Introduction
In China, the incidence of hypertension in surgical
patients has reached 20% (1). In
addition, Prys-Roberts (2)
identified that hypertension in surgical patients is 24% in
Britain. During anesthesia, hypertensive patients, in particular
the elderly, are extremely prone to fluctuations in blood pressure.
If blood pressure rises or falls by 30% away from the baseline
value, this will lead to serious complications (stroke, heart or
kidney failure etc.) and even death. Therefore, in the anesthetic
management of these patients, particular attention should be given
to maintain a stable blood pressure, and analgesics for minimizing
fluctuations in blood pressure are very important. Remifentanil
(RMF), a new ultra short effect of the µ-opioid receptor agonist,
is used for the induction and maintenance of general anesthesia.
Loading or maintenance doses of RMF can decrease peripheral
vascular resistance, thereby causing hypotension (3–6),
which may be directly associated with the relaxation of vascular
smooth muscles (7,8). Unlugenc et al (9) stated that RMF exerted a dilation
effect on isolated thoracic aortas of rats. Paris et al
(10) demonstrated that the
cerebral blood flow velocity was reduced by large doses of RMF. By
contrast, Engelhard et al (11) showed that after continuous infusion
of RMF, the mean arterial pressure, intracranial pressure, and
cerebral blood flow velocity of patients did not change. Thus, the
specific mechanisms and effects of RMF on the cerebrovascular
system remain unclear. Large-conductance calcium-activated
potassium channels (BKCa) and voltage-gated potassium channels (Kv)
are two important potassium ion channels in the vascular
smooth-muscle cell (VSMC) membrane. When the VSMC membrane
potential depolarization occurred, the opening probability of BKCa
and Kv channels increased, and the intracellular potassium efflux
also increased; the cell then hyperpolarized, the opening of
L-type-calcium channel became limited, thereby reducing the calcium
influx and decreasing the intracellular calcium concentration,
leading to vasodilation (12,13).
In the current study, the pressure myograph system and whole-cell
patch-clamp technique were used to observe the effects of RMF on
the diameter and smooth-muscle cells (SMCs) of isolated cerebral
basilar arteries (BAs) of spontaneously hypertensive rats (SHR) and
Wistar-Kyoto (WKY) rats. The effects on the BAs and the mechanisms
involved were also investigated to provide a better basis for
clinical use of these two groups of patients undergoing cranial
operation.
Materials and methods
Animals
A total of 60 SHR and 60 WKY rats were purchased
from Beijing Vital River Laboratory Animal Technology Co., Ltd.
(Beijing) [Animal certificate of conformity: SCXK (Beijing)
2012–0001, weighing ~200–250 g, aged 16–20 weeks old, male or
female]. Rats were housed in separate cages in a specific
pathogen-free environment at 24±3°C, relative humidity of 40–70%,
in a 12 h light-dark cycle, and were provided with free access to
food and water. All protocols were approved by the Institutional
Animal Care and Use Committee (IACUC) at the Medical College of
Shihezi University and consistent with the Guidelines for the Care
and Use of Laboratory Animals published by the US National
Institutes of Health (14).
Reagents
RMF was purchased from Hubei Yichang People Fook
Pharmaceutical Co., Ltd. (batch number, 6141211; Yichang, China).
Phenylephrine (PE), acetylcholine (ACh), ethylenediaminetetraacetic
acid (EDTA), tetraethylammonium (TEA), 4-aminopyridine (4-AP),
collagenase, papain, bovine serum albumin (BSA), DTT, and DMEM
culture medium were purchased from Sigma-Aldrich (Merck Millipore,
Darmstadt, Germany). KCl and other reagents were acquired locally.
All reagents used in the pressure myograph system and whole-cell
patch-clamp technique were prepared using sugar-free physiological
saline solution (PSS). Extracellular solution was a stock sample
prepared before being further diluted with external solution to
achieve the final concentration. The formulas of PSS/saline
solution with high kalium and the external solution were in
accordance with the literature (15,16).
Instruments
Pressure myograph system (110P; Danish Myo
Technology A/S, Aarhus, Denmark), MyoVIEW software (Danish Myo
Technology A/S), Axon MultiClamp 700B patch-clamp amplifier (Axon;
Molecular Devices LLC, Sunnyvale, CA, USA), micromanipulator
(PCS5001; Siskiyou Design, Oregon, USA), P-97 microelectrode
pullers (Sutter Instrument, Novato, CA, USA), heated water bath
(HSS-1B; Chengdu Science Instrument Factory, Chengdu, China), and
multiple perfusion administration system (supplied in-house by
Huazhong University of Science and Technology, China).
Pressure myograph measurement
Cerebral BA (CBA) segments were placed in a 4°C
oxygen-saturated physiological solution containing 118.9 mM NaCl,
4.7 mM KCl, 1.2 mM MgSO4, 1.2 mM
KH2PO4, 2.5 mM CaCl2, 25 mM
NaHCO3 and 5.5 mM glucose. CBA segments were tied to a
glass tube using 12–0 nylon monofilament sutures, and placed in a
microvascular chamber (Pressure Myograph System; Danish Myo
Technology A/S). The chamber was perfused with a physiological
solution (pH 7.4, bubbled with 95% O2 and 5%
CO2) and heated to 37°C. CBA segments were pressurized
to a constant transmural pressure of 60 mmHg. The diameter was
continuously determined and recorded via video dimension analyzer
and the DMT Vessel Acquisition Suite. CBA segments were treated
with progressively increasing doses of RMF (10−4-10 µM),
followed by PE (0.1 mM). The results were evaluated via the changes
in vascular diameter recorded on the DMT (17).
Whole-cell patch-clamp recording
To isolate single SMCs from CBAs, CBAs were
incubated in a low-Ca2+ solution for 20 min containing
142 mM NaCl, 5 mM KCl, 0.05 mM CaCl2, 1.0 mM
MgCl2, 4.0 mM Na-HEPES, 5.0 mM HEPES and 7.5 mM glucose
and cut into 1 mm segments and digested with low-Ca2+
solution containing papain (1 mg/ml), collagenase A (0.5 mg/ml),
BSA (1 mg/ml) and DL-dithiothreitol (1 mg/ml) for 10 min at 37°C.
Specific operations were performed according to the literature
(18).
