Introduction
Stroke is the fourth major cause of morbidity and
remains a leading challenge to public health (1). Intracerebral hemorrhage (ICH) is a
life-threatening type of stroke that affects 2 million individuals
annually worldwide (2). The
symptoms, which differ in accordance to the site and spectrum of
brain damage, have the potential to be seriously disabling and may
lead to impaired motor and sensorimotor functions, along with
cognitive impairments (3). ICH is
characterized by extravasation of blood into the brain parenchyma,
leading to hematoma formation, concomitant brain edema and
secondary brain damage (4).
Previous studies regarding secondary brain damage have identified
possible treatments that target brain edema, blood-brain barrier
disruption, inflammation and the decomposition product of
erythrocytes (5–7). However, despite considerable progress
being made in animal models and preclinical research in recent
years, there are currently no available therapeutic strategies in
clinical practice for ICH (8).
Growth-associated protein-43 (GAP-43) is a protein
associated with neurite growth during development and regeneration,
which allows differentiating neurons to respond to extracellular
signals of the central nervous system (CNS) (9). A previous study demonstrated that
mice lacking this protein have a low survival rate during the early
postnatal period (10). In human
neuropathologies and their associated animal models, complex
alterations in GAP-43 expression are frequently detected,
suggesting axonal damage or attempts of regenerative axonal
sprouting. A previous in vivo study demonstrated the
requirement of GAP-43 in sustaining synaptic stability and
accelerating the initiation of axonal regrowth (11). In particular, one of the most
marked regenerative events following the onset of stroke is axonal
sprouting in the penumbra, which is accompanied by high expression
of GAP-43 (12–14). Therefore, it was hypothesized that
GAP-43 may be a prospective therapeutic target for the treatment of
ICH.
Bone marrow mesenchymal stem cells (BMSCs) are
multipotent cells with self-renewal capabilities, which are able to
differentiate into various cell types, including adipocytes,
osteoblasts, chondroblasts and neural cells (15–17).
In addition, it has previously been indicated that BMSCs may modify
the tissue microenvironment via secreting soluble factors to
rejuvenate or reinstate diseased cells (18). Such factors secreted from BMSCs
serve a considerable role in various respects of hematopoiesis and
are collectively termed trophic factors (19). Therefore, BMSC-induced reinstating
of dysfunctional tissues could result from secretion of these
trophic factors and/or their differentiation. Previous studies have
demonstrated that intraparenchymal transplantation of BMSCs can
exert anti-inflammatory and regenerative effects, and ameliorate
neurological deficits in numerous CNS diseases, including stroke
(20–22). However, the mechanisms by which
BMSC implantation provides neuroprotection under CNS pathological
circumstances are not well understood. Since transplantation of
stem cells is often associated with poor differentiation and
survival rates, utilization of conditioned medium (CM), which
includes trophic factors from stem cells, has emerged as an
alternative method (23,24). A recent study confirmed that CM
from human umbilical cord MSCs has considerable potential for
stroke treatment without requiring stem cell transplantation
(25). Therefore, the application
of stem cell-CM in place of direct transplantation of stem cells
may overcome the restrictions of current cell-based therapies.
However, it remains unknown whether the therapeutic action of BMSCs
is due to their differentiation or the trophic factors secreted
into their CM.
In the present study, BMSC-CM was tested as an
alternative to direct BMSC transplantation. To investigate the
effects of BMSC-CM, the alterations in expression of GAP-43,
phosphorylated-extracellular signal-regulated kinase (p-ERK1/2),
p-c-Jun N-terminal kinase (JNK) and inflammatory factors
[interleukin (IL)-1β, tumor necrosis factor (TNF)-α, IL-6 and
IL-10] in the perihematomal tissues were assessed. The mechanisms
underlying the effects of BMSC-CM were also examined, in order to
identify whether it was associated with, at least in part, the
secretion of neurotrophic factors that activate the ERK1/2
signaling pathway and thereby elevate GAP-43 expression.
Materials and methods
Animals
All rat experiments were performed in compliance
with the guidelines of the National Institutes of Health Guide
(2011) for the Care and Use of Laboratory Animals (Bethesda, MD,
USA). The present experimental procedures were approved by the
Hebei Medical University Experimental Ethics Committee
(Shijiazhuang, China). A total of 151 male Sprague-Dawley rats (8
weeks old; weight, 250–280 g) and 6 male Sprague-Dawley rats (3
weeks old; weight, 90–110 g) purchased from the Hebei United
University Experimental Animal Center (Tangshan, China) were used
in present study. Rats were anesthetized with 10% chloral hydrate
(100 mg/kg, Bio-Rad Laboratories, Inc., Shanghai, China). All rats
were maintained under a 12 h light/12 h dark cycle at 22–24°C with
ad libitum access to food and water. Great efforts were made to
minimize the number of animals used and alleviate animal
suffering.
Primary culture of BMSCs
BMSCs were generated from 6 male Sprague-Dawley rats
(3 weeks old; weight, 90–110 g). The medullary cavities of rat
femurs were flushed with Dulbecco's modified Eagle's medium (DMEM;
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) to collect
fresh bone marrow cells. Cells were centrifuged at 1,000 × g for 5
min. Purified cells were dispersed in cell culture flasks at 37°C
(Corning Life Sciences, Tewksbury, MA, USA), and were cultured in
DMEM supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at 37°C in an
atmosphere containing 5% CO2. After 48 h, non-adherent
cells were removed and fresh media was added. Media was replaced
every 3 days. After primary culturing, the cells were subcultured
at a density of 1×104 cells/cm2 and used for further
experimentation after three passages.
Preparation of BMSC-CM
BMSCs from three passages were cultured in culture
flasks. When cells reached 80% confluence, they were cultured in 10
ml serum-free DMEM/F12. CM from BMSCs was collected after 1 day and
centrifuged at 560 × g for 10 min at 37°C. After the supernatants
were re-centrifuged at 5,040 × g for 5 min at 37°C, the CM was
collected and used for further experimentation.
ICH model
The rat ICH model was established as described by a
previous study (26). Rats were
anesthetized with an intraperitoneal injection of chloral hydrate
(100 mg/kg body weight) and placed on stereotaxic apparatus. A hole
was drilled into the skull, and a needle was inserted into the
right basal ganglia under stereotactic guidance (coordinates: 0.2
mm anterior, 5.5 mm ventral, and 3.5 mm lateral to the midline). A
total of 100 µl autologous arterial blood was infused slowly (5
µl/min) with a microinfusion pump. Following the completion of the
infusion, the needle was left in place for 20 min prior to
withdrawal. Control rats received an infusion of 100 µl normal
saline. Bone wax was placed around the burr hole and all procedures
were conducted under aseptic conditions to avoid infection.
Experiment groups
Three experiments were performed as follows.
Experiment 1 was conducted to determine the time course of brain
water content and functional recovery. A total of 101 rats were
randomly assigned to four groups: The normal control group (n=8,
Sham); the ICH model group (n=31, ICH); the BMSC transplantation
(10 µl, tail vein injection immediately post-ICH) after ICH group
(n=31, BMSCs); and the BMSC-CM treatment (10 µl, tail vein
injection immediately post-IC) after ICH group (n=31, CM). A
forelimb-placing test, corner turn test and modified neurological
severity score (mNSS) evaluation were used to detect motor and
sensorimotor functions 1 day prior to and 1–7 days after surgery.
The time course variation of brain edema was evaluated by analysis
of brain water content 1–7 days following ICH. Cognitive impairment
of the rats was evaluated by Morris water maze test 7–10 days
following ICH.
