Introduction
In recent years, the toxicity of traditional Chinese
medicine, which has attracted increasing attention, restricts it
development and modernization. Croton tiglium L., as one of
the most poisonous traditional Chinese medicines, is officially
prescribed as the dried mature seeds of the Euphorbiaceae C.
tiglium L. (1). C.
tiglium L. is widely distributed in tropical and subtropical
zones, including India, New Guinea, Java, Indonesia, China and the
Philippine islands (2). As a
traditional herbal medicine, C. tiglium L. has been
primarily used to treat indigestion, abdominal distension, visceral
pain, constipation and other gastrointestinal disorders (3,4).
However, it also causes severe gastrointestinal syndrome and even
mortality following ingestion (5).
Although the use of C. tiglium L. has a long history, its
toxicity remains largely unknown. A preliminary study attributed
its toxicity to irritating oils and toxic proteins (6). Oral medication of croton oil is able
to cause gastrointestinal edema and diarrhea (7). The toxicity of croton proteins is
typically evaluated by intraperitoneal injection; however, C.
tiglium L. is orally administrated in clinical practice.
Therefore, evaluating the toxicity of croton proteins by
intraperitoneal injection is not sufficiently accurate. Chen et
al (8) demonstrated that edema
and diarrhea of the digestive tract were primarily caused by the
inflammatory response.
Inflammation is one of the most important protective
defensive reactions to tissue injury. Once the body is stimulated
by chemical drugs, appropriate inflammatory responses are activated
to maintain homeostasis (9,10).
In response to stimulation in the process of inflammation,
macrophages serve a key role as defending cells and release large
amounts of inflammatory cytokines (11,12).
Simultaneously, the injured tissues or cells release inflammatory
mediator prostaglandin E2 (PGE2) and a large amount of
proinflammatory cytokines, including tumor necrosis factor (TNF)-α
and interleukin (IL)-1β, in order to maintain balance (13,14).
The release of proinflammatory cytokines is associated with the
activation of cellular signaling pathways (15). The mitogen-activated protein kinase
(MAPK) signaling pathways, as key signaling molecules, serve an
important role in the inflammatory response, mediating the
generation of cytokines (16). The
MAPK family primarily comprises p38-MAPK, extracellular
signal-regulated kinase (ERK)1/2 and c-Jun N-terminal kinases
(JNKs), therefore the three primary pathways should be studied to
analyze the inflammatory responses (17,18).
In the present study, the toxic targets of proteins from C.
tiglium L. were determined by oral medication and
intraperitoneal injection. The inflammatory response was evaluated
by intraperitoneal injection and in RAW264.7 macrophages, and the
proinflammatory effects of crude protein on the release of PGE2
were evaluated through intraperitoneal injection and production of
proinflammatory cytokines in macrophages. The inflammatory response
induced by crude protein was associated with MAPK signaling.
Materials and methods
Materials
C. tiglium L. was collected from Sichuan
province (China). The PGE2 kit was purchased from Nanjing Jiancheng
Bioengineering Institute (Nanjing, China).
Dulbecco's modified Eagle's medium (DMEM), fetal
bovine serum (FBS), penicillin and streptomycin were obtained from
Gibco (Thermo Fisher Scientific, Inc.). Primary antibodies,
including anti-phosphorylated (p)-ERK (cat. no. 9101S), anti-ERK
(cat. no. 9102S), anti-p-JNK (cat. no. 9251S), anti-JNK (cat. no.
9252S), anti-p-p38 (cat. no. 9211S), and anti-p38 antibodies (cat.
no. 9212S), were obtained from Cell Signaling Technology, Inc.
(Danvers, MA, USA). Anti-GAPDH (cat. no. YFPA1059) was obtained
from Yi Fei Xue Biotech Co., Ltd. (Nanjing, China). Goat
anti-rabbit secondary antibodies (cat. no. 10285-1-AP) were
purchased from ProteinTech Group, Inc. (Chicago, IL, USA).
Animals
A total of 230 ICR mice, (115 female and 115 male,
aged 8–10 weeks old; weight, 18–22 g) were purchased from Shanghai
SLAC Laboratory Animal Co., Ltd. (Shanghai, China). The mice were
separated by sex in plastic cages with stainless steel mesh lids in
a ventilated room. The room was maintained at standard temperature
(20°C) and humidity (50–60%) with a 12 h light-dark cycle. The mice
were given sterilized water and food. Prior to treatment, the mice
were not fed for >12 h. The mice were fed with a standard diet
ad libitum. The present study was approved by the Animal
Ethics Committee of Nanjing University of Chinese Medicine
(Nanjing, China) and performed according to the Guidelines for the
Care and Use of Laboratory Animals (19).
Cell culture
The murine RAW264.7 macrophage cell line was
obtained from the Type Culture Collection of the Chinese Academy of
Sciences (Shanghai, China), and cultured in DMEM supplemented with
10% FBS, 100 U/ml penicillin and 100 µg/ml streptomycin at 37°C in
a humidified atmosphere containing 5% CO2. The cells
were sub-cultured every 2–3 days.
Extraction of crude protein
The extraction was performed as previously described
(6). Decorticated seeds of C.
tiglium L. were powdered using a blender and extracted with
petroleum ether until colorless. The petroleum ether was
subsequently removed by leaching. The resulting powders were
allowed to dry in a ventilated room and extracted with PBS
(Concentration=0.01 mol/l; pH 7.2). The mixture was stirred on a
magnetic stirrer for 2–3 h and left overnight at 4°C. The
supernatant was collected by centrifugation at 10,000 × g for 20
min at 4°C, brought to 80% saturation with saturated
(NH4)2SO4 added slowly with constant stirring
and subsequently left overnight at 4°C. Protein precipitate was
collected by centrifugation at 10,000 × g for 20 min at 4°C,
dissolved in the minimum amount of PBS and dialyzed for 48 h
against a continuous flow of the same buffer. The supernatant was
collected by centrifugation at 10,000 × g for 20 min at 4°C,
filtered, freeze-dried and stored in a refrigerator at −80°C,
giving crude protein powders.
