Introduction
Stroke, a life-threatening condition, is the second
leading cause of death worldwide (1). With the advancement of therapeutic
strategies, the relative rate of mortality from stroke has
declined. However, it still carries a considerable emotional and
economic burden. Brain repair following stroke requires various
processes, including neurogenesis, angiogenesis and synaptogenesis
in the areas around the infarction (2–4). The
formation of new blood vessels by angiogenesis is a key feature of
stroke. Previously, a multitude of essential angiogenic inducers,
including angiopoietin-2 (Ang-2), vascular endothelial growth
factor (VEGF), fatty acid binding protein-4, glucose-6-phosphate
isomerase and neuropilin-1, were reported to promote
neovascularization following stroke (5–7).
Leucine-rich α2-glycoprotein 1 (LRG1) was initially
identified as a secreted glycoprotein in human serum by Haupt and
Baudner (8) in 1977. It consists
of a single polypeptide chain, which includes two intrachain
disulfide bonds with 312 amino acid residues (9). LRG1 is a highly conserved member of
leucine-rich repeat (LRR) family of proteins, several of which are
involved in protein-protein interactions, signaling transduction,
cell adhesion, cell survival, cell apoptosis, cell metastasis, DNA
transcription, DNA repair and RNA processing (10–13).
Zhang et al (14) reported
that LRG1 was involved in the inhibition of hepatocellular
carcinoma (HCC) metastasis and may function as a novel metastasis
suppressor in HCC. Additionally, the function of LRG1 in tumors was
investigated, and indicated that LRG1 was associated with multiple
types of tumor, including ovarian cancer, glioblastoma and squamous
cell lung cancer (15–18). Furthermore, certain previous
studies reported that LRG1 is involved in various diseases,
including inflammatory diseases, heart failure, arterial stiffness,
endothelial dysfunction and peripheral arterial disease (19–23).
Previously, LRG1 expression in the brain during aging and
neurodegenerative diseases, including idiopathic normal pressure
hydrocephalus, Parkinson's disease with dementia, dementia with
Lewy bodies, progressive nuclear palsy and Alzheimer's disease was
investigated (24,25). LRG1 was also considered as a novel
biomarker of neurodegenerative disease in human cerebrospinal fluid
and may cause neurodegeneration in mouse cerebral cortex (24,25).
Wang et al (26) reported that, as a novel angiogenic
factor, LRG1 was significantly elevated in models of retinal
neovascularization and expression was restricted almost exclusively
to the vasculature system. Increased LRG1 was also observed in a
mouse model of choroidal neovascularization compared with control
groups, whereas antibody blockade of LRG1 reduced lesion sizes
during the progression of choroidal neovascularization (26). The results suggested that LRG1 was
a potential therapeutic target for the treatment of macular
degeneration (26). Furthermore,
the investigation demonstrated that LRG1 promoted
neovascularization through the transforming growth factor (TGF)-β1
signaling pathway in pathological blood vessel formation (26). In addition, LRG1 overexpression
induced cell migration and bound to TGF-β1 in endothelial venules
(27). The previous studies
indicated that LRG1 may be a potential therapeutic target for
pathological de novo blood vessel formation. However,
whether LRG1 has a critical role in the brain during angiogenesis
following ischemic stroke and the mechanism underlying its
promotion of neovascularization is remains unknown. In the present
study, the changes of LRG1, TGF-β1, VEGF and Ang-2 a rat model of
middle cerebral artery occlusion (MCAO) were determined. The
results demonstrated that LRG1 may promote angiogenesis through
upregulating the TGF-β1 signaling pathway in ischemic rat brain
after MCAO.
Materials and methods
Animals and MCAO
Adult male Sprague-Dawley rats weighing 200–250 g
were purchased from the Center for Experimental Animals, the Second
Affiliated Hospital of Harbin Medical University (Harbin, China).
Animals were housed at a controlled temperature (23°C) and humidity
(50%), under a 12-h light/dark cycle with access to food and water
ad libitum throughout the experiment. All experiments were
approved by the Institutional Animal Care and the Ethics Committee
of Harbin Medical University.
Rats were divided randomly into 2 groups: the
control and MCAO groups. In the control group (n=6), no surgical
procedure was performed. For the rat model of MCAO, rats were
anesthetized with 10% chloral hydrate (400 mg/kg; intraperitoneal
injection; Zhanyun Institute of Biotechnology, Shanghai, China) and
a line model (diameter=0.234 mm) was inserted into the internal
carotid artery to block the middle cerebral artery as previously
described (28). Brain tissue
collection was performed postoperatively at 2 h, 6 h, 12 h, 1 day,
3 days, 7 days and 14 days (n=6 at each time point) to collect
infarcted brain samples for quantitative polymerase chain reaction
(qPCR), western blot analysis and immunohistochemical staining at
each time interval. The rats were anesthetized and sacrificed with
10% chloral hydrate (800 mg/kg; intraperitoneal injection) at each
of the time points, and the rat brain was perfused with 4%
paraformaldehyde (Zhanyun Institute of Biotechnology).
Reverse transcription-qPCR
(RT-qPCR)
Total mRNA was extracted from control or ischemic
rat brains using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) according to the manufacturer's
instructions. The purity of mRNA samples was detected using the
NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Inc.,
Wilmington, DE, USA). cDNA was synthesized using the Transcriptor
First Stand cDNA Synthesis kit (Roche Diagnostics, Basel,
Switzerland). qPCR was performed using SYBR Green (Roche
Diagnostics) on a thermocycler. The primers used are listed as
follows: LRG1, forward 5′-GCATCAAGGGAGAACCCTGT-3′, reverse
5′-CGTGCTTCCCTTAACCGACT-3′; TGF-β1, forward
5′-CAGTGGCTGAACCAAGGAGA-3′, reverse 5′-GGAAGGGTCGGTTCATGTCA-3′;
VEGF, forward 5′-CTGCTCTCTTGGGTGCACT-3′, reverse
5′-ATACACTATCTCATCGGGGTACT-3′; Ang-2. forward
5′-TTGCGACCCCTTCAACTCTG-3′, reverse 5′-CCCTTGGGCTTGGCATCATA-3′;
GAPDH, forward 5′-GCATCTTCTTGTGCAGTGCC-3′, reverse
5′-TACGGCCAAATCCGTTCACA-3′. The samples were amplified out using a
Smart Cycler (Cepheid, Inc., Sunnyvale, CA, USA) as follows:
Denatured at 95°C for 2 min, 40 cycles of amplification (95°C for
20 sec, 60°C for 30 sec and 72°C for 20 sec), and a final extension
at 72°C for 10 min. Expression data were normalized to GAPDH mRNA
expression. Data were calculated using the 2−ΔΔCq method
(29).
