Introduction
Bone remodeling is a balance between bone resorption
by osteoclasts and bone formation by osteoblasts (1). When this balance tips toward excess
resorption, the risk of osteoporosis is increased, whereas
osteoclast dysfunction increases the risk of osteopetrosis
(2). Thus, osteoclasts have an
important function in bone homeostasis. In orthodontic treatment,
osteoclasts are also important for tooth movement (3,4).
Promotion of osteoclast differentiation accelerates bone
resorption. By contrast, inhibiting osteoclast differentiation
accelerates formation. Orthodontic force consists of tensile and
compressive force. On the pressure side, osteoclasts are subjected
to compressive force. By contrast, on the tension side, osteoclasts
are subjected to tensile force. During orthodontic treatment,
numerous stimuli are applied to osteoclasts; certain signals
promote osteoclast differentiation, whereas others inhibit
osteoclast differentiation (3).
Osteoclasts are multinucleated, bone-resorbing cells
that are differentiated from the monocyte/macrophage hematopoietic
lineage (5). Receptor activator of
nuclear factor-κB (RANK)-ligand (RANKL) is an essential factor for
osteoclast differentiation. RANKL binding to RANK induces
expression of nuclear factor of activated T cells 1 (NFATc1).
NFATc1 is the master transcription factor for osteoclast
differentiation (6). An increase
in the expression of NFATc1 promotes transcription of various
osteoclast-specific genes. Numerous studies have previously
reported that stimulation of osteoclasts leads to secretion of
cytokines associated with bone resorption and formation (7–10).
In a previous study, various types of mechanical
stimuli were applied to osteoclasts, and it was reported that these
were influential factors in bone remodeling. Mechanical stimuli
include tensile force (8,9,11–14),
compressive force (7,10,15–17),
hydrostatic pressure (18), sheer
stress (19,20), rotative stress (21) and others (22,23).
Stimulation with tensile force using a Flexercell tension system
suppresses osteoclast differentiation and fusion. The number of
osteoclasts increases rapidly after the release of tensile force.
Additionally, optimal compressive force induces osteoclast
differentiation (9). In this
experiment, osteoclasts on slips are reversed and placed on
collagen gel layers, and compressive force was adjusted using
weights. Optimal compressive force is defined as the weight that
induces the largest increase in the number of osteoclasts, and is
280 mg/cm2 according to Hayakawa et al (10). Thus, the present study investigated
the effects of release from optimal compressive force on
osteoclasts.
Materials and methods
Cell culture
The current study used the murine
monocyte/macrophage cell line RAW264.7 cells (TIB-71TM; American
Type Culture Collection, Manassas, VA, USA) as osteoclast
precursors. Cells were maintained in Dulbecco's modified Eagle's
medium (Wako Pure Chemical Industries, Ltd., Osaka, Japan)
containing 10% heat-inactivated fetal bovine serum (FBS;
Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and
66.7 µg/ml kanamycin sulfate (Meiji Seika Kaisha, Ltd., Tokyo,
Japan) at 37°C in 5% CO2 in humidified air. Cells were seeded in
100-mm standard dishes (Corning Incorporated, Corning, NY, USA) and
incubated overnight. Subsequently, for osteoclast differentiation,
RAW cells (1×104 cells/well) were transferred into 24-well culture
plates (Corning Incorporated) and cultured on 12-mm diameter glass
cover slips (Fisher Microscope Cover Glass; Thermo Fisher
Scientific, Inc.) placed on the 24-well culture plate, in α-minimum
essential medium (α-MEM; Wako Pure Chemical Industries, Ltd.)
supplemented with 10% heat-inactivated FBS, 2 mM
L-alanyl-L-glutamine (Wako Pure Chemical Industries, Ltd.), 284 µM
L-ascorbic acid phosphate magnesium salt n-hydrate (Wako Pure
Chemical Industries, Ltd.), 66.7 µg/ml kanamycin sulfate and 50
ng/ml RANKL (Oriental Yeast Co., Ltd., Tokyo Japan) at 37°C under
5% CO2 in humidified air. Medium was changed every other day.
Preparation of collagen gels
The collagen mixture comprised acid-soluble collagen
solution (Cellmatrix; Nitta Gelatin NA Inc., Morrisville, NC, USA)
mixed with 10-fold concentrated α-MEM and reconstruction buffer
(2.2 g NaHCO3 + 4.77 g HEPES in 100 ml 0.05 N NaOH; Nitta Gelatin
NA Inc.) at a volume ratio of 8:1:1, and supplemented with 10%
heat-inactivated FBS (Invitrogen; Thermo Fisher Scientific, Inc.),
284 µM L-ascorbic acid 2-phosphate (Sigma-Aldrich; Merck Millipore,
Darmstadt, Germany) and 2 mM L-alanyl-L-glutamine (Wako Pure
Chemical Industries, Ltd.) at below 4°C. This collagen mixture was
placed into 24-well culture plates at a volume of 500 µl/well, and
was solidified in a CO2 incubator at 37°C for 30 min. After
gelation, 1 ml/well culture medium supplemented with 50 ng/ml RANKL
was overlaid in each well.