Animal treatment and sample
collection
Acute toxicity study
i) Oral administration. For the oral acute toxicity
study, a series of doses were set based on a median lethal dose
(LD50) estimated in the present study:
Dmax=4,996.59 mg/kg; Dmin=2,094.56 mg/kg. A
total of 60 mice of either sex were exposed to 4,996.59, 3,999.67,
3,198.82 2,558.89 2,050.68 and 0 mg/kg crude protein. Crude protein
in physiological saline solution was administered. The mice were
also intragastrically administrated with sterile physiological
saline as a control. Following the intragastric administration,
symptoms and mortality were observed and recorded throughout the
experiment. At the end of the experiment, all animals were
sacrificed for subsequent study and histological analysis of main
organs. LD50 was estimated according to the Bliss method
(20) using LD50CALC version 2.0,
developed by Professor Jianhua Wang (Zhongshan School of Medicine,
Guangzhou, China).
ii) Intraperitoneal administration. Acute toxicity
was studied using intraperitoneal injection and based on an
LD50 estimating study: Dmax=523.64 mg/kg;
Dmin=53.46 mg/kg. A total of 90 mice of either sex were
exposed to 523.64, 414.95, 331.68, 265.54, 212.24, 169.65, 135.49,
108.22 and 0 mg/kg crude protein. Crude protein in physiological
saline solution was administered intraperitoneally. The following
steps were the same as those for intragastric administration.
LD50 was estimated according to the Bliss method (Bliss,
1935).
Histopathological examination
The digestive tract, heart, liver, spleen, lung,
kidney and brain were removed, fixed in 10% formalin, embedded in
paraffin, sectioned, and stained with hematoxylin and eosin for
histological examination using an optical microscope at ×400
magnification. A professional pathologist was blind to the identity
and analysis of the pathological sections, and score for the
injuries.
Gut permeability
Following overnight fasting, a total of 40 ICR mice
of either sex were orally administrated with 0, 0.75, 1.5 and 3
g/kg crude protein (10 g/0.2 ml). After 1 h, they intragastrically
received fluorescein isothiocyanate (FITC)-conjugated dextran (0.2
mg/g body weight; mean molecular weight, 3,000–5,000 kDa;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) by oral
administration. Plasma (200 µl) was sampled from the mouse eyes
every 1 h. The FITC-dextran concentration in the plasma was
determined with a fluorescent microplate reader (excitation, 485
nm; emission, 535 nm).
Inflammatory mediator PGE2 assay
A total of 60 ICR mice of either sex were
administrated intraperitoneally with 25, 50, 100, 200 and 400 mg/kg
crude protein. The control group was injected with physiological
saline. The dose was 0.2 ml/10 g. After 30 min, all animals were
sacrificed for subsequent experimentation. The dead mice were
dissected, with the abdominal membrane exposed and subsequently
injected with 1 ml physiological saline. The peritoneal fluid was
thereafter collected in 5 min. The content of inflammatory mediator
PGE2 was determined according to the instructions of the PGE2
kit.
TNF-α and IL-1β assay
RAW264.7 macrophages were plated on 48-well plates
at a density of 1×105 cells/well and cultured in
serum-free DMEM medium (Hyclone; GE Healthcare Life Sciences,
Logan, UT, USA). To analyze the dose dependence, the cells were
treated with crude protein (0.78125–400 µg/ml) stimulation for 3 h
to determine the highest concentration of proinflammatory effect.
To study time dependence, the association between the
proinflammatory effect and time at the highest concentration was
investigated. The release of TNF-α (cat. no. 88-7324-86;
eBioscience; Thermo Fisher Scientific, Inc.) and IL-1β (cat. no.
EK201B2; Multi Sciences Biotech Co., Ltd.) by the cells was
determined by ELISA using commercial kits and following
manufacturer's protocol. The culture supernatant (100 µl) was used
for the estimation, with its absorbance determined at 450 nm and
570 nm. All estimations were performed in triplicate.
Western blotting
Macrophages were inoculated at a density of
1×107 cells/ml in a cell culture dish and cultured in
the serum starvation state. The cultured macrophages were divided
into four groups (experiments for each group were performed in
triplicate), of which three groups were given croton protein
solutions with the same volume but at different concentrations,
making the final concentrations 25, 50 and 100 µg/ml. The blank
group was given DMEM of the same volume and the cells were cultured
for 3 h in 5% CO2 at 37°C following shaking. After 3 h,
RAW264.7 cells were washed with cord buffer (10 mM PBS, 0.0067 M
(PO4), Ca, Mg; pH 7.4) and subsequently lysed with
radioimmunoprecipitation assay buffer (Nanjing KeyGen Biotech Co.,
Ltd.) containing a protease inhibitor mixture (0.1 mM
phenylmethylsulfonyl fluoride, 5 mg/ml aprotinin, 5 mg/ml pepstatin
A and 1 mg/ml chymostatin). Following the centrifugation at 825 ×
g, 4°C for 10 min, the supernatant was collected and protein
concentrations were determined using a bicinchoninic acid kit
(Thermo Fisher Scientific, Inc.). A total of 40 µg cellular protein
was subjected to SDS-PAGE on a 10% gel and transferred to a PVDF
membrane, which was activated in methanol. The membrane was
incubated with 5% bovine serum albumin (BSA; Biosharp, Hefei,
China) for 2 h at room temperature, followed by incubation with
primary antibodies at 1:1,000 (v/v) dilution in 5% BSA overnight at
4°C. Blots were washed three times with Tween-20/TBS (TTBS) and
incubated with a 1:10,000 (v/v) dilution of horseradish peroxidase
(HRP)-conjugated secondary antibody for 2 h at room temperature.
GAPDH was used as a loading control. The blots were washed three
times again with TTBS, probed by enhanced chemiluminescence HRP
substrate and autoradiographed using Image Lab version 3.0 (Bio-Rad
Laboratories, Hercules, CA, USA).
Statistical analysis
In order to compare the control and treated groups,
the Student's t-test was used. The data are presented as the mean ±
standard error and were statistically analyzed using SPSS software
(version 19; IBM SPSS, Armonk, NY, USA). P<0.05 was considered
to indicate a statistically significant difference.
Results
LD50
According to the Bliss analysis, LD50 of
oral medication (Feiller correction) at 95% credibility was
2,752.8–3,407.5 mg/kg (Table I)
and that of intraperitoneal injection (Feiller correction) at 95%
credibility was 195.8–272.69 mg/kg (Table II).
 | Table I.Bliss analysis for the oral median
lethal dose of Croton tiglium L. protein. |
Table I.
