Introduction
Acute promyelocytic leukemia (APL) is a subtype of
acute myeloid leukemia (AML), accounting for 10–15% of all cases of
AML (1). APL is characterized by
the selective expansion of immature hematopoietic precursors
inhibited at the promyelocytic stage (1). Severe bleeding associated with
disseminated intravascular coagulation and other complications can
lead to early mortality rates in patients with APL (2). APL is characterized by the t (15;17)
translocation, which fuses the retinoic acid receptor α (RARα) gene
to the promyelocytic leukemia (PML) gene on chromosome 15, leading
to the production of the PML/RARα fusion protein. So far, the
majority of investigations have focused on the PML/RARα fusion
protein as a whole, which interferes with the function of wild-type
RARα and PML, and suppresses the differentiation of promyelocytic
cells (3,4); however, the specific underlying
mechanism remains to be fully elucidated. A study by Lane and Ley
(5) found that the PML/RARα fusion
protein can be cleaved by neutrophil elastase (NE), and APL cells
derived from human patients contain this PML-RARα cleaving
activity. This cleavage produces the novel protein, NLS-RARα. A
previous study showed that the expression of PML/RARα is associated
with cellular toxicity, which is dependent on the expression of NE
in early myeloid cells (6).
In addition to the PML/RARα fusion gene, a number of
other gene translocations have been reported in rare cases of APL.
These fusion genes all have the same RARα protein as a fusion
partner, and all the resulting fusion proteins are fused with RARα
B through F domains (7). NLS-RARα
also has these domains (5),
therefore, the present study hypothesized that NLS-RARα may be
important in initiating APL. Complete remission can be achieved in
90% of patients with APL treated with a combination of arsenic
trioxide and all-trans retinoic acid (ATRA) (8). However, recurrence and resistance to
ATRA treatment occurs in ~20% of patients in complete remission. In
addition, 10–30% of patients with APL are resistant to ATRA
(9,10), and treatment with arsenic trioxide
may cause cardiac toxicity, hepatic toxicity and neurotoxicity
(11). Although the remission rate
is higher, compared with those of other types of malignant tumor,
the early mortality rate remains high (12). Therefore, it is important to
further probe the molecular mechanisms underlying APL, and identify
a novel target for its diagnosis and treatment.
In the present study, the role of NLS-RARα in NB4
cells was examined using ATRA to induce differentiation.
Furthermore, the potential molecular mechanisms modulated by
NLS-RARα in leukemogenesis were examined. The data revealed that
the overexpression of NLS-RARα promoted cell proliferation and
inhibited differentiation via the PI3K/AKT signaling pathway. The
results indicated that NLS-RARα had a marked effect on APL
pathogenesis and may be a novel target for the therapeutic
treatment of APL.
Materials and methods
Cell lines and cell culture
Human acute promyelocytic leukemia NB4 and 293T
cells were purchased from the Institute of Biochemistry and Cell
Biology, Chinese Academy of Sciences (Shanghai, China). The cells
were cultured in RPMI-1640 medium with 10% fetal bovine serum (all
purchased from Invitrogen Life Technologies; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) at 37°C in 5%
CO2.
Construction and packing of
lentiviruses
The NLS-RARα and RARα genes were amplified by
polymerase chain reaction (PCR) using pCMV-HA-NLS-RARα and
pCMV-Myc-RARα plasmids as templates, respectively. The PCR product
containing the sequence of the target gene was subcloned into the
shuttle plasmid, LV5 (GenePharma Co., Ltd., Shanghai, China),
between the NotI and BamHI restriction sites, whereas
the LV5 plasmid with no target gene insert was selected as a
negative control (NC). The LV5 shuttle plasmid contained the green
fluorescent protein (GFP) gene. The four plasmids, comprising the
packaging plasmids pGag/Pol, pRev, pVSV-G and the recombinant
shuttle plasmid (GenePharma Co., Ltd., Shanghai, China), were
transduced into 293T cells (5×106) using Lipofectamine™
2000 (Invitrogen Life Technologies; Thermo Fisher Scientific, Inc.)
for 48 h. The viral supernatant was later centrifuged at 6,000 ×
g for 2 h at 4°C, then concentrated by 0.45 µm filter and
the titer determined.
