Introduction
Age-related macular degeneration (AMD) is the
leading cause of irreversible blindness among the aged in advanced
countries. AMD causes ~20% of legal blindness AMD, and it is
estimated that ~80 million people worldwide will suffer from AMD by
the year 2020 (1). Late AMD has
two forms: One is the ‘dry’ form defined by degeneration of the
retinal pigment epithelium (RPE) cells and photoreceptor cells, and
the other is ‘wet’ form, associated with choroidal
neovascularization (CNV). Anti-angiogenic therapies have been
successful in managing wet AMD. However, dry AMD, which accounts
for 90% of AMD cases, currently lacks an effective treatment to
stop or even slow down disease progression.
AMD is a multi-factorial complex condition with
poorly understood molecular mechanisms. Age, smoking, specific
genetic polymorphisms, oxidative stress, the complement pathway,
inflammation and pathogenic RNA species (Alu) are
significant contributors to AMD pathogenesis (2–8).
There is much evidence that suggests that cumulative long-term
exposure to ultraviolet B (UVB) may lead to AMD irrespective of
age, because of the direct DNA damage and reactive oxygen species
(ROS) production in RPE cells (9).
Although most UVB is mostly absorbed by the cornea and lens, as the
depletion of the ozone layer increases, there is a considerable
growth in the accumulated lifetime exposure of the retina to UVB,
especially following cataract removal (10). Visual impairment in dry AMD is
associated with the degeneration of RPE cells and photoreceptor
cells (11). RPE, a polarized
monolayer epithelium cell layer, locates between the neural retina
and choroid, acting as the guardians of the photoreceptor (12). Moreover, RPE cells selectively
absorb the lower wavelength light particles (13). Therefore, it is believed that RPE
cells may be the main target of UVB reaching the retina.
Notch signaling is a conserved adjacent cell
signaling mechanism. In mammals, there are four Notch receptors
(Notch1-4) and each has a cytoplasmic domain implicated in signal
transduction (14,15). Notch activation is initiated by the
binding between Notch receptors and ligands on adjacent cells,
resulting in multiple steps of proteolytic cleavages of the
receptors, as well as the release of the Notch intracellular domain
(NICD) from the membrane, which translocates into the nucleus. In
the nucleus, NICD binds to the transcription factor CSL and the
co-activator mastermind-like proteins (MAML1-3), initiating
transcriptional activation of Notch target genes, such as those in
the Hes, Hey family (14,15).
Notch signaling serves an important role in many
cellular processes: Cell proliferation, differentiation, apoptosis,
migration and angiogenesis in many tissues, including pigmented and
non-pigmented cells in the eyes (14,16,17).
To date, the role of Notch signaling in dry AMD has been explored
to a very limited extent. In the present study, the effects of UVB
and the function of Notch signaling in RPE cells were investigated.
Surprisingly, the authors identified that NOTCH2 was the major
Notch receptor in RPE cells. More interestingly, the inhibition of
NOTCH2, but not NOTCH1, attenuated intracellular ROS
and cell apoptosis induced by UVB.
Materials and methods
Plasmids, small hairpin (sh)RNAs and
reagents
The following shRNA lentiviral constructs targeting
the human NOTCH1 and NOTCH2 were obtained from Thermo
Fisher Scientific, Inc. (Waltham, MA, USA) and were used in a
previous study (18). The hairpin
sequence numbers are TRCN0000003359, TRCN0000003362 (targeting
NOTCH1) and TRCN0000004895 and TRCN0000004896 (targeting
NOTCH2).
The following antibodies were used in western
blotting: Notch1 (cat. no. sc-6014; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA), Notch2 (cat. no. D76A6; Cell Signaling
Technology, Inc., Danvers, MA, USA) and α-tubulin (cat. no. AF7010;
Affinity Biosciences, Inc., Cincinnati, OH, USA).
Cell culture and UV light
apparatus
Human RPE cells (ARPE19 cell line) were obtained
from the American Type Culture Collection (Manassas, VA, USA). RPE
cells were cultured in Dulbecco's Modified Eagle's medium (DMEM;
HyClone, GE Healthcare Life Sciences, Chalfont, UK), supplemented
with 10% heat-inactivated fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 100 ng/ml streptomycin
in a humidified incubator at 37°C and 5% CO2. When cells
reached ~90% confluence, they were detached with 0.3% trypsin
solution (Gibco; Thermo Fisher Scientific, Inc.) and collected for
the subsequent experiments.
UV radiation apparatus used in this study is
3UV™-34UV Lamp (UVP, Inc., Upland, CA, USA), containing three
wavelengths: 254 nm (UVA), 302 nm (UVB), and 365 nm (UVC). RPE
cells (8×105) were seeded in 60 mm plates at ~50%
confluence and subjected to UVB radiation 8 h later. Prior to UV
radiation, cells were washed once with 2 ml pre-warmed PBS, and 1
ml PBS was left in the dish. Cells were radiated by UVB at various
doses (0, 25, 50, 100 mJ/cm2) without the lid of petri
dish, in the dark. Following UVB radiation, cells were cultured in
fresh culture medium for 36 h.
Flow cytometry analysis of ROS and
apoptosis
RPE cells were trypsinized and washed twice in
pre-warmed PBS prior to the analyses of ROS and apoptosis. The
harvested cells (2×105) were incubated for 30 min in a
humidified incubator with dihydroethidium (EMD Millipore,
Billerica, MA, USA), a well characterized reagent that has been
extensively used for the detection of reactive oxidative species,
according to the manufacturer's instructions. Following incubation,
the fluorescence intensity was measured using a Muse™ Cell Analyzer
(EMD Millipore). For apoptosis, the harvested cells
(2×105) were incubated for 30 min in a humidified
incubator with MultiCaspase (EMD Millipore), according to the
manufacturer's protocol, and incubated for 5 min at room
temperature with 7-AAD (EMD Millipore) and analyzed with a Muse™
Cell Analyzer.
