Introduction
Methamphetamine (MA) abuse is a growing health
problem worldwide, and there has been an increase in the number of
medical complications and fatalities associated with the toxicity
of MA (1,2). A previous study (3) reported that the widest distribution
and the highest uptake of MA in the human body occurred in the
lungs, and that this may render the lungs vulnerable to infection,
pulmonary hypertension and pulmonary edema (3,4). The
long-lasting pulmonary toxic effects of MA have placed an increased
burden on healthcare costs (5).
Therefore, it is particularly important to investigate the
mechanism of pulmonary toxicity for drug targets. A previous study
(6) demonstrated that serotonin is
possibly associated with MA-induced pulmonary toxicity. Pulmonary
toxicity is pathologically characterized by parenchymal damage,
recruitment of inflammatory cells and a progression of the
inflammatory processes (7).
Inflammatory changes in lung tissue are the key to the pulmonary
toxicity. However, problems remain about the exact pathogenesis of
MA-induced chronic pulmonary inflammation through serotonin.
The neurotransmitter serotonin (5-hydroxytryptamine;
5-HT) is implicated in increasing inflammatory reactions of the
skin, lungs and gastrointestinal tract (8). An etiological agent of chronic
inflammation is deregulation of the tissue macrophage polarization
balance (9). The modulation of the
phenotypic and functional polarization of macrophages is by 5-HT,
which regulates the inflammation and the tissue restoration via a
large set of receptors (5-HTR1-7) and/or a transporter (9). Lung macrophages express the serotonin
transporter (SERT) and 5-HT receptors 2a, and 2b (10). A previous study (11) reported that the inhibition of
5-HTR2a and 5-HTR2b had no effect on efferocytosis, although
inhibiting SERT prevented 5-HT-impaired efferocytosis. A previous
epidemiological study (12)
suggested that MA abuse significantly increased the risk of
developing pulmonary arterial hypertension (PAH). The disease
severity and susceptibility to PAH may be associated with the
increased SERT activity (13).
Pulmonary vascular remodeling and pulmonary inflammation in
pulmonary hypertension were associated with a SERT-induced rapid
activation of extracellular signal-regulated kinase (ERK) 1/2
(14,15). The phosphorylation and subsequent
activation of ERK, p38 mitogen activated protein kinases (p38 MAPK)
and Akt, and the production of reactive oxygen species (ROS)
appeared to be a common mechanism of proliferation and inflammation
(16,17). A previous study (18) reported that MAPK signaling is
involved in the regulation of nuclear factor erythroid 2-related
factor 2 (Nrf2).
Nrf2 is a basic leucine zipper redox-sensitive
transcriptional factor that serves a central role in the
transcriptional regulation of antioxidant and/or detoxifying genes
(19). Nuclear localization of
Nrf2 activation efficiently protects cells from ROS-induced damage
in vivo and in vitro by inducing the expression of
numerous detoxifying enzymes and antioxidant proteins (20). As Nrf2 is a transcription factor
with potent antioxidant effects against cell death caused by
ROS-induced damage, targeting Nrf2 may serve an essential role in
the protection against various inflammatory diseases (20). However, the role of Nrf2 in
MA-induced pulmonary inflammation and the protective mechanism of
fluoxetine against MA-induced oxidative stress and pulmonary
inflammation remain to be elucidated. Therefore, the present study
was designed to further evaluate the potential role of Nrf2 and to
investigate if fluoxetine can ameliorate MA-induced oxidative
stress and pulmonary inflammation throughthep38 MAPK/Nrfr2 pathway
in rats.
Materials and methods
Drugs
MA was obtained from the China Criminal Police
University (Shenyang, Liaoning, China). The purity of the MA was
identified as 97% by a Bio-Rad REMEDi HS system (Bio-Rad, Milan,
Italy) and by liquid chromatography-mass spectrometry-mass
spectrometry (Shimadzu Corporation, Kyoto, Japan). MA was dissolved
in 0.9% sterile saline and prepared as 4 mg/ml for drug
administration.
Animal and experimental paradigm
A total of 30 male Wistar rats (180±10 g) were
obtained from the Animal Resource Center, China Medical University
(Shenyang, Liaoning, China; certificate number: Liaoning 034) and
divided into four groups: i) Control; ii) MA; iii) MA plus
fluoxetine 2 mg/kg (MA+F2); and iv) MA plus fluoxetine 10 mg/kg
(MA+F10). Rats in the MA and the two fluoxetine-treated groups were
also treated daily with intraperitoneal injection of 10 mg/kg MA
(China Criminal Police University) twice daily for 5 weeks. Rats in
the control group received the respective vehicles only. Rats in
the MA+F2 and MA+F10 groups were treated with intragastric
fluoxetine (Cadila Pharmaceuticals, Ankleshwar, India) at 2 or 10
mg/kg once daily for 5 weeks, respectively. All the rats were
housed in a controlled humidity (50–70%) and temperature (18–22°C),
and were given access to food and water ad libitum in an
alternating 12 h light/dark cycle over a period of 5 weeks. All
experimental protocols for the present study were approved by the
Institutional Animal Care and Use Committee of China Medical
University.
Morphological analysis by hematoxylin
and eosin (H&E) staining
A total of 3% sodium pentobarbital (45 mg/kg) was
used to euthanize the rats. The right lower lung tissues were
dissected, fixed with paraformaldehyde and embedded in paraffin
wax. Sections (5 µm) were stained with H&E for observation and
analysis under light microscopy. The inflammatory changes of rat
lung parenchyma were evaluated by the thickness of alveolar septum
and the destructive index (DI; three randomly selected sites were
analyzed in each section; magnification, ×200).
DI, a measure of alveolar septal damage and
emphysema, has been proposed as a sensitive index of lung
destruction that closely reflects functional abnormalities
(21). DI represents the
percentage of destroyed space as a fraction of the total alveolar
and duct space (21). The
quantification of this destruction can add greatly to the
microscopic analysis of changes due to pulmonary inflammation.
Immunohistochemistry
After processing the tissue and embedding in
paraffin wax, 5 µm thick sections were stained by
immunohistochemical (IHC) procedures using Ultrasensitive TM SP kit
(Maxin-Bio Co., Fuzhou, China) and DAB Staining kit (Zhongshan
Golden Bridge Biotechnology Co., Ltd., Beijing, China). IHC
staining followed a basic indirect protocol using a citrate antigen
retrieval method. A primary rabbit anti-interleukin-6 (IL-6; cat.
no. bs 0379R, Beijing Biosynthesis Biotechnology Co., Ltd.,
Beijing, China) was diluted at 1:50 and incubated overnight at 4°C.
For the negative control, the primary antibody was replaced by 0.01
M PBS in the incubation step. A biotin-labeled secondary antibody
from SP kit (cat. no. KIT-9706; Maxin-Bio Co.) was incubated for 10
min at room temperature to detect the primary antibody.
The positive expression of IL-6 was examined using
light microscopy. At least six visual fields of lung tissue were
examined on each slide. For the convenience of understanding and
statistical processes, the quantity of protein in the rat lung was
analyzed and calculated as optical density average by microscope
with a digital camera and MetaMorph software version 7.7 (Molecular
Devices, LLC, Sunnyvale, CA, USA).