Generation of stably transfected cell
lines
NB4 cells were subcultured into a 24-well culture
plate at a density of ~5×104 cells/well. Each well was
supplemented with 5 µg polybrene (Sigma-Aldrich; Merck Millipore,
Darmstadt, Germany), and the cells were infected with the
LV5-NLS-RARα lentivirus (LV-NLS-RARα group), LV5-RARα lentivirus
(LV-RARα group) or negative control LV5 lentivirus (LV-NC group),
respectively, at a multiplicity of infection of 150. After 24 h
incubated at 37°C in 5% CO2, the complete medium was
replaced without polybrene. A fluorescence microscope was used to
observe the expression of GFP. At 72 h post-infection, stably
transfected cell lines were selected using puromycin
(Sigma-Aldrich; Merck Millipore) at a dose of 1 µg/ml for 7 days to
establish the NLS-RARα, RARα and NC cell lines. The NB4 cell line
was selected as a blank control (control group).
Cell viability assay
The viabilities of the NB4 cells infected with the
lentiviruses were detected using a Cell Counting Kit-8 (CCK-8)
assay (7SeaPharmTech, Shanghai, China) within 5 days at different
incubation time points (0, 24, 48, 72 and 96 h). The absorbance was
measured at 450 nm using a microplate reader.
Cell cycle assay
To examine the cell cycle, 1×106 cells in
each group were washed twice with PBS and were fixed with pre-cold
75% ethanol for 2 h at 4°C. Cell cycle was examined using flow
cytometry (Beckman Coulter, Brea, CA, USA) at 488 nm. Data were
analyzed using the Multicycle DNA content and cell analysis
software program AV (Phoenix Flow System, Inc., San Diego, CA,
USA)
Cell differentiation assay
For detection of the cell differentiation antigen,
CD11b, ATRA (Sigma-Aldrich; Merck Millipore) was used to induce
cell differentiation at a concentration of 1 nM for 3 days. The
cells (1×106/group) were washed twice with PBS and
incubated with phycoerythrin (PE)-conjugated CD11b antibody (cat.
no. 12-0113-42; eBioscience, Inc., San Diego, CA, USA) at 4°C for
30 min in the dark. The cells were then analyzed using flow
cytometry (BD FACS Vantage; BD Biosciences, San Jose, CA, USA) and
CellQuest Pro software version 5.1 (BD Pharmingen, San Diego, CA,
USA).
Western blot analysis
The cells were lysed on ice in RIPA lysis buffer
(Beyotime Institute of Biotechnology, Jiangsu, China) with protease
inhibitor (PMSF) and phosphatase inhibitors (NaF and
Na3VO4; Roche Diagnostics, Basel,
Switzerland). A BCA protein assay kit (Beyotime Institute of
Biotechnology) was used to measure protein concentration. A loading
buffer (5X) was added to all protein solutions, which were then
boiled for 5 min. Equal quantities (50 µm) of proteins were
separated by 10% SDS-PAGE and then transferred onto PVDF membranes
(EMD Millipore, Billerica, MA, USA). The PVDF membranes were then
blocked with 0.1% Tween-20 in TBS containing 5% nonfat dry milk for
2 h at room temperature. The membranes were incubated with primary
antibodies against AKT (cat. no. ab32505; 1:1,000; Abcam,
Cambridge, UK), glycogen synthase kinase 3β (GSK3β; cat. no.
12456), c-myc (cat. no. 5605), cyclin D1 (cat. no. 2978; 1:1,000;
Cell Signaling Technology, Inc., Danvers, MA, USA), and β-actin
(cat. no. BM0627; 1:1,000, Boster Biological Technology Ltd.,
Wuhan, China) overnight at 4°C. The membranes were washed three
times with TBST and incubated with horseradish peroxidase
(HRP)-conjugated secondary antibodies (cat. nos. ZDR-5307 and
ZDR-5306; ZSGB-Bio, Beijing, China) for 1 h at room temperature.
Protein bands were visualized by incubating the membranes in
chemiluminescent HRP substrate were and analyzed using Adobe
Photoshop 12.0 software (Adobe Systems, Inc., San Jose, CA,
USA).
Statistical analysis
SPSS 17.0 software (SPSS, Inc., Chicago, IL, USA)
was used for statistical analysis. The results are represented as
the mean ± standard deviation (n≥3) and were compared using one-way
analysis of variance. P<0.05 was considered to indicate a
statistically significant difference.