Conventional whole-cell patch-clamp recording was
performed using an Axon 700B amplifier (Axon; Molecular Devices
LLC) (19). The pipette had a
resistance of approximately 5 MΩ after being filled with internal
solution containing 130 mM K-gluconate, 10 mM NaCl, 2 mM
CaCl2, 1.2 mM MgCl2, 10 mM HEPES, 5 mM
ethylene glycol-bis (β-aminoethyl ether) N,N',N'-tetraacetic acid
and 7.5 mM glucose. The seal resistance commonly reached 1–20 GΩ
prior to rupture of the membrane. The membrane current or voltage
signals were filtered at 10 kHz and recorded on a computer equipped
with a Digidata 1440A AD-interface and pClamp software, version,
10.2 (Axon; Molecular Devices LLC) at a sampling interval of 200
msec.
Statistical analysis
SHR and WKY rats were age-matched to minimize
individual differences. The results are expressed as the mean ±
standard error. Statistical analysis was performed using the SPSS
statistical software package, version 17.0. A two-factor multilevel
analysis of variance was used for repeated measurement data,
followed by the Neuman-Keuls post hoc test. A two-sample t-test was
used between groups, and the paired t-test was applied in the same
group. P<0.05 was considered to indicate a statistically
significant difference.
Results
CBAs of SHR and WKY rats were relaxed
by RMF in a concentration-dependent manner
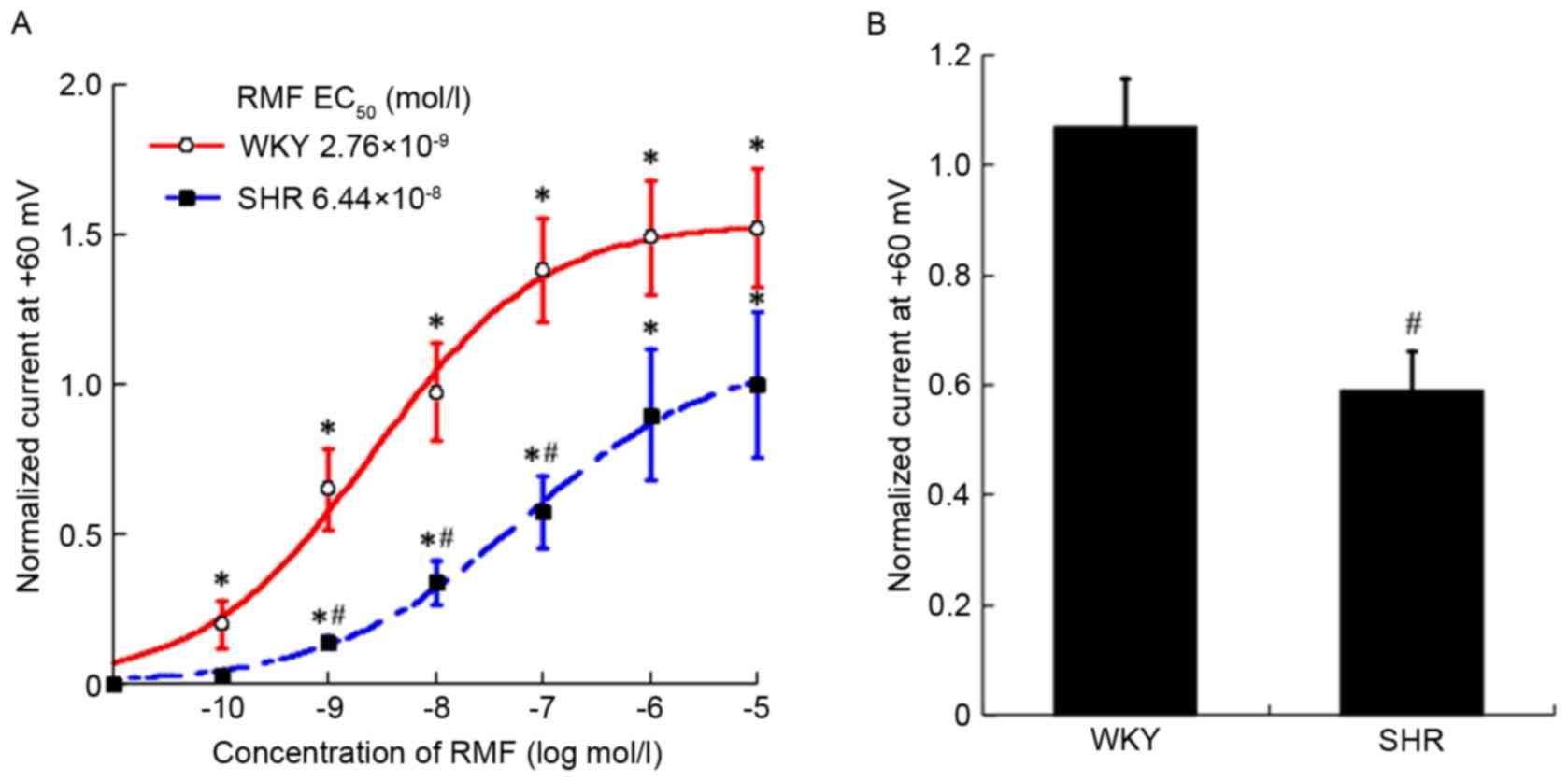
Following measuring the activity to be efficient,
restoring stability, and pre-shrinking by 10−4 mol/l PE
to steady state, the blood vessel diameters of SHR and WKY rats
were 270.11±9.79 and 242±10.27 µm. Subsequent to adding RMF
prepared by 10−4 mol/l PE from low to high
concentrations to the chamber, the diastolic range of SHR with
10−9 to 10−5 mol/l RMF was identified to be
4.29±1.56, 17.65±2.84, 30.06±6.42, 33.86±7.48 and 29.25±7.28 µm
(Fig. 1B); compared with the
pre-administration phase. Excluding the concentration of
10−9 mol/l, all concentrations were statistically
different (n=5; F=12.98; P<0.05; Fig. 1G). However, the diastolic amplitude
of WKY rats with 10−10-10−6 mol/l RMF was
12.39±7.44, 21.05±9.01, 32.13±14.71, 31.28±13.90 and 29.51±15.52
µm, respectively (Fig. 1A); and
compared with those not given RMF, a statistical significance was
observed in all concentrations (n=5; F=11.81; P<0.05; Fig. 1G). Subsequent to drawing the
dose-response curve, it was identified that the EC50
values of CBA of SHR and WKY rats relaxed by RMF were
(4.32±1.22)x10−9 mol/l and (3.09±0.58) ×10−10
mol/l, and the difference was statistically significant (n=5;
P<0.05) (Fig. 1G). Compared
with WKY rats, the vasodilation rate curve of SHR by RMF shifted
right and downwards, which indicated that the relaxation response
of BA in SHR to RMF was weaker than that of WKY rats, and the
difference was statistically significant (n=5; F=20.34; P<0.01;
Fig. 1G).