Western blot analysis
Nuclear and cytoplasmic fractions were extracted
using a Nuclear and Cytoplasmic Protein Extraction kit (Beyotime
Institute of Biotechnology, Shanghai, China). The protein
concentrations were determined using a BCA protein assay kit
(Beyotime Institute of Biotechnology) prior to storage at −80°C.
Electrophoresis was performed with 10% SDS-polyacrylamide gel using
80 µg total protein in each lane. Following electrophoresis, the
protein was transferred onto a PVDF membrane using a semi-dry
transfer unit (Bio-Rad Laboratories, Inc., Hercules, CA, USA) and
the membranes were incubated for 1 h in blocking buffer (5% non-fat
dry milk, PBS and 0.1% Tween). The membranes were probed with
primary mouse monoclonal anti-β-actin (1:2,000; cat. no. sc-130300;
Santa Cruz Biotechnology, Inc., Dallas, USA) and α-tubulin 1:2,000;
cat. no. 66031; ProteinTech Group, Inc., Chicago, IL, USA) and
rabbit polyclonal anti-SERT (1:200; cat. no. bs 1893R), IL-6
(1:200; cat. no. bs 0379R), tumor necrosis factor-α (TNF-α; 1:200;
cat. no. bs 0078R), human heme oxygenase-1 (HO-1; 1:400; cat. no.
bs 2075R) (Biosynthesis Biotechnology Co. Ltd.), anti-Nrf2 (1:600;
cat. no. 16396-1-AP; ProteinTech Group, Inc.), p38 (1:600; cat. no.
ZS-7149), and phosphorylated (p)-p38 (1:600; cat. no. ZS-101759;
Zhongshan Golden Bridge Biotechnology Co., Ltd.) overnight at 4°C.
Following washing with PBS-0.1% Tween, the membranes were incubated
in the presence of goat anti-mouse secondary antibody (cat. no.
ZB-2305; Zhongshan Golden Bridge Biotechnology Co., Ltd.) for
β-actin and α-tubulin at a dilution of 1:4,000 for 2 h, and goat
anti-rabbit secondary antibody (cat. no. SA00001-2; ProteinTech
Group, Inc.) for other proteins at a dilution of 1:2,000 for 2 h at
room temperature, followed by enhanced chemiluminescence (Pierce™
ECL Western Blotting Substrate; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). The relative protein expression was quantified
by densitometry using Molecular Dynamics Image Quant software (GE
Healthcare Life Sciences, Chalfont, UK). The result of the
expression of nuclear Nrf2 was represented by the relative yield
against α-tubulin, and the other protein expression levels were
represented by the relative yield against β-actin.
Reduced glutathione (GSH) and oxidized
glutathione (GSSG) assay
Lung tissues were homogenized with 10 ml ice-cold
lysis buffer (50 mM phosphate buffer containing 1 mM EDTA/g).
Following centrifugation at 10,000 × g for 15 min at 4°C, the
supernatant was removed, deproteinated and stored at −20°C until
further analyses. Total glutathione and oxidized glutathione levels
were determined using a GSH and GSSG Assay kit (Beyotime Institute
of Biotechnology), according to the manufacturer's protocol.
ELISA
Samples of the rat lung tissue in each group were
homogenized using a Polytron homogenizer (Kinematical, Lucerne,
Switzerland) to extract protein. The homogenate was centrifuged at
15,000 × g for 30 min at 4°C and the supernatant was collected and
stored at −80°C for ELISA. ROS concentration in the lung tissues
was measured by the rat ROS ELISA kits (R&D Systems, Inc.,
Minneapolis, MN, USA), according to the manufacturer's protocol.
The absorbance was measured at 450 nm and the corresponding
concentration was determined from the standard curve.
Statistical analysis
All data are presented as the mean ± standard
deviation. Statistical analysis was performed by one-way analysis
of variance with SPSS software, version 22.0 (IBM SPSS, Armonk, NY,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Effect of fluoxetine on the
inflammatory changes induced by MA in rat lungs
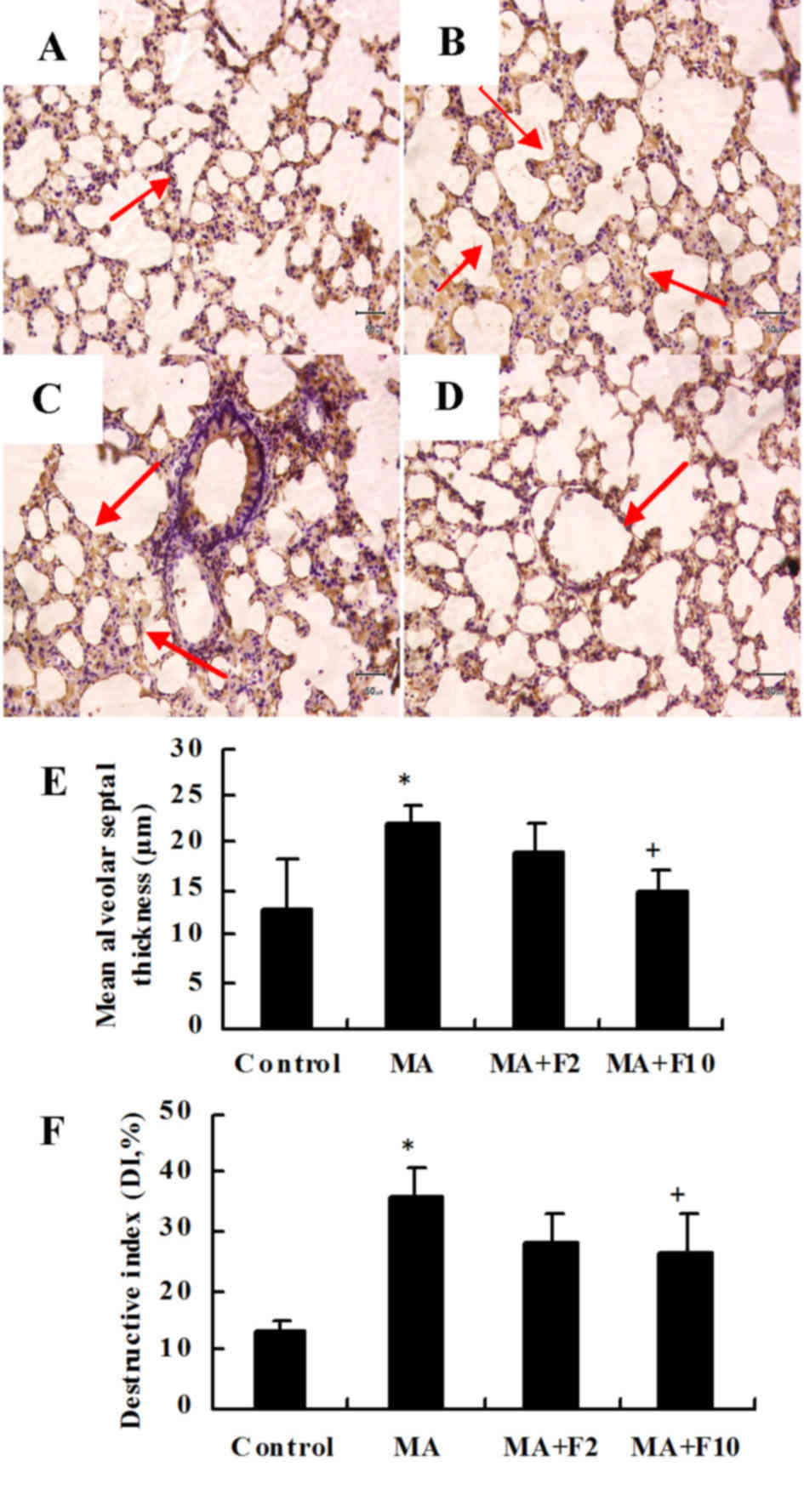
Representative H&E sections from the different
groups indicated that compared with the control group (Fig. 1A) lung injury was markedly induced
by 10 mg/kg MA (Fig. 1B) and
dose-dependently attenuated by fluoxetine (Fig. 1C and D).