Bliss analysis for the oral median
lethal dose of Croton tiglium L. protein.
| Dose, mg/kg | Logarithmic dose,
x | Mouse no. | No. of
mortalities | Percentage mortality,
% | Probability unit,
Y | Regression
probability, Y |
|---|
| 4996.59 | 3.70 | 10 | 10 | 100.00 | n/a | 7.61 |
| 3999.67 | 3.60 | 10 | 9 |
90.00 | 6.28 | 6.42 |
| 3198.82 | 3.50 | 10 | 6 |
60.00 | 5.25 | 5.23 |
| 2558.89 | 3.41 | 10 | 2 |
20.00 | 4.16 | 4.04 |
| 2050.68 | 3.31 | 10 | 0 |
0.00 | n/a | 2.84 |
| Table II.Bliss analysis for the
intraperitoneal median lethal dose of Croton tiglium L.
protein. |
Table II.
Bliss analysis for the
intraperitoneal median lethal dose of Croton tiglium L.
protein.
| Dose, mg/kg | Logarithmic dose,
x | Mouse no. | No. of
mortalities | Percentage
mortality, % | Probability unit,
Y | Regression
probability, Y |
|---|
| 523.64 | 2.72 | 10 | 10 | 100.00 | n/a | 6.77 |
| 14.95 | 2.62 | 10 | 9 | 90.00 | 6.28 | 6.27 |
| 331.68 | 2.52 | 10 | 7 | 70.00 | 5.52 | 5.78 |
| 265.54 | 2.42 | 10 | 6 | 60.00 | 5.25 | 5.30 |
| 212.24 | 2.33 | 10 | 4 | 40.00 | 4.75 | 4.81 |
| 169.65 | 2.23 | 10 | 3 | 30.00 | 4.48 | 4.33 |
| 135.49 | 2.13 | 10 | 2 | 20.00 | 4.16 | 3.84 |
| 108.22 | 2.03 | 10 | 0 |
0.00 | n/a | 3.35 |
Pathological examination
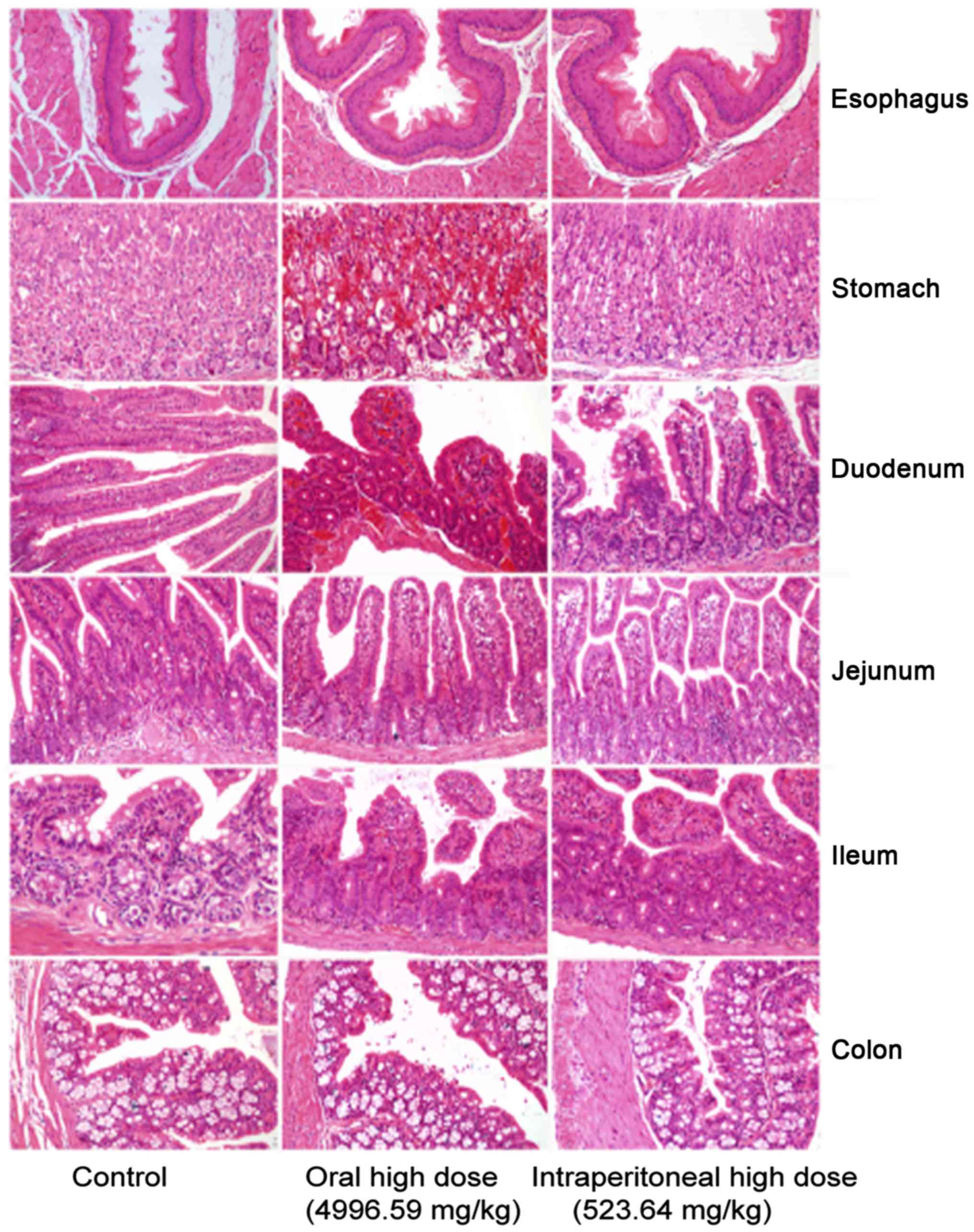
In the control group, gastrointestinal sections
exhibited healthy structural features. The oral medication and
intraperitoneal injection groups exhibited obvious mucous layer,
epithelial cell and interstitial damages. The oral medication and
intraperitoneal injection groups primarily suffered from mucosal
necrosis, epithelial cell necrosis or erosion, interstitial
hyperemia or bleeding, mucosal congestion and villous edema in the
intestine. However, they exhibited different extents of
gastrointestinal damage (Fig. 1).
