. 4C). To further test the role of
myosin, RNAi was used. The transfection efficiency was
approximately 85% following 24 h transfection (Fig. 4D and E).
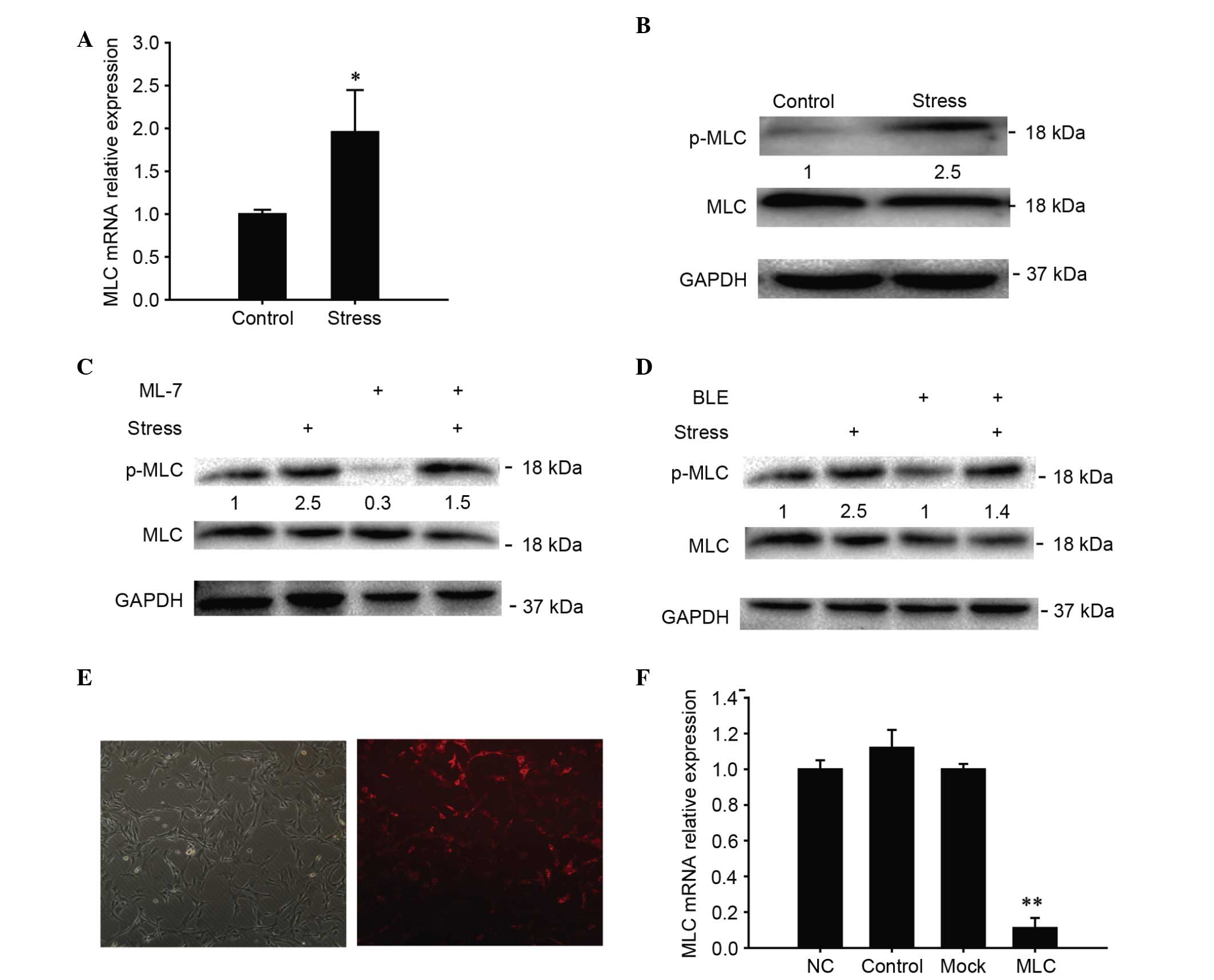 | Figure 4.Involvement of MLC in cyclical
uniaxial stress. (A) Transcriptional levels of MLC in chondrocytes
determined by RT-qPCR at 0 h after treatment. (B) Western blot
analysis of MLC phosphorylation following exposure of chondrocytes
to CUCS or the control. (C) Alterations in p-MLC subsequent to
exposure of the chondrocytes to the control, CUCS, BLE, CUCS + BLE,
ML-7 and CUCS + ML-7. (D and E) MCCs subsequent to RNA interference
transfection at 24 h (magnification, ×40). Cells stained red on the
right panel indicate the positively transfected cells (~85%). (F)
MLC silencing efficiency was assessed. The silencing efficiency of
myosin light chain was ~85% 24 h post-transfection. NC group refers
to cells transfected with a sequence that has no homology to the
target sequence. Control group refers to untransfected cells. Mock
group refers to untransfected cells treated with the transfection
reagent. Data are presented as the mean ± standard error; n=3;
*P<0.05, **P<0.01 compared with the NC. MLC, myocin light
chain; CUCS, cyclical uniaxial compressive stress; p-,
phosphorylated; BLE, blebbistatin; MCC, mandibular condylar
chondrocyte; ALP, alkaline phosphatase; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; RUNX2,