Western blotting
Tissues were obtained from control or ischemic rat
brains (n=6/group) and total cell lysates were prepared using lysis
buffer (Beyotime Institute of Biotechnology, Haimen, China).
Protein concentration was measured using the bicinchoninic acid
method with bovine serum albumin (Baoman Institute of
Biotechnology, Shanghai, China) as the standard (30). Equal amounts (50–100 µg) of whole
cell lysates were resolved by 8–12% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and transferred to
polyvinylidene difluoride membranes (EMD Millipore, Billerica, MA,
USA). The membrane was blocked with TBS-0.1% Tween 20 (TBS-T)
containing 5% skim milk at room temperature for 1 h. The membrane
was then incubated with monoclonal mouse anti-mouse LRG1 (1:50;
cat. no. sc-390920), polyclonal rabbit anti-human TGF-β1 (1:100;
cat. no. sc-146), polyclonal rabbit anti-human Ang-2 (1:200; cat.
no. sc-20718) and monoclonal mouse anti-human VEGF (1:200; cat. no.
sc-7269) primary antibody (Santa Cruz Biotechnology, Inc., Dallas,
TX, USA) diluted in blocking solution at 4°C overnight and washed
with TBS-T three times every 10 min, followed by incubation with
goat anti-mouse or goat anti-rabbit secondary antibody (1:2,000;
cat. nos. sc-2031 and sc-2030, respectively; Santa Cruz
Biotechnology, Inc.) at room temperature for 1 h. Subsequent to
washing in TBS-T, the protein bands were visualized using enhanced
chemiluminescence kits (Beyotime Institute of Biotechnology). Blots
were stained with monoclonal mouse anti-human β-actin antibody
(1:1,000; cat. no. TA-09; Zhongshan Biotechnology Co., Ltd.,
Beijing, China) as an internal control for the amount of target
proteins. Band intensity of each sample was determined using a UVP
iBox 500 imaging system (UVP, Inc., Upland, CA, USA)
Immunohistochemical staining
Tissue samples from control or ischemic rat brains
were fixed in 4% formalin at 25°C for 36 h and embedded in
paraffin, then were cut into 4 µm thick sections. Paraffin sections
were dewaxed with xylene and rehydrated by graded concentrations of
ethanol. Then endogenous peroxidase was blocked by 0.3% hydrogen
peroxide for 10 min. The sections were rehydrated with EDTA (pH
9.0) at 120°C for 2 min and cooled for 40 min at room temperature.
Subsequent to blocking for 30 min with bovine serum albumin,
primary antibody to monoclonal mouse anti-mouse LRG1 (1:50; cat.
no. sc-390920), polyclonal rabbit anti-human TGF-β1 (1:100; cat.
no. sc-146), polyclonal rabbit anti-human Ang-2 (1:200; cat. no.
sc-20718) and monoclonal mouse anti-human VEGF (1:200; cat. no.
sc-7269) primary antibody (Santa Cruz Biotechnology, Inc.) and CD34
(1:200; cat. no. ab185732; Abcam, Cambridge, MA, USA) were
incubated overnight at 4°C in humidified chambers (Santa Cruz
Biotechnology, Inc.). The goat anti-mouse/rabbit secondary antibody
(cat. no. PV-9000; Zhongshan Biotechnology Co., Ltd., Beijing,
China) was incubated for 30 min at room temperature.
Diaminobenzidine was then used to detect positive staining (cat.
no. ZLI-9017, Zhongshan Biotechnology Co., Ltd.,). Then sections
were counterstained with hematoxylin. Sections were viewed under a
light microscope and images were captured with a CCD camera
(DMI6000B; Leica Microsystems GmbH, Wetzlar, Germany) using
imaging-processing software (LAS AF6000; Leica Microsystems GmbH).
The mean percentage of positively-stained cells in 5 high power
fields per sample (magnification, ×200) of each group was used as
the final value.
Microvessel density (MVD) analysis. CD34
staining was performed as an indicator of MVD. The density of
microvessels was assessed according to the criteria proposed by
Weidner et al (31). Any
endothelial cells or endothelial cell clusters brown-stained in the
cytoplasm were considered as a countable microvessel. The average
number of microvessels in 5 high-power fields/sample (×200
magnification) of each group were used as the final value. The
assessments were performed by a single pathologist who was blinded
to the data.
Statistical analysis
SPSS 13.0 (SPSS, Inc., Chicago, IL, USA) was used
for the statistical analysis. The data are presented as the mean ±
standard deviation. Differences among the means were analyzed using
one-way analysis of variance and Student-Newman-Keuls method.
Spearman rank correlation coefficient test analysis was performed
to examine the correlations between LRG1, and TGF-β1, Ang-2, VEGF
and MVD. P<0.05 was considered to indicate a statistically
significant difference.