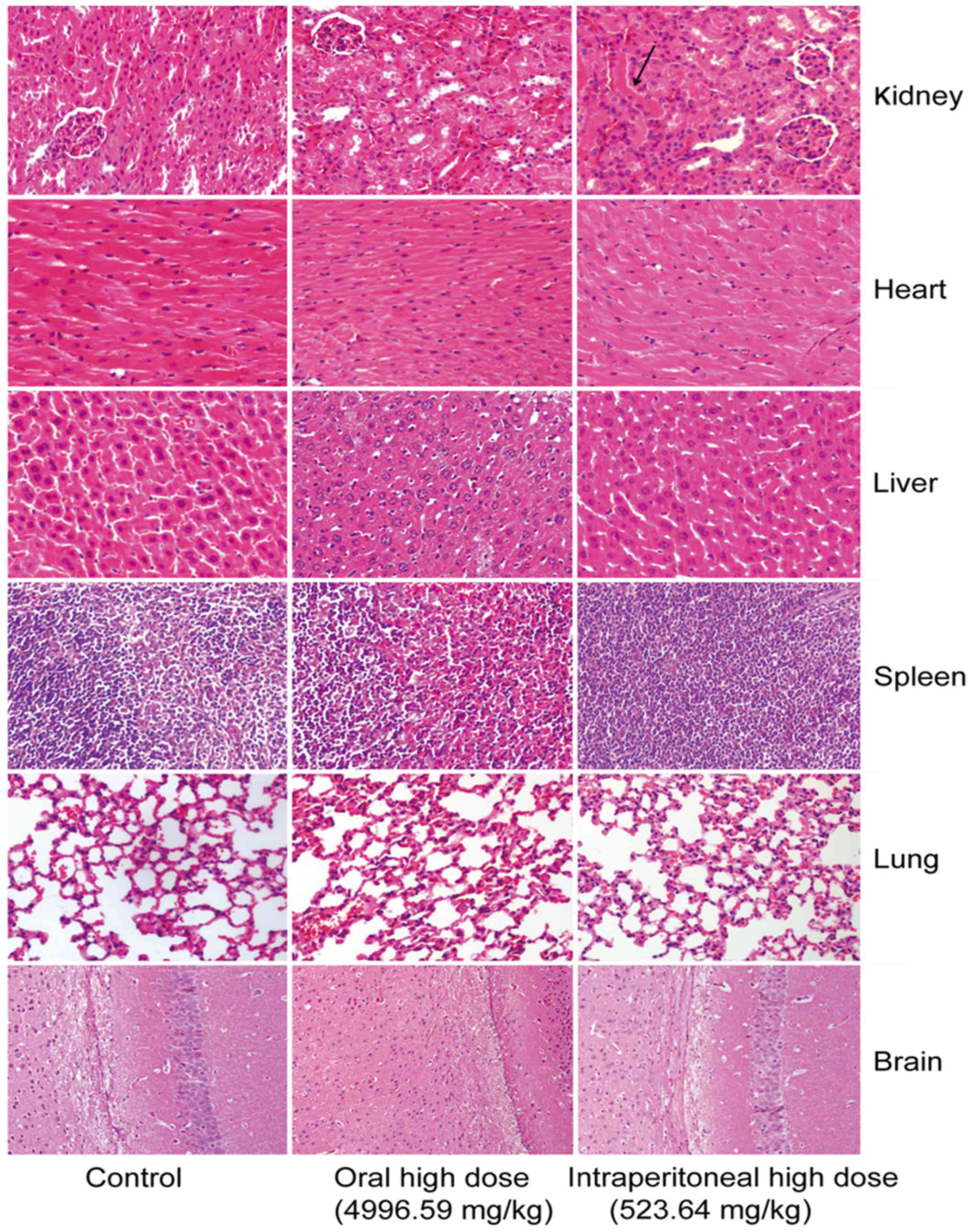
The mice intraperitoneally administrated with crude protein
underwent degeneration of renal tubular epithelial cells in the
kidney. No marked difference was observed in other tissues or
organs (Fig. 2).
Evaluation of intestinal permeability with
FITC-dextran
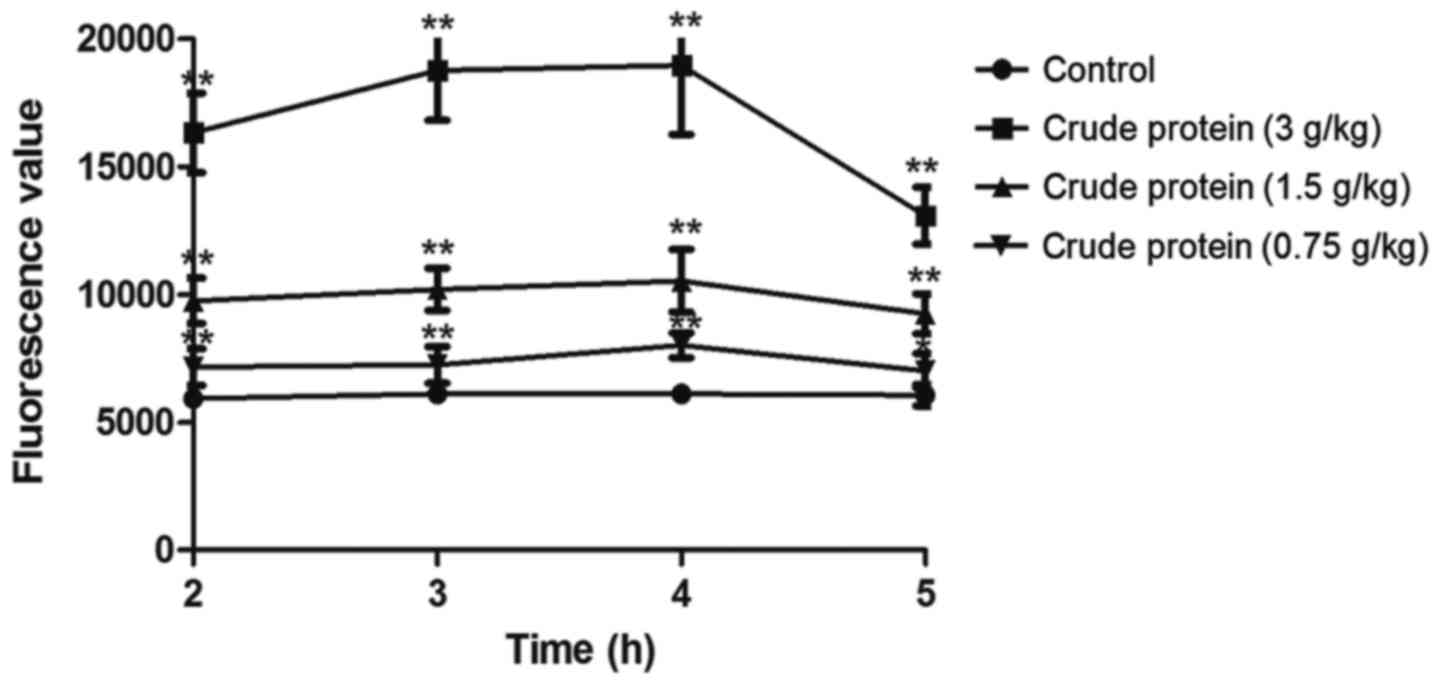
Compared with the control group (Fig. 3), croton proteins significantly
increased the intestinal permeability of the oral administration
group. When the dose reached 3 g/kg, the intestinal permeability
reached maximum. Following 4 h of treatment, the digestive tract
was most obviously damaged, and the intestinal permeability
gradually recovered. Meanwhile, the toxicity was gradually
reduced.
