Introduction
Alzheimer's disease (AD) is the most common form of
dementia that affected an estimated 44 million individuals in 2015.
This figure is expected to increase to 135 million by the year 2050
throughout the world (1). The main
clinical symptoms of AD include memory impairment, decreased
self-care ability, aphasia and visuospatial dysgnosia (2). Neuropathological studies have
confirmed that extracellular senile plaque deposition,
intracellular accumulation of neurofibrillary tangles and neuronal
degeneration loss are the main pathological hallmarks of AD
(3,4). Deposition of amyloid β-protein (Aβ)
in the brain leads to the formation of senile plaques, which
represent a primary pathological characteristic of AD (5). Aβ refers to a class of amyloid
precursor protein (APP)-derived polypeptides that are formed under
the action of β- and γ-secretase. The common subtypes of Aβ are
Aβ1–40 and Aβ1–42, and each exert neurotoxic
effects and induce apoptosis. Therefore, reducing Aβ-induced neuron
injury and intervening in AD progression are critical for the
treatment of AD.
Studies have shown that energy dysmetabolism
contributes significantly to the pathogenesis of a variety of
ageing-associated diseases and degenerative diseases of the nervous
system, including AD (6).
Homeostatic imbalance of energy metabolism develops gradually in
the brain with age, which promotes Aβ production. Aβ further
induces energy dysmetabolism in the brain, resulting in impaired
neuronal synaptic function (7,8).
Nicotinamide adenine dinucleotide (NAD+) is an important
coenzyme in the in vivo redox reaction. It participates in
energy synthesis through glycolysis, the tricarboxylic acid cycle
and mitochondrial oxidative phosphorylation (9). NAD+ may be synthesized
de novo, although the majority of NAD is synthesized via the
salvage pathway from the conversion of nicotinamide into
nicotinamide mononucleotide, which is later converted to
NAD+ (10). The
NAD+/nicotinamide adenine dinucleotide hydride (NADH)
ratio is a fundamental indicator of the cellular redox status
(11). Nicotinamide
phosphoribosyltransferase (NAMPT) is the rate-limiting enzyme in
the NAD+ salvage pathway, and it has been demonstrated
that NAMPT participates in energy metabolism and maintains the
homeostasis of energy metabolism in the brain by regulating
NAD+ synthesis; thus, increased expression levels of
NAMPT have been shown to positively regulate NAD+ levels
(12). Knockout of the NAMPT gene
enhances the formation of Aβ plaques in the brain (13). Therefore, recent studies have
focused on regulation of NAMPT expression for the purpose of
protecting against AD and other neurodegenerative diseases
(13–15).
Rhodiola rosea is a perennial herb that
belongs to the family Crassulaceae and genus Rhodiola.
Rhodiola is a traditional herbal medicine commonly used
across Asia and grows at altitudes of 1,800–2,500 m in cold,
pollution-free zones. Rhodiola stimulates the nervous
system, improves work efficiency, alleviates fatigue and prevents
mountain sickness. Salidroside is the primary active ingredient of
Rhodiola. Various studies have shown that salidroside
inhibits the inflammatory response (16), reduces oxidative stress-induced
damage (17) and exerts an
anticancer effect (18). Certain
studies demonstrated that salidroside has a neuroprotective effect,
for example, salidroside inhibited Aβ-induced neurotoxicity by
regulating the phosphatidylinositol 3-kinase/Akt signaling pathway
in primary cultured neurons (19).
In a rat model of AD, salidroside improved memory impairment of
rats (20), and salidroside
reduced mitochondrial swelling and attenuated the injury in the
central nervous system (21).
However, whether the neuroprotective effects of salidroside involve
regulation of the NAMPT/NAD+ signaling pathway in PC12
cells remains unknown.
To gain further insights into the neuroprotective
functions of salidroside, a PC12 model was used in the present
study and the underlying mechanisms were investigated. Whether the
neuroprotective effects of salidroside increasing NAD levels
against Aβ1–40-induced neurotoxicity in PC12 cells were
elucidated in the current study. Furthermore, the possible
underlying mechanisms were examined by investigating NAMPT
expression levels.
Materials and methods
Preparation of Aβ1–40
The lyophilized powder of Aβ1–40 (cat no. A1075;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was diluted with
sterile distilled water to a concentration of 200 µmol/l.
Subsequently, Aβ1–40 was incubated at 37°C for 7 days and stored at
4°C for further assays.