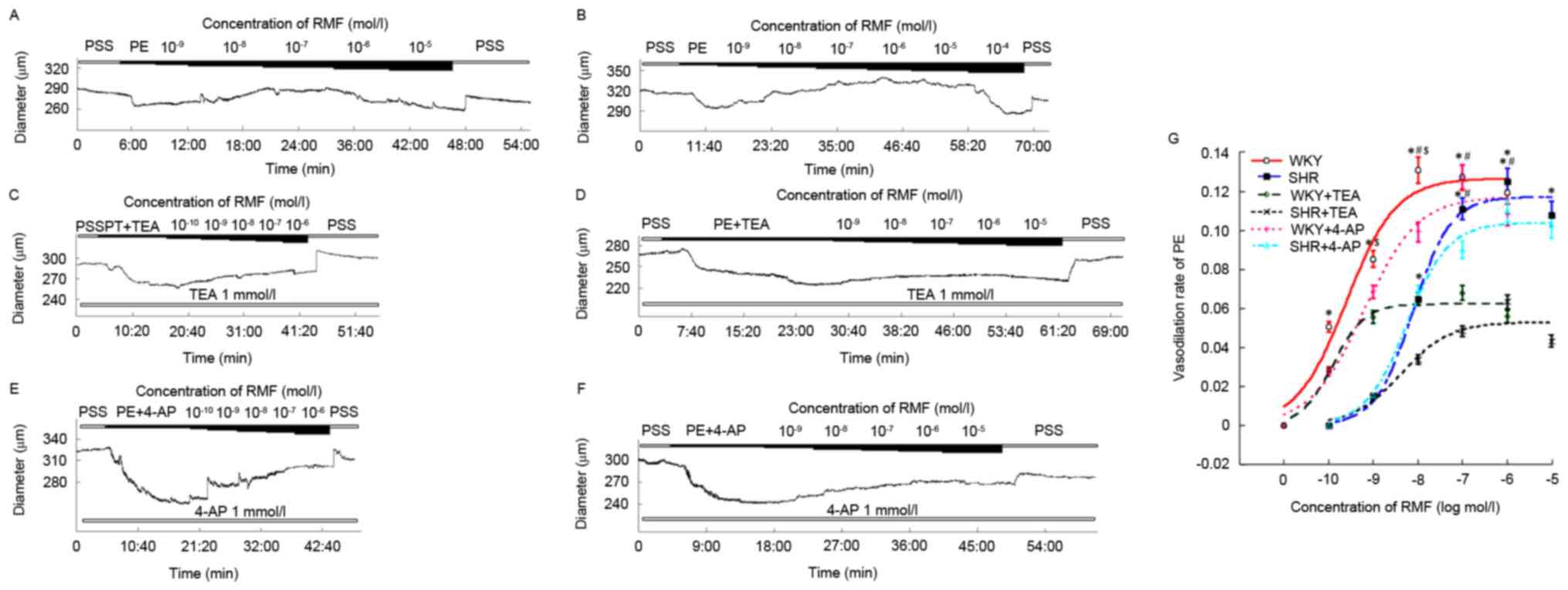 | Figure 1.Diastolic effects of RMF on CBA of
SHR and WKY rats were in a dose-dependent manner, which were
associated with the BKCa channel. The initial image about different
concentrations of RMF on cerebral CBA of WKY (A, C and E) and SHR
(B, D and F) rats. (G) The fitting curves of RMF on SHR and WKY
rats at different concentrations. All data were compared with each
other after being standardized. Results are presented as the mean ±
standard error, n=5 for WKY and SHR, n=7 for SHR+TEA, n=6 for
WKY+TEA, n=6 for SHR+4-AP and n=9 for WKY+4-AP. *P<0.05, vs.
pre-administration; #P<0.05, WKY vs. WKY + TEA, SHR
vs. SHR + TEA; $P<0.05, WKY vs. SHR. RMF,
remifentanil; CBA, cerebral basilar artery; SHR, spontaneously
hypertensive rats; WKY, Wistar-Kyoto rats; TEA, tetraethylammonium;
4-AP, 4-aminopyridine. |
The diastolic effects of RMF on CBA of
SHR and WKY rats were mediated by the BKCa channel
Following pre-incubation of the mixture of 1 mmol/l
TEA and 10−4 mol/l PE for 20 min to a steady state, the
vessel diameters of SHR and WKY rats were 231.62±5.26 and
239.75±7.04 µm, respectively. Successively adding RMF formulated by
1 mmol/l TEA and 10−4 mol/l PE from a low to high
concentration to the chamber, it was identified that the diastolic
range of SHR by 10−9-10−5 mol/l RMF was
3.74±0.59, 8.58±2.67, 12.14±3.05, 15.89±4.07 and 11.37±5.39 µm,
respectively (Fig. 1D); while the
diastolic amplitude of WKY rats with
10−10-10−6 mol/l RMF was 6.78±1.26,
13.34±1.94, 15.59±2.09, 16.34±2.37 and 13.49±2.53 µm, respectively
(Fig. 1C). Although different
concentrations of RMF produced diastolic effects on BAs in a
dose-dependent manner, compared with no administration of TEA, the
relaxation rate curve of SHR shifted down during
10−8-10−5 mol/l, and a statistically
significant difference was identified at the concentrations of
10−7 and 10−6 mol/l (n=7; F=15.47; P<0.05;
Fig. 1G). However, the
vasodilation rate curve of WKY rats significantly shifted down
during 10−9-10−6 mol/l, and the statistical
difference was observed with the concentrations of 10−8
and 10−7 mol/l (n=6; F=22.36; P<0.05; Fig. 1G). These indicated that CBA of SHR
and WKY rats were relaxed by RMF, which was associated with the
opening of BKCa channels. Prior and subsequent to incubation with
TEA, the EC50 value of CBA of SHR relaxed by RMF were
(4.32±1.22)x10−9 mol/l and (7.54±3.17)x10−9
mol/l, and the difference between them was not statistically
significant (n=7; P>0.05; Fig.