Results
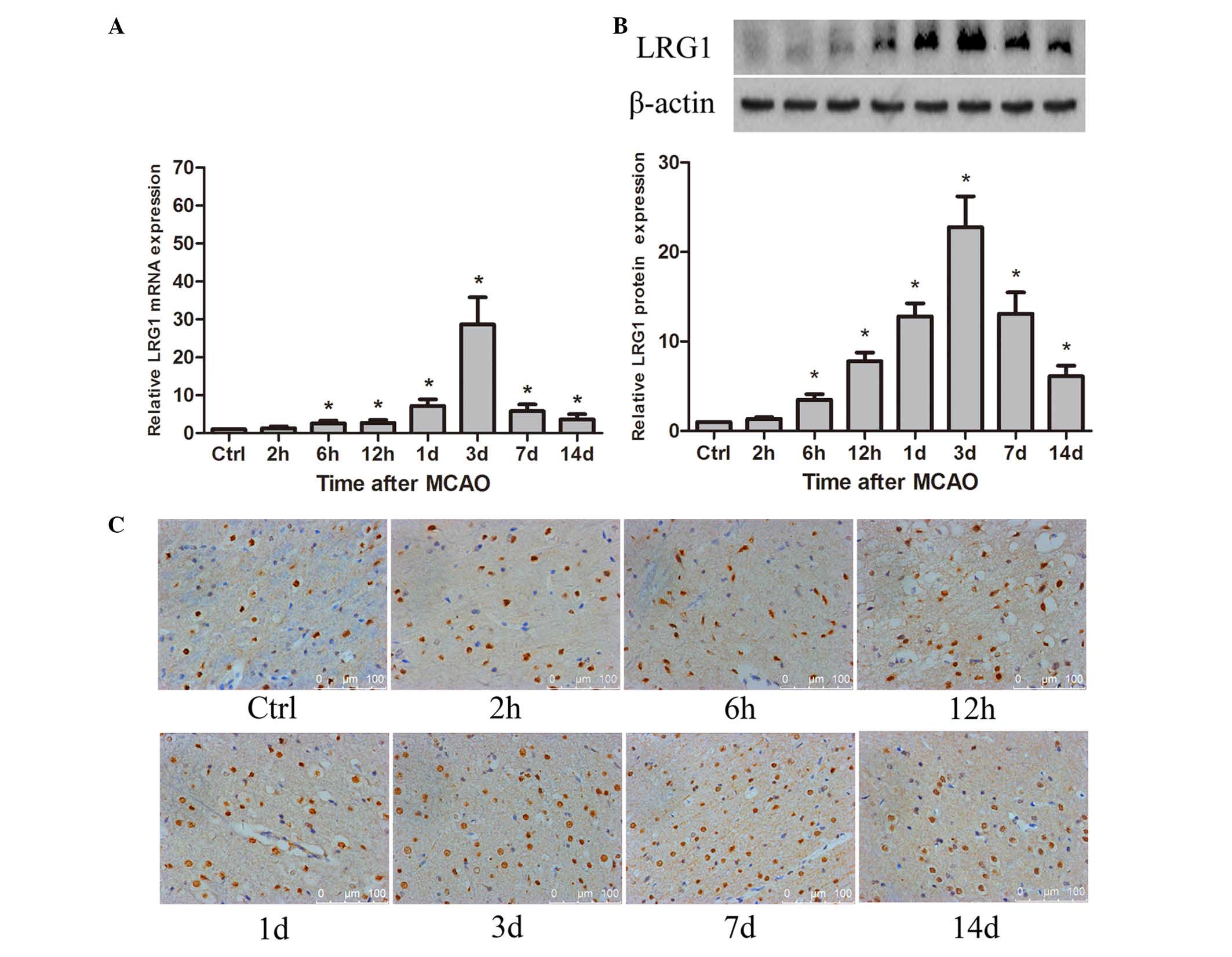
Expression of LRG1 was increased in
ischemic rat brain following MCAO
To demonstrate whether LRG1 was involved in
angiogenesis in ischemic rat brain following MCAO, the spatial and
temporal expression of LRG1 was detected. The results revealed that
LRG1 mRNA was significantly increased compared with the level in
the control as early as 6 h (P<0.05), peaked at 3 days after
MCAO, then gradually decreased, however the level of LRG1 at 14
days remained significantly higher than the control groups
(P<0.05; Fig. 1A). The level of
LRG1 protein was significantly elevated compared with the control
at 6 h (P<0.05), reached a maximum at 3 days, persisting at a
higher level compared with the control until 14 days after MCAO
(P<0.05; Fig. 1B). The
immunohistochemistry staining indicated that LRG1 protein was
located in the cellular nucleus (Fig.
1C). LRG1 was expressed in the endothelial cells of
microvessels, and also in the astrocyte-like cells (Fig. 1C). Furthermore, the percentage of
LRG1-positive cells was significantly higher than the control at 12
h in the ischemic penumbra of the rat brain following MCAO
(P<0.05), peaked at 3 days and gradually declined to the normal
level at 14 days (Table I).
 | Table I.Percentage of LRG1, TGF-β1, VEGF,
Ang-2-positive cells and MVD in ischemic penumbra of rat brain at
different time points following middle cerebral artery
occlusion. |
Table I.
Percentage of LRG1, TGF-β1, VEGF,
Ang-2-positive cells and MVD in ischemic penumbra of rat brain at
different time points following middle cerebral artery
occlusion.
| Time | LRG1+
(%) | TGF-β1+
(%) | VEGF+
(%) | Ang-2+
(%) | MVD |
|---|
| Control | 52.64±11.35 | 6.24±3.23 | 12.21±6.68 | 15.12±6.08 | 9.83±3.71 |
| 2 h | 53.19±6.68 | 6.06±2.88 |
27.36±9.18a | 29.84±10.99 | 10.63±2.09 |
| 6 h | 55.11±12.73 | 14.51±4.44 |
37.11±7.08a |
38.28±10.21a | 10.77±3.69 |
| 12 h |
59.02±7.98a |
22.02±3.99a |
54.24±13.13a |
36.87±6.72a |