Lentiviral transduction
Lentiviral transduction was performed as previously
described (19). In brief,
lentiviral vectors targeting NOTCH1, NOTCH2, along
with the packing plasmid PSPAX2 and pseudotyped envelope pMD2.G
(provided by Professor Lizi Wu, UF Health Shands Hospital,
University of Florida, Gainesville, FL, USA) were transfected into
293T cells (American Type Culture Collection, Manassas, VA, USA)
using the Effectene Transfection Reagent (Qiagen GmbH, Hilden,
Germany). RPE cells were plated at 40–50% confluence in 60 mm
plates and subsequently infected three times with 2 ml viruses plus
1 ml fresh complete medium containing 2 µg/ml polybrene
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). Finally, the RPE
cells were screened with puromycin (1.5 µg/ml; Sigma-Aldrich; Merck
KGaA) for one week.
Cell viability assays and scratch
assay
Cell viability was determined by MTT assay. Briefly,
RPE cells were plated at a density of 5,000 cells/well in 96-well
plates. Following culturing for the desired time (0, 24, 48 and 72
h), 20 µl MTT (5 mg/ml; Sigma-Aldrich; Merck KGaA) was added in
each well, and then incubated for 4 h at 37°C. Then, all fluid was
removed, and the crystallized dyes were dissolved in 150 µl
dimethyl sulfoxide (Sigma-Aldrich; Merck KGaA) per well and shaken
on a shaking table bed for 10 min. The absorbance at 490 nm
wavelength was detected with a micro-plate reader (BioTek
Instruments, Inc., Winooski, VT, USA).
For the scratch assay, RPE cells (8×105)
were seeded into a 6-well plate in growth medium at 80–90%
confluence. The scratch was drawn using a white tip and the
floating cells were removed. Then the cells were cultured in DMEM
medium with 2% fetal bovine serum. Migration of the cells into the
scratch area was observed 48 h after the scratch had been
drawn.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) and western blotting
Real-time RT-PCR was performed as described
previously (20). Total RNA was
extracted from RPE cells using TRIzol reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) following the manufacturer's instructions.
Then, cDNA was reverse transcribed using the PrimeScript II 1st
Strand cDNA Synthesis kit (Takara Bio, Inc., Japan). RT-qPCR
analysis was performed on LightCycler 96 (Roche Diagnostics, Basel,
Switzerland) using the SYBR Premix Ex TaqTM kit (Takara Bio, Inc.).
The human GAPDH gene was used as an endogenous control for sample
normalization. The sequences of the 17 pairs of primers used in
this study are not presented.
Western blot analysis was performed as described
previously (21). In brief, 50 mg
proteins were separated by using 8% SDS-PAGE, electrotransferred to
pure nitrocellulose blotting membranes, and probed with the
indicated antibodies as recommended by the manufacturer. Prior to
incubation with primary antibodies, membranes were blocked with 5%
fat-free milk for 1 h at room temperature. This was followed by
incubation with Notch1 (1:200), Notch2 (1:1,000) and α-tubulin
(1:1,000) primary antibodies overnight at 4°C. The membranes were
subsequently washed three times with TBS-Tween-20 for 10 min and
incubated with goat anti-rabbit horseradish peroxidase
(HRP)-conjugated secondary antibodies (1:5,000; cat. no. 14708;
Cell Signaling Technology, Inc.) for 1 h at 37°C. Antibody binding
was visualized using Immobilon™ Western Chemiluminescent HRP
Substrate (EMD Millipore) and detected by a Tanon5200
Chemiluminescent Imaging System (Tanon Science and Technology Co.,
Ltd., Shanghai, China). Loading was normalized with α-tubulin.
Statistical analysis
Each experiment was conducted in triplicate. The
values were expressed as the mean ± standard deviation. Statistical
analyses were performed by using Student's t-test assuming equal
variances for all data (comparison of two groups) by SPSS software
(version, 13.0; SPSS, Inc., Chicago, IL, USA). P<0.05 and
P<0.01 were determined to indicate a statistically significant
difference.