Results
NB4 cell transduction induces the
expression of NLS-RARα
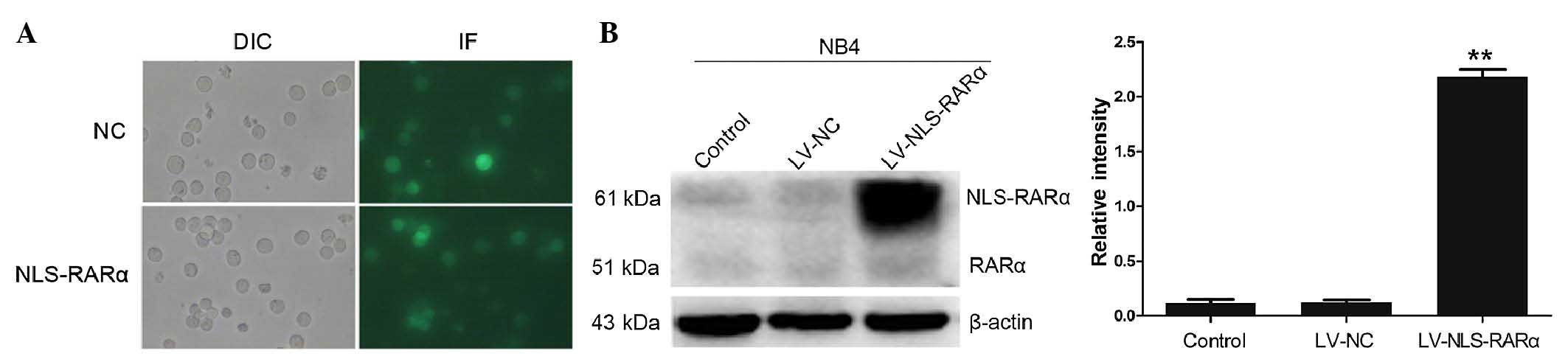
The fluorescence of the NB4 cells showed that ~90%
of the cells were transduced by the lentiviral vectors (Fig. 1A). To establish a stable NLS-RARα
cell line, puromycin was used to screen the infected cells. Western
blot analysis, with β-actin as an internal reference, showed that
the NLS-RARα/β-actin intensity ratio was 2.18±0.11, which indicated
that the NLS-RARα gene was efficiently expressed in the LV-NLS-RAR
group (Fig. 1B).
Overexpression of NLS-RARα promotes
the proliferation of NB4 cells
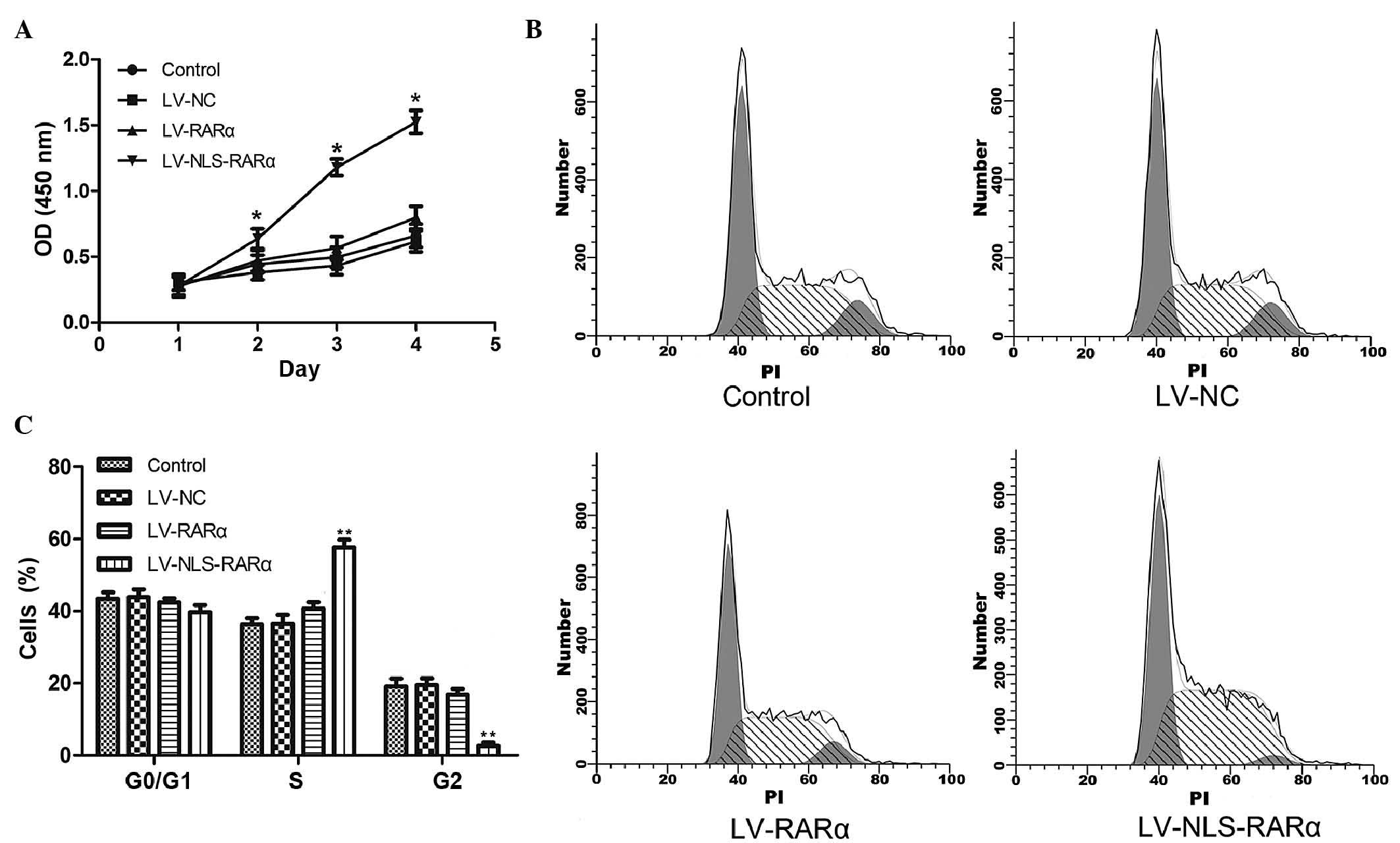
The proliferative abilities of the cells were
examined using a CCK-8 assay. As shown in Fig. 2A, the overexpression of the
NLS-RARα protein promoted the proliferation of the NB4 APL cells
(*P<0.05). No statistically significant difference were found
between the blank control, negative control and LV-RARα groups.