In vivo proinflammatory
evaluation
PGE2 assay
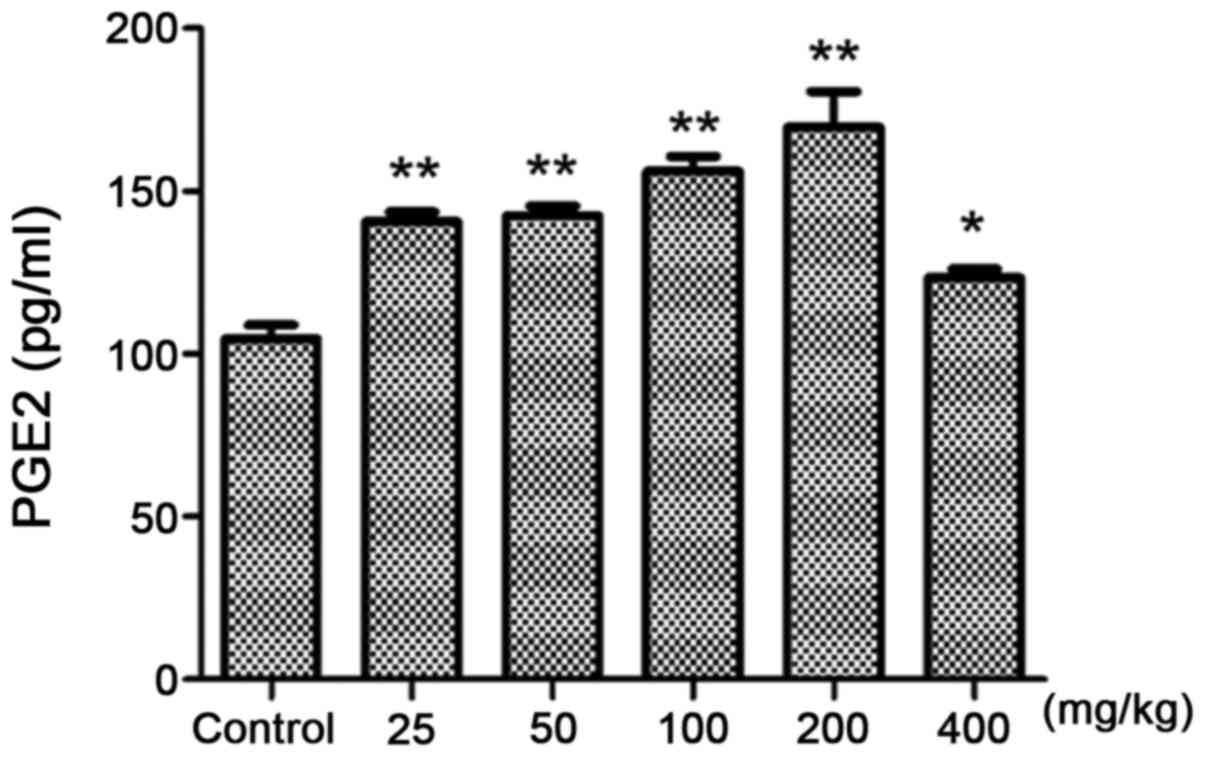
Compared with the control group (Fig. 4), 25 mg/kg croton proteins
significantly increased PGE2 in the abdominal fluid (P<0.01).
When the dose reached 200 mg/kg, the content of PGE2 reached the
maximum. Therefore, the toxicity of croton proteins is associated
with inflammation.