runt-related transcription factor 2; GAPDH, glyceraldehyde
3-phosphate dehyrdrogenase; NC, culture cell with an added sequence
with no homology to target sequence. |
Inhibition of MLCK significantly
reduced the differentiation of cultured chondrocytes
ALP activity reduced (P<0.05; Fig. 5A). Similar results were observed
for RUNX2 protein expression after CUCS and addition of the
inhibitor (Fig. 5B). The results
indicated that the inhibitor-treated cells exhibited a significant
reduction in RUNX2 mRNA expression after CUCS and inhibitor
treatment, as presented in Fig. 5C
(P<0.05).
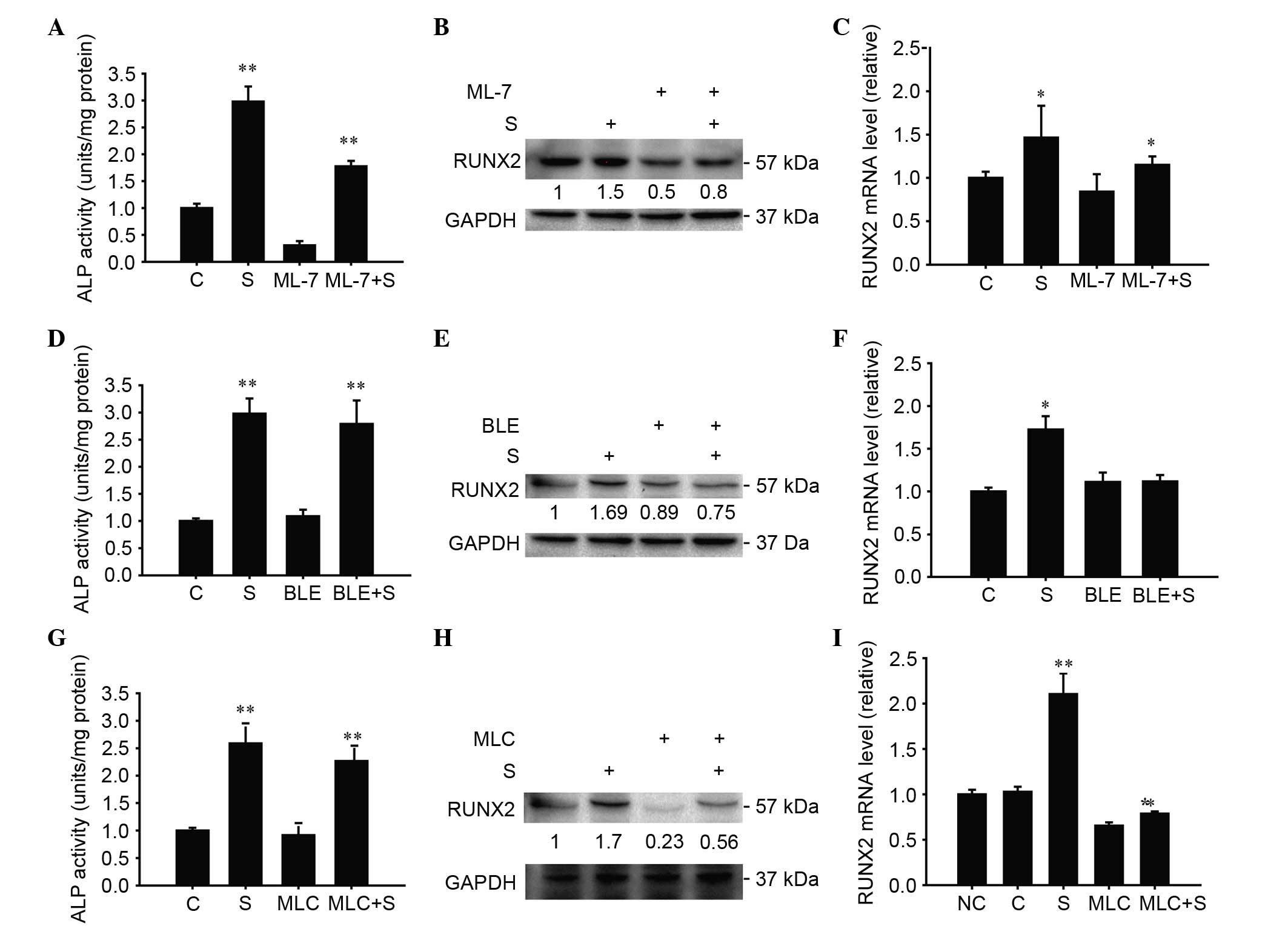 | Figure 5.MLC governs CUCS-induced
differentiation in MCCs. (A) ALP activity quantification at 48 h
subsequent to involvement of ML-7 in the CUCS stress assay. (B)
Western blot analysis of RUNX2 protein expression at 24 h
subsequent to involvement of ML-7. (C) Transcriptional levels of
RUNX2 in chondrocytes, determined by RT-qPCR at 0 h after addition
of ML-7. (D) ALP activity quantification at 48 h subsequent to
addition of BLE in the CUCS stress assay. (E) Western blot analysis
of RUNX2 protein expression at 24 h after addition of BLE. (F)
Transcriptional levels of RUNX2 in chondrocytes, determined by
RT-qPCR at 0 h after addition of of BLE. (G) ALP activity
quantification at 48 h after CUCS, a significant reduction was
observed subsequent to CUCS with RNAi treatment. (H) Alterations in