1G); while the EC50 value of WKY rats by RMF were
(3.09±0.58)x10−10 mol/l and (2.26±0.55)x10−10
mol/l, and neither was statistically different (n=6; P>0.05;
Fig. 1G).
RMF relaxed CBA, which was not related
to Kv channel
Following pre-incubation of BA with the mixture of 1
mmol/l 4-AP and 10−4 mol/l PE for 20 min to a steady
state, the diameters of SHR and WKY rats were 257.67±5.36 and
273.93±12.79 µm, respectively. Subsequently, RMF that had been
prepared with 1 mmol/l 4-AP and 10−4 mol/l PE was added
to the chamber from a low to high concentration, and it was
identified that the diastolic amplitudes of BA of SHR with
10−9-10−5 mol/l RMF were 6.73±0.77,
17.12±4.48, 22.64±4.99, 27.6±5.36 and 26.20±6.07 µm, respectively
(Fig. 1F); while the diastolic
range of WKY rats with 10−10-10−6 mol/l RMF
were 7.43±1.72, 17.71±3.51, 25.63±4.33, 34.93±5.25 and 29.66±4.53
µm, respectively (Fig. 1E).
Different concentrations of RMF still caused diastolic reactions on
the vessels of SHR and WKY rats in a dose-dependent manner.
Compared with pre-incubation without 4-AP, the diastolic rate
curves of SHR and WKY rats marginally moved down, and no
significant difference was observed (n=6 and 9; F=2.13 and 2.59;
P>0.05; Fig. 1G). Prior and
subsequent to incubation with 4-AP, the EC50 values of
RMF on BA of SHR were (4.32±1.22)x10−9 mol/l and
(4.12±1.16)x10−9 mol/l, and the difference between them
was not statistically significant (n=6; P>0.05; Fig. 1G). The EC50 values of
WKY rats produced by RMF were (3.09±0.58)x10−10 mol/l
and (10.75±3.41)x10−10 mol/l, and neither were
statistically different (n=9; P>0.05; Fig. 1G).
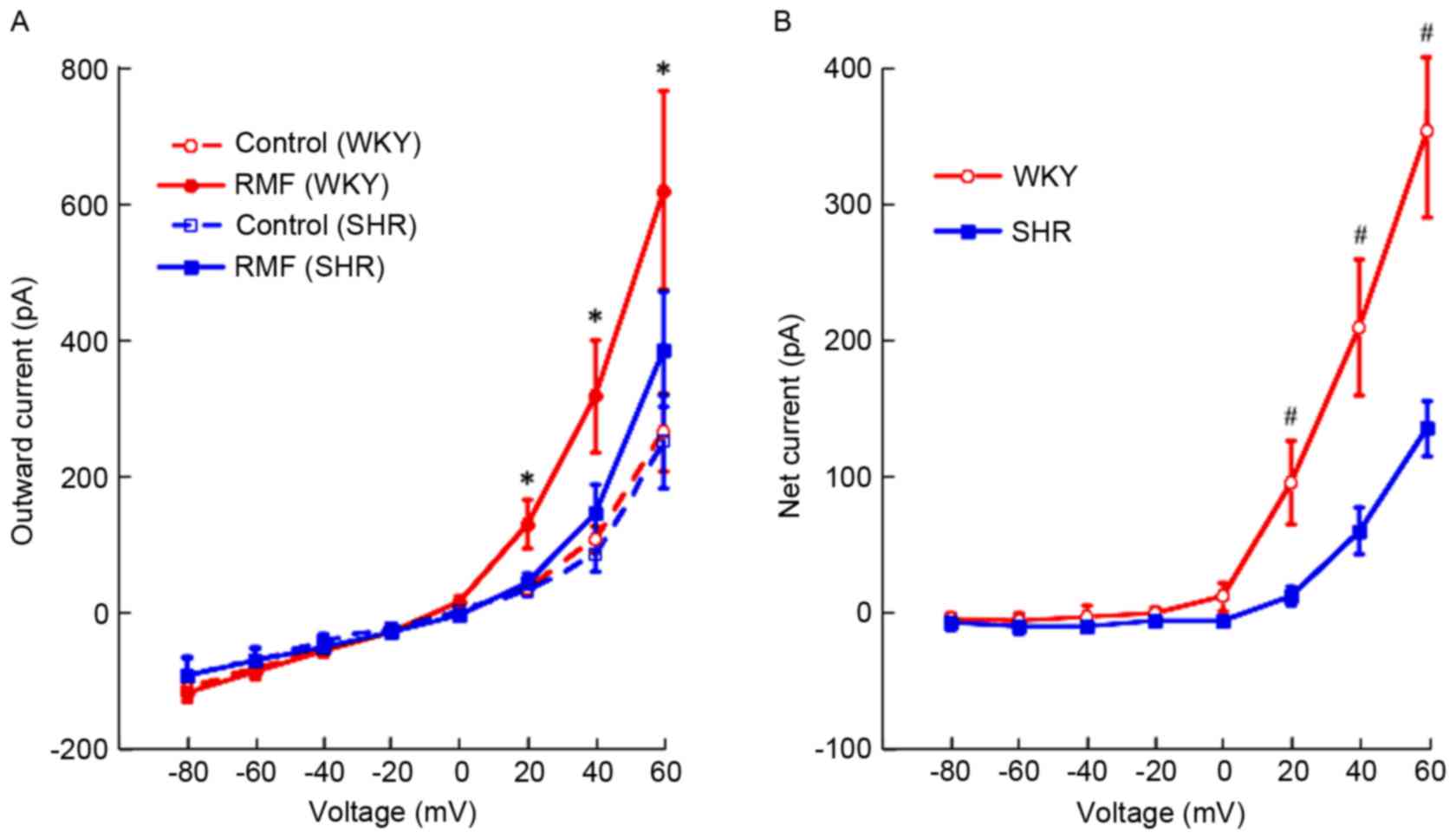
Outward currents were increased by RMF
in BASMCs of rats in a voltage-dependent manner
At 0 to 60 mV, RMF (3×10−7 mol/l)
enhanced the outward currents in BASMCs of SHR and WKY rats in a
voltage-dependent manner, however no significant difference at 0 mV
was observed (Fig. 2A). In
addition, the increasing effect of RMF on outward currents in
BASMCs of SHR was weaker than for that of WKY rats with the same
voltage (Fig. 2B).