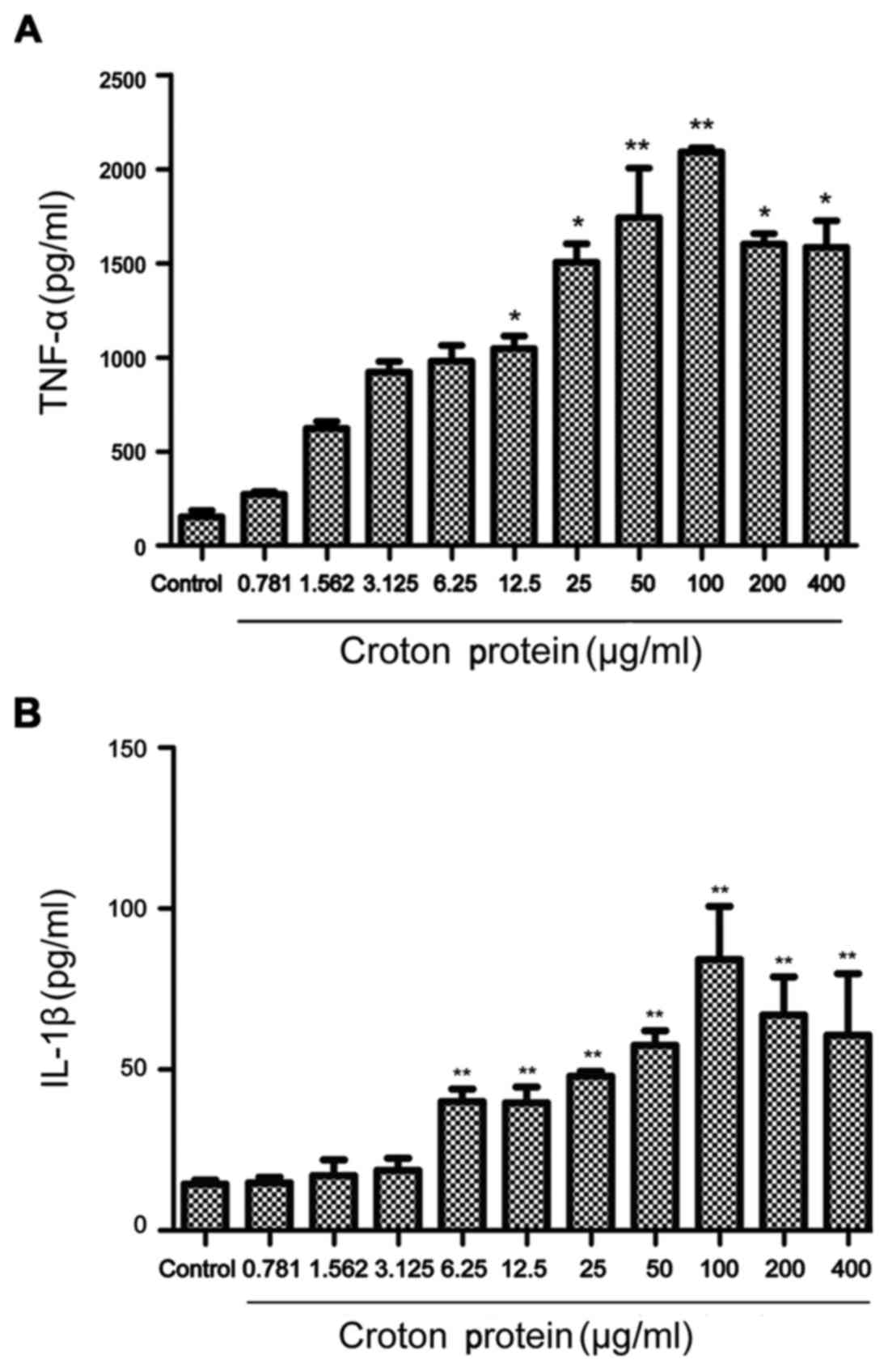
Croton proteins induces the release of the
proinflammatory cytokines TNF-α and IL-1β in RAW264.7 macrophages
in a dose-dependent manner
Croton proteins significantly stimulated the
production of proinflammatory cytokines dose-dependently (Fig. 5). Compared with the control group,
croton proteins (50, 100, 200 and 400 µg/ml) increased cytokines,
including TNF-α and IL-1β, significantly after 3 h of treatment
(P<0.01). Furthermore, when the concentration of croton proteins
reached 100 µg/ml, maximum TNF-α and IL-1β were released. In the
presence of croton proteins at >100 µg/ml the TNF-α and IL-1
levels began to decrease.
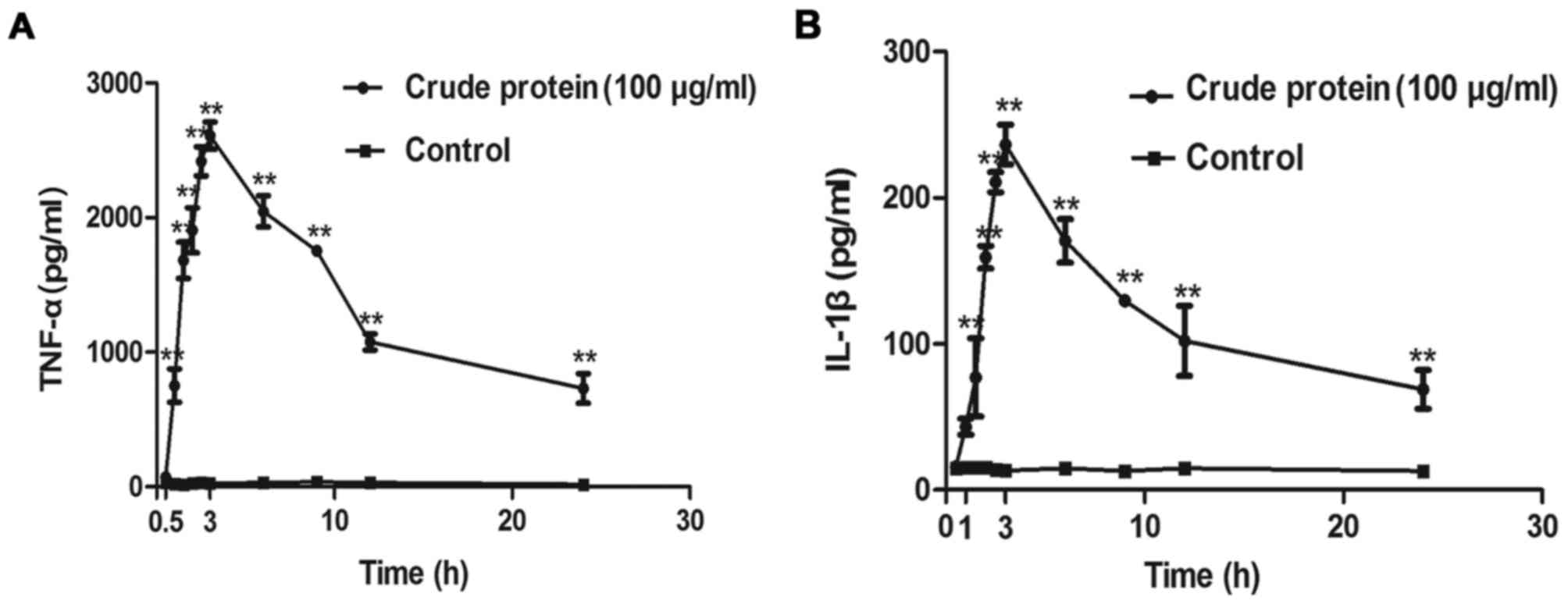
Croton proteins induces the release of the
proinflammatory cytokines TNF-α and IL-1β in RAW264.7 macrophages
in a time-dependent manner
Croton proteins also stimulated the production of
proinflammatory cytokines in a time-dependent manner (Fig. 6). After 3 h of treatment, the
release of TNF-α and IL-1β reached maximum.