RUNX2 protein expression at 24 h subsequent to chondrocyte exposure
to RNAi treatment. (I) Transcriptional levels of RUNX2 in
chondrocytes, determined by RT-qPCR at 0 h with RNAi treatment.
Data are presented as the mean ± standard error; n=3; *P<0.05,
**P<0.01 compared with the NC. MLC, myocin light chain; CUCS,
cyclical uniaxial compressive stress; MCC, mandibular condylar
chondrocyte; ALP, alkaline phosphatase; RUNX2, runt-related
transcription factor 2; RT-qPCR, reverse transcription-quantitative
polymerase chain reaction; BLE, blebbistatin; RNAi, RNA
interference; C, control; S, stress; GAPDH, glyceraldehyde
3-phosphate dehyrdrogenase; NC, culture cell with an added sequence
with no homology to target sequence. |
Inhibition of MLC partially reduced
the differentiation of cultured chondrocytes
ALP activity was upregulated 48 h post-CUCS
(P<0.05), while ALP activity was altered following addition of
the inhibitor and the stress treatment (P<0.05; Fig. 5D). At the same time, RUNX2 protein
expression and mRNA remained unchanged following addition of the
inhibitor and stress treatment (P>0.05) (Fig. 5E and F). The clear increase in ALP
RNAi-transfected chondrocytes was statistically significant
following CUCS compared with RNAi-transfected cells (Fig. 5G). The RUNX2 protein and mRNA
expression increased following addition of the RNAi and the stress
treatment (P<0.05; Fig. 5H and
I).