13.20±3.54a |
| 1 days |
63.65±13.37a |
39.59±6.14a |
56.98±14.17a |
46.78±9.90a |
21.53±4.22a |
| 3 days |
68.43±6.05a |
66.90±9.66a |
76.23±7.37a |
57.30±9.22a |
26.63±4.18a |
| 7 days |
66.38±7.22a |
58.05±7.91a |
71.36±6.86a |
38.11±6.98a |
24.83±5.06a |
| 14 days | 56.77±11.31 |
45.77±10.00a |
57.46±11.75a |
32.74±6.58a |
14.83±3.30a |
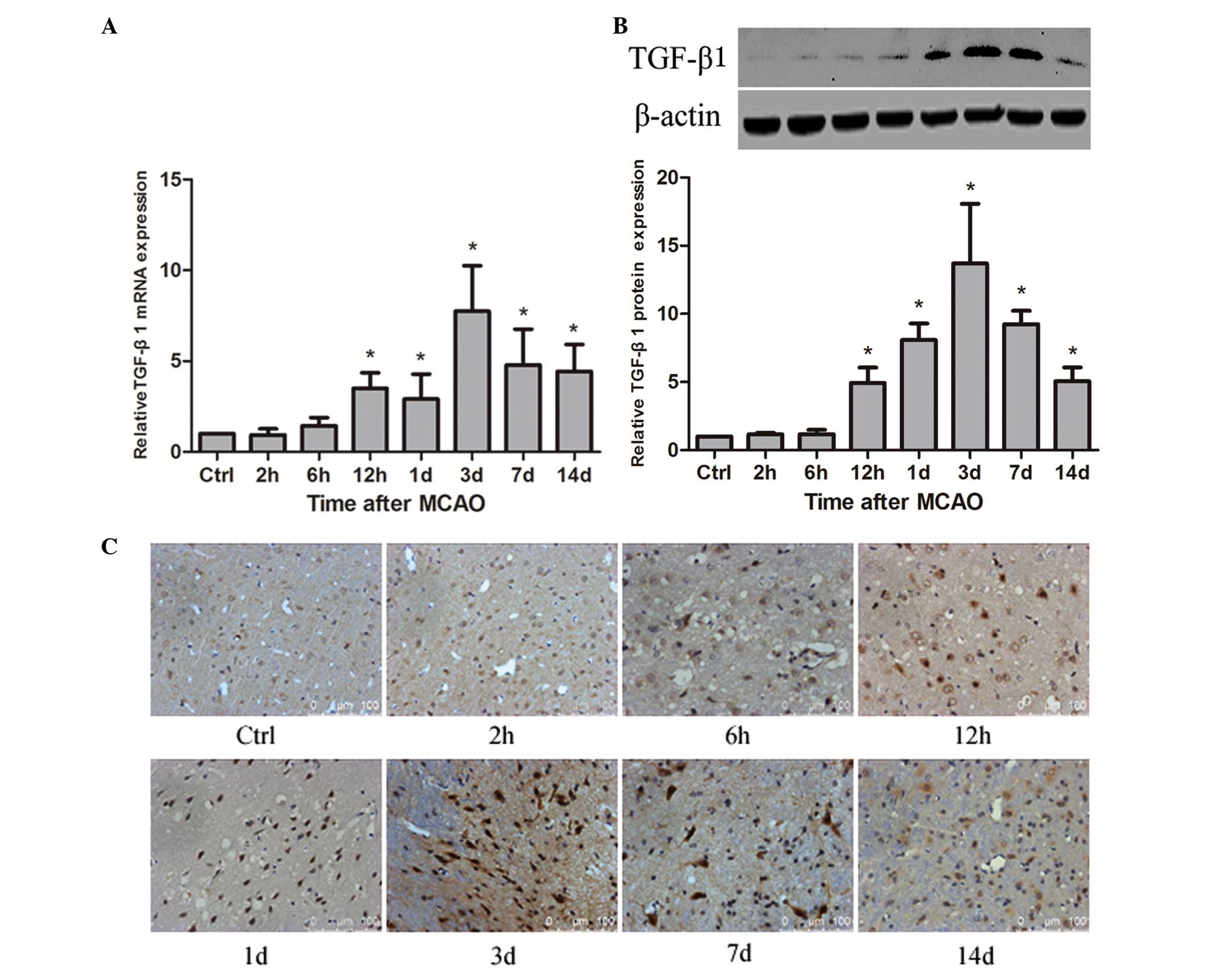
Expression of TGF-β1 is upregulated in
ischemic rat brain following MCAO
To detect whether TGF-β1 is associated with
angiogenesis in rat brain following MCAO, the spatial and temporal
expression of TGF-β1 was examined. The results demonstrated that
TGF-β1 mRNA was significantly increased at 12 h compared with the
control (P<0.05) and reached a maximum at 3 days after MCAO,
persisting at a significantly higher level until 14 days
(P<0.05; Fig. 2A). TGF-β1
protein was significantly upregulated at 12 h compared with the
control (P<0.05), peaked at 3 days after MCAO, and remained a
significantly higher level until 14 days (P<0.05; Fig. 2B). The immunohistochemistry
staining indicated that TGF-β1 protein was distributed to the
cellular nucleus (Fig. 2C).
Compared with the control group, the percentage of TGF-β1-positive
cells was significantly increased at 12 h (P<0.05), peaked at 3
days in the ischemic penumbra of the rat brain following MCAO,
persisted at a significantly higher level at 7 day (P<0.05) and
gradually decreased until 14 days (Table I).
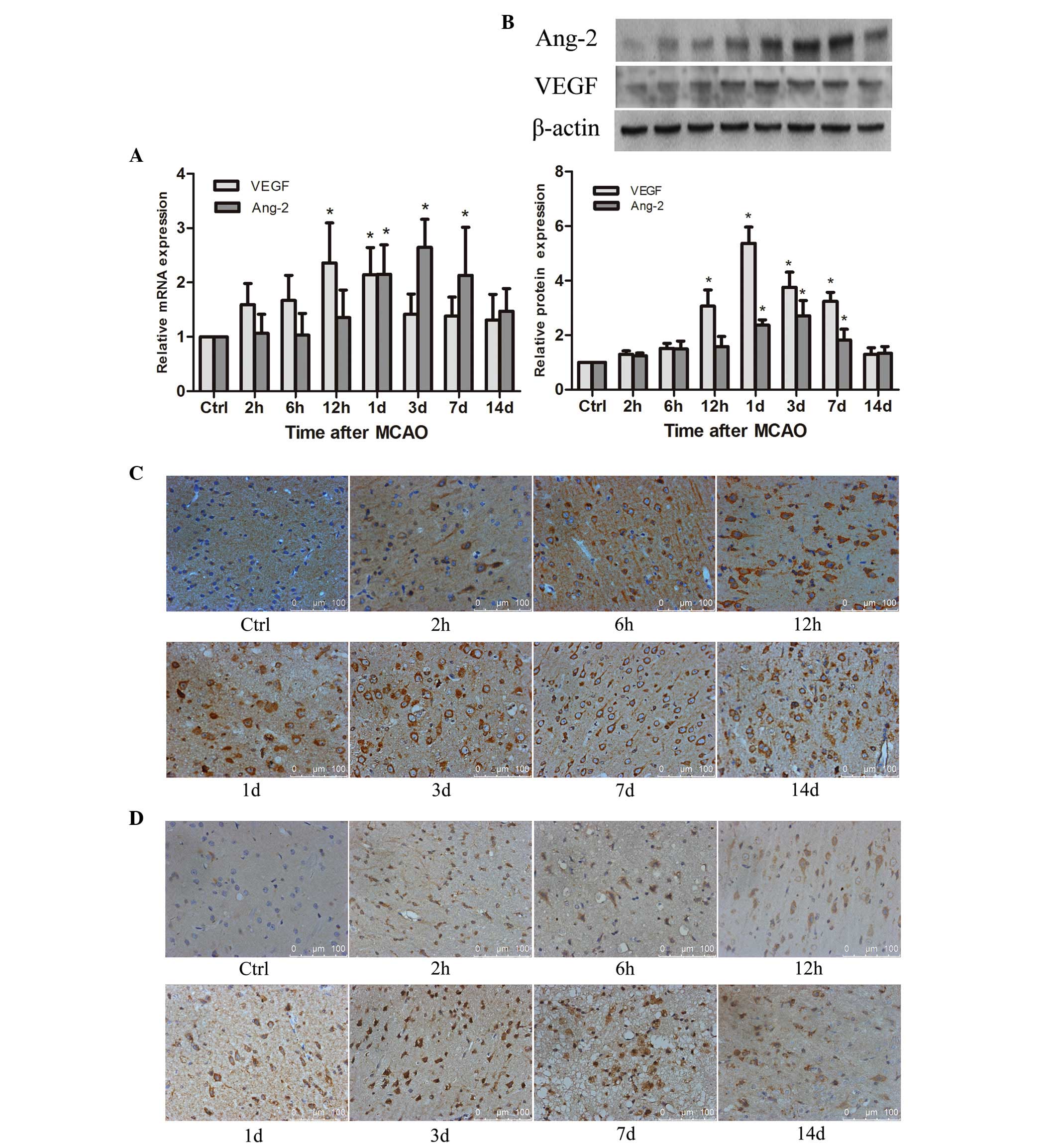
Expression of angiogenic factors, VEGF
and Ang-2, are elevated in ischemic rat brain following MCAO
As the angiogenic factors, the spatial and temporal
expression of VEGF and Ang-2 were examined in ischemic rat brain
following MCAO. The results suggested that VEGF mRNA was
significantly elevated compared with the control levels as early as
6 h (P<0.05), reached a peak at 12 h, and remained a
significantly higher level until 1 day after MCAO (P<0.05), then
decreased gradually (Fig. 3A).