Results
UVB radiation increased intracellular
ROS and induced apoptosis in RPE cells
To determine UVB-induced damage, RPE cells were
exposed to UVB (0, 25, 50, 100 mJ/cm2), and then
cultured for 36 h. Intracellular ROS and apoptotic cells were
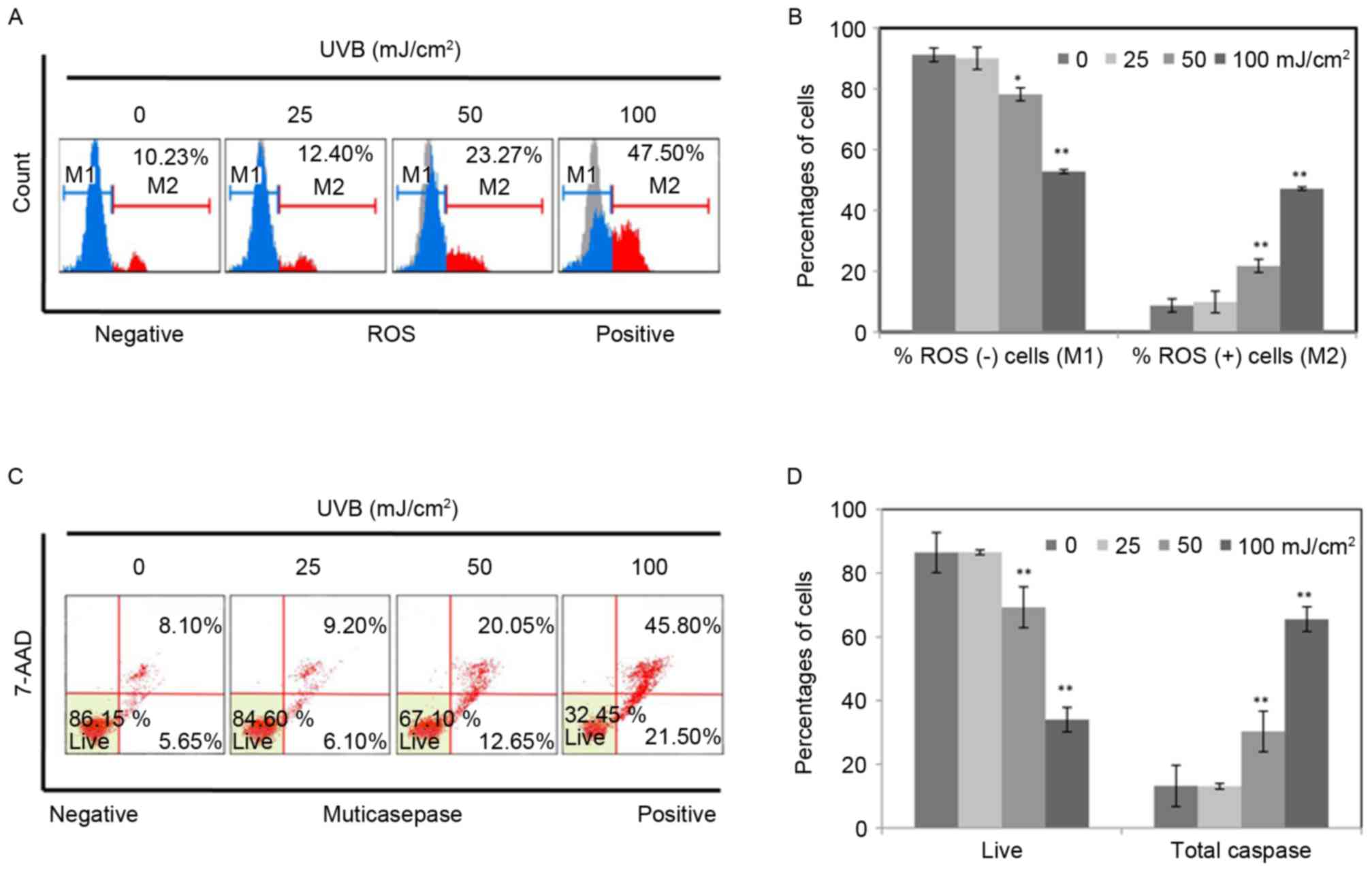
measured by flow cytometry. As presented in Fig. 1A, the percentages of ROS positive
cells were identified to increase from 10.23 to 47.50% with an
increasing intensity of UVB, indicating that UVB significantly
increased ROS production in RPE cells. In addition, the percentage
of apoptotic cells increased notably following UVB treatment in a
dose dependent manner (Fig. 1C).
The percentage of total caspase increased from 13.75 to 67.3% along
with the increase of intensity of UVB. The results are represented
as the means ± standard deviation determined from three independent
experiments (Fig. 1B and D). The
results in Fig. 1 indicated that
UVB induced damage in RPE cells.
UVB activated the Notch signaling in
RPE cells
To understand the function of Notch signaling in RPE
cells, RT-qPCR was conducted to confirm the expression levels of
all 13 key components of Notch signaling, including four receptors
(NOTCH1-4), five NOTCH ligands (JAGGED1 and 2,
DELTA-LIKE-1, 3 and 4), three transcriptional
co-activators (MAML1, 2 and 3) and the transcription
factor (CSL) (Fig. 2A).
Notably, NOTCH2 was the major Notch receptor in RPE cells.
Among them, the expression levels of NOTCH3, 4, DELTA-LIKE 1, 3,
4 and MAML 3 are too low to display.