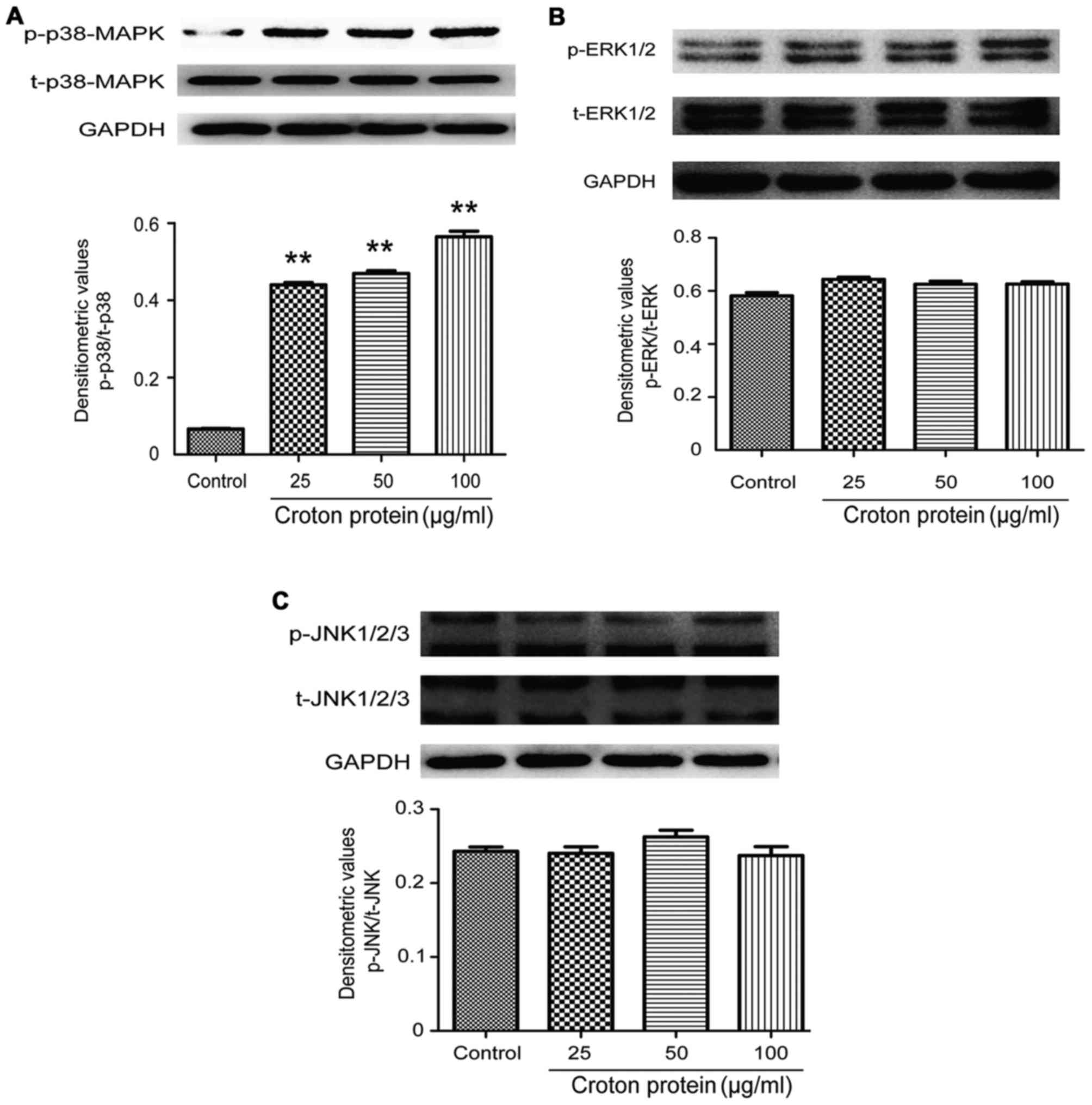
Effect of crude protein on the MAPK signaling
pathway in RAW264.7 cells
To understand the molecular mechanisms underlying
the proinflammatory effect of crude protein on the production of
various cytokines, the effects of crude protein on the MAPK
signaling pathway were examined. p38-MAPK, p-p38-MAPK, ERK1/2,
p-ERK1/2, JNK1-3 and p-JNK1-3 protein levels were decreased in
RAW264.7 cells upon croton proteins treatment (0, 25, 50 and 100
µg/ml) (Fig. 7).
Discussion
C. tiglium L. as a classic traditional
Chinese medicine, has been widely used to treat indigestion,
abdominal distension, visceral pain, constipation and other
gastrointestinal disorders; however, it exhibits toxicity. It is
essential to investigate the toxic composition and targets for
improved clinical application. To analyze the toxic components,
croton oil has been extensively studied, whereas croton proteins
has seldom been assessed. Croton oil is an active and toxic
ingredient which, following dilution to 18–20%, may be safe used in
clinical practice. Nevertheless, the dilution method induces strong
gastrointestinal edema and diarrhea (21). Therefore, the toxicity of C.
tiglium L. cannot be completely attributed to croton oil. For
the dilution method, the toxicity of C. tiglium L. is
reduced by first heating with steam and subsequent dilution, to
ensure the clinical curative effect and safety. The process of
heating, which denatures croton proteins that exhibit the majority
of the toxicity, primarily weakens the toxicity of C.
tiglium L.; therefore, it is important to study the toxicity of
these proteins to the digestive tract. Ricinus communis L.
is a plant of the same family as that of C. tiglium and
produces ricin, which is able to cause strong toxicity to the
digestive tract, manifested as edema and diarrhea primarily
associated with inflammation (22). Therefore, the toxicity of orally
administrated croton proteins to the digestive tract may be
associated with the inflammatory response. Inflammation is one of
the most important protective defensive reactions to tissue injury.
Furthermore, the proinflammatory signaling pathway in macrophages
further determines the production of these factors (23). The MAPK signaling pathway is an
important pathway responsible for inflammatory responses. However,
the process of inflammatory response is accompanied by activation
of other signaling pathways, including nuclear factor (NF)-κB. The
activation of the NF-κB signaling pathway is indicated by p65
nuclear translocation (24,25),
which will be investigated in future studies.