Experiment 2 was conducted to identify the
neuroprotective mechanism of BMSC-CM. A total of 30 rats were
randomly divided into three groups: The sham-operated group (n=10,
Sham); the ICH model group (n=10, ICH); and the BMSC-CM treatment
after ICH group (n=10, CM). A total of 4 days after ICH or sham
operation, reverse transcription-polymerase chain reaction (RT-PCR)
and western blot analysis were used to detect the expression levels
of axonal regrowth-associated protein GAP-43, p-ERK, p-JNK and
inflammatory cytokines.
Experiment 3 was conducted to determine the
regulation of GAP-43 transcription and expression following BMSC-CM
treatment. A total of 20 ICH rats treated with BMSC-CM were
randomly divided into two groups: The vehicle group, which received
an equal volume of 5% dimethyl sulfoxide (DMSO; n=10, vehicle); and
the PD98059 group, which received 5 µg p-ERK1/2 inhibitor PD98059
(Bio-Rad Laboratories, Inc.) dissolved in 5% DMSO (n=10, PD98059).
The drug injection volume was 20 µl followed by a flush with 10 µl
0.9% saline via intrathecal injection 1 day after ICH.
Neurological function tests
Forelimb-placing test, corner turn test and an mNSS
evaluation were performed prior to surgery and 1–7 days after ICH
by an investigator who was blinded to the experimental groups. A
total of 15 rats (5 per group) were used for these tests.
Forelimb placing was scored using a
vibrissae-elicited forelimb-placing test. Independent testing of
each forelimb was induced by brushing the vibrissae ipsilateral to
that forelimb on the edge of a tabletop. Intact animals placed the
forelimb quickly onto the countertop. This task was repeated 10
times and the percentage of successful placing responses was
determined.
In the corner turn test, rats were allowed to
proceed into a 30° corner. To exit the corner, the rat could turn
either left or right, and the direction was recorded. This task was
repeated 10 times.
The mNSS evaluation is a composite of motor,
sensory, balance and reflex tests. Neurological function was graded
on a scale of 0–18 (0, normal score; 18, maximal deficit score)
with one point awarded for the exhibition of specific abnormal
behavior or for lack of a tested reflex. A greater impairment of
normal function results in a higher score.
Evaluation of brain edema
Brain edema was evaluated by analysis of brain water
content, as previously described (27). A total of 66 rats were used to
evaluate brain edema (Sham, 3; ICH, 21; BMSCs, 21; CM, 21). The
percentage of water in the tissues was calculated according to the
following formula: Brain water %=[(wet weight-dry weight)/wet
weight]x100.
Morris water maze test
The spatial learning and memory of the rats was
evaluated by Morris water maze, as described previously, 7–10 days
after ICH (28). A total of 20
rats were used for this test (10 per group). Each rat was allowed
to find the submerged platform within 60 sec. A total of 4 trials
were conducted on the first testing day (7 days post-ICH), which
was considered as a training procedure. Formal testing was then
conducted daily on days 8, 9 and 10. At 5 h after the final testing
on day 10, the platform was removed from the tank and a probe trial
was performed to assess the learning and memory performance of each
rat.
RT-PCR
Semi-quantitative PCR was conducted at 4 days
post-ICH. Tissues were homogenized on ice with TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) and total RNA was
extracted, according to the manufacturer's protocol. High-quality
RNA was reverse transcribed into cDNA with a Reverse Transcription
kit (Takara Biotechnology Co., Ltd., Dalian, China). GAP-43 and
GAPDH primers were used. GAP-43 sense:
5′-CATGAGAAGTATGACAACAGCCT-3′, GAP-43 antisense
5′-AGTCCTTCCACGATACCAAAGT-3′; GAPDH sense:
5′-CTGCAACCAAAATTCAGGCTA-3′, GAPDH antisense
5′-CATCAGCAACGGGAGCAT-3′. The PCR reaction system consisted of cDNA
(10 ng), primers (0.1 µM), deoxy-ribonucleoside triphosphate (1
mM), Taq DNA polymerase (5 U), 10X buffer (2.5 µl), and double
distilled water to a final volume of 25 µl. PCR amplification was
performed using SYBR Premix Ex Taq (Takara Biotechnology Co., Ltd.)
on an ABI StepOnePlus Real-time PCR system (Thermo Fisher
Scientific, Inc.). The PCR protocol was as follows: 95°C for 2 min,
40 cycles of 95°C for 10 sec and 60°C for 40 sec. For
semi-quantitative PCR, products were analyzed using agarose gel
electrophoresis and the corresponding optical density ratio was
quantified (optical density value of the specific gene/optical
density value of GAPDH).
Western blot analysis
Western blotting was conducted at 4 days post-ICH.
Rat perihematomal tissues were lysed in Tissue Protein Lysis
Solution (Thermo Fisher Scientific, Inc.) supplemented with 5%
Proteinase Inhibitor Cocktail (Sigma-Aldrich; Merck KGaA),
incubated on ice for 30 min, and centrifuged at 15,000 × g at
22–24°C for 15 min. Protein concentrations were determined using
bicinchoninic acid protein assay reagents (Nanjing Jiancheng
Bioengineering Institute, Nanjing, China). Samples were loaded (3
µl per line) onto 10% SDS-PAGE, and transferred to polyvinylidene
difluoride membranes for 60 min. Non-specific binding sites were
blocked with 5% bovine serum albumin (Bio-Rad Biotechnology, Inc.)
at 22–24°C for 1 h, then incubated with various antibodies: Rabbit
anti-rat GAP-43 (cat. no. sc10786), IL-1β (cat. no. sc7884), IL-6
(cat. no. sc7920), IL-10 (cat. no. sc984), TNF-α (cat. no. sc8301),
ERK (cat. no. sc292838), p-ERK (cat. no. sc101760), p-JNK (cat. no.
sc135642) and β-actin (cat. no. sc130657) polyclonal antibodies
(dilution, 1:1,000; Santa Cruz Biotechnology, Inc., Dallas, TX,
USA) overnight at 4°C. The membranes were then incubated with
secondary antibodies (cat. no. sc2955, dilution, 1:5,000; Santa
Cruz Biotechnology, Inc.) for 2 h at 4°C. The immunoreactive bands
were visualized using an enhanced chemiluminescence reagent
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). Blots were scanned
by densitometry, and integrated density of pixels was quantified
using Image Quant software (version, 5.2; Molecular Dynamics; GE
Healthcare Bio-Sciences, Pittsburgh, PA, USA).
Statistical analysis
Data are expressed as the mean ± standard error. All
tests were performed using SPSS software (version 17.0; SPSS, Inc.,
Chicago, IL, USA). Statistical significance was determined using
one-way analysis of variance, and the Student-Newman-Keuls post hoc
test was used to determine differences among the groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
Rat mortality
Only two of the total 151 rats succumbed during
experimentation. One rat did not survive transplantation of BMSCs
during experiment 1, and the other did not survive treatment with
the vehicle in experiment 3.