Cell culture and treatment
Highly differentiated PC12 rat adrenal
pheochromocytoma cells (Type Culture Collection of the Chinese
Academy of Sciences, Shanghai, China) were maintained in Dulbecco's
modified Eagle's medium (DMEM; cat no. 12100046) containing Gibco
10% fetal bovine serum (cat no. 10099-141) (both from Thermo Fisher
Scientific, Inc., Waltham, MA, USA) and 1% penicillin-streptomycin.
The cells were maintained in a 37°C incubator in an atmosphere of
5% CO2 and grew adherently. The culture medium was
replaced daily. Upon reaching 80–90% confluence, the cells were
trypsinized for 1–2 min. Once the cells became round, 2 ml of
culture medium was added to the cells. The cells were then
centrifuged, 1,500 × g for 5 min at room temperature, resuspended,
and split into culture dishes. The cells were divided into the
following main groups: Normal control group, the Aβ1–40 group, the
Aβ1–40 + salidroside (cat no. B20504; Shanghai Yuanye Biological
Technology Co., Ltd., Shanghai, China) group, and the salidroside
group. The normal control group was cultured conventionally in DMEM
and did not receive any treatment. The Aβ1–40 group was subjected
to 24 h of treatment with 5 µmol/l Aβ1-40. The Aβ1–40 + salidroside
group was subjected to simultaneous Aβ1–40 (5 µmol/l) and
salidroside (100 µmol/l) treatments for 24 h. The salidroside group
was treated with 100 µmol/l salidroside for 24 h.
Cell viability assay
PC12 cells were seeded into 96-well plates and
cultured at 37°C in DMEM for 24 h. Once the cells attached stably
to the culture surface, various concentrations of Aβ1–40 (1, 5 or
10 µmol/l) or salidroside (12.5, 25, 50, 100 or 200 µmol/l) were
added to the cells. Following a 24-h treatment, each well of cells
was overlaid with 10 µl MTT solution (5 mg/ml; cat no. C0009)
obtained from Beyotime Institute of Biotechnology (Haimen, China)
and incubated at 37°C for 4 h. The supernatant was discarded, and
150 µl dimethyl sulphoxide was added to each well of cells to fully
dissolve the crystals. Subsequently, absorbance was measured at a
wavelength of 490 nm using a PowerWave XS microplate reader (BioTek
Instruments, Inc., Winooski, VT, USA).
Lactate dehydrogenase (LDH) activity
assay
The cells were subjected to different treatments
(medium only, 5 µmol/l Aβ1-40, 5 µmol/l Aβ1–40 + 100 µmol/l
salidroside or 100 µmol/l salidroside). Subsequently, the activity
of released LDH was examined according to the instructions provided
with the LDH Activity Assay kit (cat. no. G1780; Promega
Corporation, Madison, WI, USA). Absorbance was measured at a
wavelength of 490 nm. The LDH release was expressed as a percentage
relative to the positive control group.
NAD+/NADH analysis
An NAD+/NADH quantification kit was used
to determine NAD+ and NADH levels (cat no. k337-100;
BioVision, Inc., Milpitas, CA, USA) according to the manufacturer's
instructions. The cells were washed with pre-cooled
phosphate-buffered saline (PBS), homogenized in 400 µl
NAD+/NADH extraction buffer, and then centrifuged for 5
min at 18,000 × g and 4°C. The resulting supernatants were
transferred to fresh Eppendorf (EP) tubes and labelled as NADt
samples. Subsequently, 200 µl NADt sample was transferred to fresh
EP tubes and heated at 60°C for 30 min (all NAD+
decomposed, leaving only NADH to be analyzed). After cooling on
ice, the samples were quickly centrifuged 12,000 × g for 30 sec at
4°C, and the resulting pellets were discarded. The supernatants
were collected and labelled as NADH samples for further assays.
Standards (0, 2, 4, 6, 8 and 10 µl) and 40 µl samples were loaded
into the wells of a microtiter plate, and appropriate volumes of
NAD+/NADH Extraction Buffer were added to bring the
volume to 50 µl. Subsequently, 100 µl enzyme reaction mix and 10 µl
NADH developer were added to each well. The mixtures were allowed
to react at room temperature (RT) for 1–4 h, and the absorbance was
measured using a microplate reader (wavelength, 450 nm).