The present study demonstrated that oral medication
of croton proteins caused significant gastrointestinal damage with
edema and diarrhea. The toxic reactions of the crude protein were
associated with inflammation. In vivo, the crude protein
caused the release of inflammatory mediator PGE2 in mice by
intraperitoneal injection. In vitro, proinflammatory
cytokines, including TNF-α and IL-1β, were produced in macrophages
in a dose- and time-dependent manner. Furthermore, the crude
protein in macrophages was associated with MAPK signaling pathway
and the activated p38-MAPK pathway. In clinical applications, the
effective components in C. tiglium L. frequently coexist
with toxic ingredients, including croton oil and protein, that may
cause gastrointestinal toxicity. Therefore, they may have a
synergistic effect, and it is necessary to clarify the toxic
ingredients and targets, as well as the synergistic toxicity to
ensure safe use of C. tiglium L. in clinical practice. The
aim of future studies will be to solve these problems.
References
|
1
|
National Pharmacopoeia Committee:
Pharmacopoeia of the People's Republic of China. Beijing: China
Medical Science and Technology Press; 2015
|
|
2
|
Shahid M, Tayyab M, Naz F, Jamil A, Ashraf
M and Gilani AH: Activity-guided isolation of a novel protein from
Croton tiglium with antifungal and antibacterial activities.
Phytother Res. 22:1646–1649. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang JF, Yang SH, Liu YQ, Li DX, He WJ,
Zhang XX, Liu YH and Zhou XJ: Five new phorbol esters with
cytotoxic and selective anti-inflammatory activities from Croton
tiglium. Bioorg Med Chem Lett. 25:1986–1989. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang DD, Zhou B, Yu JH, Xu CH, Ding J and
Zhang H: Cytotoxic tigliane-type diterpenoids from Croton
tiglium. Tetrahedron. 71:9638–9644. 2015. View Article : Google Scholar
|
|
5
|
State Administration of Traditional
Chinese Medicine: Materia Medica of China. Shanghai: Shanghai
Scientific Technology Press; pp. 7691999
|
|
6
|
Stirpe F, Pession-Brizzi A, Lorenzoni E,
Strocchi P, Montanaro L and Sperti S: Studies on the proteins from
the seeds of Croton tiglium and of Jatropha curcas. Toxic
properties and inhibition of protein synthesis in vitro. Biochem J.
156:1–6. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cabral PH, de Morais Campos R, Fonteles
MC, Santos CF, Leal Cardoso JH and do Nascimento NR: Effects of the
essential oil of Croton zehntneri and its major components,
anethole and estragole, on the rat corpora cavernosa. Life Sci.
112:74–81. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen P, Zhou X, Zhang L, Shan M, Bao B,
Cao Y, Kang A and Ding A: Anti-inflammatory effects of Huangqin
tang extract in mice on ulcerative colitis. J Ethnopharmacol.
162:207–214. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Baines KJ, Backer V, Gibson PG, Powel H
and Porsbjerg CM: Impaired lung function is associated with
systemic inflammation and macro-phage activation. Eur Respir J.
45:557–559. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wen Z, Fan L, Li Y, Zou Z, Scott MJ, Xiao
G, Li S, Billiar TR, Wilson MA, Shi X and Fan J: Neutrophils
counteract autophagy-mediated anti-inflammatory mechanisms in
alveolar macrophage: Role in posthemorrhagic shock acute lung
inflammation. J Immunol. 193:4623–4633. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lai DM: Pathology. The reasons of
Inflammation Beijing: People's Medical Publishing House; pp. 49–52.
2014
|
|
12
|
Cronstein BN and Weissman G: The adhesion
molecules of inflammatio. Arthritis Rheum. 36:147–157. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Noah T. Ashley, Zachary M Weil and Randy
J. Nelson: Inflammation: Mechanisms, costs and natural variation.
Ann Rev Ecol Evolution and Systemat. 43:385–406. 2012. View Article : Google Scholar
|
|
14
|
Galley HF and Webster NR: The
immuno-inflammmatory cascade. Brit J Anaesth. 77:11–16. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Valko M, Leibfritz D, Moncol J, Cronin MT,
Mazur M and Telser J: Free radicals and antioxidants in normal
physiological functions and human disease. Int J Biochem Cell Biol.
39:44–84. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang J, Qin Y and Mi X: The protective
effects of bone marrow-derived mesenchymal stem cell (BMSC) on
LPS-induced acute lung injury via TLR3-mediated IFNs, MAPK and
NF-κB signaling pathways. Biomed Pharmacother. 79:176–187. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guo C, Hao LJ, Yang ZH, Chai R, Zhang S,
Gu Y, Gao HL, Zhong ML, Wang T, Li JY and Wang ZY:
Deferoxamine-mediated up-regulation of HIF-1α prevents dopaminergic
neuronal death via the activation of MAPK family proteins in
MPTP-treated mice. Exp Neurol. 280:13–23. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yıldız MT and Arslanyolu M: In silico
identification and characterization of the MAPK family members of
unicellular model eukaryoteTetrahymena thermophila. Eur J
Protistol. 50:538–550. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jones-Bolin S: Guidelines for the care and
use of laboratory animals in biomedical research. Curr Protoc
Pharmacol: Appendix 4: Appendix 4B. 2012. View Article : Google Scholar
|
|
20
|
Bliss CI: The calculation of the
dosage-mortality curve. Ann Appl Biol. 22:134–167. 1935. View Article : Google Scholar
|
|
21
|
Yi Wang and Zhang JX: New processing
method of croton cream and its acute toxicity test. Chin Herbal
Med. 4:24–27. 1993.
|
|
22
|
Linna Liu, Hongwei Gao, Jiping Li, Ying
Dong, Ning Liu, Jiayu Wan, Wensen Liu, Yucheng Sun and Ming Xu:
Analysis of intestinal injuries induced by ricin in vitro using SPR
technology and MS identification. Int J Mol Sci. 5:2431–3439.
2009.
|
|
23
|
Wang XL: Prarctical molecular
pharmacology. Inflammatory mediators and anti-inflammatory drugs.
Wang XL: Beijing: Peking Union Medical College Publishers; pp.
433–438. 2005
|
|
24
|
Kim HR, Shin DY and Chung KH: The role of
NF-κB signaling pathway in polyhexamethylene guanidine phosphate
induced inflammatory response in mouse macrophage RAW264.7 cells.
Toxicol Lett. 233:148–155. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bey Williams Y, Boularan C, Vural A, Huang
NN, Hwang IY, Shan-Shi C and Kehrl JH: Omega-3 free fatty acids
suppress macrophage inflammasome activation by inhibiting NF-κB
activation and enhancing autophagy. PLoS One. 9:e979572014.
View Article : Google Scholar : PubMed/NCBI
|