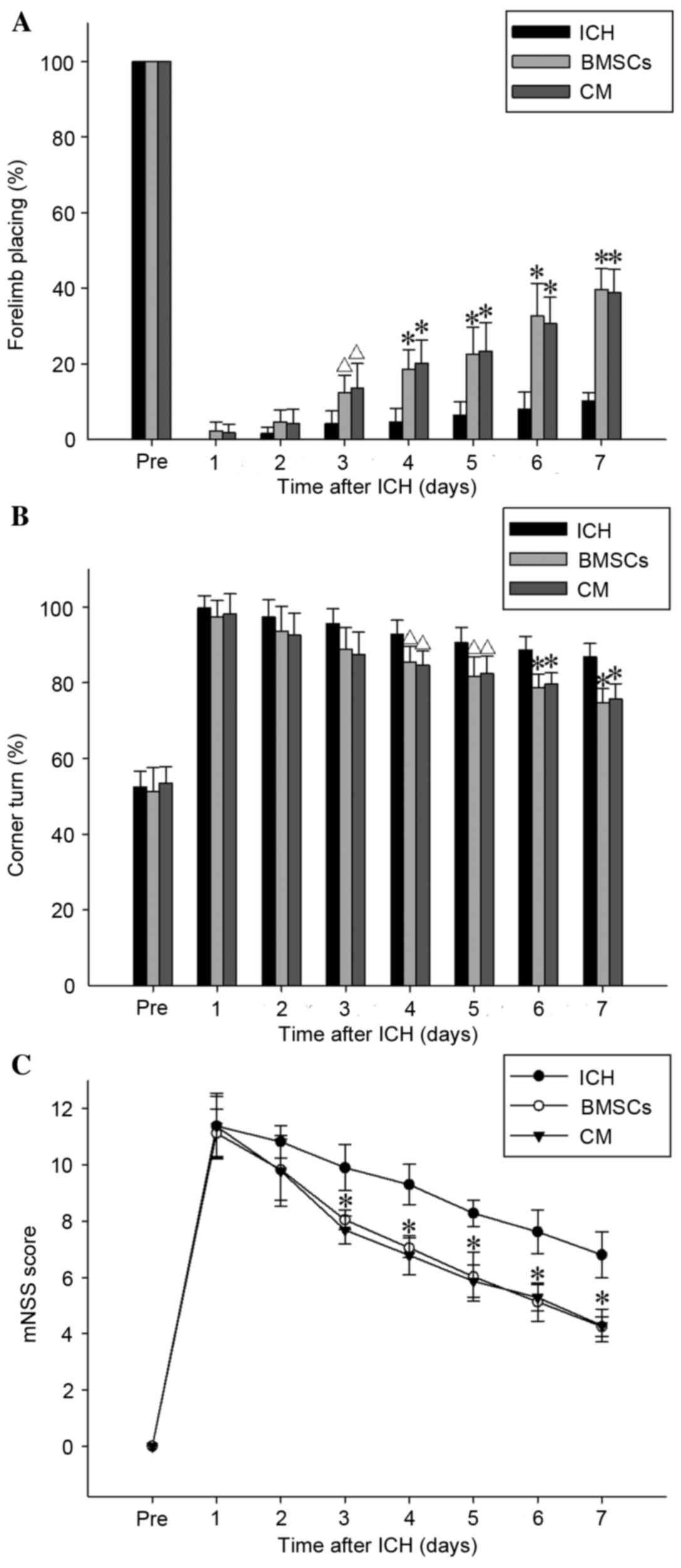
Transplantation of BMSCs and BMSC-CM
treatment ameliorates neurological deficit
Forelimb-placing and corner turn tests, and the mNSS
evaluation, were performed prior to and 1–7 days after ICH. As
presented in Fig. 1A, in the ICH
model group, there was little recovery of forelimb placing with
time. In the BMSC transplantation or BMSC-CM treatment groups,
there was a significant recovery compared with in the ICH model
group after 3–7 days (P<0.05 after 3 days; P<0.01 after 4–7
days). As depicted in Fig. 1B, in
the corner turn test, the percentage of turns to the right was
significantly decreased after 4–7 days in the BMSC transplantation
and BMSC-CM treatment groups compared with in the ICH group
(P<0.05 after 4–5 days; P<0.01 after 6–7 days). In addition,
Fig. 1C demonstrated that
transplantation of BMSCs or treatment with BMSC-CM resulted in
significantly improved mNSS scores compared with in the ICH group
after 4–7 days (P<0.01). However, there was no significant
difference (P>0.05) in motor and sensorimotor functions after
1–7 days between the BMSCs and BMSC-CM groups (Fig. 1C).
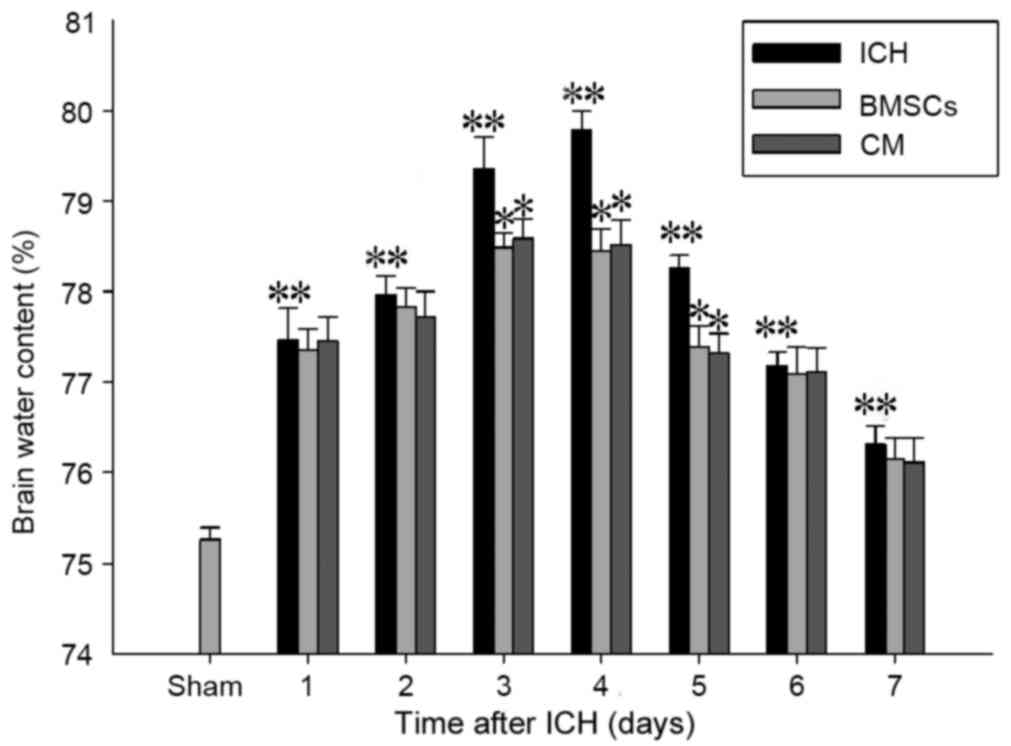
Transplantation of BMSCs and BMSC-CM
treatment attenuates the extent of brain edema
Analysis of brain water content was conducted 1–7
days post-ICH to identify the effects of BMSC transplantation and
BMSC-CM on cerebral edema. As demonstrated in Fig. 2, ICH caused a marked increase in
brain water content compared with the sham-operated group
(P<0.01). The mean brain water content of rat brains was
significantly increased in the ICH group compared with in the BMSCs
or BMSC-CM groups after 3–5 days (P<0.01). However, there was no
significant difference (P>0.05) in brain water content after 1–7
days between the BMSCs and BMSC-CM groups.