Therefore, the overexpression of NLS-RARα specifically induced the
proliferation of the APL cells.
To determine whether the NLS-RARα protein affected
the cell cycle of the NB4 cells, flow cytometry was used to assess
the cell cycle distribution (Fig.
2B). The results showed that the overexpression of NLS-RARα led
to a significant increase (57.63±1.17%) in the number of cells in
the S-phase and a decrease (2.63±0.91%) in the G2-phase of the cell
cycle suggesting that NLS-RARα protein promoted cell proliferation
by enhancing cell cycle progression (Fig. 2C).
Overexpression of NLS-RARα inhibits
the differentiation of NB4 cells
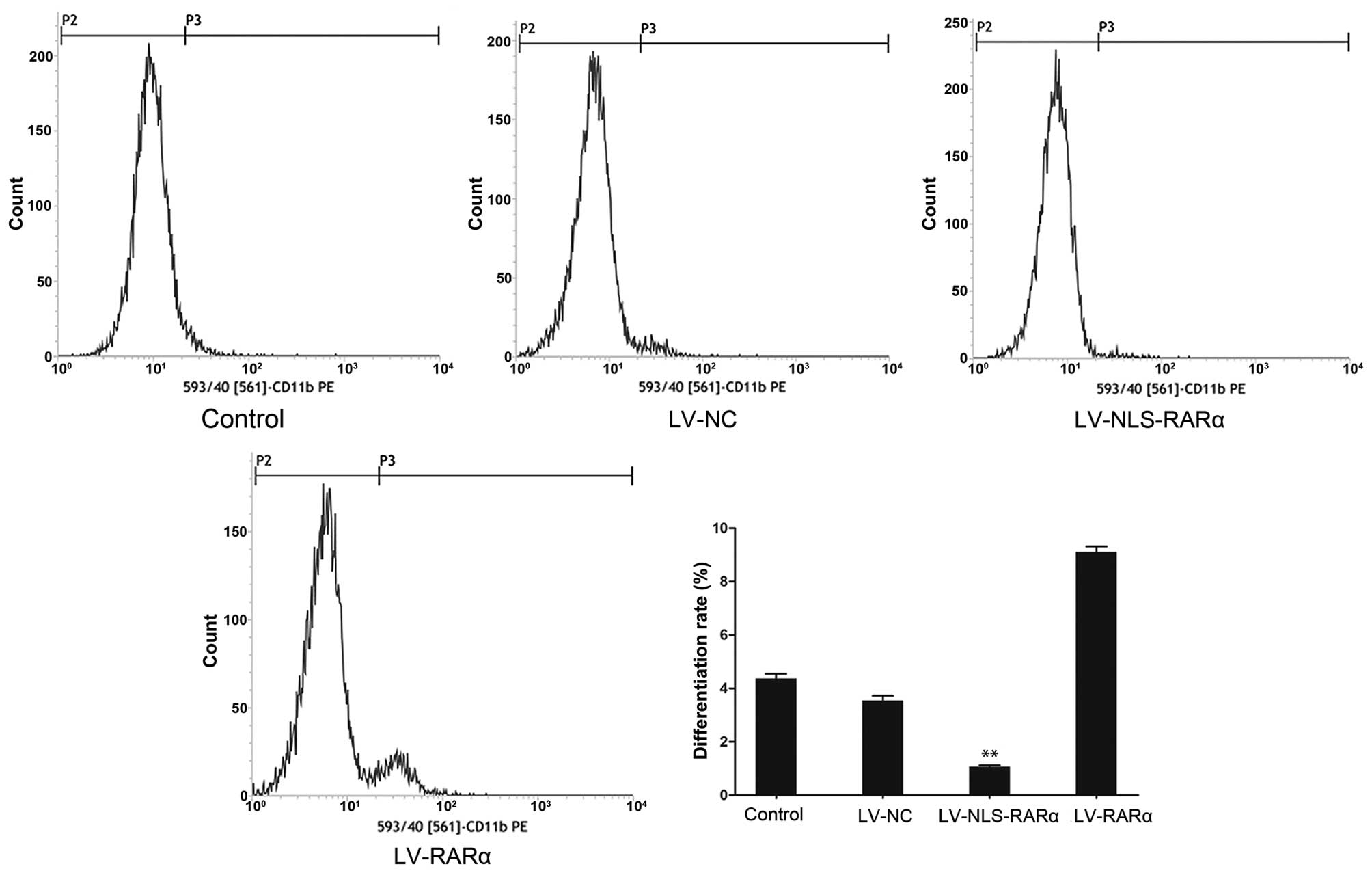
Flow cytometric analysis for the determination of
NB4 cell differentiation was based on the detection of
PE-conjugated CD11b antibody fluorescence. The percentage of total
differentiated cells was significantly decreased (0.98±0.06%) in
the LV-NLS-RARα group, compared with the blank control
(4.56±0.15%), negative control (3.48±0.11%) and LV-RARα groups
(9.49±0.19%), as shown in Fig. 3.
The analysis showed that the NLS-RARα protein inhibited the
ATRA-induced cell differentiation.
Overexpression of NLS-RARα activates
the PI3K/AKT pathway
To further define the molecular mechanisms modulated
by NLS-RARα in the proliferation and differentiation of NB4 cells,
the protein expression of proliferation- and
differentiation-associated genes in the NLS-RARα-overexpressing
cells were detected. It was found that the protein expression of
c-myc and the phosphorylation of AKT (Ser473) and GSK3β (Ser9)
increased, although no difference was found in the protein