Outward currents were increased by RMF
in BASMCs of rats in a dose-dependent manner
Approximately 10−10-10−5 mol/l
RMF made outward currents (+60 mV) in BASMCs of SHR increase from
317±33 to 326±30, 363±28, 430±38, 504±32, 572±72 and 619±89 pA,
respectively; the outward currents in BASMCs of WKY rats were from
381±68 to 542±49, 632±45, 721±58, 859±69, 1217±19 and 1610±50 pA,
respectively. Subsequent to fitting the dose-response curve, the
EC50 values of the enhancing effect of RMF on outward
currents in BASMCs of SHR and WKY rats were
(9.58±5.1)x10−8 mol/l and (2.93±1.4)x10−9
mol/l, and the two exhibited a statistically significant difference
(n=6; P<0.01). Compared with pre-administration, the outward
currents of SHR were enhanced in a dose-dependent manner, with
exception of the concentration of 10−10 mol/l (n=6;
F=13.21; P<0.05). These effects were additionally observed in
the WKY rats at all concentrations (n=6; F=21.57; P<0.05).
Compared with WKY rats, the increasing effect of RMF on outward
currents of SHR was weaker at the same concentration. In addition
to 10−10, 10−6 and 10−5 mol/l, the
remaining concentrations had a statistically significant difference
(n=6; F=8.65; P<0.05; Fig. 3A).
Fig. 3B presents the comparison of
the effect of RMF (3×10−7 mol/l) on outward currents in
BASMCs of SHR and WKY rats, which suggested that the relaxant
effect on CBA of SHR by RMF was weaker in WKY rats (n=6;
P<0.05).
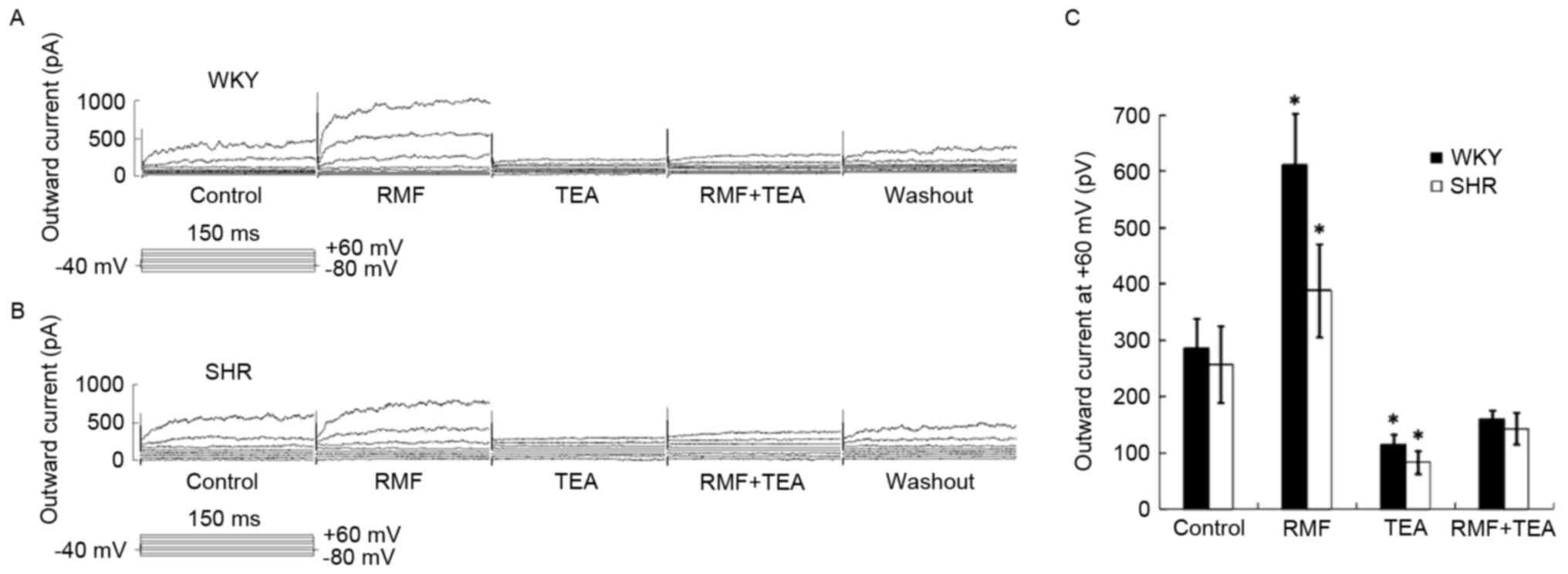
The enhanced effect of RMF on BASMCs
outward current was mediated by the BKCa channel
Approximately 10−3 mol/l TEA was
pre-perfused for 3 min. The mixture of 10−3 mol/l TEA
and 3×10−7 mol/l RMF was administered. The increasing
effect of RMF on BASMCs outward current was partially blocked and
outward current (at +60 mV) of WKY rats reduced to 115.15±19.88 pA,
which was only 0.44±0.03 times higher than that of the control
group (n=6; P<0.05; Fig. 4A);
while outward current of SHR decreased to 83.21±20.69 pA, which was
only 0.33±0.02 times higher than that of the control group (n=6;
P<0.05; Fig. 4B); these
indicated that the BKCa channel-mediated outward current
contributed to the outward current of the BASMCs of SHR and WKY
rats induced by RMF (Fig. 4C). In
addition, the inhibition rate of TEA on IBKCa in BASMCs
of SHR and WKY rats was 0.67±0.02 and 0.56±0.03, and the difference
between them was statistically significant (n=6, P<0.05), which
suggested that the influence of BKCa channels in the BASMCs of SHR
was more than in WKY rats.