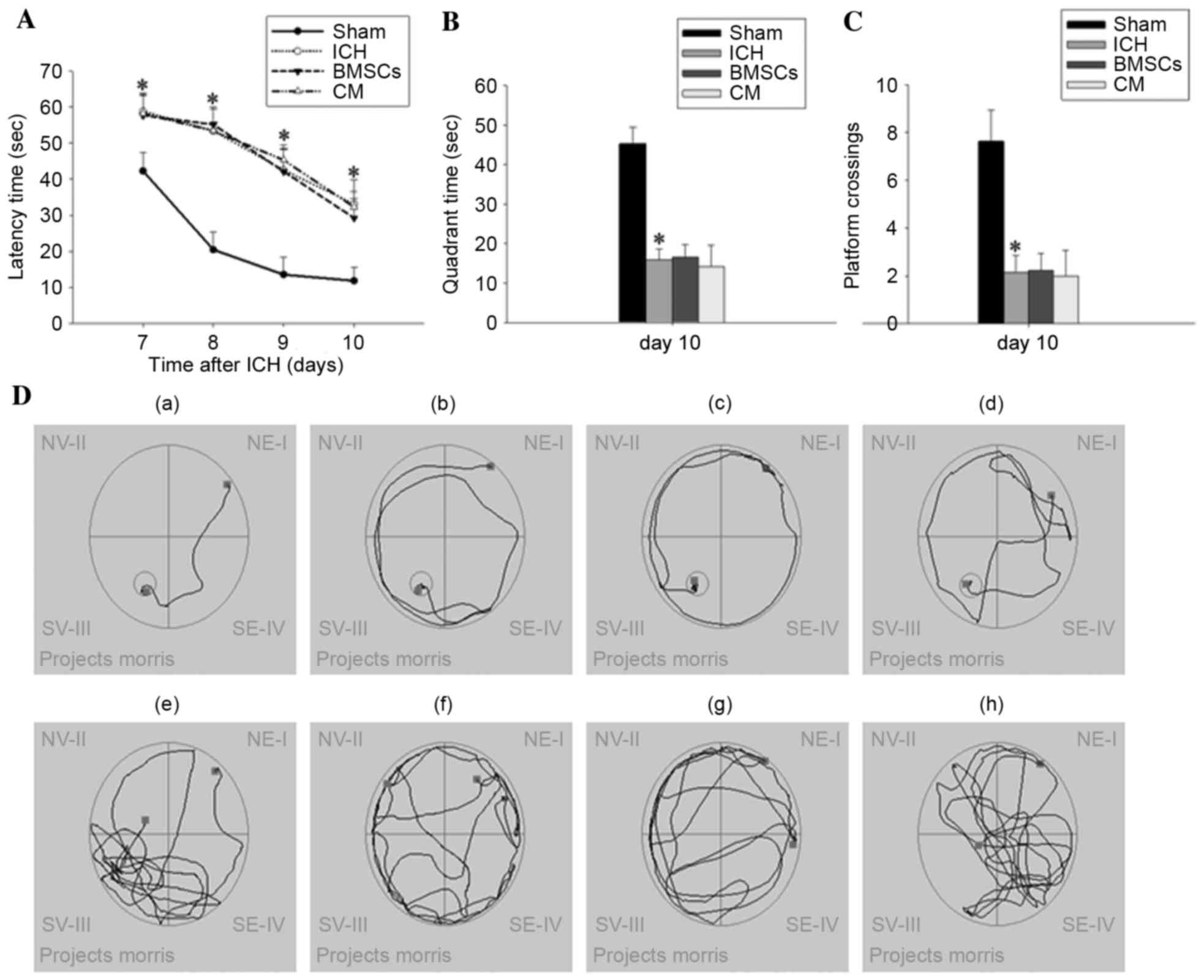
Transplantation of BMSCs or BMSC-CM
treatment has no effect on ICH-induced cognitive function
deficits
To determine whether BMSC transplantation and
BMSC-CM treatment led to improved cognitive functional outcome
post-ICH, a Morris water maze study was conducted 7–10 days after
ICH (Fig. 3). Fig. 3A illustrates the effects of BMSC
transplantation and BMSC-CM on latency time for finding the hidden
platform during latency trials. Latency time in the ICH groups was
progressively longer compared with in the sham group, in a
time-dependent manner; however, there was no significant difference
between the ICH, BMSCs and BMSC-CM treatment groups. Representative
trace diagrams indicating the latency time for finding the
submerged platform on day 10 are depicted in Fig. 3Da-d. In probe trials, which are
characterized by the removal of the hidden platform (Fig. 3B and C), ICH rats displayed a
reduced bias towards the goal quadrant, which previously contained
the platform. They spent less time in the goal quadrant and had
less platform crossings compared with their sham counterparts. In
addition, this phenomenon was not altered in the BMSCs and BMSC-CM
groups. Representative traces obtained during the specified probe
trials are depicted in Fig.
3De-h.
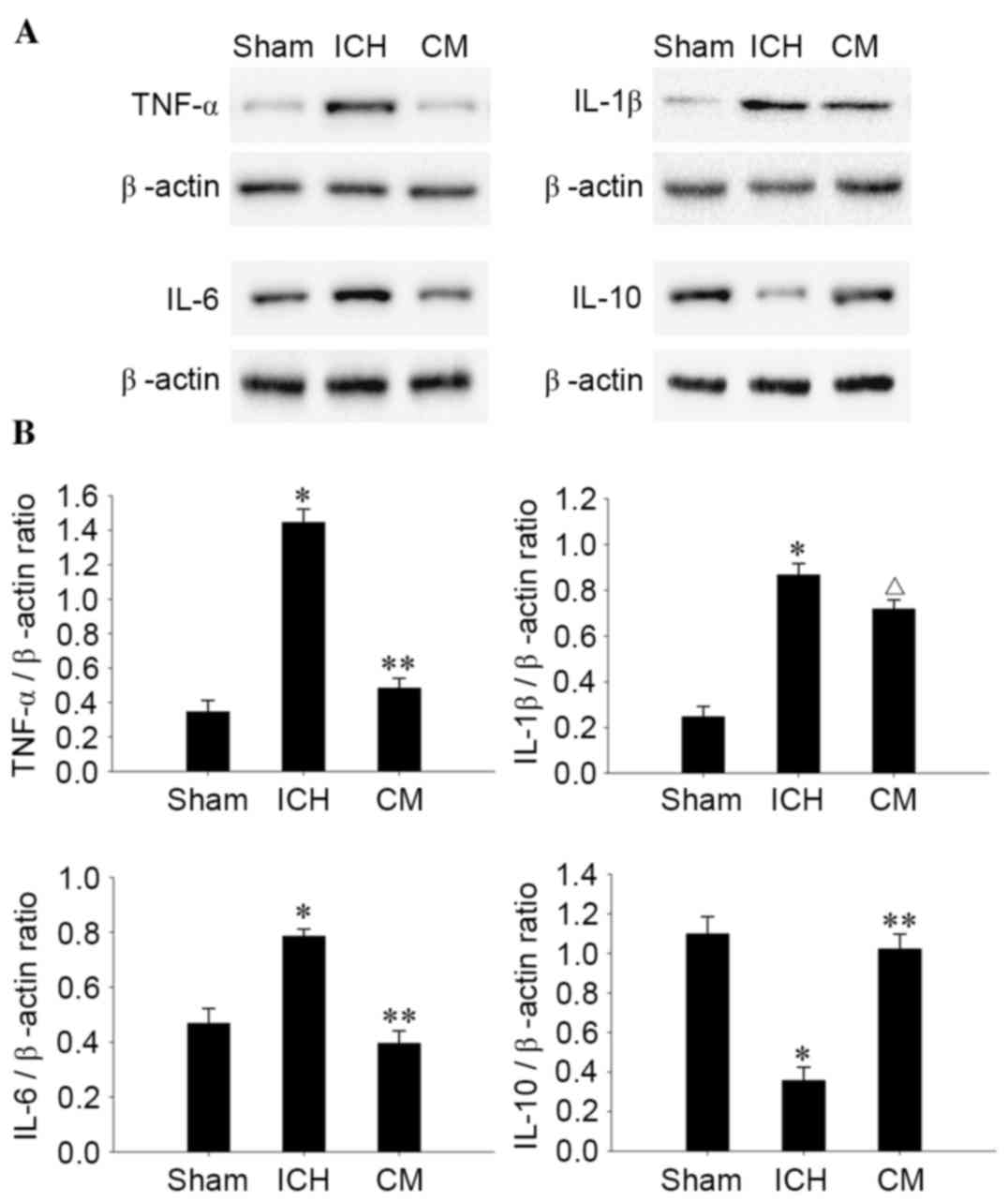
BMSC-CM treatment reduces the
expression of IL-1β, IL-6 and TNF-α, but not IL-10, in
perihematomal tissues
To explore the anti-inflammatory effects of BMSC-CM
treatment on ICH, the expression levels of inflammatory cytokines,
including TNF-α, IL-1β and IL-6, and the anti-inflammatory cytokine
IL-10, were measured 4 days after ICH in the perihematomal tissues
by western blot analysis. As demonstrated in Fig. 4, the protein expression levels of
IL-1β, IL-6 and TNF-α were significantly reduced in the brains of
BMSC-CM-treated rats compared with in the ICH group (IL-6 and
TNF-α, P<0.01; IL-1β, P<0.05). Conversely, the expression
levels of IL-10 were increased compared with in the ICH group
(P<0.01).