Application of compressive forces
RAW264.7 cells (1×104 cells/well) were
cultured for 3 days on thin glass slips (diameter, 12 mm;
thickness, 0.1 mm). Osteoclasts on slips were then inverted and
placed on collagen gel layers that were prepared in other 24-well
culture plates. Compressive force was adjusted by placing a weight
on a slip (10). Seven layered
slips (~40 mg/slip) were used as a weight. Cells were subjected to
280 mg/cm2 of compressive force for 24 h, whereas control cells
were inverted and placed on collagen gel layers without
weights.
Release from compressive forces
After cells had been compressed and incubated for 24
h, weights were removed, and cells on slips were inverted.
Subsequently, cells were incubated for 24 h.
Tartrate-resistant acid phosphatase
(TRAP) staining
After cells were cultured for a 2, 3, 4, 5 or 6
days, and fixed with 10% neutral formalin for 30 min at room
temperature, they were washed with distilled water and treated with
TRAP staining solution (pH 5.0) supplemented with Fast Red Violet
LB Salt (Sigma Aldrich; Merck Millipore) (24). TRAP staining solution contained
acetate buffer (pH 5.0; Sigma-Aldrich; Merck Millipore), naphthol
AS-MX phosphate (Sigma-Aldrich; Merck Millipore) as a substrate,
red violet LB (Sigma-Aldrich; Merck Millipore) as a stain in the
presence of 50 mM sodium tartrate (Wako Pure Chemical Industries,
Ltd.). TRAP-positive cells with ≥2 nuclei were counted under the
microscope as osteoclasts. TRAP-positive cells with 2–7 nuclei were
considered to be small osteoclasts, and those with ≥8 nuclei to be
large osteoclasts (8,9).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Cells were incubated for 1, 3, 6, 12 and 24 h in
24-well culture plates, after compression for 24 h. Total RNA was
isolated from cultured cells under each set of conditions using
TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.) (25) according to the manufacturer's
instructions, and aliquots containing equal amounts of mRNA were
subjected to RT-qPCR. First-strand cDNA synthesis was performed
using 1 µg total RNA, 25 pmol oligo dT (Toyobo Co., Ltd., Osaka,
Japan), 1 mM dNTP (Toyobo Co., Ltd.), 100 U ReverTra
Ace® (Toyobo Co., Ltd.), 20 U RNase inhibitor (Toyobo
Co., Ltd.) and 4 µl 5X reaction mixture (Toyobo Co., Ltd.) in 20 µl
with annealing at 30°C for 10 min, enzyme reaction at 42°C for 20
min, denaturation at 99°C for 5 min and cooling at 4°C. qPCR was
performed using the ABI Prism 7300 sequence detection system
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The reactions
were incubated at 50°C for 2 min and 95°C 10 min, followed by 40
cycles of 95°C for 15 sec and annealing at 60°C for 1 min. The
following specific TaqMan probes (Applied Biosystems; Thermo Fisher
Scientific, Inc.) for osteoclast-associated genes were used: NFATc1
(ID no. Mm00479445_ml), TRAP (ID no. Mm00475698_ml), matrix
metalloproteinase-9 (MMP-9; ID no. Mm00432271_ml), RANK (ID no.
Mm-00437135_ml), osteoclast stimulatory trans membrane protein
(OC-STAMP; ID no. Mm00512445_ml), dendritic cell specific trans
membrane protein (DC-STAMP; ID no. Mm01168058_ml), cathepsin-K
(Cath-K; ID no. Mm00484036_ml), chloride channel 7 (ClC-7; ID no.
Mm00442400_ml) and ATPase H+ transporting vacuolar proton pump
member I (ATP6i; ID no. Mm00469395_gl). Levels of mRNA expression
were calculated and standardized against the level of
glyceraldehyde 3-phosphate dehydrogenase (GAPDH; ID no.
Mm99999915_gl) mRNA. Following each PCR run, data were analyzed by
the system and amplification plots were obtained. qPCR results were
calculated using the 2-ΔΔCq method (26).
Statistical analysis
Values represent the mean ± standard deviation.