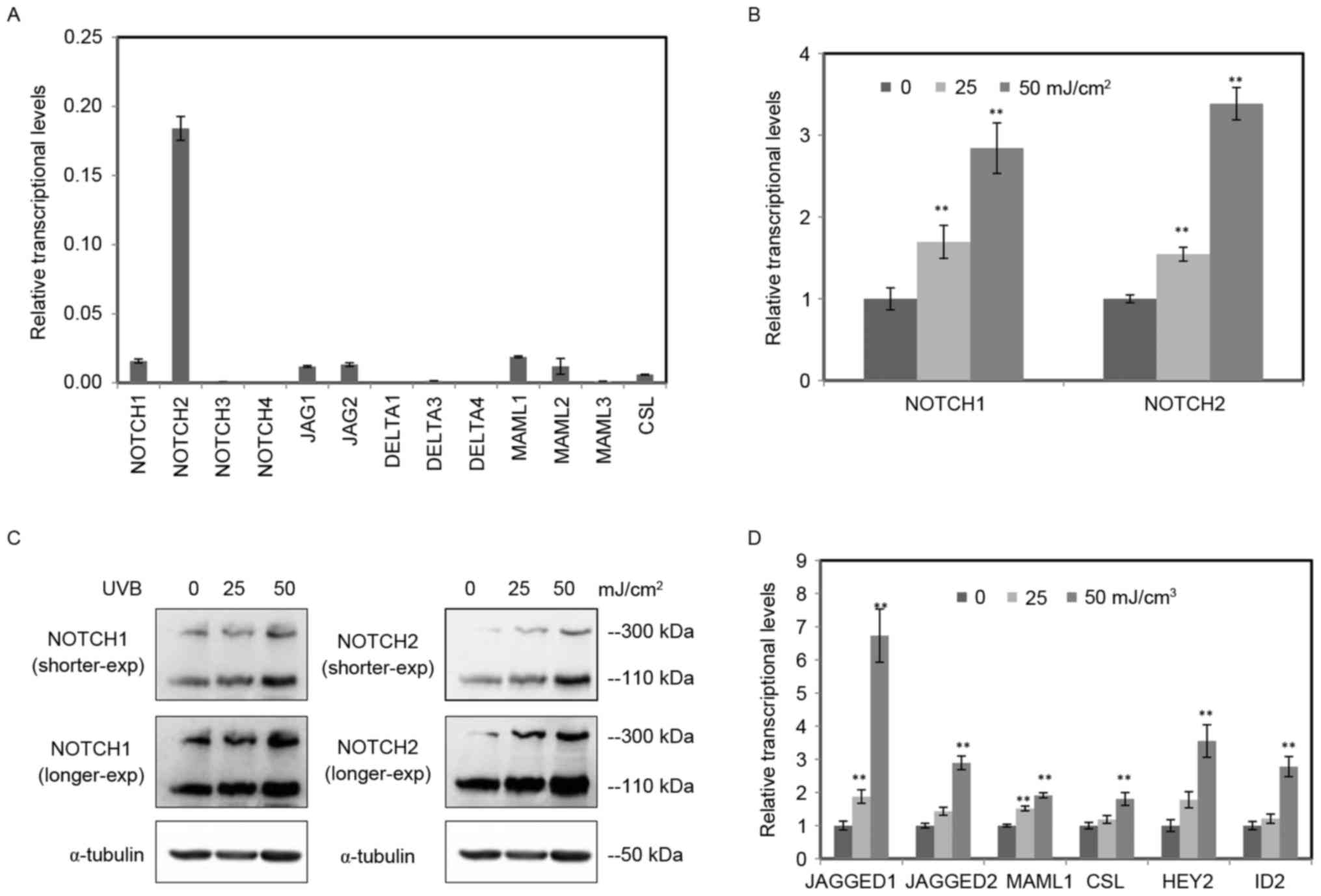 | Figure 2.UVB activated the Notch signaling in
RPE cells. (A) The relative expression levels of Notch signaling
components in RPE cells were determined by RT-qPCR. The detected
components included four receptors (NOTCH1-4), five ligands (JAGGED
1, 2 and DELTA-LIKE1, 3 and 4), three transcriptional coactivators
(MAML1, 2 and 3) and the transcription factor CSL. Among them, the
expression of NOTCH3, 4, DELTA-LIKE 1, 3 and 4 and MAML 3 are too
low to display. (B) The relative expression levels of NOTCH1 and
NOTCH2 in RPE cells radiated by UVB (0, 25, 50 mJ/cm2)
were tested by RT-qPCR. (C) The protein level of NOTCH1 and NOTCH2
in RPE cells radiated by UVB (0, 25, 50 mJ/cm2) were
tested by western blotting using α-tubulin as an internal control.
The full length of NOTCH is 300 kDa, and the cleaved Notch
intracellular domain was 110 kDa. (D) The other genes of Notch
signaling (JAGGED1, JAGGED2, MAML1, CSL, HEY2 and ID2) in RPE cells
treated by UVB were determined by RT-qPCR. The data are expressed
as the mean ± standard deviation of three independent experiments.
Statistical analysis was performed using Student's t-test.
**P<0.01 vs. control. UVB, ultraviolet B; RPE, retinal pigment
epithelium; RT-qPCR, reverse transcription-quantitative polymerase
chain reaction. |
Among the four receptors, the expression levels of
NOTCH1 and NOTCH2 were obviously higher compared with
NOTCH3 and NOTCH4 receptors (Fig. 2A), indicating they were main
receptors of Notch signaling in RPE cells. To study the function of
Notch signaling in UVB-induced damage of RPE cells, RT-qPCR was
performed to investigate the mRNA expression of NOTCH1 and
NOTCH2 in RPE cells treated with various doses of UVB (0,
25, 50 mJ/cm2; Fig.
2B). The exposure of UVB enhanced the mRNA expression of
NOTCH1 and NOTCH2 in a dose dependent manner. The
protein levels of NOTCH1 and NOTCH2 were tested by western blot
analysis using α-tubulin as an internal control (Fig. 2C). Consistent with the RT-qPCR
data, both the full length and the cleaved NICD increased in RPE
cells radiated by UVB.
Moreover, expression of other important Notch
signaling components was detected by RT-qPCR. All six important
components increased following UVB radiation, including NOTCH
ligands (JAGGED 1 and 2), transcriptional
co-activator (MAML1), transcription factor (CSL) and
target genes (HEY2 and ID2) (Fig. 2D). These data suggested that the
exposure of UVB activated the Notch signaling in RPE cells.