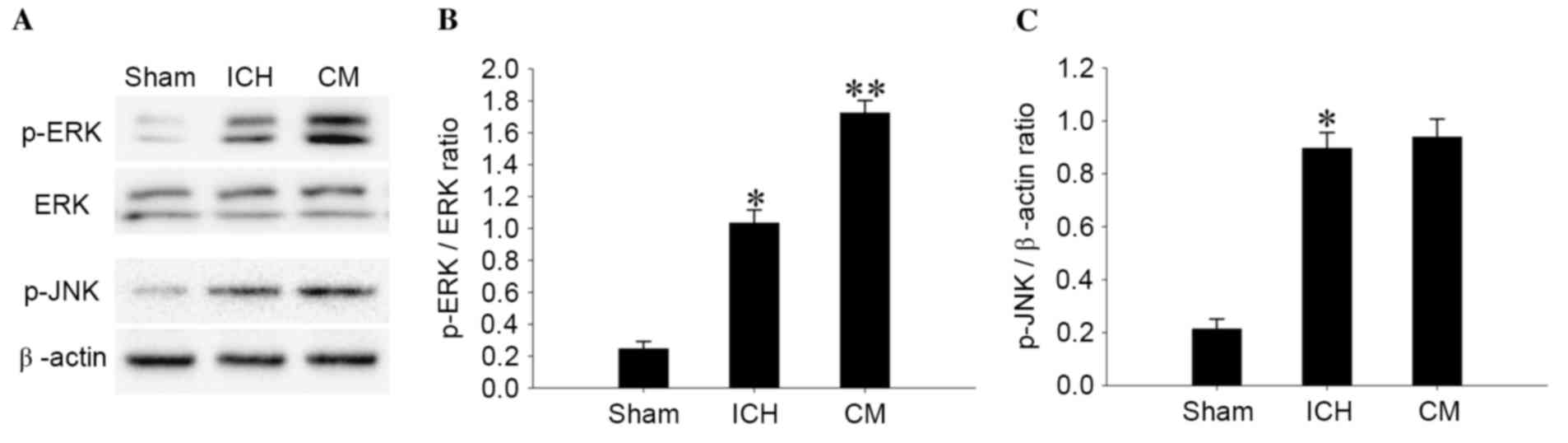
BMSC-CM treatment enhances
phosphorylation of ERK1/2 in perihematomal tissues
The present study explored whether BMSC-CM treatment
could enhance the activation of cell signaling. As demonstrated in
Fig. 5, phosphorylation of ERK1/2
and JNK was significantly elevated in the perihematomal region 4
days after ICH (P<0.01), whereas BMSC-CM treatment further
enhanced ERK1/2 phosphorylation (Fig.
5B, P<0.01). However, BMSC-CM treatment had no effect on JNK
activation in the perihematomal region 4 days after ICH (Fig. 5C, P>0.05).
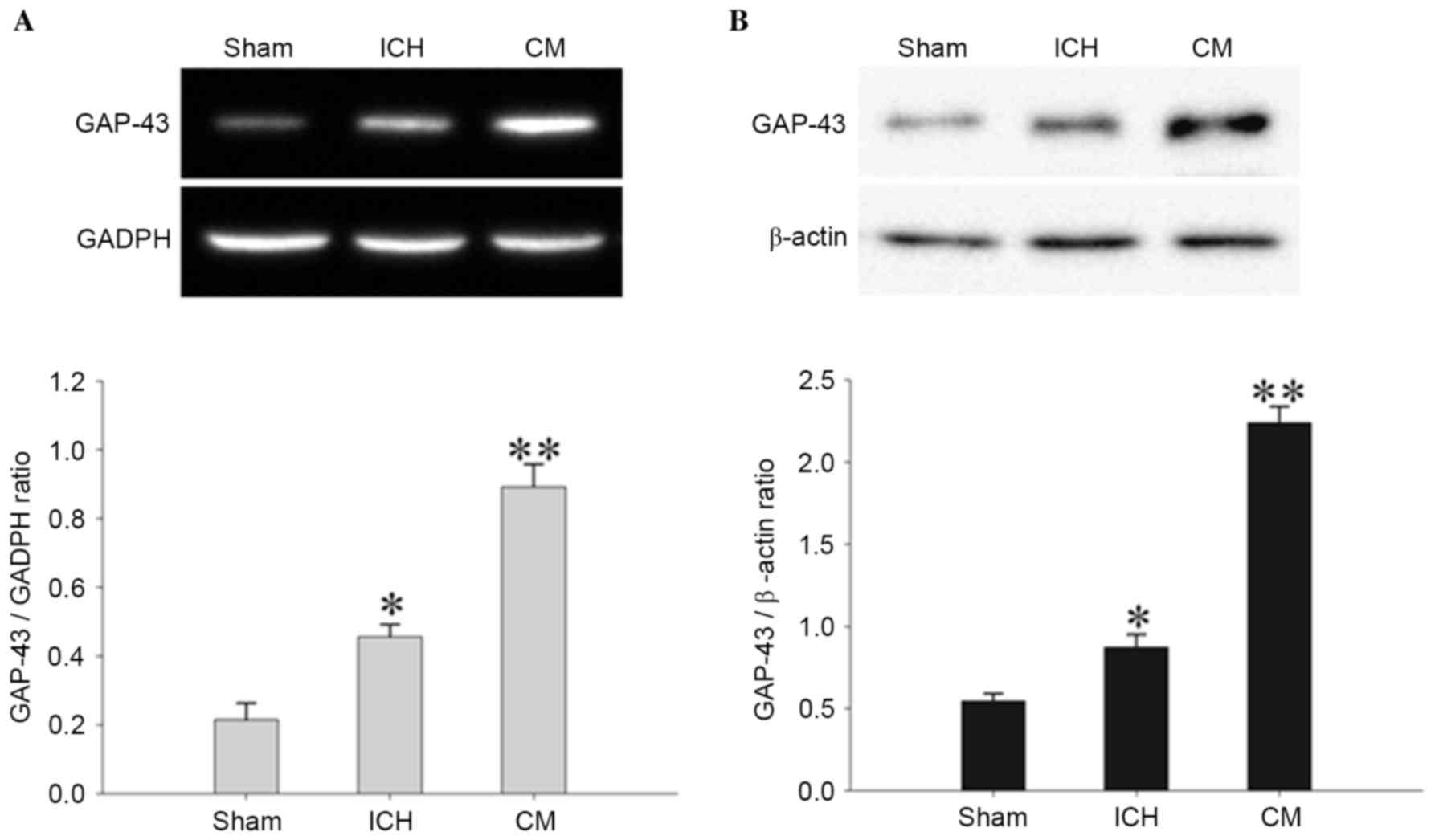
BMSC-CM rescues GAP-43 transcription
and expression in perihematomal tissues
To determine the effects of BMSC-CM on GAP-43
transcription and expression post-ICH, the expression levels of
GAP-43 were detected by RT-PCR and western blot analysis. RT-PCR
(Fig. 6A) demonstrated that the
mRNA expression levels of GAP-43 in the ICH group were markedly
increased 4 days following the operation (P<0.01). In the CM
group, BMSC-CM treatment significantly enhanced the mRNA expression
levels of GAP-43 compared with in the ICH rats (P<0.01). The
results of western blot analysis (Fig.