Under the microscope, the rat lung tissue was
infiltrated by inflammatory cells in the MA group, the lung
parenchyma was more compact and the septum thickened (P=0.003, MA
vs. control; Fig. 1E). The DI was
significantly increased in the MA group compared with the control
group (P=0.007, MA vs. control; Fig.
1F). The inflammatory damage to the lung tissues was partly
ameliorated following the administration of fluoxetine at 10 mg/kg
(P=0.028 vs. MA).
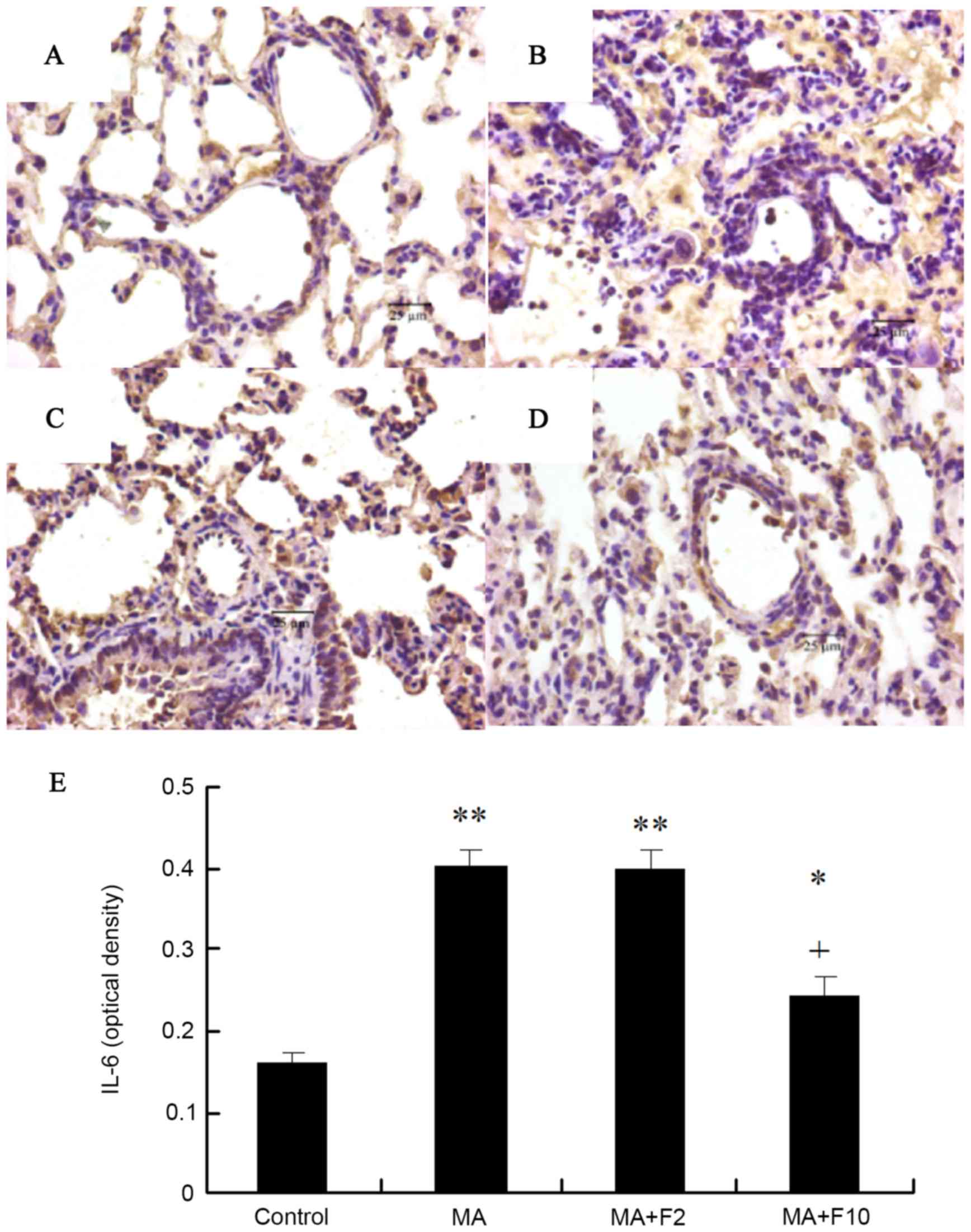
Immunohistochemical analysis of IL-6
protein expression in pulmonary arteries
Sections were stained with rabbit anti-IL-6 (brown)
and counterstained with hematoxylin (blue) in the different groups.
Compared with control group, IL-6 protein expression in the lungs
in the rats of the MA group was significantly increased (P=0.003),
and administration of fluoxetine dose-dependently decreased IL-6
protein expression. IL-6 expression in the rats of MA+F10 group was
significantly decreased, compared with the MA group (P=0.0174;
Fig. 2).