expression of cyclin D1. The cells transduced with LV-NLS-RARα,
which were pretreated with 20 µM of the PI3K inhibitor, LY294002,
showed that it significantly inhibited the phosphorylation of AKT
(Ser473) and its downstream target, GSK3β (Ser9). These results
indicated that the NLS-RARα protein exerted proliferative and
anti-differentiation effects in the APL cells, at least partly
through the PI3K/AKT pathway (Fig.
4).
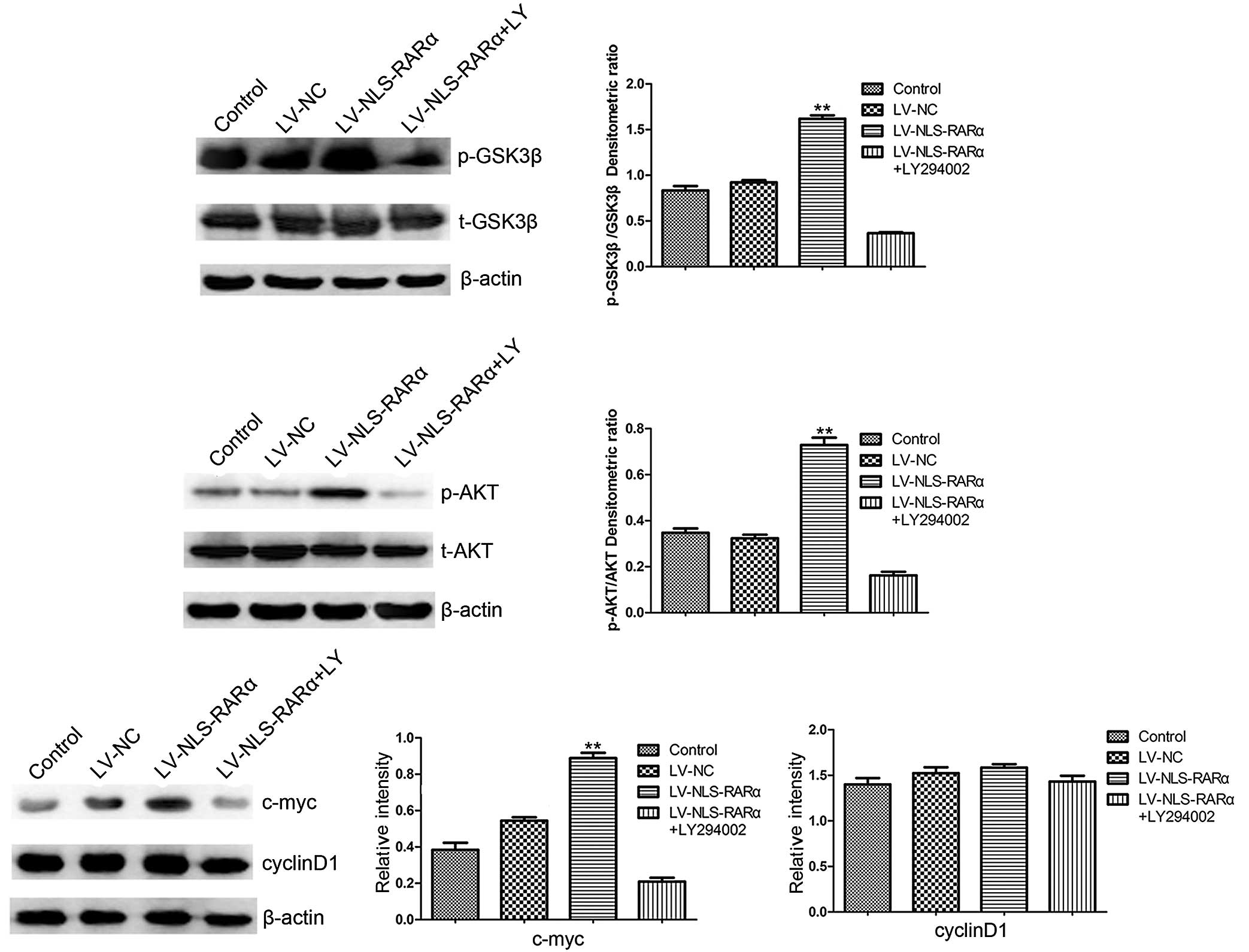 | Figure 4.Western blot analysis detection of the
expression of p-AKT and p-GSK3β, c-myc and cyclin D1. Each group of
cells were lysed and analyzed using western blot analysis. The
LV-NLS-RARα group was pretreated with 20 µM PI3K inhibitor
(LY294002) for 24 h. Date are presented as the mean ± standard
deviation (n=3) and analyzed using one-way analysis of variance.
p-AKT, p-GSK3β, and c-myc: **P<0.01, LV-NLS-RARα group, vs.
control, LV-NC or LV-NLS-RARα+LY294002 group. Cyclin D1: P>0.05
LV-NLS-RARα group, vs. control, LV-NC or LV-NLS-RARα+LY294002
group. NLS-RARα, nuclear localization signal-retinoic acid receptor
α; NC, negative control; LV, lentivirus; p-, phosphorylated. |
Discussion
Previous studies have shown that NE is important for
the ability of PML-RARα to initiate APL in transgenic mice models
of this disease. Primary mouse and human APL cells also exhibit
NE-dependent PML-RARα cleaving activity (5). In the present study, to further
investigate the functional role of NLS-RARα, the overexpression of
NLS-RARα was induced in APL NB4 cells through the process of
lentiviral transfection. The results showed that NLS-RARα promoted
cell proliferation, and caused a significant increase in S-phase