Discussion
Mechanical stress of the TMJ is commonly caused by
talking, mastication or orthodontic treatment (4–6). The
ability chondrocytes to respond to mechanical load is important for
the maintenance of normal function of articular cartilage (16–18).
Previous studies have indicated that exogenous mechanical stress
may induce a series of cellular alterations including gene
expression, protein phosphorylation and cellular function (19–24).
In the current study, in order to simulate natural joint status,
primary MCCs from rats were exposed to cyclic forces of 2,000
µstrain (12).
RUNX2 is one of the most important transcription
factors of hypertrophic differentiation (25–27).
It is expressed in pre-hypertrophic and hypertrophic chondrocytes,
and RUNX2 has been reported to promote chondrocyte hypertrophy
prior to endochondral ossification (28). In RUNX2 knockout mice, a previous
study observed that bone maturation was delayed (29,30).
Overexpression of RUNX2 induces hypertrophic differentiation
(31), and it was reported that
stress may increase the expression of RUNX2 in human chondrocytes
(32). In the current study, the
mRNA and protein levels of RUNX2 were identified to be
significantly upregulated following CUCS treatment.
ALP is predominantly expressed in hypertrophic
chondrocytes (33) and it is
commonly used as a marker for osteogenic differentiation. In ALP
knockout mice, a previous study observed that cartilage
mineralization was reduced (34)
and additional studies reported that stress increased ALP activity
in MC3T3 cells (35) and human
periodontal ligament cells (36).
The results of the current study demonstrated that CUCS promoted
the differentiation of primary MCCs in vitro, indicated by
significantly increased expression levels of RUNX2 and increased
ALP activity.
It has been previously demonstrated that the
cytoskeleton is associated with mechanical stress (37–39).
In the present study, it was observed that cytoskeletal
rearrangement occurred immediately subsequent to CUCS treatment,
with part of the actin filaments accumulating together around the
cell membrane. Li et al (11) reported that cytoskeletal
rearrangement was induced with higher levels of stress (4,000
µstrain) compared with that used in the current study (2,000
µstrain). It is suggested that the age of rat may affect the
ability of the chondrocytes to sense and respond to different level
of exogenous mechanical stress (40). The viscoelastic properties of older
cells are commonly lower than that of younger cells (41) and it has been reported that the
cytoskeleton is closely linked with with cellular
differentiation-associated proteoglycan and collagen synthesis in
chondrocytes (42). In the present
study, when the cytoskeleton was disrupted with cytochalasin D, the
differentiation associated with alterations of the MCCs was
observed to be significantly reduced, indicating that the
cytoskeleton was involved in signal transduction of MCCs in the
experimental system (data not shown).
MLC-II has been demonstrated to control actomyosin
contractility in various types of cells (43), and phosphorylated MLC-II adjusts
the activity of bone cells and acts as an important effector of
cell function (44,45). In drosophila, the differentiation
of cells has been inhibited by replacing phosphorylated MLC-II with
MLC-II (46). In the current
study, silencing MLC-II using RNAi resulted in downregulation of
chondrocyte differentiation, as presented by the reduction in ALP
activity and RUNX2 mRNA levels. This indicated that MLC-II serves
an important role in MCC differentiation. The MLCK inhibitor, ML-7,
aimed to inhibit the phosphorylation of MLC-II, and induced a
reduction of ALP activity and downregulation of RUNX2 expression.
Taken together, the results suggest that CUCS promotes rat primary
MCC differentiation through phosphorylated MLC-II.