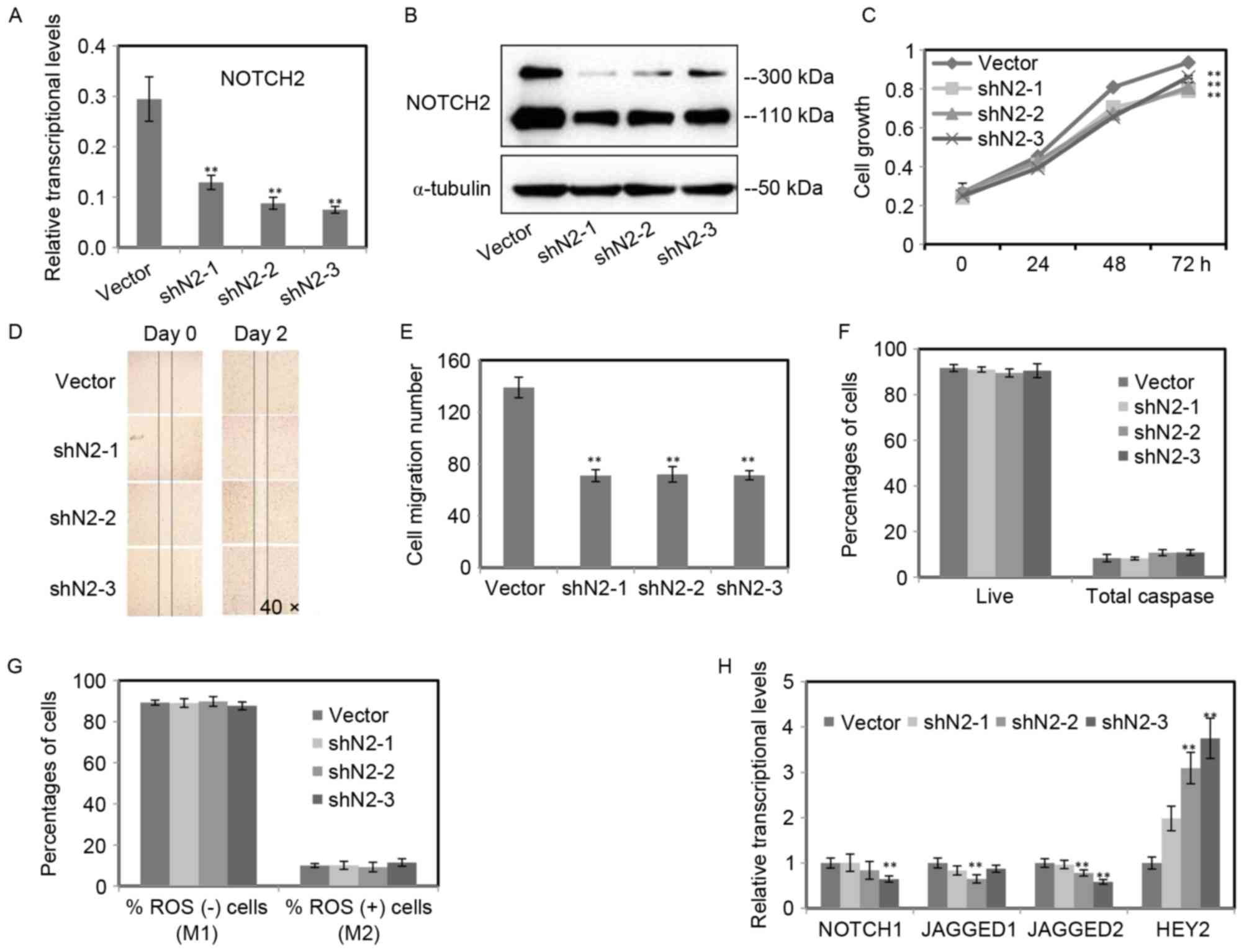
The inhibition of NOTCH2 reduced cell
growth and cell migration, but had no impacts on intracellular ROS
and cell apoptosis
As presented in Fig.
2, NOTCH2 was the major Notch receptor and increased
following UVB radiation in RPE cells. To gain an insight into the
role of NOTCH2 in RPE cells, the expression of NOTCH2
was inhibited by lentiviral-based small hairpin RNA (shRNA). Three
shRNAs (shN2-1, 2, 3) targeting NOTCH2 were selected. Then,
the efficiency of NOTCH2 knockdown was determined by RT-qPCR
(Fig. 3A) and western blot
analysis (Fig. 3B). Both mRNA and
protein levels of NOTCH2 were inhibited in the three stable
RPE cell lines.
To determine whether the inhibition of NOTCH2
has an effect on RPE cell, the cell growth, migratory capacity,
intracellular ROS and cell apoptosis of three stable RPE cell lines
were monitored. An obvious reduction of cell growth was detected in
RPE cells with NOTCH2 inhibited, when compared with the
control (Fig. 3C). The migratory
capacity of RPE cells was confirmed by scratch assay, and the
migration of the three NOTCH2-knockdown RPE cell lines were
significantly inhibited, while the control scratch wound almost
recovered following two days incubation (Fig. 3D). The number of migration cells
significantly decreased in three NOTCH2 knockdown RPE cell
lines (Fig. 3E).
Surprisingly, there were no statistical changes for
the percentage of ROS positive cells and apoptotic cells between
RPE cells with NOTCH2 inhibited and the control (P>0.05;
Fig. 3F and G). To investigate the
mechanism of the effects on the migration and proliferation of RPE
cells, RT-qPCR was performed to confirm the expression levels of
other genes of the Notch signaling. The target gene HEY2
increased notably. Moreover, JAGGED1 and 2 and
NOTCH1 decreased (Fig.
3H).