cells and decrease in G2-phase cells, thus promoting cell cycle
progression. Human wild-type RARα is a ligand-dependent
transcription factor, which requires combination with retinoid X
receptor α (RXRα) to respond to ATRA. The RARα/RXRα heterodimer
binds to specific DNA sequences of the retinoic acid response
element in the promoter regions of target genes and regulates the
transcription of these genes (13). RARα target genes, including p21,
RARβ, transglutaminase II and homeobox genes, are important in the
regulation of cell proliferation and differentiation (4). PML-RARα can compete with wild-type
RARα for RXRα and interfere in the regulation of transcription
(14). The function of NLS-RARα
may be similar to PML-RARα, which interferes with RARα.
Our previous experiments showed that NLS-RARα, which
has PML nuclear localization signal, shows increased distribution
in the nucleus, compared with wild-type RARα (15). Several studies have found that
alterations in protein localization can affect cell function
(16,17). The occurrence of tumors may be
associated with the abnormal localization of proteins; therefore,
it was hypothesized that alterations in protein location were also
implicated in the pathogenesis of APL. The CD11b myeloid
differentiation antigen is a marker of mature myeloid cell
differentiation (18). The results
of the present study confirmed that, following ATRA treatment to
induce cell differentiation, the expression of CD11b was reduced
significantly in the LV-NLS-RARα group, compared with that in the
control group, demonstrating that the overexpression of NLS-RARα
inhibited cell maturation.