Comparisons between two groups were analyzed by 2-tailed unpaired
Student's t test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Number of osteoclasts on various
culture days
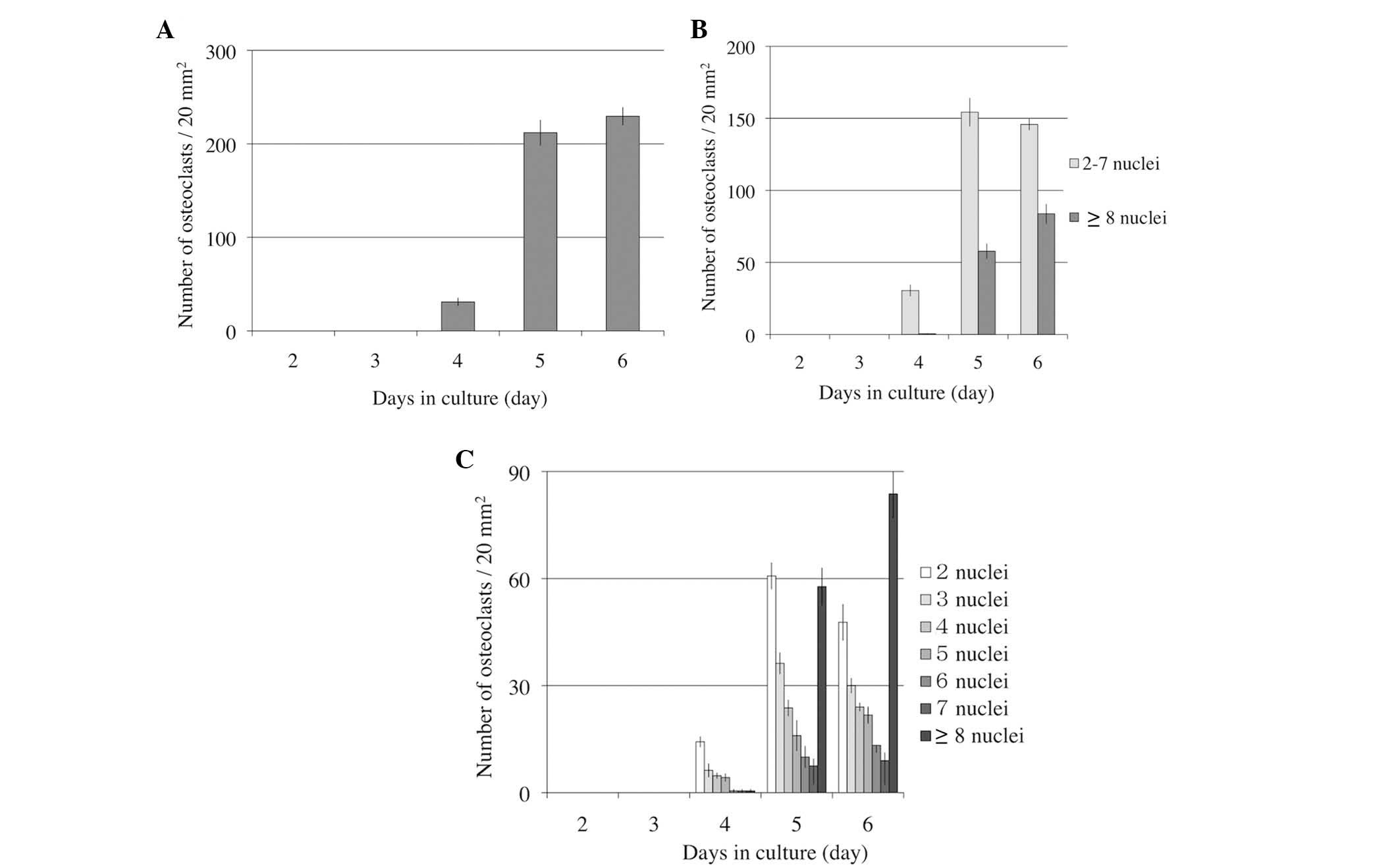
In order to determine when osteoclast
differentiation and fusion were activated, the number of
osteoclasts and the nuclei in osteoclasts were observed for 6 days.
Osteoclasts induced from RAW264.7 cells were cultured with 50 ng/ml
RANKL on 24-well culture plates for 6 days. The number of
osteoclasts increased rapidly on day 5, and marginally on day 6
(Fig. 1A). Small osteoclasts (2–7
nuclei) did not increase in number, but large osteoclasts (≥8
nuclei) increased on day 6 (Fig.
1B). Even in small osteoclasts, osteoclasts with 2–3 nuclei
decreased, however osteoclasts with 4–7 nuclei did not increase
(Fig. 1C).
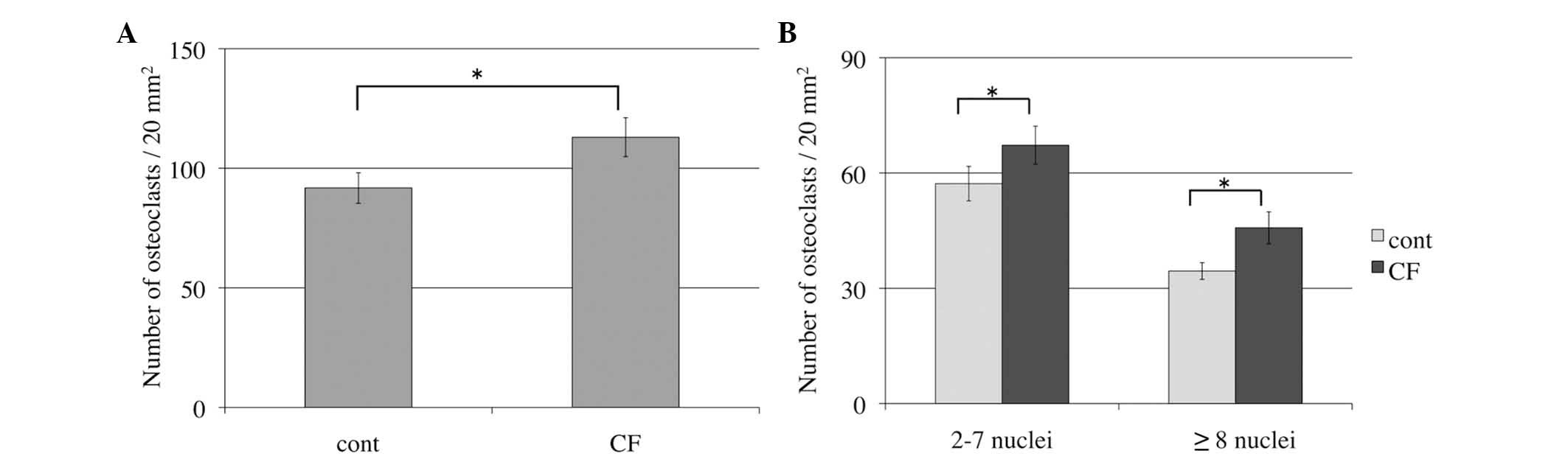
Number of osteoclasts increases in
response to compressive force during osteoclast
differentiation
In order to determine whether compressive force
induced osteoclast differentiation, the number of TRAP-positive
multinucleated osteoclasts was assessed by TRAP staining.