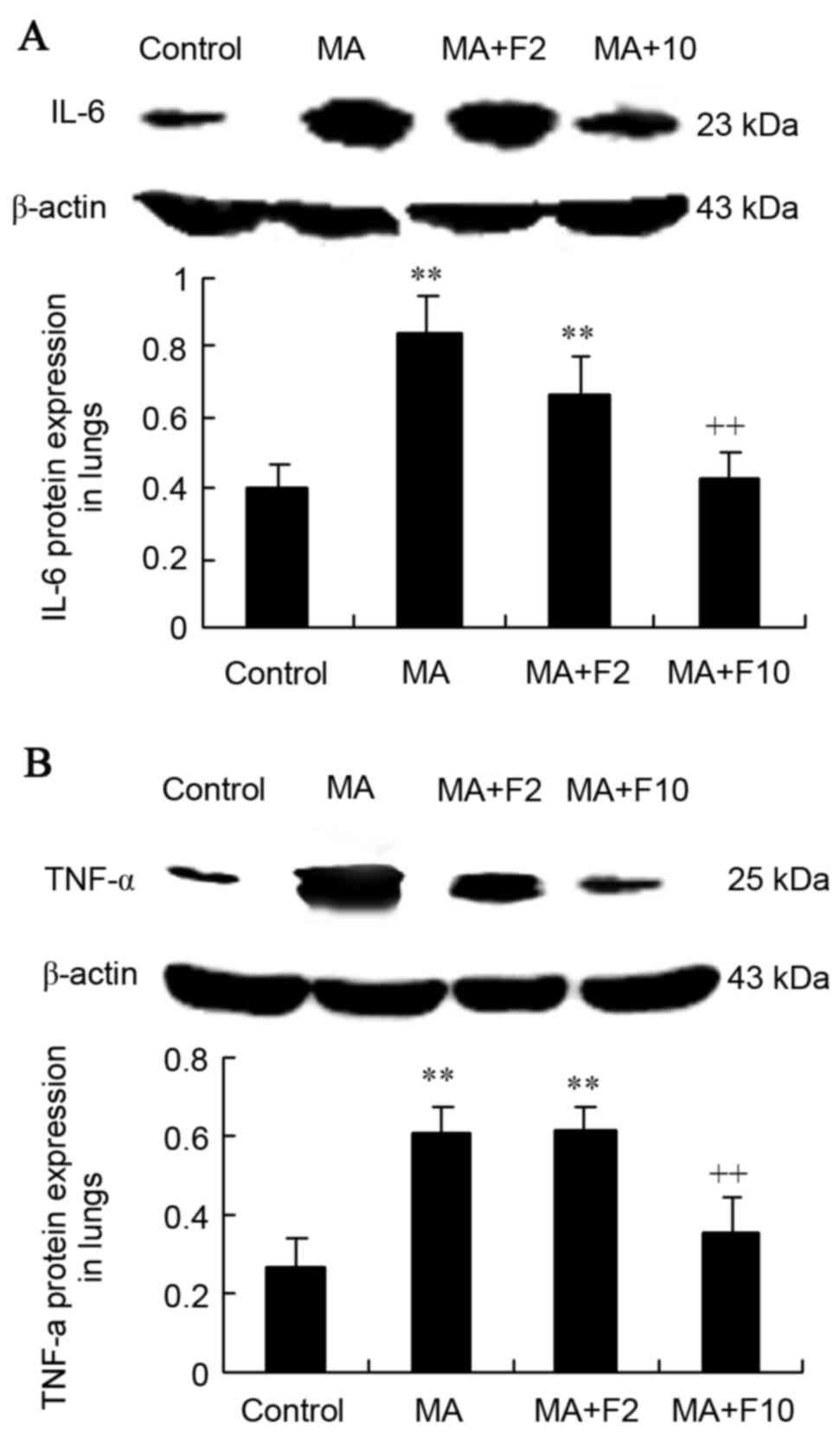
Western blot analysis of IL-6 and
TNF-α expression in rat lungs
Results from western blot analysis demonstrated that
compared with the control group, IL-6 protein expression was
significantly increased in the MA and MA+F2 groups (0.84±0.20 vs.
0.40±0.10, 0.67±0.10 vs. 0.40±0.10; P=0.004). IL-6 expression was
decreased following administration of fluoxetine at 10 mg/kg.
Compared with the MA group, relative IL-6 protein level was
significantly decreased in the MA+F10 group (0.43±0.10 vs.
0.84±0.20, P=0.006; Fig. 3A).
Western blot analysis was performed to further
demonstrate that TNF-α protein expression was significantly
increased in the MA group compared with the control group
(0.60±0.07 vs. 0.26±0.08; P=0.003). Compared with the MA group,
relative TNF-α protein expression in the lungs was not markedly
changed in the MA+F2 group, but was significantly decreased in the
MA+F10 group (0.35±0.09 vs. 0.60±0.07, P=0.032; Fig. 3B).
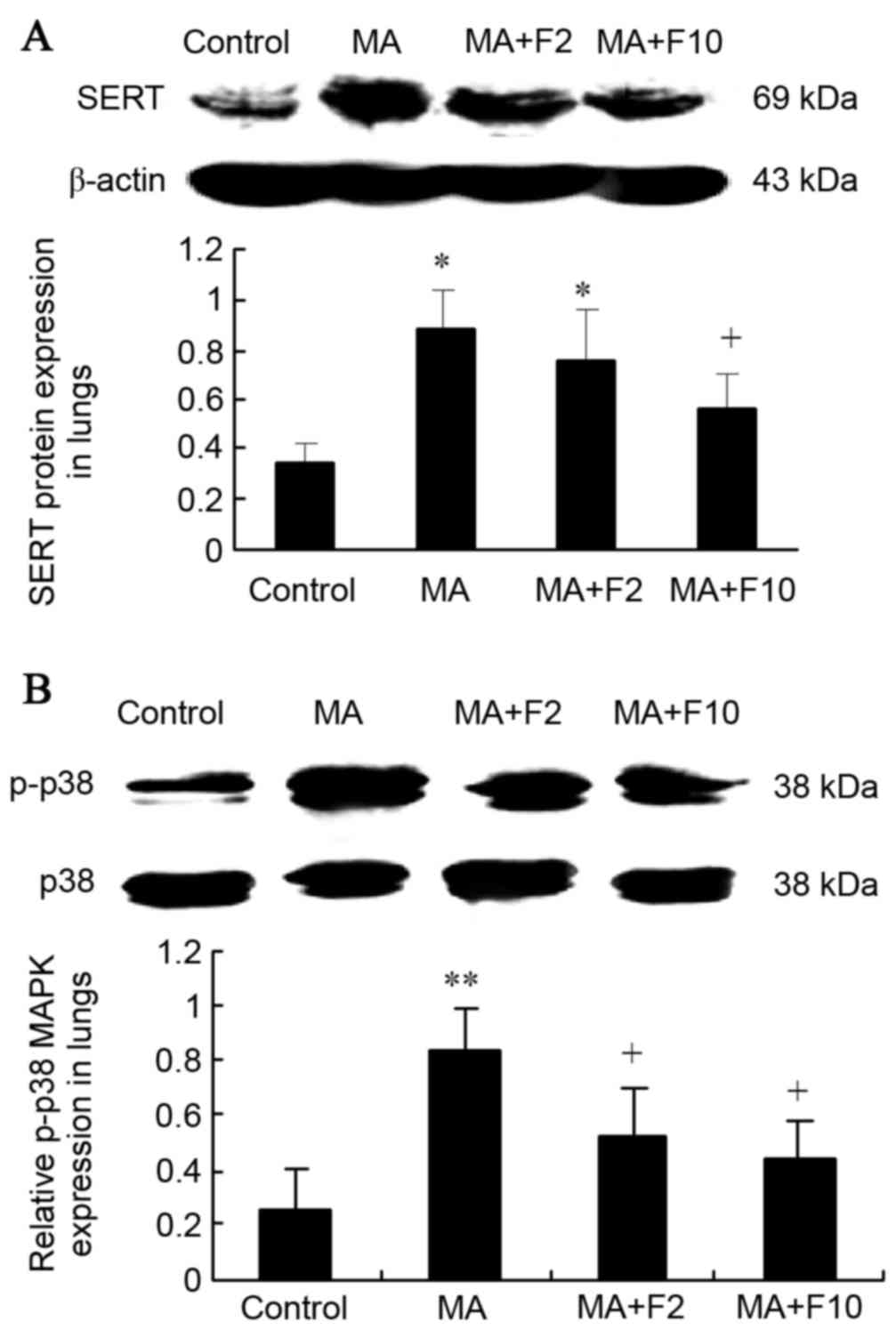
SERT and p38 MAPK protein expression
in lungs
Results from western blot analysis also demonstrated
that SERT expression was significantly upregulated in lungs in the
MA and MA+F2 groups, compared with the control group (0.89±0.15 vs.