To determine the effects of NLS-RARα on
proliferation and differentiation, the present study focused on the
AKT-GSK3β-c-myc-cyclin D1 pathway. Previous studies have shown that
AKT is activated abnormally in several types of cancer, including
leukemia (19–21). The PI3K/AKT pathway coordinately
regulates diverse cellular programs, including proliferation,
differentiation and apoptosis. In the present study, it was
demonstrated that the over-expression of NLS-RARα increased the AKT
serine/threonine phosphorylation and caused upregulation of the
downstream proteins, pGSK and c-myc. As an important oncogene,
constitutively expressing a high level of c-myc leads to a reduced
growth factor requirement, shorter duration in the G1 phase and an
increased growth rate. Furthermore, c-myc can inhibit
differentiation by preventing cells from leaving the cell cycle
(22). AKT phosphorylates numerous
substrates to modify their function (19), including GSK-3β associated with the
regulation of cell proliferation and differentiation (23). Additionally, GSK-3β has been shown
to negatively regulate the stability and expression of the
cell-cycle regulator, cyclin D1 (24). Cyclin D1 is an important downstream
of AKT, which can promote G1-S progression associated with cell
growth (25). Previous studies
have showed that PI3K/AKT activity is also critical in cellular
differentiation processes (26,27);
it negatively regulates granulocyte differentiation. ATRA, through
deactivation of the PI3K/AKT signaling pathways, induces the
differentiation of APL cells (28). Previously, Srinivas et al
(29) demonstrated that RARα
interacts with AKT through the DNA binding domain, which is also
present in NLS-RARα. Our previous study showed that NLS-RARα
interacts with AKT, which may be one of the AKT pathway-activating
factors (30). Thus, it is
essential to further determine whether their direct interaction
activates the AKT pathway.
In conclusion, the data obtained in the present
study demonstrated that the overexpression of NLS-RARα promoted the
proliferation of APL cells and inhibited their differentiation via
the PI3K/AKT signaling pathway. These results provide novel insight
into the mechanisms of leukemogenesis. Future investigations aim to
specifically target NLS-RARα as a therapeutic option for diagnosis
and treatment response.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant no. 81171658) and the Natural
Science Foundation Project of CQ CSTC (grant no. 2011BA5037).
References
|
1
|
Warrell RP Jr, de Thé H, Wang ZY and Degos
L: Acute promyelocytic leukemia. N Engl J Med. 329:177–189. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lo-Coco F and Cicconi L: History of acute
promyelocytic leukemia: A tale of endless revolution. Mediterr J
Hematol Infect Dis. 3:e20110672011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Grignani F, Ferrucci PF, Testa U, Talamo
G, Fagioli M, Alcalay M, Mencarelli A, Grignani F, Peschle C,
Nicoletti I, et al: The acute promyelocytic leukemia-specific
PML-RAR alpha fusion protein inhibits differentiation and promotes
survival of myeloid precursor cells. Cell. 74:423–431. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Laurenzana A, Pettersson F and Miller WH
Jr: Role of PML-RARα in the pathogenesis of APL. Drug Discov Today
Dis Mech. 3:499–505. 2006. View Article : Google Scholar
|
|
5
|
Lane AA and Ley TJ: Neutrophil elastase
cleaves PML-RARalpha and is important for the development of acute
promyelocytic leukemia in mice. Cell. 115:305–318. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lane AA and Ley TJ: Neutrophil elastase is
important for PML-retinoic acid receptor alpha activities in early
myeloid cells. Mol Cell Biol. 25:23–33. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
di Masi A, Leboffe L, De Marinis E, Pagano
F, Cicconi L, Rochette-Egly C, Lo-Coco F, Ascenzi P and Nervi C:
Retinoic acid receptors: From molecular mechanisms to cancer
therapy. Mol Aspects Med. 41:1–115. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang ZY and Chen Z: Acute promyelocytic
leukemia: From highly fatal to highly curable. Blood.
111:2505–2515. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang ME, Ye YC, Chen SR, Chai JR, Lu JX,
Zhao L, Gu LJ and Wang ZY: Use of all-trans retinoic acid in the
treatment of acute promyelocytic leukemia. Blood. 72:567–572.
1988.PubMed/NCBI
|
|
10
|
Breccia M, Cicconi L and Lo-Coco F: ATRA+
ATO: Has a new standard of care been established in low-risk acute
promyelocytic leukaemia? Curr Opin Hematol. 21:95–101. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Iland HJ and Seymour JF: Role of arsenic
trioxide in acute promyelocytic leukemia. Curr Treat Options Oncol.