Immunofluorescence assay
Following various treatments and interventions, all
groups of cells were washed with PBS for 5 min. The cells were then
fixed with 4% paraformaldehyde for 15 min, blocked with a blocking
solution containing 10% calf serum for 45 min at RT, incubated with
anti-NAMPT antibody (dilution, 1:200; cat no. ab45890; Abcam,
Cambridge, MA, USA) at 37°C for 1 h, and placed in a 4°C
refrigerator overnight. After washing with PBS, the cells were
incubated with fluorescein isothiocyanate-labelled goat anti-rabbit
IgG (dilution, 1:500; cat no. A0562; Beyotime Institute of
Biotechnology) for 1 h in a 37°C incubator. After 3 rinses with
PBS, the cells were covered with DAPI staining solution (cat no.
C1005; Beyotime Institute of Biotechnology) and incubated at RT for
5 min. The cells were rinsed 3 more times with PBS (5 min each
time). Subsequently, the cells were mounted onto glass slides in
PBS and glycerol (1:1). The fluorescence signal was examined using
an Olympus BX51 biological microscope at a magnification of
×200.
Western blot analysis
At the end of the treatment period, all groups of
cells were collected. Following centrifugation 1,500 × g for 5 min
at RT, the cells were lysed in lysis buffer. The resulting
supernatants were extracted and the protein concentrations in the
supernatants were determined using the bicinchoninic acid (BCA)
method. Subsequently, the supernatants were mixed thoroughly with
5X sodium dodecyl sulphate (SDS) loading buffer, boiled at 100°C
for 5 min, cooled on ice and then stored at −20°C. Immediately
before the experiments, the protein samples were thawed at RT. The
protein samples (50 µg each) were heated at 95°C for 5 min, then
separated by SDS-polyacrylamide gel electrophoresis and transferred
to polyvinylidene fluoride membranes. The membranes were blocked
with 2% bovine serum albumin for 1 h and then incubated with the
primary antibody (anti-NAMPT antibody, dilution, 1:250; cat no.
ab45890; Abcam) at 4°C overnight. Subsequently, the membranes were
washed with PBS for 45 min, incubated with horseradish
peroxidase-labelled secondary antibody (dilution, 1:2,000; cat no.
SA00001-2; ProteinTech Group, Inc., Chicago IL, USA) at 37°C for 1
h, and washed again in PBS for 45 min. Protein bands were examined
using an ImageQuant LAS 4000 mini chemiluminescence detector (GE
Healthcare Life Sciences, Little Chalfont, UK). GAPDH (dilution,
1:1,000, cat no. 5174S; Cell Signaling Technology, Inc., Danvers,