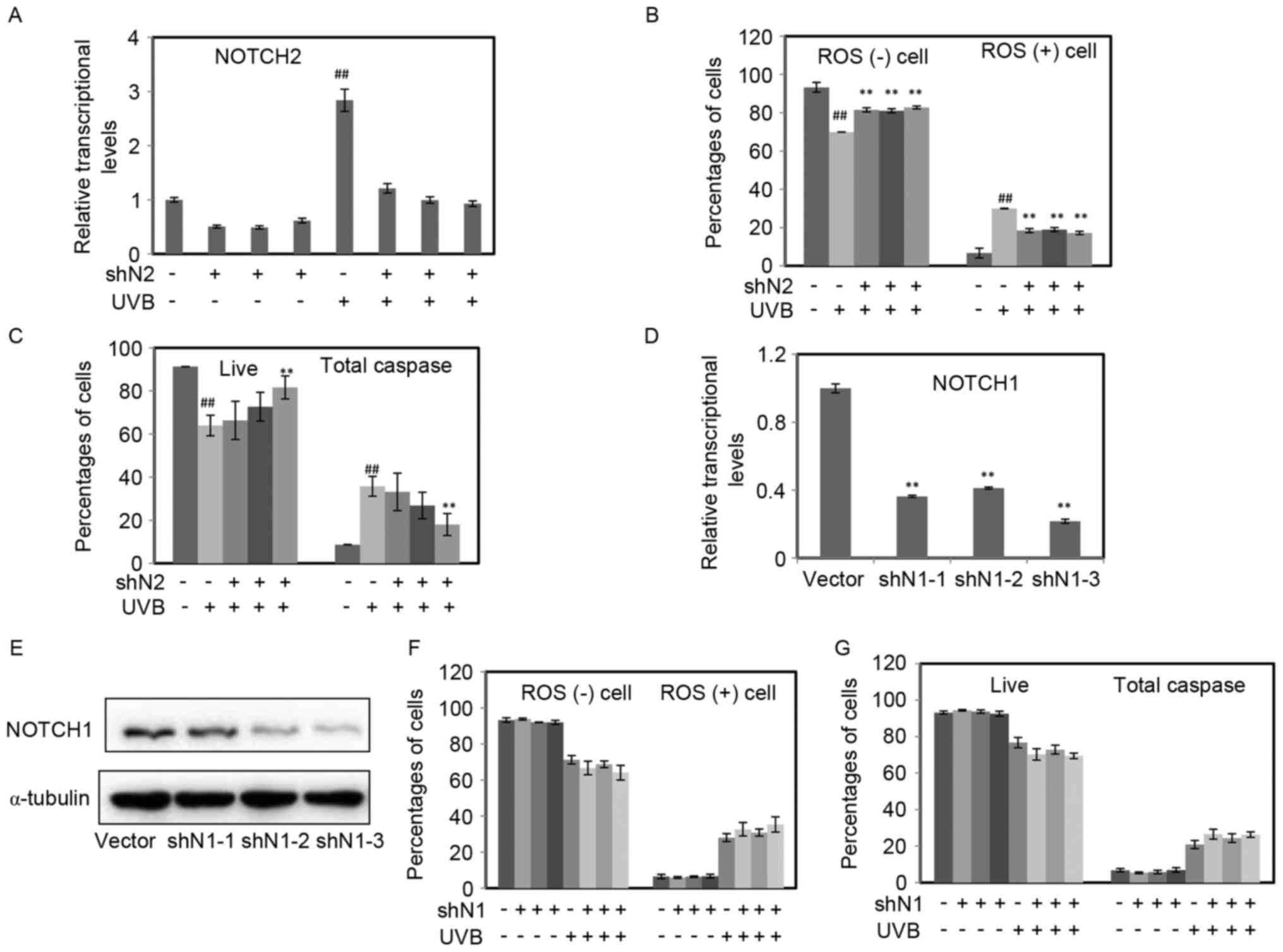
The inhibition of NOTCH2 attenuated
the intracellular ROS and cell apoptosis induced by UVB
To investigate the role of Notch signaling on the
damage induced by UVB, RPE cells with NOTCH1 or
NOTCH2 inhibited were exposed to UVB (50 mJ/cm2),
and then incubated for 36 h. The expression of NOTCH2 in each group
was measured by RT-qPCR. As demonstrated in Fig. 4A, UVB increased NOTCH2,
whereas the inhibition of NOTCH2 was kept at a similar level
to the blank control. The percentage of ROS positive cells was
significantly increased following UVB treatment, nevertheless, the
enhanced effect of UVB was decreased in RPE cells with the
inhibition of NOTCH2 (P<0.01; Fig. 4B). Similar to the above results,
cell apoptosis increased following UVB radiation, and then
decreased because of the inhibition of NOTCH2 (Fig. 4C).
In addition, the authors constructed three stable
cell lines with NOTCH1 inhibited. The efficiency of
NOTCH1 knockdown was determined by RT-qPCR (Fig. 4D) and western blot analysis
(Fig. 4E). The inhibition of
NOTCH1 had no effects on intracellular ROS and cell
apoptosis, moreover, it could not protect RPE cells from UVB
induced damage (P>0.05; Fig. 4F and
G).
Discussion
The present study provided evidence that UVB induced
damage and activated Notch signaling in RPE cells, and the
inhibition of NOTCH2, which presented the highest expression
among various Notch receptors, reduced the damage induced by UVB.
To the best of the authors' knowledge, the effect of Notch
signaling on the protection of RPE cells against UVB damage has not
yet been reported.