Compressive force was applied for 24 h, whereas control cells were
incubated under the same conditions, but without compressive force.
TRAP-positive multinucleated cells with >2 nuclei were counted
under the microscope. The number of osteoclasts with compressive
force was increased significantly compared with the control group
(P=0.03; Fig. 2A). Furthermore,
the number of small osteoclasts (2–7 nuclei) and large osteoclasts
(≥8 nuclei) with compressive force was significantly increased
compared with the control groups (P=0.04 and P=0.03, respectively;
Fig. 2B).
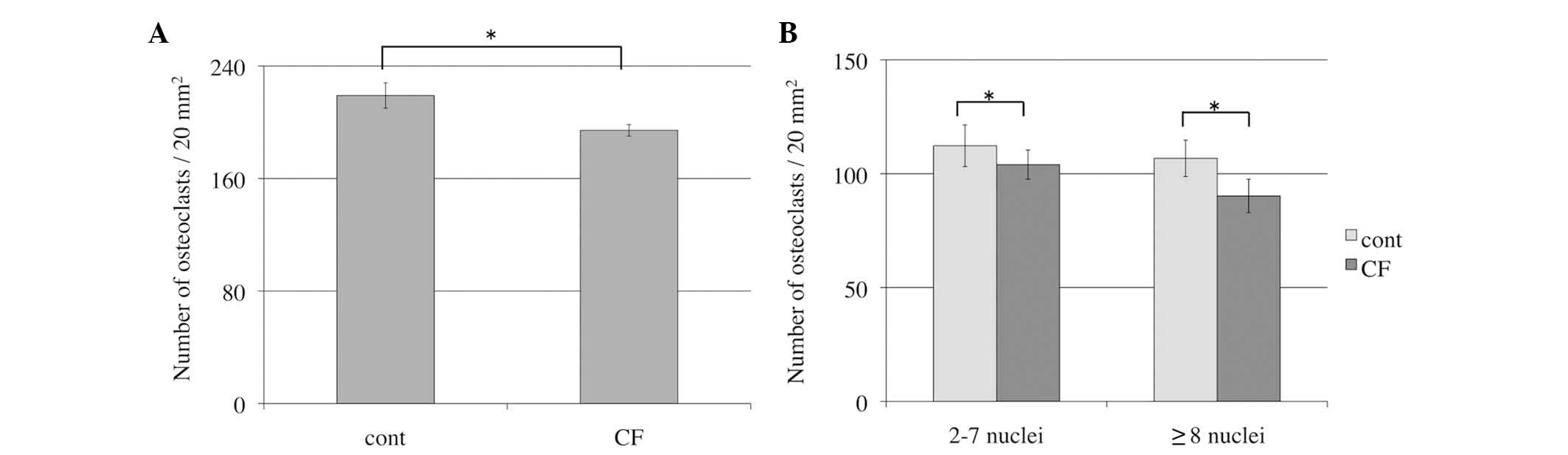
Number of osteoclasts following
release from compressive force
The effect of the release from compressive force on
osteoclasts was also examined. Following release from compressive
force, osteoclast differentiation and fusion were decreased in
comparison with the control groups. The number of osteoclasts
release from compressive force increased 1.7 fold. However, the
number in the control groups increased by 2.4 fold. The total
number of osteoclasts in the control groups was significantly
greater than the cells released from compressive force (P=0.02;
Fig. 3A). The number of small
osteoclasts (2–7 nuclei) and large osteoclasts (≥8 nuclei) in the
control group was increased compared with the cells released from
compressive force (P=0.04 and P=0.03, respectively; Fig. 3B).