VEGF protein was significantly increased compared with the control
at 12 h (P<0.05), peaked at 1 days, persisted at a higher level
until 7 days (P<0.05), then declined to the normal level
(Fig. 3B). Immunohistochemistry
demonstrated that VEGF protein was predominantly located in the
cellular cytoplasm of neuron-like and glial-like cells in the
ischemic penumbra of the rat brain (Fig. 3C). Compared with the control group,
the percentage of VEGF-positive cells was significantly higher in
the ischemic penumbra of the rat brain at 2 h after MCAO
(P<0.05), reached a maximum at 3 days and remained at a
significantly higher level at 14 days (P<0.05; Table I).
The results also revealed that Ang-2 mRNA was
significantly increased at 1 day after MCAO compared with the
control (P<0.05), reached a maximum at 3 d and remained a higher
level at 7 days after MCAO (P<0.05), then gradually declined to
the normal baseline (Fig. 3A). The
Ang-2 protein was significantly upregulated at 1 day compared with
the control (P<0.05), peaked at 3 days after MCAO, persisting at
a significantly higher level until 7 days (P<0.05), and declined
to the baseline (Fig. 3B).
Immunohistochemistry demonstrated that Ang-2 protein was
predominantly located in the cellular cytoplasm of neuron-like and
glial-like cells in the ischemic penumbra (Fig. 3D). Furthermore, the percentage of
Ang-2-positive cells was significantly higher in the ischemic
penumbra of the rat brain at 6 h after MCAO compared with the
control (P<0.05), reached a peak at 3 days, still persisted at a
significantly higher level until 14 days (P<0.05; Table I).
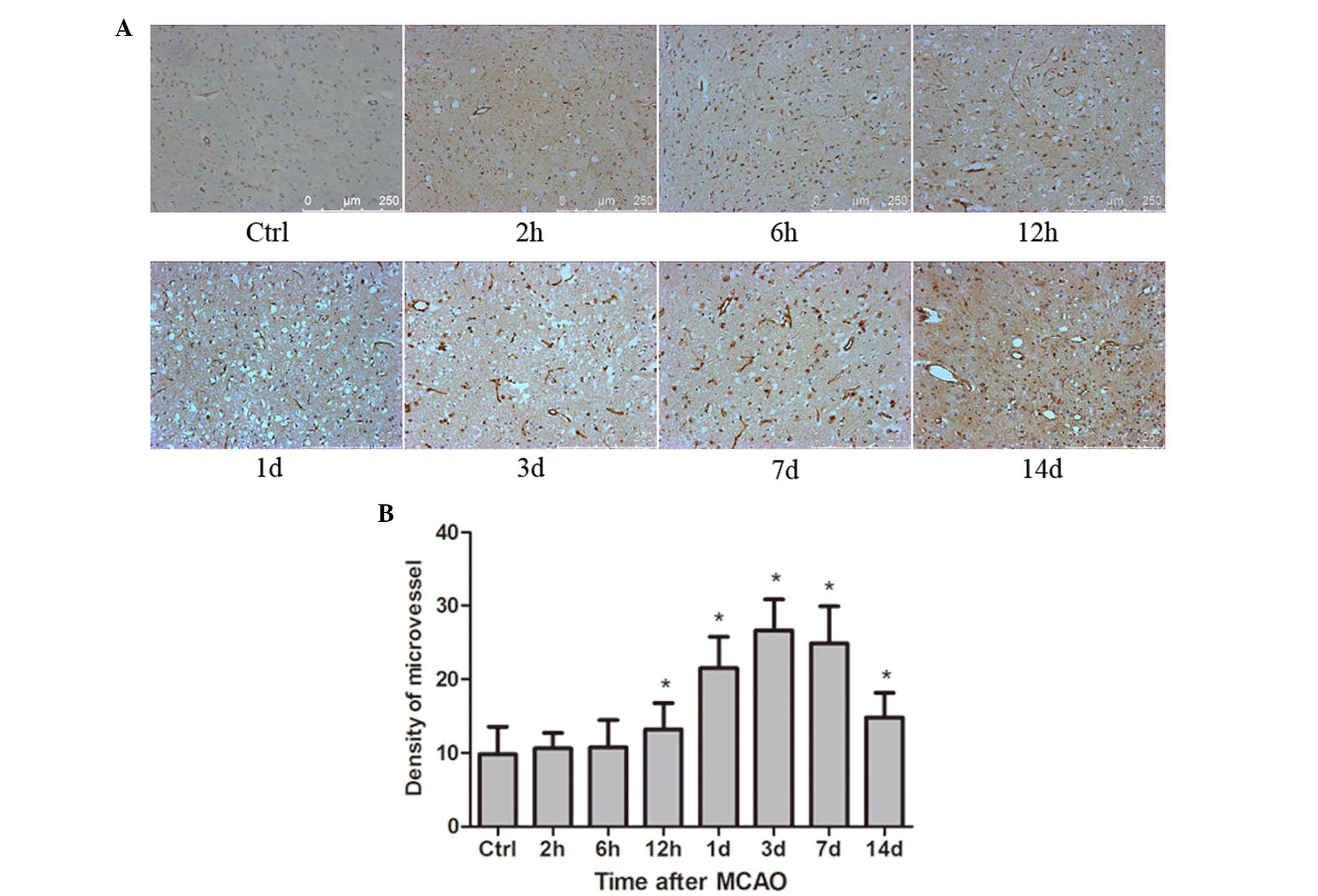
Formation of blood vessels in response
to the ischemic rat brain following MCAO
Ischemic rat brains at the different time points
after MCAO (n=6 per group) were paraffin-embedded and
formalin-fixed. Sections (4 µm thick) from the brain tissues were
used to detect the expression of CD34 protein. CD34 staining was
performed as an indicator of MVD (Fig.