0.34±0.08, P=0.005; 0.76±0.20 vs. 0.34±0.08, P=0.017), and that it
was downregulated in the MA+F10 group (0.57±0.14 vs. 0.89±0.18,
P=0.034), compared with the MA group (Fig. 4A).
Western blot analysis was performed to further
demonstrate that in the M10 group p38 MAPK was phosphorylated
compared with the control group (0.84±0.14 vs. 0.25±0.06; P=0.003).
In addition, p-p38 MAPK expression was decreased following
administration of fluoxetine. Compared with the M10 group, the
relative p-p38 MAPK protein level in lungs was markedly decreased
in the M10+F2 group (0.52±0.18 vs. 0.84±0.14; P=0.034) and in the
M10+F10 group (0.44±0.13 vs. 0.84±0.14, P=0.028; Fig. 4B).
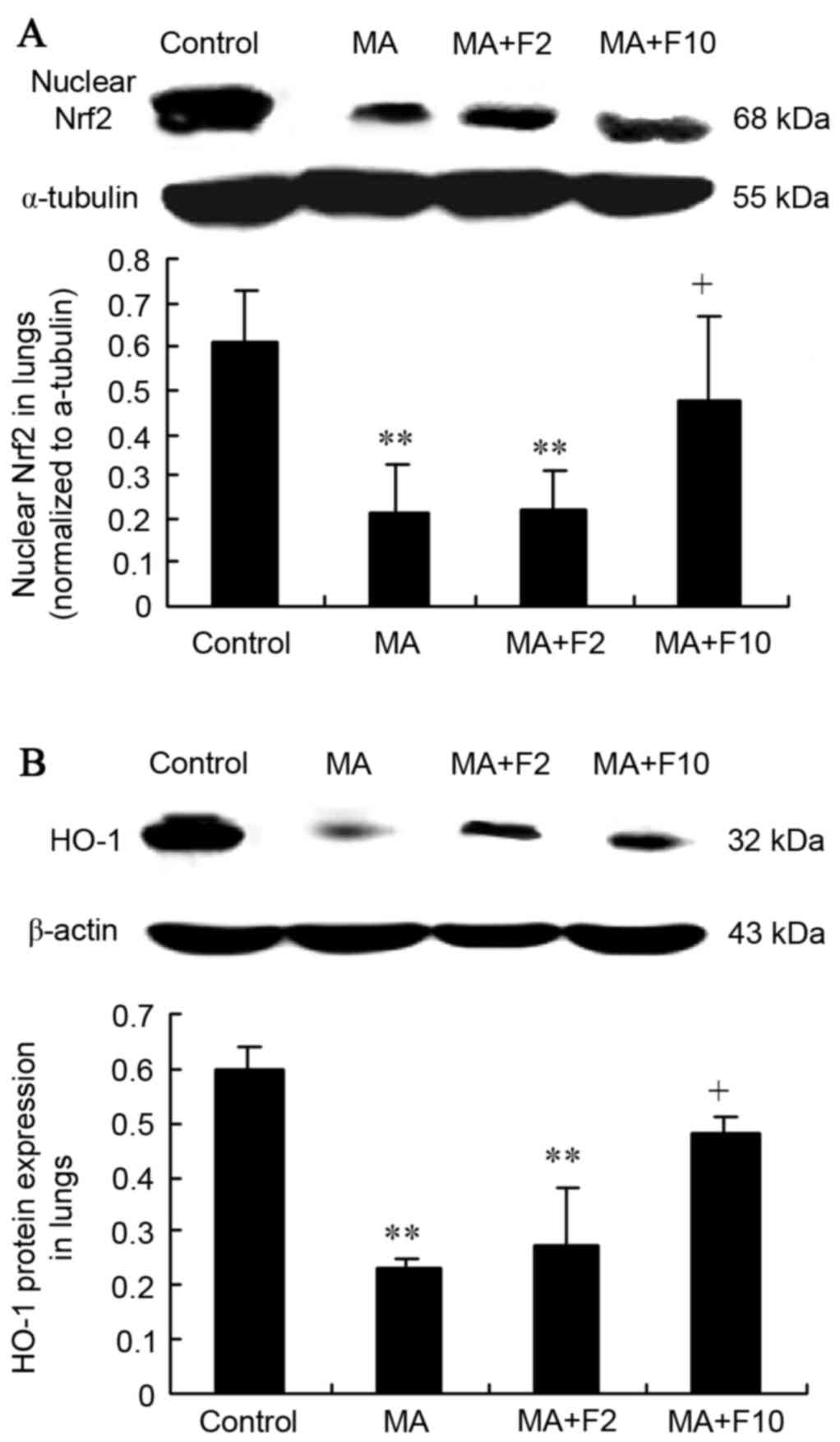
Nrf2/HO-1 expression in the lungs from
the different groups
Results from western blot analysis also demonstrated
that nuclear Nrf2 expression was significantly decreased in the
lungs in the MA and MA+F2 groups, compared with the control group
(0.21±0.11 vs. 0.61±0.12, 0.22±0.09 vs. 0.61±0.12, respectively;
P=0.004) and that it was increased in the MA+F10 group (0.48±0.19
vs. 0.21±0.0.11, P=0.032), compared with the MA group (Fig. 5A).
Western blot analysis further demonstrated that the
HO-1 expression was significantly downregulated in the lungs of the
MA and MA+F2 groups, compared with the control group (0.23±0.02 vs.
0.60±0.04, P=0.004; 0.27±0.11 vs. 0.60±0.04; P=0.005). However, it
was markedly upregulated in the MA+F10 group, compared with the MA
group (0.48±0.03 vs. 0.23±0.02, P=0.036; Fig. 5B).