Effects of release from compressive
force on the expression of osteoclast differentiation genes
How the expression of osteoclast differentiation
genes was altered by release from compressive force was
investigated using RT-qPCR analysis for osteoclast-specific genes
(NFATc1, TRAP, RANK, MMP-9, Cath-K, ClC7 and ATP6i) and
fusion-associated factors (DC-STAMP and OC-STAMP). Analysis
demonstrated that NFATc1 mRNA levels in the control groups were
increased at 0, 1, 3 and 6 h compared with the cells released from
compressive force (P=0.0003, P=0.003, P=0.0005 and P=0.05,
respectively). However, TRAP, RANK, MMP-9, Cath-K, ClC7, ATP6i,
DC-STAMP, OC-STAMP mRNA levels peaked at 3 h in control groups; and
were significantly increased compared with the cells that had been
released from compressive force (all P<0.01; Fig. 4).
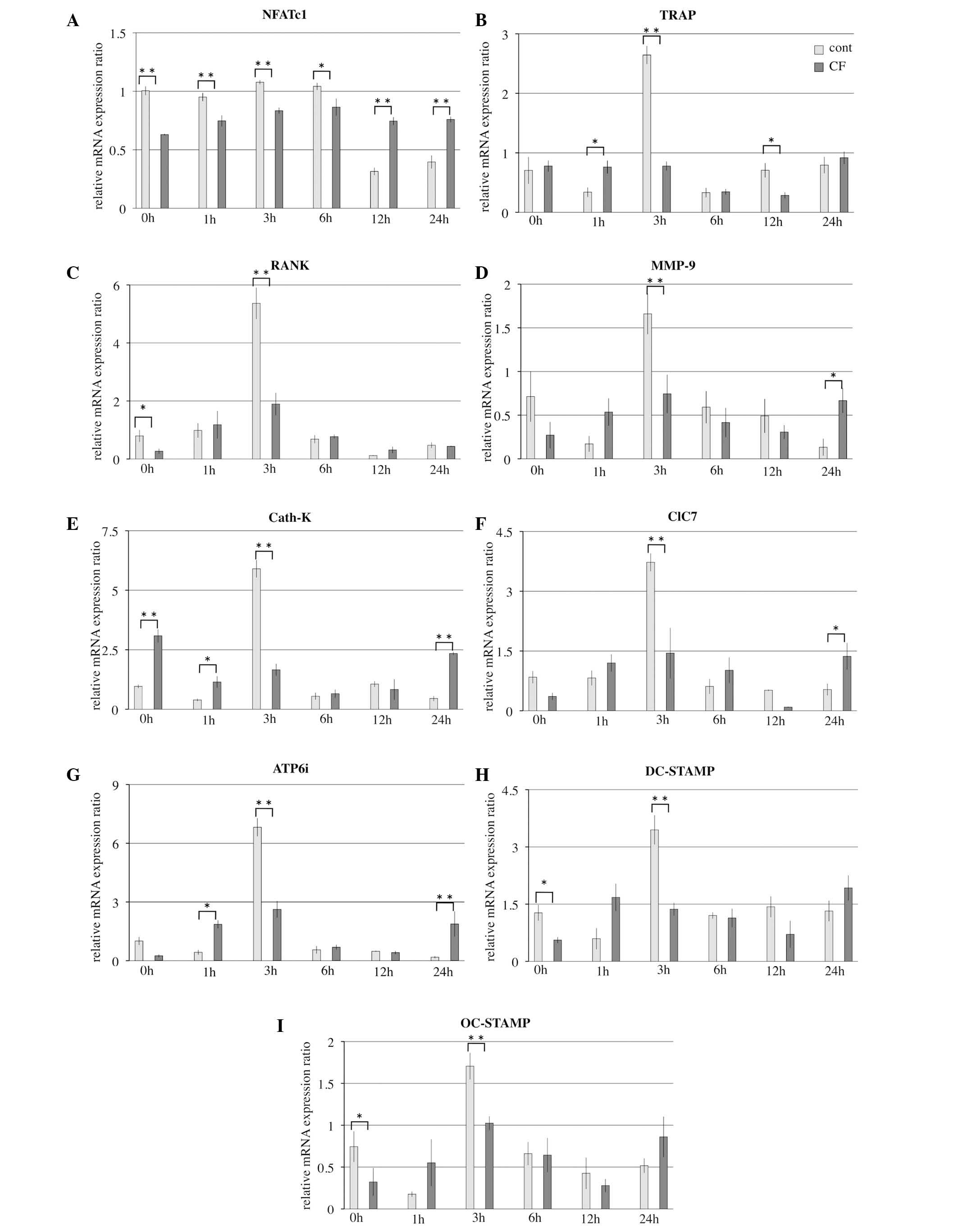 | Figure 4.Effects of release from optimal CF on
mRNA levels of osteoclast-associated genes for 1–24 h. mRNA levels
of osteoclast-associated genes (A) NFATc1 (B) TRAP (C) RANK (D)
MMP-9 (E) Cath-K (F) ClC7 (G) ATP6i (H) DC-STAMP and (I) OC-STAMP
were evaluated by reverse transcription-quantitative polymerase
reaction. Results are presented as the mean ± standard deviation
(n=4). *P<0.05, **P<0.01, comparison indicated by brackets.
cont, control; CF, compressive force; NFATc1, nuclear factor of
activated T cells 1; TRAP, tartrate-resistant acid phosphatase;
RANK, receptor activator of nuclear factor-κB; MMP-9, matrix
metalloproteinase-9; Cath-K, cathepsin-K; ClC7, chloride channel 7;
ATP6i, ATPase H+ transporting vacuolar proton pump
member I; DC-STAMP, dendritic cell-specific transmembrane protein;
OC-STAMP, osteoclast stimulatory transmembrane protein. |
Discussion
Orthodontic tooth movement is achieved by the
remodeling of periodontal ligament and alveolar bone in response to
mechanical pressure and tension (3,4).