The four-point bending system used in the current
study provides an in vitro model for the study of cellular
responses to mechanical stress. The signaling transferred from
outside mechanical stress to inside of cells was also reported to
be responsible by monolayer chondrocyte. Sanz-Ramos et al
(47) described that the
expression levels of integin α1, β1 and β3 in monolayer
chondrocytes were sensitive to matrix stiffness, similar to the
response of 3D-cultured chondrocytes, however this mechanism
remains unclear. In the current study, the chondrocytes cultured on
the slides were monolayer, however it remains unclear whether the
culture strategy may influence how these cells perceive mechanical
signals. Further study with chondrocytes subjected to compressive
strain cultured in 3D matrices may address this issue.
Acknowledgements
The current study was supported by Grants from the
National Natural Science Foundation of China (grant no. 81070860)
and the Natural Science Foundation of Guangdong Province (grant no.
s2013010016753). The authors would like to thank Professor Guangmei
Yan (Sun Yat-sen University) and his group for providing technical
support.
References
|
1
|
Maruthamuthu V, Aratyn-Schaus Y and Gardel
ML: Conserved F-actin dynamics and force transmission at cell
adhesions. Curr Opin Cell Biol. 22:583–588. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Papusheva E and Heisenberg CP: Spatial
organization of adhesion: Force-dependent regulation and function
in tissue morphogenesis. EMBO J. 29:2753–2768. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sarasa-Renedo A, Tunç-Civelek V and
Chiquet M: Role of RhoA/ROCK-dependent actin contractility in the
induction of tenascin-C by cyclic tensile strain. Exp Cell Res.
312:1361–1370. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tanaka E and van Eijden T: Biomechanical
behavior of the temporomandibular joint disc. Crit Rev Oral Biol
Med. 14:138–150. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mori H, Horiuchi S, Nishimura S, Nikawa H,
Murayama T, Ueda K, Ogawa D, Kuroda S, Kawano F, Naito H, et al:
Three-dimensional finite element analysis of cartilaginous tissues
in human temporomandibular joint during prolonged clenching. Arch
Oral Biol. 55:879–886. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kuroda S, Tanimoto K, Izawa T, Fujihara S,
Koolstra JH and Tanaka E: Biomechanical and biochemical
characteristics of the mandibular condylar cartilage.
Osteoarthritis Cartilage. 17:1408–1415. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tanaka E, Sasaki A, Tahmina K, Yamaguchi
K, Mori Y and Tanne K: Mechanical properties of human articular
disk and its influence on TMJ loading studied with the finite
element method. J Oral Rehabil. 28:273–279. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hu K, Radhakrishnan P, Patel RV and Mao
JJ: Regional structural and viscoelastic properties of
fibrocartilage upon dynamic nanoindentation of the articular
condyle. J Struct Biol. 136:46–52. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Walsh AJ and Lotz JC: Biological response
of the intervertebral disc to dynamic loading. J Biomech.
37:329–337. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ronziere MC, Roche S, Gouttenoire J,
Démarteau O, Herbage D and Freyria AM: Ascorbate modulation of
bovine chondrocyte growth, matrix protein gene expression and
synthesis in three-dimensional collagen sponges. Biomaterials.
24:851–861. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li H, Yang HS, Wu TJ, Zhang XY, Jiang WH,
Ma QL, Chen YX, Xu Y, Li S and Hua ZC: Proteomic analysis of
early-response to mechanical stress in neonatal rat mandibular
condylar chondrocytes. J Cell Physiol. 223:610–622. 2010.PubMed/NCBI
|
|
12
|
Owan I, Burr DB, Turner CH, Qiu J, Tu Y,
Onyia JE and Duncan RL: Mechanotransduction in bone: Osteoblasts
are more responsive to fluid forces than mechanical strain. Am J
Physiol. 273:C810–C815. 1997.PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Saitoh M, Ishikawa T, Matsushima S, Naka M
and Hidaka H: Selective inhibition of catalytic activity of smooth
muscle myosin light chain kinase. J Biol Chem. 262:7796–7801.