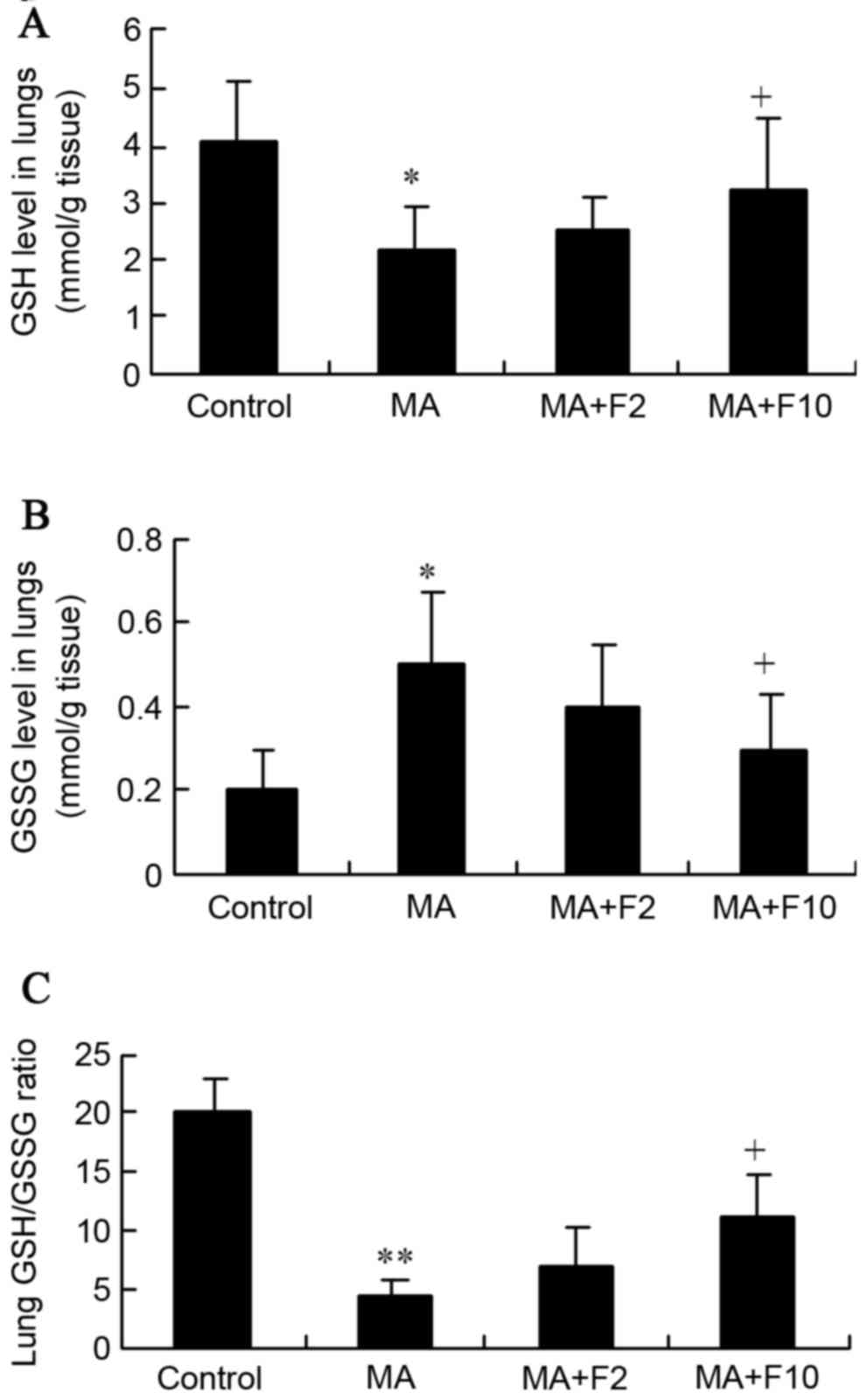
Effect of fluoxetine and MA on GSH and
GSSG in lungs
In the present study, MA markedly downregulated the
GSH level and upregulated the GSSG level in rat lungs, compared
with the control group (2.2±0.7 vs. 4.1±1.0, P=0.035; 0.5±0.18 vs.
0.2±0.1, P=0.027; Fig. 6A and B).
However, both were markedly reversed in the MA+F10 group compared
with the MA group.
Together with the decreased GSH level, the GSH/GSSG
ratios were significantly reduced by MA, as is presented in
Fig. 6C (4.4±1.4 vs. 20±3.0,
P=0.001). However, the ratio was markedly reversed from 4.4±1.4 in
the MA group to 11±3.8 in the MA+F10 group. These findings
supported the hypothesis that the protective effect of fluoxetine
was possibly performed by suppressing the Nrf2-mediated
antioxidative stress in the lungs.
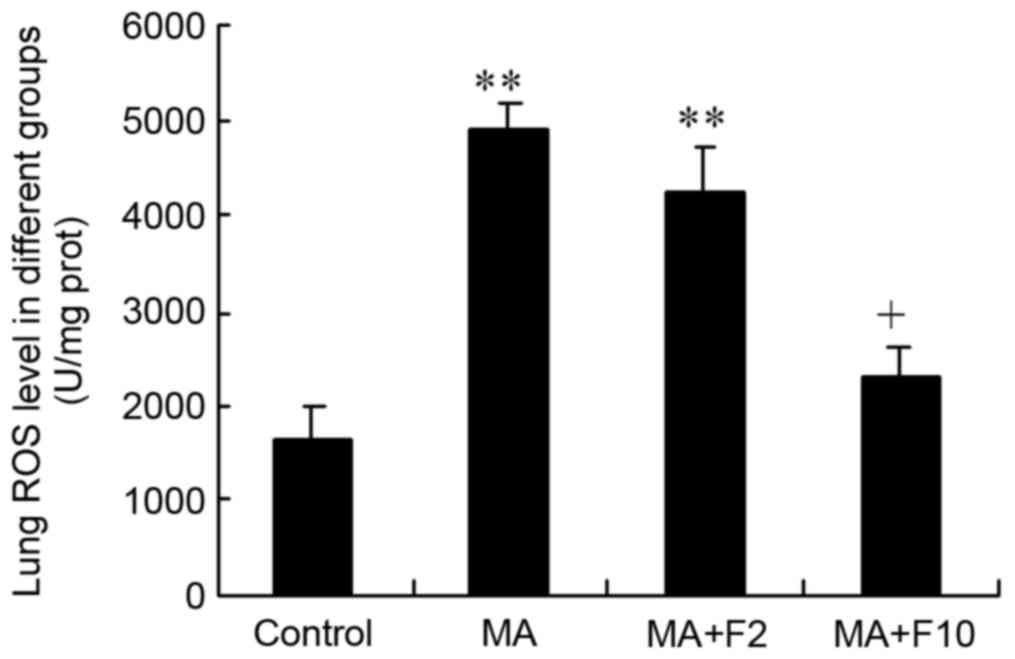
Effect of fluoxetine and MA on lung
ROS level
The results of the ELISA analysis demonstrated that
ROS in the lungs were at a low level in the control group,
(control: 1,608.4±364.6 U/mg protein). However, in the MA and MA+F2
groups, lung ROS levels were markedly enhanced to 4,938.8±265.1 and
4,256.2±470.1 U/mg protein (P<0.0001, vs. control). The ROS
level in lungs was significantly decreased in the MA+F10 group
(2,271.4±334.5 vs. 4,938.8±265.1 U/mg protein, P=0.015 vs. MCT;
Fig. 7).
Discussion
Results from the present study demonstrated that
chronic use of MA caused rat pulmonary inflammation; inflammatory
cell infiltration, crowded lung parenchyma, thickened septum and
increased DI. Fluoxetine attenuated the inflammatory changes and
the expression of the inflammatory factors IL-6 and TNF-α in rat
lungs. Fluoxetine also inhibited MA-induced increases in the
expression of SERT and p-p38 MAPK, and reversed the MA-induced
decrease in nuclear Nrf2 and HO-1 in lungs. Additionally,
fluoxetine at 10 mg/kg significantly reversed the increases in GSH
level, GSH/GSSG ratio and the ROS level in rat lungs from the MA
group. These findings suggested that fluoxetine, a SERT inhibitor,
has a protective effect against MA-induced lung inflammation by
suppressing oxidative stress through the SERT/p38 MAPK/Nrf2 pathway
in rats.