During tooth movement, osteoclasts remove bone on the pressure side
and osteoblasts create new bone on the tension side of the tooth.
Following the release of orthodontic force from teeth, orthodontic
relapse occurs. Orthodontic relapse can be defined as the tendency
for teeth to return to their pre-treatment position. This is
considered to be due to gingival fibers and unbalanced lip-tongue
force (27). However, effects of
release from orthodontic force on osteoclasts are unknown.
In a previous study, osteoclasts were subjected to
various types of force in vitro. During orthodontic
treatment, osteoclasts are subjected to compressive force on the
pressure side, and tensile force on the tension side. The number of
osteoclasts was observed to decrease following the application of
tensile force using a Flexercell tension system (9). Osteoclast differentiation was
upregulated subsequent to release from tensile force (8). By contrast, optimal compressive force
induced osteoclast differentiation and fusion (10). Thus, the aim of the present study
was to investigate the effects of release from compressive force on
the pressure side of tooth movement in vitro.
In order to investigate the effects of release from
optimal compressive force on osteoclast differentiation, the number
of TRAP-positive cells was counted and the expression of NFATc1,
TRAP, MMP-9, Cath-K, ClC7, ATP6i, DC-STAMP and OC-STAMP mRNA was
examined.
The results presented in Fig. 1A suggest that osteoclastogenesis
advances rapidly after 4–5 days. Osteoclastogenesis continued to
increase up to day 6, therefore, indicating that the most
appropriate time to apply compressive force was at days 4–5 and to
release compressive force at day 5.
As described above, the optimal compressive force
was defined as 280 mg/cm2, which induced the greatest
increase in osteoclasts in a previous experiment using collagen
gels (10). Optimal compressive
force had been applied to osteoclasts for 24 h; the number of
TRAP-positive multinucleated osteoclasts was increased in the
compressive force cells compared with the control groups. This
suggests that optimal compressive force promoted osteoclast
differentiation and fusion.
Subsequent to the release from optimal compressive
force, the number of TRAP-positive multinucleated osteoclasts were
counted. The number of osteoclasts in the control group was
increased compared with the cells that were released from
compressive force. This suggests that release from optimal
compression suppresses osteoclast differentiation. Thus, how the
expression of osteoclast differentiation genes was altered by
release from compressive force was subsequently investigated using
RT-qPCR analysis.
NFATc1 mRNA levels in the control groups were
increased at 0, 1, 3 and 6 h compared with cells that were release
from compressive force. Thus, NFATc1 mRNA expression was inhibited
for 6 h subsequent to release from compressive force. NFATc1 is the
master switch for osteoclast differentiation (28). The increasing of expression of
NFATc1 lead to expression of other osteoclast-specific genes
increased in the control group. TRAP, RANK, MMP-9, Cath-K, ClC7,
ATP6i, DC-STAMP and OC-STAMP mRNA levels peaked at 3 h in the
control groups and were significantly increased compared with the
cells released from compressive force. Although the NFATc1 mRNA
level in the control group was higher than the compressive force
group at 0 h, these data were consistent with the results of
Hayakawa et al (10) and it
was noted that compressive force affected the expression of NFATc1
at 0 h. However, release from compressive force also affected the
expression of NFATc1 at 1, 3 and 6 h.
The inhibition of the expression levels of TRAP,
RANK, MMP-9, Cath-K, ClC7, ATP6i, DC-STAMP and OC-STAMP mRNA peaked
at 3 h after release from compressive force. TRAP and RANK are
histochemical markers of osteoclasts (29). Inhibition of TRAP and RANK
expression indicates a reduction in the number of osteoclasts.
MMP-9 expression is essential for the migration of osteoclasts
through collagen in the periosteum and developing marrow cavity of
primitive long bones (30,31). Bone resorption may be significantly
reduced by inhibition of MMP-9 (32,33).
Cath-K, ClC7 and ATP6i directly affect bone
resorption within the ruffled border of osteoclasts (34–36).
This expression is associated with bone resorption and was
inhibited following the release from compressive force. In addition
to inhibition of MMP-9 mRNA expression, osteoclast migration and
resorption may be decreased.
DC-STAMP and OC-STAMP modulate cell-cell fusion in
osteoclasts, and are induced by the RANKL-NFATc1 axis (37). Decreased expression of DC-STAMP and
OC-STAMP inhibit the fusion of osteoclasts. Thus, the number of
large osteoclasts was decreased. The current study demonstrated
that optimal compressive force promoted osteoclast differentiation
and fusion; however, release from this compressive force suppressed
osteoclast differentiation and fusion.