4A). The average number of microvessels in 5 high-power
fields/sample (×200 magnification) of each group was used as the
final value. The results indicated that the mean value of MVD was
significantly increased compared with the control at 12 h
(P<0.05), reached a peak at 3 days, and remained a significantly
increased level until 14 days (P<0.05; Fig. 4B).
Using Spearman correlation analysis, the
immunohistochemistry results demonstrated that the percentage of
LRG1-positive cells was positively correlated with the percentage
of TGF-β1-positive, VEGF-positive, Ang-2-positive cells and the MVD
(Table II). Furthermore, the
results also suggested that TGF-β1-positive cells had a positively
correlation with the percentage of VEGF-positive cells and the MVD
(Table II). In addition, the
percentage of VEGF-positive cells and the percentage of
Ang-2-positive cells were positively correlated with the MVD
(Table II).
| Table II.Correlations of LRG1, TGF-β1, VEGF,
Ang-2-positive cells and MVD in ischemic penumbra of rat brain
following middle cerebral artery occlusion. |
Table II.
Correlations of LRG1, TGF-β1, VEGF,
Ang-2-positive cells and MVD in ischemic penumbra of rat brain
following middle cerebral artery occlusion.
| Parameter |
LRG1+% |
TGF-β1+% |
VEGF+% |
Ang-2+% | MVD |
|---|
| LRG1+% | – | 0.002a | 0.001a | 0.015a | 0.000a |
| TGF-β1+% | – | – | 0.000a | 0.071 | 0.000a |
| VEGF+% | – | – | – | 0.058 | 0.000a |
| Ang-2+% | – | – | – | – | 0.021a |
Discussion
Stroke is subdivided into ischemic, intracerebral
putic hemorrhagic and subarachnoid hemorrhagic stroke (32). It is a serious cause of human
mortality and disability. Opening the collateral circulation and
recovering the perfusion of blood flow in the ischemic penumbra
following stroke plays is crucial for protecting the structure and
function of the ischemic brain tissue (33,34).
Therefore, it is important to investigate the mechanisms of
angiogenesis following stroke.
A novel angiogenic factor, LRG1, is a highly
conserved member of the LRR family of proteins. It is a secreted
glycoprotein. The LRR structure of LRG1 has been proposed to be
important for its biological function (20). It regulates diverse biological
processes, including cell proliferation, cell apoptosis, cell
mobility and cell adhesion (10,11,13).
Wang et al (26) reported that LRG1 expression was
significantly increased during retinal neovascular remodeling of
mouse eye compared with in the normal mouse retina. The study also
observed that the expression of LRG1 was not restricted to the
mouse retina, but also expressed in the vasculature of mouse
choriocapillaries (26).
Consistent with the data, the study reported that LRG1 was
expressed in the vessels of retina, breast, skin and intestine in
normal adult human at a low level (26). Based on the previous study, LRG1
was declared as a novel regulator of neovascularization with
expression restricted almost exclusively to the vasculature system
(26). Thus, it was speculated
that LRG1 may be involved in the regulation of angiogenesis
following stroke. To demonstrate whether LRG1 is involved in
cerebral ischemia, the current study detected LRG1 expression in
ischemic rat brain at 2 h, 6 h, 12 h, 1 day, 3 days, 7 days and 14
days after MCAO. The results suggested that LRG1 mRNA and protein
were significantly upregulated as early as 6 h after MCAO and
peaked at 3 days, persisting at a significantly higher level
compared with control until 14 days (Fig. 1A). In addition to increased LRG1
expression, the mean value of MVD was also significantly increased
(Fig. 4A). The above results
indicated that LRG1 may promote de novo blood formation in
ischemic rat brain following MCAO. The blood formation will benefit
the recovery of blood flow in the ischemic penumbra, which may
protect the ischemic brain from injury. Thus, the data of the
current study further indicated that LRG1 may protect the ischemic
brain by recovering the blood flow due to the formation of novel
blood in the ischemic penumbra.
LRG1 expression has diverse cellular locations,
including the nucleus, cytoplasm, plasma membrane, extracellular
matrix, serum and cerebrospinal fluid (12,14,20).
Nakajima et al (35)
reported that LRG1 is distributed throughout the entire brain.
Double staining of CD31 and LRG1 revealed marked expression of LRG
in the capillaries, and the cells expressing LRG1 exhibited similar
morphology to astrocytes (35).
The results of the present investigation revealed that LRG1 protein
was predominantly located in the cellular nucleus of ischemic rat
brain (Fig. 1C). LRG1 was not only
expressed in vasculature system, but also in astrocyte-like cells.
The results were in accordance with the previous study (35). Thus, we hypothesize that in
ischemic rat brain after MCAO, the expression of LRG1 was
significantly increased in endothelial and astrocyte cells, and
secreted into the extracellular environment, then involved in the
regulation of angiogenesis and protected the ischemic brain.
Wang et al (26) reported that in vitreous samples of
human subjects with proliferative diabetic retinopathy, in addition
to increased LRG1 protein expression, TGF-β1 protein level was also
significantly upregulated. Furthermore, with increased LRG1 gene
expression, the TGF-β1 mRNA transcript level was also concomitantly
elevated in the models of choroidal and retinal neovascularization
(26). Takemoto et al
(36) reported that LRG1
potentiated the effect of TGF-β1 in Lewis lung carcinoma cells.
Furthermore, LRG1 was implicated as an upstream regulator of the
TGF-β1 signaling pathway in non-small-cell lung carcinoma (37). In addition, LRG1 has a strong
affinity with TGF-β1 in high endothelial venules (27). These data indicated that LRG1 may
act as a modulator of the TGF-β signaling pathway. This prompted us
to investigate whether LRG1 was involved in angiogenesis via the
TGF-β signaling pathway in ischemic rat brain following MCAO. In
the present study, TGF-β1 mRNA and protein were significantly
increased compared with the control at 12 h and reached a maximum
at 3 days after MCAO, persisting at a significantly higher level
until 14 days (Fig. 2A and B).
Consistent with increased LRG1 expression, TGF-β1 expression was
also significantly elevated. This indicated that in ischemic rat
brain following MCAO, LRG1 may protect the ischemic rat brain from
injury through promoting angiogenesis via upregulating the TGF-β1
signaling pathway.