Pulmonary toxicity is pathologically characterized
by parenchymal damage, the recruitment of inflammatory cells and
the progression of the inflammatory process (7). Inflammatory changes in lung tissue
are the key to pulmonary toxicity. Endogenous IL-6 serves a
critical role in the inflammatory response to injury in the lungs
of mice (22). TNF-α is involved
in lung matrix fragmentation, macrophage activation and endothelial
cell apoptosis (23). Thus, both
IL-6 and TNF-α are the classical markers of pulmonary inflammation
(24). It was demonstrated in the
present study that the expression of IL-6 protein was significantly
increased in MA-induced lung inflammation in rats, that fluoxetine
dose-dependently inhibited the expression of IL-6 and TNF-α, and
that fluoxetine significantly attenuated the inflammatory cell
infiltration and reduced the inflammation of the alveoli with
thickened septum; this all suggested the protective effect of
fluoxetine on MA-induced lung inflammation.
Fluoxetine inhibited the SERT-mediated reuptake of
cytoplasmic 5-HT in the lungs (25). 5-HT, released from enterochromaffin
cells, can act on various innate and adaptive immune cells, and,
under inflammatory conditions, increased 5-HT production can
promote local inflammation (26).
In a previous study (6), it was
identified that MA induced the increasedconcentration of 5-HT,
which indicated that the serotonin mechanism is involved in
MA-induced pulmonary toxicity. MA, as a substrate for SERT, was
transported into cells and subsequently inhibited the metabolism of
5-HT by inhibiting monoamine oxidase A (6,27).
The activation of SERT can promote 5-HT-impaired efferocytosis
(11). SERT is the predominant
protein responsible for the uptake and release of serotonin by
transporting serotonin in either direction (28). Accordingly, SERT contributes to the
modulation of extracellular and intracellular 5-HT concentration
(29). In the present study, it
was demonstrated that fluoxetine at 10 mg/kg attenuated MA-induced
pulmonary inflammation and inhibited MA-induced upregulation of
SERT expression in the lung. These findings suggested that SERT may
serve an important role in MA-induced pulmonary inflammation.
SERT-induced rapid activation of MAPK is associated
with 5-HT-induced pulmonary inflammation (30). The p38 MAPK signaling pathways are
involved in the regulation of the Nrf2-mediated cytoprotective
effect (31). The present study
demonstrated that MA induced the upregulation of SERT and the
phosphorylation of p38 MAPK in the lungs. In addition, MA inhibited
the expression of Nrf2 in nucleoprotein. Therefore, it is
hypothesized that Nrf2 expression is possibly regulated by the
activation of SERT/p38 MAPK signaling in MA-induced pulmonary
inflammation.
Nrf2 is a transcription factor that controls the
expression of a variety of antioxidant and detoxification genes
(32). Nrf2 is an important
protein involved in the transcriptional upregulation of numerous
target genes in phase II drug metabolizing enzymes, including
γ-glutamylcysteine synthetase and HO-1 (32). Oxidant/antioxidant balance may
serve an important role in a number of the processes of
inflammation and fibrosis (33).
GSH is an abundant endogenous antioxidant and a critical regulator
of oxidative stress (34,35). The GSH level and the ratio of GSH
to GSSG (GSH/GSSG) serve a role in the Nrf2-mediated antioxidative
stress (36). ROS activation in
oxidative stress is considered to be the key factor in inflammatory
amplification (37). The present
study demonstrated that MA significantly inhibited the protein
expression levels of Nrf2 and HO-1 in rat lungs, and that the GSH
level and the ratio of GSH/GSSG were decreased in the MA group,
accompanied by increasing ROS level in the lungs. The above indices
of antioxidative stress from MA were significantly reversed by
fluoxetine at 10 mg/kg. Taken together, it is suggested that
fluoxetine may alleviate pulmonary inflammation, and may be
associated with the inhibition of oxidative stress including
upregulation of the endogenous antioxidative factor GSH and the
downregulation of ROS levels in the lungs by the activation of the
Nrf2/HO-1 pathway.
In conclusion, long-term administration of MA
induces chronic pulmonary inflammation. Fluoxetine, as a SERT
inhibitor, alleviated the MA-induced lung inflammation in rats. The
potential protective mechanisms of fluoxetine may be associated
with suppressing oxidative stress through the SERT/p38 MAPK/Nrf2
pathways in rats.
Acknowledgements
The present study was funded by the National Natural
Science Foundation of China (grant no. 81503058) and the Natural
Science Foundation of Liaoning Province (grant no.
2014021,065).
Glossary
Abbreviations
Abbreviations:
|
MA
|
methamphetamine
|
|
5-HT
|
serotonin (5-hydroxytryptamine)
|
|
SERT
|
serotonin transporter
|
|
PAH
|
pulmonary arterial hypertension
|
|
ERK1/2
|
extracellular signal-regulated kinase
1/2
|
|
p38 MAPK
|
p38 mitogen activated protein
kinases
|
|
ROS
|
reactive oxygen species
|
|
Nrf2
|
nuclear factor erythroid 2-related
factor 2
|
|
HO-1
|
human heme oxygenase-1
|
|
IL-6
|
interleukin-6
|
|
TNF-α
|
tumor necrosis factor-α
|
|
GSH
|
reduced glutathione
|
|
GSSG
|
oxidized glutathione
|
References
|
1
|
Rawson RA: Current research on the
epidemiology, medical and psychiatric effects, and treatment of
methamphetamine use. J Food Drug Anal. 21:S77–S81. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Albertson TE, Derlet RW and Van Hoozen BE:
Methamphetamine and the expanding complications of amphetamines.
West J Med. 170:214–219. 1999.PubMed/NCBI
|
|
3
|
Volkow ND, Fowler JS, Wang GJ, Shumay E,
Telang F, Thanos PK and Alexoff D: Distribution and
pharmacokinetics of methamphetamine in the human body: Clinical
implications. PLoS One. 5:e152692010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Peerzada H, Gandhi JA, Guimaraes AJ,
Nosanchuk JD and Martinez LR: Methamphetamine administration
modifies leukocyte proliferation and cytokine production in murine
tissues. Immunobiology. 218:1063–1068. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ma J, Wan J, Meng J, Banerjee S,
Ramakrishnan S and Roy S: Methamphetamine induces autophagy as a
pro-survival response against apoptotic endothelial cell death
through the Kappa opioid receptor. Cell Death Dis. 5:e10992014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang Y, Liu M, Wang HM, Bai Y, Zhang XH,
Sun YX and Wang HL: Involvement of serotonin mechanism in
methamphetamine-induced chronic pulmonary toxicity in rats. Hum Exp
Toxicol. 32:736–746. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hollinger MA: Drug-induced lung toxicity.
Int J Toxicol. 12:31–47. 1993. View Article : Google Scholar
|
|
8
|
Kushnir-Sukhov NM, Gilfillan AM, Coleman
JW, Brown JM, Bruening S, Toth M and Metcalfe DD:
5-hydroxytryptamine induces mast cell adhesion and migration. J
Immunol. 177:6422–6432. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
de Las Casas-Engel M and Corbí AL, .