6B) indicated that ICH induced the elevation of GAP-43 protein
compared with the Sham group (P<0.01), and the expression of
GAP-43 was markedly reinforced following BMSC-CM treatment
(P<0.01).
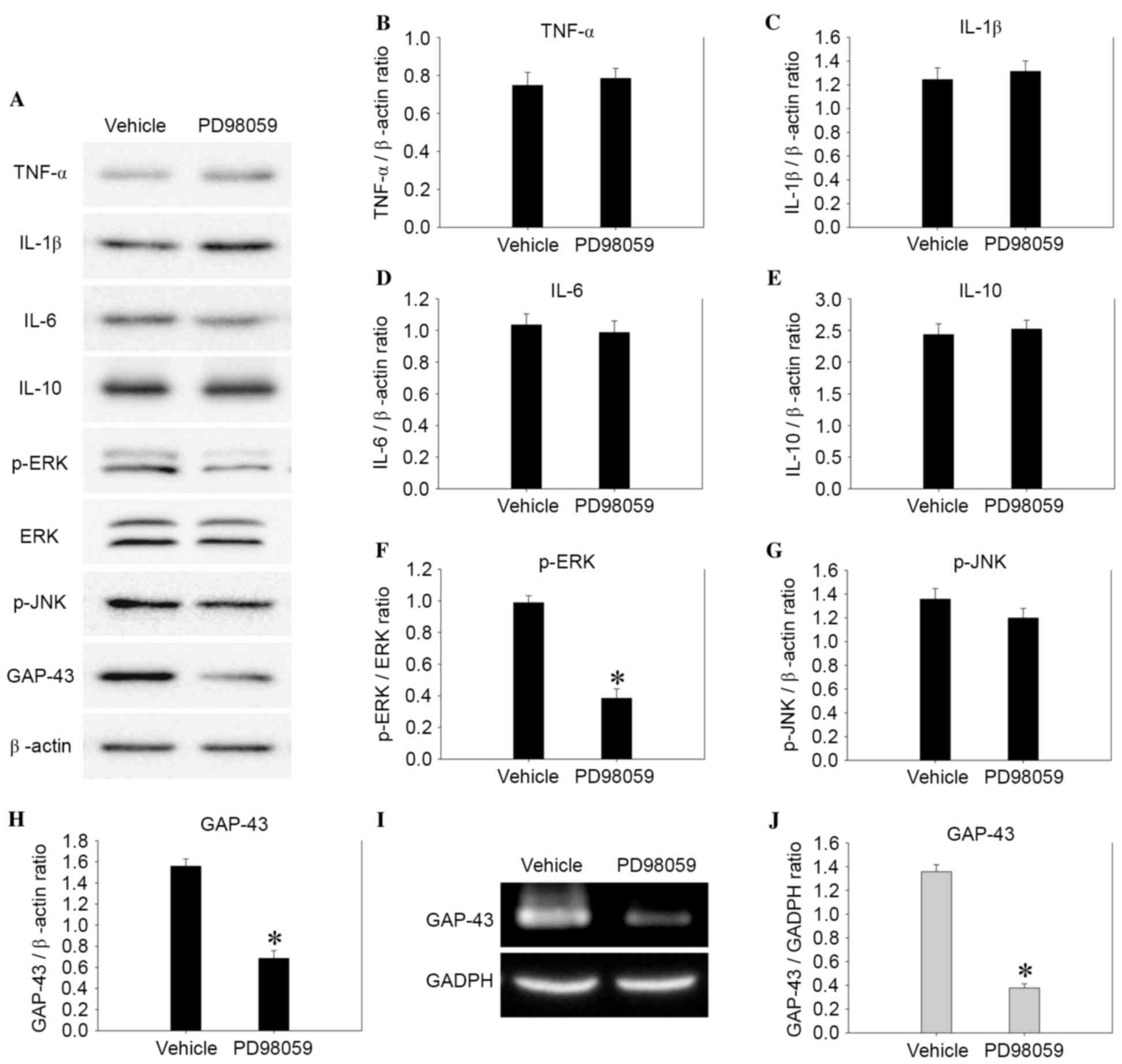
PD98059 reverses the elevation of
GAP-43 transcription and expression following BMSC-CM treatment in
perihematomal tissues
To further investigate whether the activation of
GAP-43 following BMSC-CM treatment was regulated by the ERK1/2
signaling pathway, a p-ERK1/2 inhibitor, PD98059, was intrathecally
administered. Fig. 7 depicts the
alterations in inflammatory cytokine level, cell signaling, and
GAP-43 transcription and expression following the administration of
PD98059 4 days after ICH. Notably, PD98059 inhibited the
phosphorylation of ERK1/2 (Fig.
7F, P<0.01). The mRNA and protein expression levels of
GAP-43 were also significantly decreased in the PD598059 group
compared with in the vehicle group (Fig. 7H and J, P<0.01). However, there
was no significant difference (P>0.05) in the expression of
inflammatory cytokines between the vehicle and PD98059 treatment
groups.
Discussion
The present study demonstrated that intraparenchymal
treatment with BMSC-CM 1 day after ICH exerted neuroprotective
effects in rats. Treatment with BMSC-CM attenuated post-ICH
cerebral edema after 3–5 days and improved the motor functions of
ICH rats after 3–7 days. Furthermore, the current study
investigated the mechanisms underlying the protective effects of
BMSC-CM on ICH. At the molecular level, the expression levels of
GAP-43 and inflammatory cytokines in the perihematomal tissues were
detected 4 days after ICH. Treatment with BMSC-CM resulted in a
marked elevation in the expression of GAP-43 and IL-10, as well as
a significant reduction in the expression levels of IL-1β, TNF-α
and IL-6. These findings suggested that BMSC-CM acts by inhibiting
neuroinflammatory activation, and regulating the expression of
GAP-43. Following administration of the p-ERK1/2 inhibitor,
PD98059, in BMSC-CM-treated rats, the mRNA and protein expression
levels of GAP-43 were significantly decreased, suggesting that the
ERK1/2 signaling pathway is essential for GAP-43 expression. A
previous study reported that CM from human adipose-derived MSCs
mediated protection in neurons following glutamate excitotoxicity
by regulating energy metabolism and GAP-43 expression (29). In addition, a recent in vivo
study demonstrated that CM from human umbilical cord MSCs
contributed to vascular remodeling in the ischemic brain, which
serves an important role in functional outcome in a rat model of
stroke (25). These findings,
together with the observations of the present study, may aid to
improve current understanding regarding stem cell-CM-mediated
neuroprotection in neurological disorders.
ICH has a complex pathology with numerous diverse
and incompletely clarified etiologies and prognoses. Despite
various promising preclinical experiments, including the evaluation
of anti-inflammatory, anti-hypertensive and neuroprotective drugs,
only symptomatic treatments are currently effective (8). In this context, the search for novel
alternatives is a necessity for ICH treatment. A previous study
regarding stem cell transplantation in ICH animal models
demonstrated that transplantation attenuated hematoma and improved
neurological behaviors (30).