Wang et al (26) demonstrated that in the presence of
TGF-β1, LRG1 binds to activin A receptor like type 1 (ALK1), a TGF-
β signaling pathway receptor, and activated its downstream
mediators, Smads (26,38), which are involved in the
angiogenesis by inducing the expression and secretion of vascular
growth factors, including VEGF and Ang-2 (39,40).
Angiogenic factors, VEGF and Ang-2, promote the proliferation of
endothelial cells and the initiation of angiogenesis (41). In the current study, the levels of
VEGF and Ang-2 mRNA and protein expression were significantly
increased at 12 h and 1 day in ischemic rat brain after MCAO, with
the peak at 1 day and 3 days, respectively. Then they gradually
declined to the baseline. Therefore, we hypothesize that the
expression of LRG1 was upregulated in endothelial and
astrocyte-like cells and secreted during the angiogenesis process
following cerebral ischemia. Additionally, LRG1 bound to the TGF-β1
receptor, ALK1, and activated Smads downstream. Then, the ischemic
brain secreted angiogenic factors, including VEGF and Ang-2, which
promoted angiogenesis.
A recent publication implied that LRG1 was
positively correlated with TGF-β receptor 1 in cardiac fibrosis
(19). In the current study, using
Spearman correlation analysis, the immunohistochemistry results
revealed that the percentage of LRG1-positive cells was positively
correlated with the percentage of TGF-β1-positive cells.
Furthermore, the percentage of LRG1-positive and TGF-β1-positive
cells had a positively correlation with the MVD (Table II). The results further indicated
that LRG1 may promote angiogenesis through upregulating the TGF-β1
signaling pathway. The immunohistochemistry results were consistent
with the RT-qPCR and western blot results.
In summary, the results of the present study
indicated that in ischemic rat brain following MCAO, LRG1
expression was increased in astrocyte-like cells and secreted from
cells, then may have promoted the de novo blood vessel
formation by inducing the expression angiogenic factors, including
VEGF and Ang-2, through upregulating TGF-β1 signaling. In order to
fully identify the mechanisms that LRG1 regulating the angiogenesis
in ischemic brain, future studies are required.
Acknowledgements
We would like to thank Mr. Xiang Fang, Mr. Hongda
Wang, Ms. Miaomiao Jiang and Ms. Yunhe Gu (Department of Pathology,
The First Affiliated Hospital of Harbin Medical University, Harbin,
China) for technical support. This study was supported by the
Natural Science Fund of China (grant no. 30973106).
References
|
1
|
Lundberg GP and Volgman AS: Burden of
stroke in women. Trends Cardiovasc Med. 26:81–88. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Verma V, Samanthapudi K and Raviprakash R:
Classic studies on the potential of stem cell neuroregeneration. J
Hist Neurosci. 25:123–141. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Esposito E, Hayakawa K, Maki T, Arai K and
Lo EH: Effects of postconditioning on neurogenesis and angiogenesis
during the recovery phase after focal cerebral ischemia. Stroke.
46:2691–2694. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang ZG and Chopp M: Promoting brain
remodeling to aid in stroke recovery. Trends Mol Med. 21:543–548.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Moss A: The angiopoietin: Tie 2
interaction: A potential target for future therapies in human
vascular disease. Cytokine Growth Factor Rev. 24:579–592. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kwon HS, Kim YS, Park HH, Choi H, Lee KY,
Lee YJ, Heo SH, Chang DI and Koh SH: Increased VEGF and decreased
SDF-1α in patients with silent brain infarction are associated with
better prognosis after first-ever acute lacunar stroke. J Stroke
Cerebrovasc Dis. 24:704–710. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shaikh H, Boudes E, Khoja Z, Shevell M and
Wintermark P: Angiogenesis dysregulation in term asphyxiated
newborns treated with hypothermia. PLoS One. 10:e01280282015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Haupt H and Baudner S: Isolation and
characterization of an unknown, leucine-rich
3.1-S-alpha2-glycoprotein from human serum (author's transl). Hoppe
Seylers Z Physiol Chem. 358:639–646. 1977.(In German). View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Takahashi N, Takahashi Y and Putnam FW:
Periodicity of leucine and tandem repetition of a 24-amino acid
segment in the primary structure of leucine-rich alpha
2-glycoprotein of human serum. Proc Natl Acad Sci USA.
82:1906–1910. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kobe B and Kajava AV: The leucine-rich
repeat as a protein recognition motif. Curr Opin Struct Biol.
11:725–732. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kobe B and Deisenhofer J: Proteins with
leucine-rich repeats. Curr Opin Struct Biol. 5:409–416. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Serada S, Fujimoto M, Terabe F, Iijima H,
Shinzaki S, Matsuzaki S, Ohkawara T, Nezu R, Nakajima S, Kobayashi
T, et al: Serum leucine-rich alpha-2 glycoprotein is a disease
activity biomarker in ulcerative colitis. Inflamm Bowel Dis.
18:2169–2179. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wong CC, Tse AP, Huang YP, Zhu YT, Chiu
DK, Lai RK, Au SL, Kai AK, Lee JM, Wei LL, et al: Lysyl
oxidase-like 2 is critical to tumor microenvironment and metastatic
niche formation in hepatocellular carcinoma. Hepatology.
60:1645–1658. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang Y, Luo Q, Wang N, Hu F, Jin H, Ge T,
Wang C and Qin W: LRG1 suppresses the migration and invasion of
hepatocellular carcinoma cells. Med Oncol. 32:1462015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wu J, Yin H, Zhu J, Buckanovich RJ, Thorpe
JD, Dai J, Urban N and Lubman DM: Validation of LRG1 as a potential
biomarker for detection of epithelial ovarian cancer by a blinded
study. PLoS One. 10:e01211122015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhong D, Zhao S, He G, Li J, Lang Y, Ye W,
Li Y, Jiang C and Li X: Stable knockdown of LRG1 by RNA
interference inhibits growth and promotes apoptosis of glioblastoma
cells in vitro and in vivo. Tumour Biol. 36:4271–4278. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu YS, Luo XY, Li QR, Li H, Li C, Ni H,
Li RX, Wang R, Hu HC, Pan YJ, et al: Shotgun and targeted
proteomics reveal that pre-surgery serum levels of LRG1, SAA and
C4BP may refine prognosis of resected squamous cell lung cancer. J
Mol Cell Biol. 4:344–347. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wen SY, Zhang LN, Yang XM, Zhang YL, Ma L,
Ge QL, Jiang SH, Zhu XL, Xu W, Ding WJ, et al: LRG1 is an
independent prognostic factor for endometrial carcinoma. Tumour
Biol. 35:7125–7133. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kentsis A, Ahmed S, Kurek K, Brennan E,
Bradwin G, Steen H and Bachur R: Detection and diagnostic value of
urine leucine-rich α-2-glycoprotein in children with suspected
acute appendicitis. Ann Emerg Med. 60:78–83.e1. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shirai R, Hirano F, Ohkura N, Ikeda K and
Inoue S: Up-regulation of the expression of leucine-rich
alpha(2)-glycoprotein in hepatocytes by the mediators of
acute-phase response. Biochem Biophys Res Commun. 382:776–779.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Song W and Wang X: The role of TGFβ1 and
LRG1 in cardiac remodeling and heart failure. Biophys Rev.