1987.PubMed/NCBI
|
|
15
|
Engler AJ, Sen S, Sweeney HL and Discher
DE: Matrix elasticity directs stem cell lineage specification.
Cell. 126:677–689. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu M, Xu T, Zhou Y, Lu H and Gu Z:
Pressure and inflammatory stimulation induced increase of
cadherin-11 is mediated by PI3K/Akt pathway in synovial fibroblasts
from temporomandibular joint. Osteoarthritis Cartilage.
21:1605–1612. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Palmoski MJ, Colyer RA and Brandt KD:
Joint motion in the absence of normal loading does not maintain
normal articular cartilage. Arthritis Rheum. 23:325–334. 1980.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kiviranta I, Jurvelin J, Tammi M, Säämänen
AM and Helminen HJ: Weight bearing controls glycosaminoglycan
concentration and articular cartilage thickness in the knee joints
of young beagle dogs. Arthritis Rheum. 30:801–809. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wong M, Siegrist M and Goodwin K: Cyclic
tensile strain and cyclic hydrostatic pressure differentially
regulate expression of hypertrophic markers in primary
chondrocytes. Bone. 33:685–693. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Heyland J, Wiegandt K, Goepfert C,
Nagel-Heyer S, Ilinich E, Schumacher U and Pörtner R:
Redifferentiation of chondrocytes and cartilage formation under
intermittent hydrostatic pressure. Biotechnol Lett. 28:1641–1648.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Batra NN, Li YJ, Yellowley CE, You L,
Malone AM, Kim CH and Jacobs CR: Effects of short-term recovery
periods on fluid-induced signaling in osteoblastic cells. J
Biomech. 38:1909–1917. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liedert A, Kaspar D, Blakytny R, Claes L
and Ignatius A: Signal transduction pathways involved in
mechanotransduction in bone cells. Biochem Biophys Res Commun.
349:1–5. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sibonga JD, Zhang M, Evans GL, Westerlind
KC, Cavolina JM, Morey-Holton E and Turner RT: Effects of
spaceflight and simulated weightlessness on longitudinal bone
growth. Bone. 27:535–540. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wong M and Carter DR: Articular cartilage
functional histomorphology and mechanobiology: A research
perspective. Bone. 33:1–13. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wuelling M and Vortkamp A: Chondrocyte
proliferation and differentiation. Endocr Dev. 21:1–11.
2011.PubMed/NCBI
|
|
26
|
Ding M, Lu Y, Abbassi S, Li F, Li X, Song
Y, Geoffroy V, Im HJ and Zheng Q: Targeting Runx2 expression in
hypertrophic chondrocytes impairs endochondral ossification during
early skeletal development. J Cell Physiol. 227:3446–3456. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang S, Xiao Z, Luo J, He N, Mahlios J
and Quarles LD: Dose-dependent effects of Runx2 on bone
development. J Bone Miner Res. 24:1889–1904. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Goldring MB, Tsuchimochi K and Ijiri K:
The control of chondrogenesis. J Cell Biochem. 97:33–44. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ducy P, Zhang R, Geoffroy V, Ridall AL and
Karsenty G: Osf2/Cbfa1: A transcriptional activator of osteoblast
differentiation. Cell. 89:747–754. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Komori T, Yagi H, Nomura S, Yamaguchi A,
Sasaki K, Deguchi K, Shimizu Y, Bronson RT, Gao YH, Inada M, et al:
Targeted disruption of Cbfa1 results in a complete lack of bone
formation owing to maturational arrest of osteoblasts. Cell.