The major causes of orthodontic relapse are
considered to be gingival fibers and unbalanced lip-tongue force.
We hypothesize that the suppression of osteoclast differentiation
and fusion following release from optimal compressive force also
has a role in orthodontic relapse. Release from compressive force
suppresses osteoclast differentiation, accelerating bone formation,
whereas, release from tensile force promotes osteoclast
differentiation, accelerating bone resorption (8).
This mechanism may be a factor involved in
orthodontic relapse, however, further research is necessary to
clarify the mechanism of the relapse following active orthodontic
treatment.
Acknowledgements
We are grateful to Dr K. Shibata (Department of
Orthodontics Dentistry, Hokkaido University Graduate School of
Dental Medicine, Sapporo, Japan) for technical advice and support.
This study was supported in part by the Japan Society for the
Promotion of Science Grant-in-Aid for Scientific Research (grant
nos. 25463161, 24659906 and 24593071) and for Young Scientists
(grant no. 2586199403).
References
|
1
|
Karsenty G and Wagner EF: Reaching a
genetic and molecular understanding of skeletal development. Dev
Cell. 2:389–406. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Teitelbaum SL and Ross FP: Genetic
regulation of osteoclast development and function. Nat Rev Genet.
4:638–649. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Roberts-Harry D and Sandy J: Orthodontics.
Part 11: Orthodontic tooth movement. Br Dent J. 196:391–394. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bourauel C, Vollmer D and Jäger A:
Application of bone remodeling theories in the simulation of
orthodontic tooth movements. J Orofac Orthop. 61:266–279. 2000.(In
English, German). View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Boyle WJ, Simonet WS and Lacey DL:
Osteoclast differentiation and activation. Nature. 423:337–342.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Takayanagi H: The role of NFAT in
osteoclast formation. Ann N Y Acad Sci. 1116:227–237. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang F, Wang CL, Koyama Y, Mitsui N,
Shionome C, Sanuki R, Suzuki N, Mayahara K, Shimizu N and Maeno M:
Compressive force stimulates the gene expression of IL-17s and
their receptor in MC3T3-E1 cells. Connect Tissue Res. 51:359–369.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shibata K, Yoshimura Y, Kikuiri T,
Hasegawa T, Taniguchi Y, Deyama Y, Suzuki K and Iida J: Effect of
the release from mechanical stress on osteoclastogenesis in
RAW264.7 cells. Int J Mol Med. 28:73–79. 2011.PubMed/NCBI
|
|
9
|
Kameyama S, Yoshimura Y, Kameyama T,
Kikuiri T, Matsuno M, Deyama Y, Suzuki K and Iida J: Short-term
mechanical stress inhibits osteoclastogenesis via suppression of
DC-STAMP in RAW264.7 cells. Int J Mol Med. 31:292–298.
2013.PubMed/NCBI
|
|
10
|
Hayakawa T, Yoshimura Y, Kikuiri T,
Matsuno M, Fukushima K, Shibata K, Deyama Y, Suzuki K and Iida J:
Optimal compressive force accelerates osteoclastogenesis in
RAW264.7 cells. Mol Med Rep. 12:5879–5885. 2015.PubMed/NCBI
|
|
11
|
Suzuki N, Yoshimura Y, Deyama Y, Suzuki K
and Kitagawa Y: Mechanical stress directly suppresses osteoclast
differentiation in RAW264.7 cells. Int J Mol Med. 21:291–296.
2008.PubMed/NCBI
|
|
12
|
Rubin J, Murphy TC, Fan X, Goldschmidt M
and Talor WR: Activation of extracellular signal-regulated kinase
is involved in mechanical strain inhibition of RANKL expression in
bone stromal cells. J Bone Miner Res. 17:1452–1460. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Koike M, Shimokawa H, Kanno Z, Ohya K and
Soma K: Effects of mechanical strain on proliferation and
differentiation of bone marrow stromal cell line ST2. J Bone Miner
Metab. 23:219–225. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kanzaki H, Chiba M, Sato A, Miyagawa A,
Arai K, Nukatsuka S and Mitani H: Cyclical tensile force on
periodontal ligament cells inhibits osteoclastogenesis through OPG
induction. J Dent Res. 85:457–462. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nishijima Y, Yamaguchi M, Kojima T, Aihara
N, Nakajima R and Kasai K: Levels of RANKL and OPG in gingival
crevicular fluid during orthodontic tooth movement and effect of
compression force on release from periodontal ligament cells in
vitro. Orthod Cranifac Res. 9:63–70. 2006. View Article : Google Scholar
|
|
16
|
Ichimiya H, Takahashi T, Ariyoshi W,
Takano H, Matayoshi T and Nishihara T: Compressive mechanical
stress promotes osteoclast formation through RANKL expression on
synovial cells. Oral Surg Oral Med Oral Pathol Oral Radiol Endod.