It was previously demonstrated that UVB exposure
increased (22) or downregulated
(23) the Notch signaling in
keratinocytes. These contrary results urged us to investigate the
change of Notch signaling and the mechanism upon UVB stimulation in
RPE cells. Interestingly, in the current study, the exposure of UVB
activated the Notch signaling in RPE cells. But the mechanism
requires further research. To investigate the function of Notch
signaling in RPE cells, the expression of all of 13 key components
of Notch signaling were determined. It was discovered that in RPE
cells NOTCH2 demonstrated the highest expression among various
Notch receptors. NOTCH2 is strongly expressed in the pigmented
epithelium of eye, including the RPE, but the investigation of
NOTCH2 only focused on the roles in development and morphogenesis
(24–26). In the present study, the inhibition
of NOTCH2 had no impacts on intracellular ROS and cell
apoptosis under normal conditions. However, in the presence of UVB,
the inhibition of NOTCH2 attenuated intracellular ROS and
cell apoptosis.
More interestingly, the inhibition of NOTCH2,
but not NOTCH1, attenuated intracellular ROS and cell
apoptosis in RPE cells stimulated with UVB. However, in
keratinocytes, the apoptotic response is increased by deletion of
the NOTCH1 gene (22). The
results indicated above could be explained by the fact that
although NOTCH1 and NOTCH2 are closely related paralogs and
function through the same canonical signaling, they contribute to
different outcomes in cell and disease contexts.
In the current study, the inhibition of
NOTCH2 reduced the cell growth and cell migration. Based on
previous work of the authors, blockage of NOTCH1 also
inhibited the migration and proliferation of RPE cells (27). The increase of cell migration is
linked to epithelial to mesenchymal transition leading to wet AMD
and proliferative vitreoretinopathy (PVR). The downregulation of
cleaved NOTCH1 blocked the activation of migration-related
signaling molecules (28). It
revealed that the blockage of Notch signaling may contribute to the
treatment of wet AMD and PVR.
As shown in the present study and in a previous
report (29), the exposure of UVB
increased intracellular ROS and induced cell apoptosis in RPE
cells. It was reported that RPE cells exposed to UVB exhibit
several cellular pathological features, such as the reduction of
cell viability, loss of phagocytotic activity and activation of
inflammatory signaling, which are features of dry AMD (30,31).
Some of these effects may be mediated through the production of
ROS. UVB usually produces ROS and DNA damage. Beyond the repair
capacity, both them may activate signaling pathways which determine
the death or survival of a cell (32,33).
Notch signaling was key regulator of CNV and a
molecular target for therapy in wet AMD (28). However, there are no explicit
evidences studying the function of Notch signaling in the
pathogenesis and therapies of dry AMD. To date, the therapy
available for dry AMD is an intake of antioxidant formulations,
which presents limited efficacy. It is hoped that, through these
efforts, understanding the therapies of AMD will sooner be
elucidated.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81270914, 81670874
and 81170891 to Professor Huangxuan Shen). The authors would like
to thank Professor Lizi Wu (UF Health Shands Hospital, University
of Florida, Gainesville, FL, USA), who provided the plasmid and
helped to review the paper.
References
|
1
|
Clemons TE, Milton RC, Klein R, Seddon JM
and Ferris FL III: Age-Related Eye Disease Study Research Group:
Risk factors for the incidence of advanced age-related macular
degeneration in the age-related eye disease study (AREDS) AREDS
report no. 19. Ophthalmology. 112:533–539. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Biarnés M, Monés J, Alonso J and Arias L:
Update on geographic atrophy in age-related macular degeneration.
Optom Vis Sci. 88:881–889. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Klein R, Klein BE, Knudtson MD, Meuer SM,
Swift M and Gangnon RE: Fifteen-year cumulative incidence of
age-related macular degeneration: The beaver dam eye study.
Ophthalmology. 114:253–262. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Klein R, Klein BE, Knudtson MD, Wong TY,
Cotch MF, Liu K, Burke G, Saad MF and Jacobs DR Jr: Prevalence of
age-related macular degeneration in 4 racial/ethnic groups in the
multi-ethnic study of atherosclerosis. Ophthalmology. 113:373–380.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Thornton J, Edwards R, Mitchell P,
Harrison RA, Buchan I and Kelly SP: Smoking and age-related macular
degeneration: A review of association. Eye (Lond). 19:935–944.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yates JR, Sepp T, Matharu BK, Khan JC,
Thurlby DA, Shahid H, Clayton DG, Hayward C, Morgan J, Wright AF,
et al: Complement C3 variant and the risk of age-related macular
degeneration. N Engl J Med. 357:553–561. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ambati J, Atkinson JP and Gelfand BD:
Immunology of age-related macular degeneration. Nat Rev Immunol.
13:438–451. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tarallo V, Hirano Y, Gelfand BD, Dridi S,
Kerur N, Kim Y, Cho WG, Kaneko H, Fowler BJ, Bogdanovich S, et al:
DICER1 loss and Alu RNA induce age-related macular
degeneration via the NLRP3 inflammasome and MyD88. Cell.
149:847–859. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kraljević Pavelić S, Klobučar M, Sedić M,
Micek V, Gehrig P, Grossman J, Pavelić K and Vojniković B:
UV-induced retinal proteome changes in the rat model of age-related
macular degeneration. Biochim Biophys Acta. 1852:1833–1845. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kerr JB and McElroy CT: Evidence for large
upward trends of ultraviolet-B radiation linked to ozone depletion.
Science. 262:1032–1034. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Forest DL, Johnson LV and Clegg DO:
Cellular models and therapies for age-related macular degeneration.