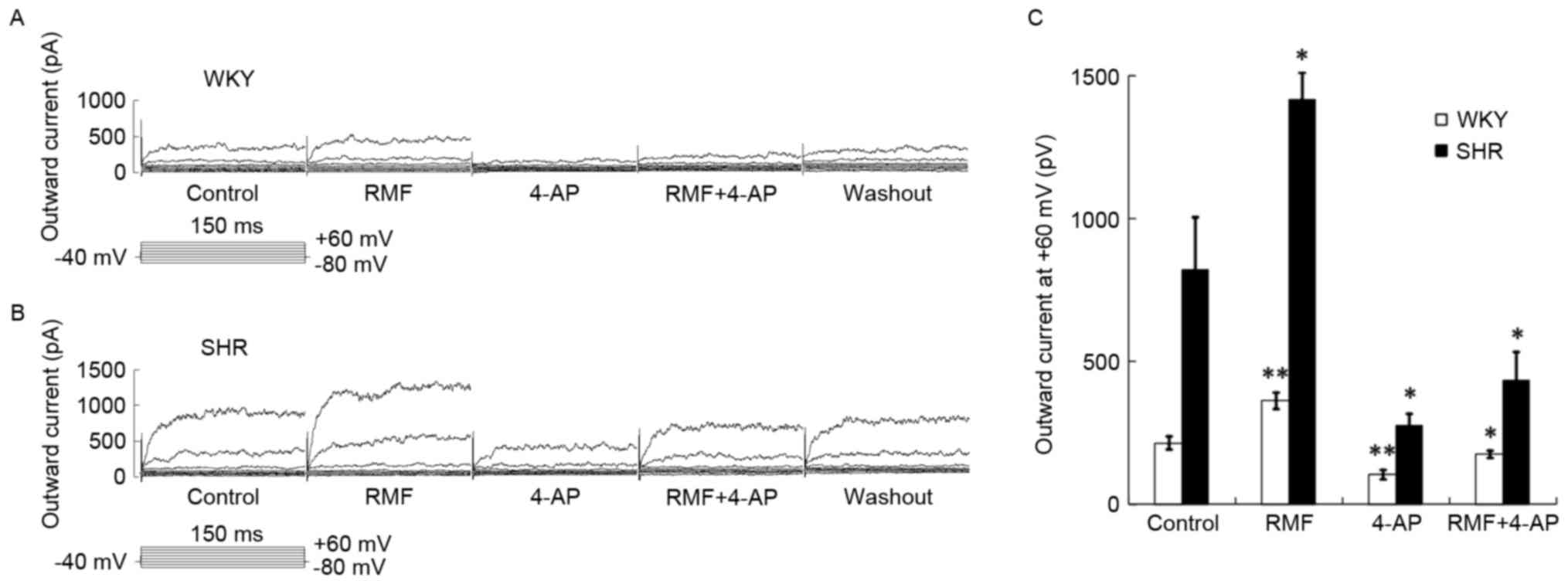
Enhanced effect of RMF on BASMCs
outward current was partially blocked by 4-AP (Kv channel
blocker)
Approximately 10−3 mol/l of 4-AP was
pre-perfused for 3 min, and the mixture of 10−3 mol/l
4-AP and 3×10−7 mol/l RMF was administered. The
increasing effect of RMF on the BASMCs outward current was still
partially blocked, and the outward current (at +60 mV) of WKY rats
was reduced to 105.12±12.18 pA, which was 0.52±0.04 times greater
than that of the control group (n=9; P<0.01) (Fig. 5A); while outward current of SHR
decreased to 274.96±81.92 pA, which was only 0.36±0.05 times higher
than that of the control group (n=6; P<0.05; Fig. 5B); these suggested that the outward
current of the BASMCs of SHR and WKY rats were enhanced by RMF, and
involved the outward current mediated by Kv channel (Fig. 5C).
Enhanced effect of RMF on BASMCs
outward current was totally inhibited by the mixture of TEA and
4-AP
The mixture of 10−3 mol/l TEA and 4-AP
was pre-perfused for 3 min, and the mixture of 10−3
mol/l TEA, 4-AP and 3×10−7 mol/l RMF were administered.
The increasing effect of RMF on BASMCs outward current was totally
inhibited and outward current (at +60 mV) of WKY rats reduced to
139.69±18.04 pA, which was 0.32±0.034 times higher than that of the
control group (n=7; P<0.05; Fig.
6A); while outward current of SHR decreased to 99.23±27.23 pA,
which was only 0.32±0.04 times higher than that of the control
group (n=7; P<0.05; Fig. 6B);
these suggested that the enhanced effects of RMF on BASMCs outward
current of SHR and WKY rats were completely and collectively
mediated by BKCa and Kv channels (Fig.
6C).
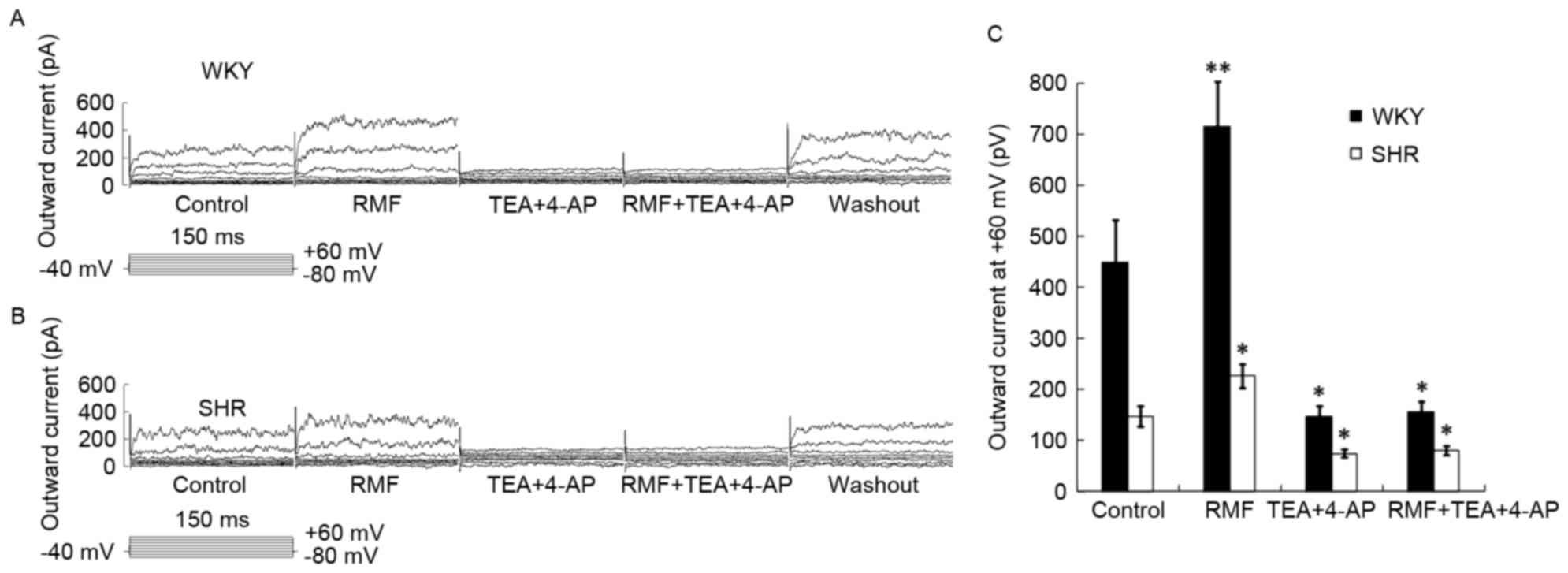 | Figure 6.Enhanced effect of RMF can be totally
blocked by 10−3 mol/l TEA and 4-AP. Raw current images
with respect to the effect of RMF, TEA and 4-AP on (A) WKY and (B)
SHR rats. (C) Bar chart showing the effect of RMF, TEA and 4-AP on
SHR and WKY rats. Results are presented as the mean ± standard
error, n=7. *P<0.05, **P<0.01, vs. control group. RMF,
remifentanil; TEA, tetraethylammonium; 4-AP, 4-aminopyridine; WKY,
Wistar-Kyoto rats; SHR, spontaneously hypertensive rats. |
Discussion
In the current study, RMF was observed to relax BA
in a concentration-dependent manner, however this can be suppressed
by TEA. The results suggested that the relaxant effect of RMF on BA
was ascribed to the BKCa channel activity. The whole-cell
patch-clamp technique was used to show that RMF enhanced
IBKCa and IKv in BASMCs in a voltage- and
dose-dependent manner. In conclusion, RMF served a diastolic role
in the basilar arteries of rats, likely via activation of BKCa and
Kv channels. The two techniques in the present study demonstrated
different responses to the inhibitory effects of 4-AP on
vasodilation of the RMF, possibly due to different drug
sensitivities at tissue and cellular levels.
K+ efflux results in smooth-muscle cell
membrane hyperpolarization following channel opening, and it
additionally limits the opening of the L-type-calcium channel,
reducing Ca2+ influx, decreasing the intracellular
calcium concentration, and thus leading to vasodilation (12,13).
A previous study confirmed that BKCa and Kv channels exist in the
BASMCs of rats (16). Lin et
al (20) identified that RMF
relaxed human mesenteric arteries by activating BKCa channels in
SMCs in a concentration-dependent manner via whole-cell patch-clamp
technique, which was in agreement with the results of the current
study.
However, in view of the current literature, the
effects of RMF on cerebral arteries remain controversial. Paris
et al (10) identified that
RMF reduced the cerebral blood flow velocity with large doses,
however Engelhard et al (11) suggested that the mean arterial
pressure, intracranial pressure, and cerebral blood flow velocity