14:170–184. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Park JH, Qiao B, Panageas KS, Schymura MJ,
Jurcic JG, Rosenblat TL, Altman JK, Douer D, Rowe JM and Tallman
MS: Early death rate in acute promyelocytic leukemia remains high
despite all-trans retinoic acid. Blood. 118:1248–1254. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
De Braekeleer E, Douet-Guilbert N and De
Braekeleer M: RARA fusion genes in acute promyelocytic leukemia: A
review. Expert Rev Hematol. 7:347–357. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Soprano DR, Qin P and Soprano KJ: Retinoic
acid receptors and cancers. Annu Rev Nutr. 24:201–221. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang H, Zhong L, Jiang KL, Zhu XY, Ma PP,
Yang XQ and Liu BZ: Location verification of NLS-RARα protein in
infected NB4 cell line with adenovirus Ad-NE. Chin J Cell Biol.
36:331–337. 2014.(In Chinese).
|
|
16
|
Marshall KS, Cohen MJ, Fonseca GJ,
Todorovic B, King CR, Yousef AF, Zhang Z and Mymryk JS:
Identification and characterization of multiple conserved nuclear
localization signals within adenovirus E1A. Virology.
454-455:206–214. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun Z, Wu T, Zhao F, Lau A, Birch CM and
Zhang DD: KPNA6 (Importin {alpha}7)-mediated nuclear import of
Keap1 represses the Nrf2-dependent antioxidant response. Mol Cell
Biol. 31:1800–1811. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rizzatti EG, Garcia AB, Portieres FL,
Silva DE, Martins SL and Falcão RP: Expression of CD117 and CD11b
in bone marrow can differentiate acute promyelocytic leukemia from
recovering benign myeloid proliferation. Am J Clin Pathol.
118:31–37. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vivanco I and Sawyers CL: The
phosphatidylinositol 3-kinase-AKT pathway in human cancer. Nat Rev
Cancer. 2:489–501. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cumberbatch M, Tang X, Beran G, Eckersley
S, Wang X, Ellston RP, Dearden S, Cosulich S, Smith PD, Behrens C,
et al: Identification of a subset of human non-small cell lung
cancer patients with high PI3Kβ and low PTEN expression, more
prevalent in squamous cell carcinoma. Clin Cancer Res. 20:595–603.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xie X, Tang B, Zhou J, Gao Q and Zhang P:
Inhibition of the PI3K/Akt pathway increases the chemosensitivity
of gastric cancer to vincristine. Oncol Rep. 30:773–782.
2013.PubMed/NCBI
|
|
22
|
Henriksson M and Lüscher B: Proteins of
the Myc network: Essential regulators of cell growth and
differentiation. Adv Cancer Res. 68:109–182. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wray J, Kalkan T, Gomez-Lopez S, Eckardt
D, Cook A, Kemler R and Smith A: Inhibition of glycogen synthase
kinase-3 alleviates Tcf3 repression of the pluripotency network and
increases embryonic stem cell resistance to differentiation. Nat
Cell Biol. 13:838–845. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ougolkov AV and Billadeau DD: Targeting
GSK-3: A promising approach for cancer therapy? Future Oncol.
2:91–100. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Diehl JA, Cheng M, Roussel MF and Sherr
CJ: Glycogen synthase kinase-3beta regulates cyclin D1 proteolysis
and subcellular localization. Gene Dev. 12:3499–3511. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qiao J, Paul P, Lee S, Qiao L, Josifi E,
Tiao JR and Chung DH: PI3K/AKT and ERK regulate retinoic
acid-induced neuroblastoma cellular differentiation. Biochem Biophs
Res Commun. 424:421–426. 2012. View Article : Google Scholar
|
|
27
|
Kim JM, Yoon M, Kim J and Kim SS, Kang I,
Ha J and Kim SS: Phosphatidylinositol 3-kinase regulates
differentiation of H9c2 cardiomyoblasts mainly through the protein
kinase B/Akt-independent pathway. Arch Biochem Biophys. 367:67–73.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ozpolat B, Akar U, Steiner M,
Zorrilla-Calancha I, Tirado-Gomez M, Colburn N, Danilenko M,
Kornblau S and Berestein GL: Programmed cell death-4 tumor
suppressor protein contributes to retinoic acid-induced terminal
granulocytic differentiation of human myeloid leukemia cells. Mol
Cancer Res. 5:95–108. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Srinivas H, Xia D, Moore NL, Uray IP, Kim
H, Ma L, Weigel NL, Brown PH and Kurie JM: Akt phosphorylates and
suppresses the transactivation of retinoic acid receptor alpha.
Biochem J. 395:653–662. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jiang K, Yang X, Ma P, Wang H, Zhu X, Liu
B and Zhong L: NLS-RARα: A novel transcriptional factor. Oncol
Lett. (In press).
|