103:334–341. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu J, Zhao Z, Zou L, Li J, Wang F, Li X,
Zhang J, Liu Y, Chen S, Zhi M and Wang J: Pressure-loaded MSCs
during early osteodifferentiation promote osteoclastogenesis by
increase of RANKL/OPG ratio. Ann Biomed Eng. 37:794–802. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rubin J, Biskobing D, Fan X, Rubin C,
McLeod K and Taylor WR: Pressure regulates osteoclast formation and
MCSF expression in marrow culture. J Cell Physiol. 170:81–87. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mehrotra M, Saegusa M, Wadhwa S,
Voznesensky O, Peterson D and Pilbeam C: Fluid flow induceds RANKL
expression in primary murine calvarial osteoblasts. J Cell Biochem.
98:1271–1283. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tan SD, Vries TJ, Kujipers-Jagtman AM,
Semeins CM, Everts V and Klein-Nulend J: Osteocytes subjected to
fluid flow inhibits osteoclast formation and formation and bone
resorption. Bone. 41:745–751. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hong Z Qing, Tao L Meng, Yi Z, Wei L, Ju
Xiang S and Li L: The effect of rotative stress on CAII, FAS, FASL,
OSCAR, and TRAP gene expression in osteoclasts. J Cell Biochem.
114:388–397. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Makihira S, Kawahara Y, Yuge L, Mine Y and
Nikawa H: Impact of the microgravity environment in a 3-dimentional
clinostat on osteoblast- and osteoclast-like cells. Cell Biol Int.
32:1176–1181. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kadow-Romacker A, Haffman JE, Duga G,
Wildemann B and Schmidmaier G: Effect of mechanical stimulation on
osteoblast- and osteoclast-like cells in vitro. Cell Tissues
Organs. 190:61–68. 2009. View Article : Google Scholar
|
|
24
|
Takeyama S, Yoshimura Y, Deyama Y,
Sugawara Y, Fukuda H and Matsumoto A: Phosphate decreases
osteoclastogenesis in coculture of osteoblast and bone marrow.
Biochem Biophys Res Commum. 282:798–802. 2001. View Article : Google Scholar
|
|
25
|
Nakamura K, Deyama Y, Yoshimura Y, Suzuki
K and Morita M: Toll-like receptor 3 ligand-induced antiviral
response in mouse osteoblastic cells. Int J Mol Med. 19:771–775.
2007.PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Reitan K: Tissue behavior during
orthodontic tooth movement. Am J Orthod. 46:881–900. 1960.
View Article : Google Scholar
|
|
28
|
Zhao Q, Wang X, Liu Y, He A and Jia R:
NFATc1: Functions in osteoclasts. Int J Biochem Cell Biol.
42:576–579. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nakagawa N, Kinosaki M, Yamaguchi K, Shima
N, Yasuda H, Yano K, Morinaga T and Higashio K: RANK is essential
signaling receptor for osteoclast differentiation factor in
osteoclastogenesis. Biochem Biophys Res Commun. 253:395–400. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Blavier L and Delaissé JM: Matrix
metalloproteinases are obligatory for the migration of
preosteoclasts to the developing marrow cavity of primitive long
bones. J Cell Sci. 108:3649–3659. 1995.PubMed/NCBI
|
|
31
|
Sato T, Foged NT and Delaissé JM: The
migration of purified osteoclasts through collagen is inhibited by
matrix metalloproteinase inhibitors. J Bone Miner Res. 13:59–66.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hill PA, Murphy G, Docherty AJ, Hembry RM,
Millican TA, Reynolds JJ and Meikle MC: The effects of selective
inhibitors of matrix metalloproteinase (MMPs) on bone resorption
and the identification of MMPs and TIMP-1 in isolated osteoclasts.
J Cell Sci. 107:3055–3064. 1994.PubMed/NCBI
|
|
33
|
Spessotto P, Rossi FM, Degan M, Di Francia
R, Perris R, Colombatti A and Gattei V: Hyaluronan-CD44 interaction
hampers migration of osteoclast-like cells by down-regulation
MMP-9. J Cell Biol. 158:1133–1144. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Saftig P, Hunziker E, Wehmeyer O, Jones S,
Boyde A, Rommerskirch W, Moritz JD, Schu P and von Figura V:
Impaired osteoclastic bone resorption leads to osteopeterosis in
cathepsin-K-deficient mice. Proc Natl Acad Sci USA. 95:13453–13458.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kornak U, Kasper D, Bösl MR, Kaiser E,
Schweizer M, Schulz A, Friedrich W, Delling G and Jentsch TJ: Loss
of the ClC-7 chloride channel leads to osteopetrosis in mice and
man. Cell. 104:205–215. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li YP, Chen W, Liang Y, Li E and Stashenko
P: Atp6i-deficient mice exhibit severe osteopetrosis due to loss of
osteoclast-mediated extracellular acidfiction. Nat Genet.
23:447–451. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
37
|
Miyamoto H, Suzuki T, Miyauchi Y, Iwasaki
R, Kobayashi T, Sato Y, Miyamoto K, Hoshi H, Hashimoto K, Yoshida
S, et al: Osteiclasts stimulatory transmembrane protein and
dendritic cell-specific transmembrane protein cooperatively
modulate cell-cell fusion to form osteoclasts and foreign body
giant cells. J Bone Miner Res. 27:1289–1297. 2012. View Article : Google Scholar : PubMed/NCBI
|