Dis Model Mech. 8:421–427. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Strauss O: The retinal pigment epithelium
in visual function. Physiol Rev. 85:845–881. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cruickshanks KJ, Klein R, Klein BE and
Nondahl DM: Sunlight and the 5-year incidence of early age-related
maculopathy: The beaver dam eye study. Arch Ophthalmol.
119:246–250. 2001.PubMed/NCBI
|
|
14
|
Artavanis-Tsakonas S, Rand MD and Lake RJ:
Notch signaling: Cell fate control and signal integration in
development. Science. 284:770–776. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kopan R and Ilagan MX: The canonical Notch
signaling pathway: Unfolding the activation mechanism. Cell.
137:216–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schouwey K, Aydin IT, Radtke F and
Beermann F: RBP- Jk-dependent Notch signaling enhances retinal
pigment epithelial cell proliferation in transgenic mice. Oncogene.
30:313–322. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bolós V, Grego-Bessa J and de la Pompa JL:
Notch signaling in development and cancer. Endocr Rev. 28:339–363.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen J, Kesari S, Rooney C, Strack PR,
Chen J, Shen H, Wu L and Griffin JD: Inhibition of notch signaling
blocks growth of glioblastoma cell lines and tumor neurospheres.
Genes Cancer. 1:822–835. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lin S, Tian L, Shen H, Gu Y, Li JL, Chen
Z, Sun X, You MJ and Wu L: DDX5 is a positive regulator of
oncogenic NOTCH1 signaling in T cell acute lymphoblastic leukemia.
Oncogene. 32:4845–4853. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen Z, Jin G, Lin S, Lin X, Gu Y, Zhu Y,
Hu C, Zhang Q, Wu L and Shen H: DNA methyltransferase inhibitor
CDA-II inhibits myogenic differentiation. Biochem Biophys Res
Commun. 422:522–526. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vatsyayan R, Lelsani PC, Chaudhary P,
Kumar S, Awasthi S and Awasthi YC: The expression and function of
vascular endothelial growth factor in retinal pigment epithelial
(RPE) cells is regulated by 4-hydroxynonenal (HNE) and glutathione
S-transferaseA4-4. Biochem Biophys Res Commun. 417:346–351. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mandinova A, Lefort K, di Vignano Tommasi
A, Stonely W, Ostano P, Chiorino G, Iwaki H, Nakanishi J and Dotto
GP: The FoxO3a gene is a key negative target of canonical Notch
signalling in the keratinocyte UVB response. EMBO J. 27:1243–1254.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fukunaga-Kalabis M, Hristova DM, Wang JX,
Li L, Heppt MV, Wei Z, Gyurdieva A, Webster MR, Oka M, Weeraratna
AT and Herlyn M: UV-induced Wnt7a in the human skin
microenvironment specifies the fate of neural crest-like cells via
suppression of Notch. J Invest Dermatol. 135:1521–1532. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bao ZZ and Cepko CL: The expression and
function of Notch pathway genes in the developing rat eye. J
Neurosci. 17:1425–1434. 1997.PubMed/NCBI
|
|
25
|
Zhou Y, Tanzie C, Yan Z, Chen S, Duncan M,
Gaudenz K, Li H, Seidel C, Lewis B, Moran A, et al: Notch2
regulates BMP signaling and epithelial morphogenesis in the ciliary
body of the mouse eye. Proc Natl Acad Sci USA. 110:8966–8971. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
McCright B, Gao X, Shen L, Lozier J, Lan
Y, Maguire M, Herzlinger D, Weinmaster G, Jiang R and Gridley T:
Defects in development of the kidney, heart and eye vasculature in
mice homozygous for a hypomorphic Notch2 mutation. Development.
128:491–502. 2001.PubMed/NCBI
|
|
27
|
Liu W, Jin G, Long C, Zhou X, Tang Y,
Huang S, Kuang X, Wu L, Zhang Q and Shen H: Blockage of Notch
signaling inhibits the migration and proliferation of retinal
pigment epithelial cells. ScientificWorldJournal. 2013:1787082013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Park GB, Kim D, Kim YS, Kim JW, Sun H, Roh
KH, Yang JW and Hur DY: Regulation of ADAM10 and ADAM17 by
sorafenib inhibits epithelial-to-mesenchymal transition in
Epstein-Barr virus-infected retinal pigment epithelial cells.
Invest Ophthalmol Vis Sci. 56:5162–5173. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Balaiya S, Murthy RK, Brar VS and Chalam
KV: Evaluation of ultraviolet light toxicity on cultured retinal
pigment epithelial and retinal ganglion cells. Clin Ophthalmol.
4:33–39. 2010.PubMed/NCBI
|
|
30
|
Roduit R and Schorderet DF: MAP kinase
pathways in UV-induced apoptosis of retinal pigment epithelium
ARPE19 cells. Apoptosis. 13:343–353. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Roehlecke C, Schaller A, Knels L and Funk
RH: The influence of sublethal blue light exposure on human RPE
cells. Mol Vis. 15:1929–1938. 2009.PubMed/NCBI
|
|
32
|
Kulms D and Schwarz T: Independent
contribution of three different pathways to ultraviolet-B-induced
apoptosis. Biochem Pharmacol. 64:837–841. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Valerie K, Yacoub A, Hagan MP, Curiel DT,
Fisher PB, Grant S and Dent P: Radiation-induced cell signaling:
Inside-out and outside-in. Mol Cancer Ther. 6:789–801. 2007.
View Article : Google Scholar : PubMed/NCBI
|