Among miscellaneous stem cells, neural stem cells (NSCs) and MSCs
are the most widely used and are considered promising in the
treatment of ICH. As a result of the distinct capacity of NSCs to
differentiate into functional neural cells, the neuroprotective
effect may result from transplantation of NSCs replacing damaged
neurons. Conversely, the efficacy of MSCs, as demonstrated in
models of neurological disease, may be due to the cells' capacity
to secrete various neurotrophins, such as brain-derived
neurotrophic factor (BDNF), nerve growth factor (NGF) and vascular
endothelial growth factor (VEGF), all of which potentially
contribute to functional recovery following ICH (31–33).
In particular, BMSCs have been regarded as candidates for cell
therapy as they can express an extensive ability for expansion
in vitro and can be readily procured by a simple bone marrow
aspiration (34). The present
study indicated that the neuroprotective effects of BMSC
implantation and BMSC-CM treatment have no significant difference
to each other and both prevent post-ICH cerebral edema and improve
neurological function. Therefore, the observed beneficial effects
of BMSCs are mediated by soluble factors secreted by these BMSCs
into CM.
The present results also suggested that BMSC-CM was
able to exert anti-inflammatory effects in perihematomal tissues of
ICH rats. Previous studies have focused on therapies for secondary
brain injury following ICH and indicated that inflammatory events
within the first 7 days are the crucial contributor to secondary
brain injury (20,35). These findings indicated that
attenuation of inflammatory factors may improve neurological
recovery following ICH. It has been demonstrated that inflammatory
cytokines, including TNF-α, IL-1β and IL-6, produce cerebral edema
and brain damage post-ICH by recruiting lymphocyte and neutrophil
infiltration into perihematomal tissues (20,36,37).
Conversely, the anti-inflammatory cytokine IL-10 exerts beneficial
effects against brain damages post-ICH (38). A previous study indicated that
compared with the pathophysiology of traumatic brain injury and
cerebral ischemia, the pathophysiology of ICH was not associated
with the expression of TNF-α, IL-1β and IL-6 in the acute phase (1
h after ICH) in the perihematomal region (35). In the present study, 4 days after
ICH, TNF-α, IL-1β and IL-6 levels were decreased, whereas IL-10
expression was significantly elevated in the perihematomal tissues
of BMSC-CM-treated rats. These results provided evidence to suggest
that BMSC-CM is able to effectively regulate the production and
secretion of inflammatory cytokines in the perihematomal regions of
the ICH-injured brain.
The present study focused on the association between
ERK1/2 signaling and GAP-43 transcription and expression. It
appears that the phosphorylation of ERK1/2 is a major mechanism in
BMSC-CM-induced GAP-43 expression in ICH rats. A previous study
suggested that GAP-43 was required for BDNF-induced ERK1/2
activation (39). The findings of
the present study, together with previous hypotheses, highlight the
possibility that heterogeneous factors, including BDNF, NGF and
VEGF, are involved in a similar signaling pathway leading to ERK1/2
phosphorylation (40). To the best
of our knowledge, the present study is the first to suggest that
the ERK1/2-GAP-43 signaling pathway may be considered a potential
novel application target of BMSC-CM for the treatment of CNS
diseases.
In addition to motor deficits post-ICH, learning and
memory impairments may evolve that also adversely impair lifestyle
(41). Previous studies have
confirmed the role of the striatum in cognition, and extravasation
of blood into the caudate putamen of rats produces a learning and
memory impairment (42,43). In the present study, the rat
cognitive test was conducted 7–10 days after ICH. BMSC
transplantation and BMSC-CM treatment produced no significant
effect on the learning and memory functions of ICH rats. However,
it remains uncertain as to whether BMSC or BMSC-CM treatment has
beneficial effects on learning and memory functions in the ICH
model used in the present study, since the study was conducted
during a relatively short time period. In addition, the majority of
patients with ICH are elderly; however, the rats used in the
present study were selected from a fixed age group with a high
proportion of young rats. Therefore, future studies are required to
explore the long-term effects of BMSC-CM-mediated neuroprotection
in aged animal models.
In conclusion, treatment with BMSC-CM may be
considered an alternative to direct BMSC transplantation in a rat
model of ICH. The mechanism underlying BMSC-CM-mediated
neuroprotection may involve anti-inflammatory effects, and
activation of GAP-43 transcription and expression via ERK1/2
phosphorylation.
Acknowledgements
The present study was supported by a grant from the
Natural Science Foundation of Hebei Province (grant no.
H2014105079).
Glossary
Abbreviations
Abbreviations:
|
ICH
|
intracerebral hemorrhage
|
|
GAP-43
|
growth- associated protein-43
|
|
CNS
|
central nervous system
|
|
BMSCs
|
bone marrow mesenchymal stem cells
|
|
CM
|
conditioned medium
|
|
mNSS
|
modified neurological severity
score
|
|
DMSO
|
dimethyl sulfoxide
|
|
NSCs
|
neural stem cells
|
|
MSCs
|
mesenchymal stem cells
|
|
BDNF
|
brain-derived neurotrophic factor
|
|
NGF
|
nerve growth factor
|
|
VEGF
|
vascular endothelial growth factor
|
References
|
1
|
Towfighi A and Saver JL: Stroke declines
from third to fourth leading cause of death in the United States:
Historical perspective and challenges ahead. Stroke. 42:2351–2355.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Qureshi AI, Mendelow AD and Hanley DF:
Intracerebral haemorrhage. Lancet. 373:1632–1644. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cheon SH: The effect of a skilled reaching
task on hippocampal plasticity after intracerebral hemorrhage in
adult rats. J Phys Ther Sci. 27:131–133. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lok J, Leung W, Murphy S, Butler W,
Noviski N and Lo EH: Intracranial hemorrhage: Mechanisms of
secondary brain injury. Acta Neurochir Suppl. 111:63–69. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wasserman J and Schlichter L: Neuron death
and inflammation in a rat model of intracerebral hemorrhage:
Effects of delayed minocycline treatment. Brain Res. 1136:208–218.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kidwell CS, Latour LL, Hsia AW, Merino JC,
Burgess RE, Copenhaver BR, Castle A and Warach S: Demonstration of
blood-brain barrier disruption in humans with primary intracerebral
hemorrhage. Stroke. 38:4642007.
|
|
7
|
Xi G, Keep RF and Hoff JT: Erythrocytes
and delayed brain edema formation following intracerebral
hemorrhage in rats. J Neurosurg. 89:991–996. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Katsuki H: Exploring neuroprotective drug
therapies for intracerebral hemorrhage. J Pharmacol Sci.
114:366–378. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Meiri KF, Pfenninger KH and Willard MB:
Growth-associated protein, GAP-43, a polypeptide that is induced
when neurons extend axons, is a component of growth cones and
corresponds to pp46, a major polypeptide of a subcellular fraction
enriched in growth cones. Proc Natl Acad Sci USA. 83:3537–3541.
1986. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Maier DL, Mani S, Donovan SL, Soppet D,
Tessarollo L, McCasland JS and Meiri KF: Disrupted cortical map and
absence of cortical barrels in growth-associated protein (GAP)-43
knockout mice. Proc Natl Acad Sci USA. 96:9397–9402. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mascaro AL Allegra, Cesare P, Sacconi L,
Grasselli G, Mandolesi G, Maco B, Knott GW, Huang L, De Paola V and
Pavone FS: In vivo single branch axotomy induces GAP-43-dependent
sprouting and synaptic remodeling in cerebellar cortex. Proc Natl
Acad Sci USA. 110:10824–10829. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Carmichael ST, Archibeque I, Luke L, Nolan
T, Momiy J and Li S: Growth-associated gene expression after
stroke: Evidence for a growth-promoting region in peri-infarct
cortex. Exp Neurol. 193:291–311. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Stroemer RP, Kent TA and Hulsebosch CE:
Acute increase in expression of growth associated protein GAP-43
following cortical ischemia in rat. Neurosci Lett. 162:51–54. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gorup D, Bohaček I, Miličević T, Pochet R,
Mitrečić D, Križ J and Gajović S: Increased expression and
colocalization of GAP43 and CASP3 after brain ischemic lesion in
mouse. Neurosci Lett. 597:176–182. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ashton BA, Allen TD, Howlett CR, Eaglesom
CC, Hattori A and Owen M: Formation of bone and cartilage by marrow
stromal cells in diffusion chambers in Vivo. Clin Orthop Relat Res.