of patients remained unchanged following continuous infusion of
RMF. The current study demonstrated that RMF relaxed the BA of rats
in vitro, and this inconsistency may be attributed to the
following: More uncontrollable factors of macro-indicators detected
in clinical studies, differences between the species, or possible
existence of contraction mechanisms of RMF on BA which remain to be
identified. The effect of this contraction mechanism should be
greater than the relaxation mechanism, and it should be a new
direction for future research.
In addition, two opposite views were presented on
the comparison between SHR and WKY rats from local and
international studies. Among these views, Liu et al
(21) showed that the KCa current
density sensitive to IBTX in cerebral artery smooth-muscle cell
membrane of SHR was 4.7 times higher than that of WKY rats, and Hu
et al (22) identified that
the BKCa current density of SHR was 2.5 times higher than that of
WKY rats. Other views were represented by Yang et al
(23); they suggested the
functions of BKCa channels in mesenteric artery smooth-muscle cells
of Han hypertensive patients. It was observed that whole-cell
current density, spontaneous transient outward currents, and the
Ca2+ sensitivity of hypertensive patients were
significantly reduced (23). In
the current study, the BKCa channel activity in BASMCs of SHR was
higher than those in WKY rats. Yang et al (23) presented a different reason
regarding the difference between experimental objects. In the
current study, it was concluded that the diastolic effect of RMF on
BASMCs of SHR was weaker than that on WKY rats, and this
observation may be ascribed to SHR being less sensitive than WKY
rats; RMF served a role in SHR via another contraction mechanism,
which required further experiments to be confirmed. It was
hypothesized that RMF may serve a diastolic role in the BA of
normotensive patients and produce an increase in cerebral blood
flow, which increases intracranial pressure. When intracranial
pressure exceeded 30~40 mmHg, cerebral blood flow declined, and
eventually caused cerebral ischemia and hypoxia, which can lead to
the formation of brain midline shift or herniation in severe cases
(24). However, the diastolic
effect of RMF on cerebral arteries of hypertensive patients was
weaker than in normotensive ones, which was more likely to cause
serious complications in chronic hypertension patients with risk
for ischemia and hypoxia. Therefore, RMF should be carefully used
in hypertensive patients with increased intracranial pressure
(traumatic brain injury) under anesthesia.
RMF is a new µ-opioid receptor agonist with ultra
short effect. Previous studies have indicated that the opioid
peptides significantly increased the whole-cell IBKCa
and the opening probability of BKCa channels and inhibited
Ca+ channels of neurons and chromaffin cells through the
opioid receptors, which indicated a tight coupling between
receptors and channels (25,26).
In addition, G-protein-coupled receptors can exist in equilibrium
with active and inactive states (27). Consequently, it was hypothesized
that RMF was likely to serve an activation effect on BKCa and Kv
channels via G-protein-coupled approach, however, the specific
mechanisms require further elucidation.
In summary, RMF served a diastolic role in the CBA
of rats, likely by activating BKCa and Kv channels in avoltage- and
dose-dependent manner. SHR demonstrated a weaker diastolic reaction
to RMF than WKY rats. These results may provide guidance regarding
the two groups, particularly in hypertensive patients undergoing
cranial surgery.
References
|
1
|
Guo J, Yu CQ, Lyu J, Guo Y, Bian Z, Zhou
H, Tan Y, Pei P, Chen J, Chen Z, et al: Status of prevalence,
awareness, treatment and controll on hypertension among adults in
10 regions, China. Zhonghua Liu Xing Bing Xue Za Zhi. 37:469–474.