7:91–104. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Watson CJ, Ledwidge MT, Phelan D, Collier
P, Byrne JC, Dunn MJ, McDonald KM and Baugh JA: Proteomic analysis
of coronary sinus serum reveals leucine-rich α2-glycoprotein as a
novel biomarker of ventricular dysfunction and heart failure. Circ
Heart Fail. 4:188–197. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pek SL, Tavintharan S, Wang X, Lim SC,
Woon K, Yeoh LY, Ng X, Liu J and Sum CF: Elevation of a novel
angiogenic factor, leucine-rich-α2-glycoprotein (LRG1), is
associated with arterial stiffness, endothelial dysfunction, and
peripheral arterial disease in patients with type 2 diabetes. J
Clin Endocrinol Metab. 100:1586–1593. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nakajima M, Miyajima M, Ogino I, Watanabe
M, Miyata H, Karagiozov KL, Arai H, Hagiwara Y, Segawa T, Kobayashi
K and Hashimoto Y: Leucine-rich α-2-glycoprotein is a marker for
idiopathic normal pressure hydrocephalus. Acta Neurochir (Wien).
153:1339–1346; discussion 1346. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Miyajima M, Nakajima M, Motoi Y, Moriya M,
Sugano H, Ogino I, Nakamura E, Tada N, Kunichika M and Arai H:
Leucine-rich α2-glycoprotein is a novel biomarker of
neurodegenerative disease in human cerebrospinal fluid and causes
neurodegeneration in mouse cerebral cortex. PLoS One. 8:e744532013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang X, Abraham S, McKenzie JA, Jeffs N,
Swire M, Tripathi VB, Luhmann UF, Lange CA, Zhai Z, Arthur HM, et
al: LRG1 promotes angiogenesis by modulating endothelial TGF-β
signaling. Nature. 499:306–311. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Saito K, Tanaka T, Kanda H, Ebisuno Y,
Izawa D, Kawamoto S, Okubo K and Miyasaka M: Gene expression
profiling of mucosal addressin cell adhesion molecule-1+ high
endothelial venule cells (HEV) and identification of a leucine-rich
HEV glycoprotein as a HEV marker. J Immunol. 168:1050–1059. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zan L, Wu H, Jiang J, Zhao S, Song Y, Teng
G, Li H, Jia Y, Zhou M, Zhang X, et al: Temporal profile of Src,
SSeCKS, and angiogenic factors after focal cerebral ischemia:
Correlations with angiogenesis and cerebral edema. Neurochem Int.
58:872–879. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yu Q, Zhao Z, Sun J, Guo W, Fu J,
Burnstock G, He C and Xiang Z: Expression of P2X6 receptors in the
enteric nervous system of the rat gastrointestinal tract. Histochem
Cell Biol. 133:177–188. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Weidner N, Folkman J, Pozza F, Bevilacqua
P, Allred EN, Moore DH, Meli S and Gasparini G: Tumor angiogenesis:
A new significant and independent prognostic indicator in
early-stage breast carcinoma. J Natl Cancer Inst. 84:1875–1887.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mozaffarian D, Benjamin EJ, Go AS, Arnett
DK, Blaha MJ, Cushman M, de Ferranti S, Després JP, Fullerton HJ,
Howard VJ, et al: Heart disease and stroke statistics-2015 update:
A report from the American heart association. Circulation.
131:e29–e322. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yoo SY and Kwon SM: Angiogenesis and its
therapeutic opportunities. Mediators Inflamm. 2013:1271702013.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Manoonkitiwongsa PS: Critical questions
for preclinical trials on safety and efficacy of vascular
endothelial growth factor-based therapeutic angiogenesis for
ischemic stroke. CNS Neurol Disord Drug Targets. 10:215–234. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nakajima M, Miyajima M, Ogino I, Watanabe
M, Hagiwara Y, Segawa T, Kobayashi K and Arai H: Brain localization
of leucine-rich α2-glycoprotein and its role. Acta Neurochir Suppl.
113:97–101. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Takemoto N, Serada S, Fujimoto M, Honda H,
Ohkawara T, Takahashi T, Nomura S, Inohara H and Naka T:
Leucine-rich α-2-glycoprotein promotes TGFβ1-mediated growth
suppression in the Lewis lung carcinoma cell lines. Oncotarget.
6:11009–11022. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li Y, Zhang Y, Qiu F and Qiu Z: Proteomic
identification of exosomal LRG1: A potential urinary biomarker for
detecting NSCLC. Electrophoresis. 32:1976–1983. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Oh SP, Seki T, Goss KA, Imamura T, Yi Y,
Donahoe PK, Li L, Miyazono K, ten Dijke P, Kim S and Li E: Activin
receptor-like kinase 1 modulates transforming growth factor-beta 1
signaling in the regulation of angiogenesis. Proc Natl Acad Sci
USA. 97:2626–2631. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Orlova VV, Liu Z, Goumans MJ and ten Dijke
P: Controlling angiogenesis by two unique TGF-β type I receptor
signaling pathways. Histol Histopathol. 26:1219–1230.
2011.PubMed/NCBI
|
|
40
|
Ueki Y and Reh TA: Activation of
BMP-Smad1/5/8 signaling promotes survival of retinal ganglion cells
after damage in vivo. PLoS One. 7:e386902012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Poittevin M, Bonnin P, Pimpie C, Rivière
L, Sebrié C, Dohan A, Pocard M, Charriaut-Marlangue C and Kubis N:
Diabetic microangiopathy: Impact of impaired cerebral
vasoreactivity and delayed angiogenesis after permanent middle
cerebral artery occlusion on stroke damage and cerebral repair in
mice. Diabetes. 64:999–1010. 2015. View Article : Google Scholar : PubMed/NCBI
|