151:294–307. 1980.
|
|
16
|
Pereira RF, Halford KW, O'Hara MD, Leeper
DB, Sokolov BP, Pollard MD, Bagasra O and Prockop DJ: Cultured
adherent cells from marrow can serve as long-lasting precursor
cells for bone, cartilage, and lung in irradiated mice. Proc Natl
Acad Sci USA. 92:4857–4861. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen J, Li Y, Wang L, Zhang Z, Lu D, Lu M
and Chopp M: Therapeutic benefit of intravenous administration of
bone marrow stromal cells after cerebral ischemia in rats. Stroke.
32:1005–1011. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Caplan AI and Dennis JE: Mesenchymal stem
cells as trophic mediators. J Cell Biochem. 98:1076–1084. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Majumdar MK, Thiede MA, Haynesworth SE,
Bruder SP and Gerson SL: Human marrow-derived mesenchymal stem
cells (MSCs) express hematopoietic cytokines and support long-term
hematopoiesis when differentiated toward stromal and osteogenic
lineages. J Hematother Stem Cell Res. 9:841–848. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee ST, Chu K, Jung KH, Kim SJ, Kim DH,
Kang KM, Hong NH, Kim JH, Ban JJ, Park HK, et al: Anti-inflammatory
mechanism of intravascular neural stem cell transplantation in
haemorrhagic stroke. Brain. 131:616–629. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Daan VP, Biju P, Cho CH, Berthiaume F,
Nahmias Y, Tilles AW and Yarmush ML: Mesenchymal stem cell-derived
molecules directly modulate hepatocellular death and regeneration
in vitro and in vivo. Hepatology. 47:1634–1643. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lda S Meirelles, Fontes AM, Covas DT and
Caplan AI: Mechanisms involved in the therapeutic properties of
mesenchymal stem cells. Cytokine Growth Factor Rev. 20:419–427.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen L, Tredget EE, Wu PY and Wu Y:
Paracrine factors of mesenchymal stem cells recruit macrophages and
endothelial lineage cells and enhance wound healing. PLoS One.
3:e18862008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li X, Luo Q, Sun J and Song G: Conditioned
medium from mesenchymal stem cells enhances the migration of
hepatoma cells through CXCR4 up-regulation and F-actin remodeling.
Biotechnol Lett. 37:511–521. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhao Q, Hu J, Xiang J, Gu Y, Jin P, Hua F,
Zhang Z, Liu Y, Zan K, Zhang Z, et al: Intranasal administration of
human umbilical cord mesenchymal stem cells-conditioned medium
enhances vascular remodeling after stroke. Brain Res. 1624:489–496.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Teng W, Wang L, Xue W and Guan C:
Activation of TLR4-mediated NFkappaB signaling in hemorrhagic brain
in rats. Mediators Inflamm. 2009:4732762009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cui C, Cui Y, Gao J, Sun L, Wang Y, Wang
K, Li R, Tian Y, Song S and Cui J: Neuroprotective effect of
ceftriaxone in a rat model of traumatic brain injury. Neurol Sci.
35:695–700. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Raz L, Zhang QG, Zhou CF, Han D, Gulati P,
Yang LC, Yang F, Wang RM and Brann DW: Role of Rac1 GTPase in NADPH
oxidase activation and cognitive impairment following cerebral
ischemia in the rat. PLoS One. 5:e126062010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Peng H, Liang Z, Piao H, Ji X, Wang Y, Liu
Y, Liu R and Liu J: Conditioned medium of human adipose-derived
mesenchymal stem cells mediates protection in neurons following
glutamate excitotoxicity by regulating energy metabolism and GAP-43
expression. Metab Brain Dis. 29:193–205. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hu Y, Liu N, Zhang P, Pan C, Zhang Y, Tang
Y, Deng H, Aimaiti M, Zhang Y, Zhou H, et al: Preclinical studies
of stem cell transplantation in intracerebral hemorrhage: A
systemic review and meta-analysis. Mol Neurobiol. 53:5269–5277.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Seyfried D, Ding J, Han Y, Li Y, Chen J
and Chopp M: Effects of intravenous administration of human bone
marrow stromal cells after intracerebral hemorrhage in rats. J
Neurosurg. 104:313–318. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li Y, Chen J, Chen XG, Wang L, Gautam SC,
Xu YX, Katakowski M, Zhang LJ, Lu M, Janakiraman N and Chopp M:
Human marrow stromal cell therapy for stroke in rat: Neurotrophins
and functional recovery. Neurology. 59:514–523. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gerdoni E, Gallo B, Casazza S, Musio S,
Bonanni I, Pedemonte E, Mantegazza R, Frassoni F, Mancardi G,
Pedotti R and Uccelli A: Mesenchymal stem cells effectively
modulate pathogenic immune response in experimental autoimmune
encephalomyelitis. Ann Neurol. 61:219–227. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yan C, Lian X, Dai Y, Wang X, Qu P, White
A, Qin Y and Du H: Gene delivery by the hSP-B promoter to lung
alveolar type II epithelial cells in LAL-knockout mice through bone
marrow mesenchymal stem cells. Gene Ther. 14:1461–1470. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee IN, Cheng WC, Chung CY, Lee MH, Lin
MH, Kuo CH, Weng HH and Yang JT: Dexamethasone reduces brain cell
apoptosis and inhibits inflammatory response in rats with
intracerebral hemorrhage. J Neurosci Res. 93:178–188. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ziai WC: Hematology and inflammatory
signaling of intracerebral hemorrhage. Stroke. 44(6 Suppl 1):
S74–S78. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Aronowski J and Zhao X: Molecular
pathophysiology of cerebral hemorrhage: Secondary brain injury.
Stroke. 42:1781–1786. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jiang Y, Wei N, Lu T, Zhu J, Xu G and Liu
X: Intranasal brain-derived neurotrophic factor protects brain from
ischemic insult via modulating local inflammation in rats.
Neuroscience. 172:398–405. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gupta SK, Mishra R, Kusum S, Spedding M,
Meiri KF, Gressens P and Mani S: GAP-43 is essential for the
neurotrophic effects of BDNF and positive AMPA receptor modulator
S18986. Cell Death Differ. 16:624–637. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Matsuoka Y and Yang J: Selective
inhibition of ERK1/2 blocks NGF to BDNF signaling and suppresses
the development of and reverses already established pain behavior
in rats. Neuroscience. 206:224–236. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nys GM, van Zandvoort MJ, De Kort PL,
Jansen BP, deHaan EH and Kappelle LJ: Cognitive disorders in acute
stroke: Prevalence and clinical determinants. Cerebrovasc Dis.
23:408–416. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lekic T, Hartman R, Rojas H, Manaenko A,
Chen W, Ayer R, Tang J and Zhang JH: Protective effect of melatonin
upon neuropathology, striatal function, and memory ability after
intracerebral hemorrhage in rats. J Neurotrauma. 27:627–637. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ragozzino ME: The contribution of the
medial prefrontal cortex, orbitofrontal cortex and dorsomedial
striatum to behavioral flexibility. Ann N Y Acad Sci. 1121:355–375.
2007. View Article : Google Scholar : PubMed/NCBI
|