2016.(In Chinese). PubMed/NCBI
|
|
2
|
Prys-Rroberts C: Anaesthesia and
hypertension. Br J Anaesth. 56:711–724. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Elliott P, O'Hare R, Bill KM, Phillips AS,
Gibson FM and Mirakhur RK: Severe cardiovascular depression with
remifentanil. Anesth Analg. 91:58–61. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sebel PS, Hoke JF, Westmoreland C, Hug CC
Jr, Muir KT and Szlam F: Histamine concentrations and hemodynamic
responses after remifentanil. Anesth Analg. 80:990–993. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ouattara A, Boccara G, Köckler U, Lecomte
P, Leprince P, Léger P, Riou B, Rama A and Coriat P: Remifentanil
induces systemic arterial vasodilation in humans with a total
artificial heart. Anesthesiology. 100:602–607. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kazmaier S, Hanekop GG, Buhre W, Weyland
A, Busch T, Radke OC, Zoelffel R and Sonntag H: Myocardial
consequences of remifentanil in patients with coronary artery
disease. Br J Anaesth. 84:578–583. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Noseir RK, Ficke DJ, Kundu A, Arain SR and
Ebert TJ: Sympathetic and vascular consequences from remifentanil
in humans. Anesth Analg. 96:1645–1650. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gursoy S, Bagcivan I, Yildirim MK, Berkan
O and Kaya T: Vasorelaxant effect of opioid analgesics on the
isolated human radial artery. Eur J Anaesthesiol. 23:496–500. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Unlugenc H, Itegin M, Ocal I, Ozalevli M,
Güler T and Isik G: Remifentanil produces vasorelaxation in
isolated rat thoracic aorta strips. Acta Anaesthesiol Scand.
47:65–69. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Paris A, Scholz J, von Knobelsdorff G,
Tonner PH, Schulte AM and Esch J: The effect of remifentanil on
cerebral blood flow velocity. Anesth Analg. 87:569–573. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Engelhard K, Reeker W, Kochs E and Werner
C: Effect of remifentanil on intracranial pressure and cerebral
blood flow velocity in patients with head trauma. Acta Anaesthesiol
Scand. 48:396–399. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hill MA, Yang Y, Ella SR, Davis MJ and
Braun AP: Large conductance, Ca2+-activated K+ channels
(BKCa) and arteriolar myogenic signaling. FEBS Lett. 584:2033–2042.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nelson MT and Quayle JM: Physiological
roles and properties of potassium channels in arterial smooth
muscle. Am J Physiol. 268:C799–C822. 1995.PubMed/NCBI
|
|
14
|
Laboratory animal welfare: Public Health
Service policy on humane care and use of laboratory animals by
awardee institutions; notice. Federal register. 50:19584–19585.
1985.PubMed/NCBI
|
|
15
|
Zhang W, Ma KT, Wang Y, Si JQ and Li L:
Inhibitory effect of 18beta-glycyrrhetinic acid on KCl- and
PE-induced constriction of rat renal interlobar artery in vitro.
Sheng Li Xue Bao. 66:195–202. 2014.PubMed/NCBI
|
|
16
|
Tian WW, Zhao L, Ma KT, Li L and Si JQ:
Isoliquiritigenin relaxes the cerebral basilar artery by enhancing
BKCa current in spontaneously hypertensive rat: Role of sGC/cGMP.
Sheng Li Xue Bao. 67:329–334. 2015.PubMed/NCBI
|
|
17
|
Li L, Wang R, Ma KT, Li XZ, Zhang CL, Liu
WD, Zhao L and Si JQ: Differential effect of calcium-activated
potassium and chloride channels on rat basilar artery vasomotion. J
Huazhong Univ Sci Technolog Med Sci. 34:482–490. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li L, Zhang W, Shi WY, Ma KT, Zhao L, Wang
Y, Zhang L, Li XZ, Zhu H, Zhang ZS, et al: The enhancement of Cx45
expression and function in renal interlobar artery of spontaneously
hypertensive rats at different age. Kidney Blood Press Res.
40:52–65. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li XZ, Ma KT, Guan BC, Li L, Zhao L, Zhang
ZS, Si JQ and Jiang ZG: Fenamates block gap junction coupling and
potentiate BKCa channels in guinea pig arteriolar cells. Eur J
Pharmacol. 703:74–82. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lin PT, Liao DQ and Luo NF: Effect of
remifentanil on calcium- activated potassium currents in human
mesenteric arterial smooth muscle cells. Chin J Anesthesiol.
30:1307–1309. 2006.
|
|
21
|
Liu Y, Hudetz AG, Knaus HG and Rusch NJ:
Increased expression of Ca2+-sensitive K+ channels in
the cerebral microcirculation of genetically hypertensive rats:
Evidence for their protection against cerebral vasospasm. Circ Res.
82:729–737. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hu Z, Ma A, Tian HY, Xi YT, Fan LH and
Wang TZ: Activity and changed expression of calcium-activated
potassium channel in vascular smooth muscle cells isolated from
mesenteric arteries of spontaneous hypertensive rats. J Xi'an Jiao
tong University (Medical Sciences). 31:424–428. 2010.
|
|
23
|
Yang Y, Li PY, Cheng J, Mao L, Wen J, Tan
XQ, Liu ZF and Zeng XR: Function of BKCa channels is reduced in
human vascular smooth muscle cells from Han Chinese patients with
hypertension. Hypertension. 61:519–525. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu QM: Clinical Anesthesiology. People's
Medical Publishing House; Beijing: pp. 242–243. 2008, (In
Chinese).
|
|
25
|
Su X, Wachtel RE and Gebhart GF:
Inhibition of calcium currents in rat colon sensory neurons by K-
but not mu- or delta-opioids. J Neurophysiol. 80:3112–3119.
1998.PubMed/NCBI
|
|
26
|
Twitchell WA and Rane SG: Opioid peptide
modulation of Ca(2+)-dependent K+ and voltage-activated
Ca2+ currents in bovine adrenal chromaffin cells.
Neuron. 10:701–709. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Leff P: The two-state model of receptor
activation. Trends Pharmacol Sci. 16:89–97. 1995. View Article : Google Scholar : PubMed/NCBI
|