89:755–764. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Komori T: Regulation of skeletal
development by the Runx family of transcription factors. J Cell
Biochem. 95:445–453. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Saito T, Nishida K, Furumatsu T, Yoshida
A, Ozawa M and Ozaki T: Histone deacetylase inhibitors suppress
mechanical stress-induced expression of RUNX-2 and ADAMTS-5 through
the inhibition of the MAPK signaling pathway in cultured human
chondrocytes. Osteoarthritis Cartilage. 21:165–174. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pfander D, Swoboda B and Kirsch T:
Expression of early and late differentiation markers (proliferating
cell nuclear antigen, syndecan-3, annexin VI, and alkaline
phosphatase) by human osteoarthritic chondrocytes. Am J Pathol.
159:1777–1783. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mackie EJ, Ahmed YA, Tatarczuch L, Chen KS
and Mirams M: Endochondral ossification: How cartilage is converted
into bone in the developing skeleton. Int J Biochem Cell Biol.
40:46–62. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mai Z, Peng Z, Wu S, Zhang J, Chen L,
Liang H, Bai D, Yan G and Ai H: Single bout short duration fluid
shear stress induces osteogenic differentiation of MC3T3-E1 cells
via integrin β1 and BMP2 signaling cross-talk. PLoS One.
8:e616002013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tang M, Peng Z, Mai Z, Chen L, Mao Q, Chen
Z, Chen Q, Liu L, Wang Y and Ai H: Fluid shear stress stimulates
osteogenic differentiation of human periodontal ligament cells via
the extracellular signal-regulated kinase 1/2 and p38
mitogen-activated protein kinase signaling pathways. J Periodontol.
85:1806–1813. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Durrant LA, Archer CW, Benjamin M and
Ralphs JR: Organisation of the chondrocyte cytoskeleton and its
response to changing mechanical conditions in organ culture. J
Anat. 194:343–353. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Broom ND and Myers DB: A study of the
structural response of wet hyaline cartilage to various loading
situations. Connect Tissue Res. 7:227–237. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ohashi N, Robling AG, Burr DB and Turner
CH: The effects of dynamic axial loading on the rat growth plate. J
Bone Miner Res. 17:284–292. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chahine NO, Blanchette C, Thomas CB, Lu J,
Haudenschild D and Loots GG: Effect of age and cytoskeletal
elements on the indentation-dependent mechanical properties of
chondrocytes. PLoS One. 8:e616512013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Duan W, Wei L, Zhang J, Hao Y, Li C, Li H,
Li Q, Zhang Q, Chen W and Wei X: Alteration of viscoelastic
properties is associated with a change in cytoskeleton components
of ageing chondrocytes from rabbit knee articular cartilage. Mol
Cell Biomech. 8:253–274. 2011.PubMed/NCBI
|
|
42
|
Mallein-Gerin F, Garrone R and van der
Rest M: Proteoglycan and collagen synthesis are correlated with
actin organization in dedifferentiating chondrocytes. Eur J Cell
Biol. 56:364–373. 1991.PubMed/NCBI
|
|
43
|
Alenghat FJ and Ingber DE:
Mechanotransduction: All signals point to cytoskeleton, matrix and
integrins. Sci STKE. 2002:pe62002.PubMed/NCBI
|
|
44
|
Matsumura F and Hartshorne DJ: Myosin
phosphatase target subunit: Many roles in cell function. Biochem
Biophys Res Commun. 369:149–156. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Goeckeler ZM, Bridgman PC and Wysolmerski
RB: Nonmuscle myosin II is responsible for maintaining endothelial
cell basal tone and stress fiber integrity. Am J Physiol Cell
Physiol. 295:C994–C1006. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Jordan P and Karess R: Myosin light
chain-activating phosphorylation sites are required for oogenesis
in Drosophila. J Cell Biol. 139:1805–1819. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Sanz-Ramos P, Mora G, Ripalda P,
Vicente-Pascual M and Izal-Azcárate I: Identification of signalling
pathways triggered by changes in the mechanical environment in rat
chondrocytes. Osteoarthritis Cartilage. 20:931–939. 2012.
View Article : Google Scholar : PubMed/NCBI
|