Serotonin modulation of macrophage polarization: Inflammation and
beyond. Adv Exp Med Biol. 824:89–115. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mann DA and Oakley F: Serotonin paracrine
signaling in tissue fibrosis. Biochim Biophys Acta. 1832:905–910.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tanaka T, Doe JM, Horstmann SA, Ahmad S,
Ahmad A, Min SJ, Reynolds PR, Suram S, Gaydos J, Burnham EL and
Vandivier RW: Neuroendocrine signaling via the serotonin
transporter regulates clearance of apoptotic cells. J Biol Chem.
289:10466–10475. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu M, Wang Y, Wang HM, Bai Y, Zhang XH,
Sun YX and Wang HL: Fluoxetine attenuates chronic
methamphetamine-induced pulmonary arterial remodelling: Possible
involvement of serotonin transporter and serotonin 1B receptor.
Basic Clin Pharmacol Toxicol. 112:77–82. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dempsie Y and MacLean MR: Pulmonary
hypertension: Therapeutic targets within the serotonin system. Br J
Pharmacol. 155:455–462. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Morrell NW, Adnot S, Archer SL, Dupuis J,
Jones PL, MacLean MR, McMurtry IF, Stenmark KR, Thistlethwaite PA,
Weissmann N, et al: Cellular and molecular basis of pulmonary
arterial hypertension. J Am Coll Cardiol. 54(1 Suppl): S20–S31.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dizeyi N, Hedlund P, Bjartell A, Tinzl M,
Austild-Taskén K and Abrahamsson PA: Serotonin activates MAP kinase
and PI3K/Akt signaling pathways in prostate cancer cell lines. Urol
Oncol. 29:436–445. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang Y, Han DD, Wang HM, Liu M, Zhang XH
and Wang HL: Downregulation of osteopontin is associated with
fluoxetine amelioration of monocrotaline-induced pulmonary
inflammation and vascular remodelling. Clin Exp Pharmacol Physiol.
38:365–372. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Singh M, Foster CR, Dalal S and Singh K:
Osteopontin: Role in extracellular matrix deposition and myocardial
remodeling post-MI. J Mol. Cell Cardiol. 48:538–543. 2010.
View Article : Google Scholar
|
|
18
|
Sahu BD, Mahesh Kumar J and Sistla R:
Baicalein, a bioflavonoid, prevents cisplatin-induced acute kidney
injury by up-regulating antioxidant defenses and down-regulating
the MAPKs and NF-κB pathways. PLoS One. 10:e01341392015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pandurangan AK, Mohebali N, Norhaizan ME
and Looi CY: Gallic acid attenuates dextran sulfate sodium-induced
experimental colitis in BALB/c mice. Drug Des Devel Ther.
9:3923–3934. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ryu J, Kwon MJ and Nam TJ: Nrf2 and NF-κB
signaling pathways contribute to porphyra-334-mediated inhibition
of UVA-induced inflammation in skin fibroblasts. Mar Drugs.
13:4721–4732. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Saetta M, Shiner RJ, Angus GE, Kim WD,
Wang NS, King M, Ghezzo H and Cosio MG: Destructive index: A
measurement of lung parenchymal destruction in smokers. Am Rev
Respir Dis. 131:764–769. 1985.PubMed/NCBI
|
|
22
|
Rincon M and Irvin CG: Role of IL-6 in
asthma and other inflammatory pulmonary diseases. Int J Biol Sci.
8:1281–1290. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lockett AD, Kimani S, Ddungu G, Wrenger S,
Tuder RM, Janciauskiene SM and Petrache I: α1-Antitrypsin modulates
lung endothelial cell inflammatory responses to TNF-α. Am J Respir
Cell Mol Biol. 49:143–150. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Stromps J, Fuchs P, Demir E, Grieb G,
Reuber K and Pallua N: Intraalveolar TNF-α in combined burn and
inhalation injury compared with intraalveolar interleukin-6. J Burn
Care Res. 36:e55–e61. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rothman RB and Baumann MH: Therapeutic and
adverse actions of serotonin transporter substrates. Pharmacol
Ther. 95:73–88. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cloëz-Tayarani I and Changeux JP: Nicotine
and serotonin in immune regulation and inflammatory processes: A
perspective. J Leukoc Biol. 81:599–606. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wells SM, Buford MC, Porter VM, Brunell
HL, Bunderson-Schelvan M, Nevin AB, Cardozo-Pelaez F and Holian A:
Role of the serotonergic system in reduced pulmonary function after
exposure to methamphetamine. Am J Respir Cell Mol Biol. 42:537–544.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tavoulari S, Forrest LR and Rudnick G:
Fluoxetine (Prozac) binding to serotonin transporter is modulated
by chloride and conformational changes. J Neurosci. 29:9635–9643.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rose'Meyer R: A review of the serotonin
transporter and prenatal cortisol in the development of autism
spectrum disorders. Mol Autism. 4:372013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bai Y, Wang HM, Liu M, Wang Y, Lian GC,
Zhang XH, Kang J and Wang HL: 4-Chloro-DL-phenylalanine protects
against monocrotaline-induced pulmonary vascular remodeling and
lung inflammation. Int J Mol Med. 33:373–382. 2014.PubMed/NCBI
|
|
31
|
Chen HH, Wang TC, Lee YC, Shen PT, Chang
JY, Yeh TK, Huang CH, Chang HH, Cheng SY, Lin CY, et al: Novel
Nrf2/ARE activator, trans-coniferylaldehyde, induces a
HO-1-mediated defense mechanism through a dual p38α/MAPKAPK-2 and
PK-N3 signaling pathway. Chem Res Toxicol. 28:1681–1692. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ishikawa T: Genetic polymorphism in the
NRF2 gene as a prognosis marker for cancer chemotherapy. Front
Genet. 5:3832014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ni S, Wang D, Qiu X, Pang L, Song Z and
Guo K: Bone marrow mesenchymal stem cells protect against
bleomycin-induced pulmonary fibrosis in rat by activating Nrf2
signaling. Int J Clin Exp Pathol. 8:7752–7761. 2015.PubMed/NCBI
|
|
34
|
Richie JP Jr, Nichenametla S, Neidig W,
Calcagnotto A, Haley JS, Schell TD and Muscat JE: Randomized
controlled trial of oral glutathione supplementation on body stores
of glutathione. Eur J Nutr. 54:251–263. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sinha-Hikim I, Shen R, Lee WN Paul, Crum
A, Vaziri ND and Norris KC: Effects of a novel cystine-based
glutathione precursor on oxidative stress in vascular smooth muscle
cells. Am J Physiol Cell Physiol. 299:C638–C642. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Qin T, Yin Y, Yu Q and Yang Q: Bursopentin
(BP5) protects dendritic cells from lipopolysaccharide-induced
oxidative stress for immunosuppression. PLoS One. 10:e01174772015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang AL, Niu Q, Shi N, Wang J, Jia XF,
Lian HF, Liu Z and Liu CX: Glutamine ameliorates intestinal
ischemia-reperfusion Injury in rats by activating the Nrf2/Are
signaling pathway. Int J Clin Exp Pathol. 8:7896–7904.
2015.PubMed/NCBI
|