MA, USA) served as a loading control.
Statistical analysis
The results were statistically analyzed using
GraphPad Prism 5.0 software (GraphPad Software, Inc., La Jolla, CA,
USA). All data are expressed as the mean ± standard error. One-way
analysis of variance was used to assess multiple groups of data and
Student's t-test was used to compare between two groups and
P<0.05 was considered to indicate a statistically significant
difference.
Results
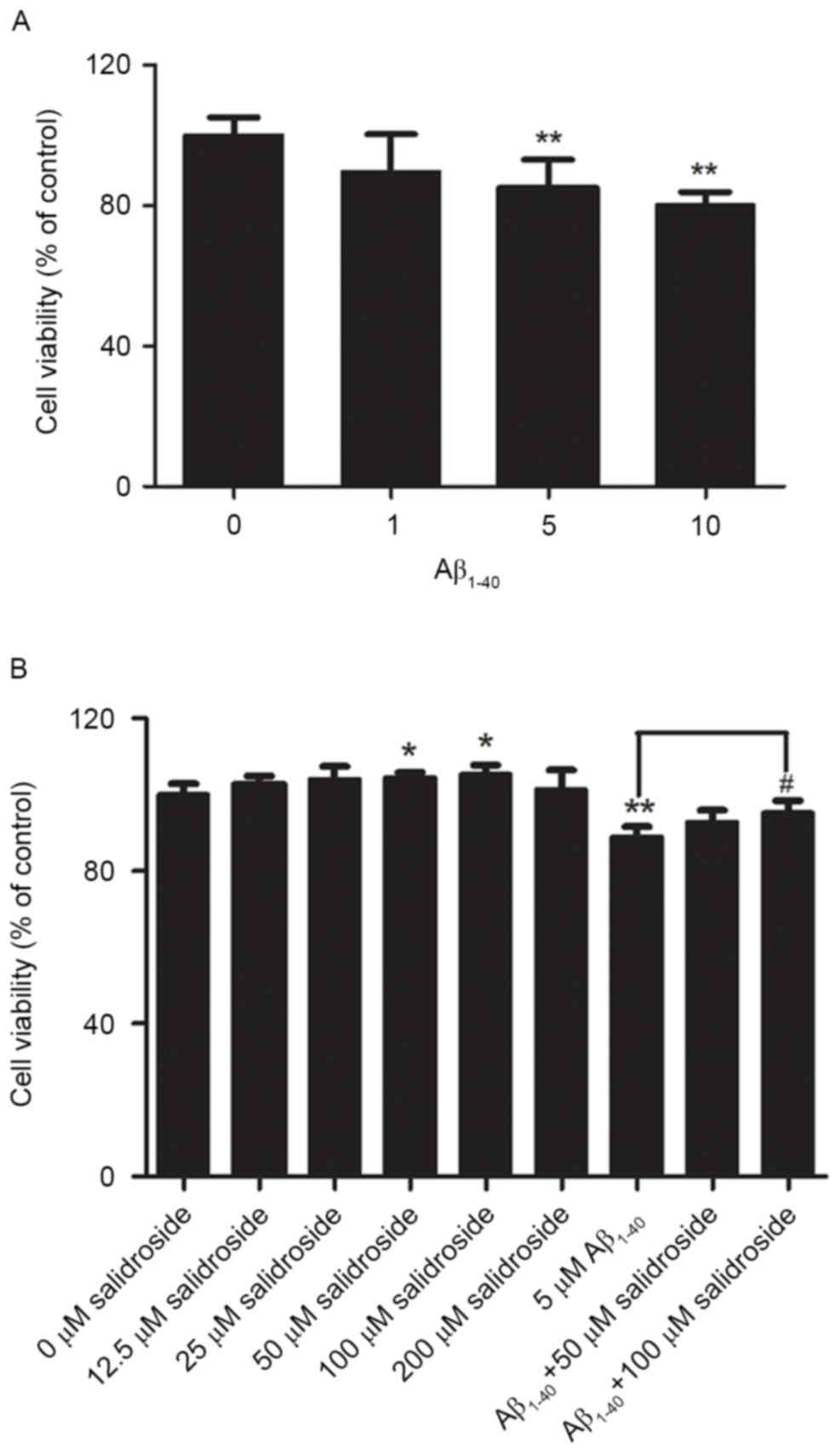
Impact of Aβ1–40 treatment
on the viability of PC12 cells and the protective effect of
salidroside
PC12 cells were treated with various concentrations
of Aβ1–40 (1, 5 or 10 µmol/l) for 24 h, which enabled the analysis
of changes in PC12 cell viability under the action of different
concentrations of Aβ1-40. The results demonstrated that cell
viability declined as the Aβ1–40 concentration increased. The
viability of PC12 cells decreased by ~15% after treatment with 5
µmol/l Aβ1-40. Therefore, a concentration of 5 µmol/l Aβ1–40 was
selected to establish the cell model of AD (Fig. 1A).
Before investigation of the protective effect of
salidroside, the present study first examined whether salidroside
would exert a neurotoxic effect on normal PC12 cells at certain
concentrations. PC12 cells were treated with various concentrations
of salidroside (12.5, 25, 50, 100 or 200 µmol/l). Compared with the
normal control group, 24-h incubation with 12.5, 25 or 200 µmol/l
salidroside exerted no significant effect on normal PC12 cells,
whereas incubation with 50 and 100 µmol/l salidroside improved cell
viability (P<0.05). Therefore, the two concentrations of
salidroside (50 and 100 µmol/l) were selected for further
investigation of the effect of salidroside on the viability of PC12
cells that were damaged by Aβ1–40 exposure. The results
indicated that cell viability was significantly decreased in the
Aβ1–40 group compared with the normal control group
(P<0.01). Compared with the Aβ1–40 group, treatment
with 50 µmol/l salidroside failed to significantly improve the
viability of Aβ1–40-damaged cells, whereas treatment
with 100 µmol/l salidroside improved the viability of cells damaged
by Aβ1–40 (P<0.05). The results demonstrated that 100
µmol/l salidroside exerted an effect on cell viability and
alleviated the Aβ-induced PC12 cell injury. Therefore, this
salidroside concentration (100 µmol/l) was selected for further
experiments (Fig. 1B).
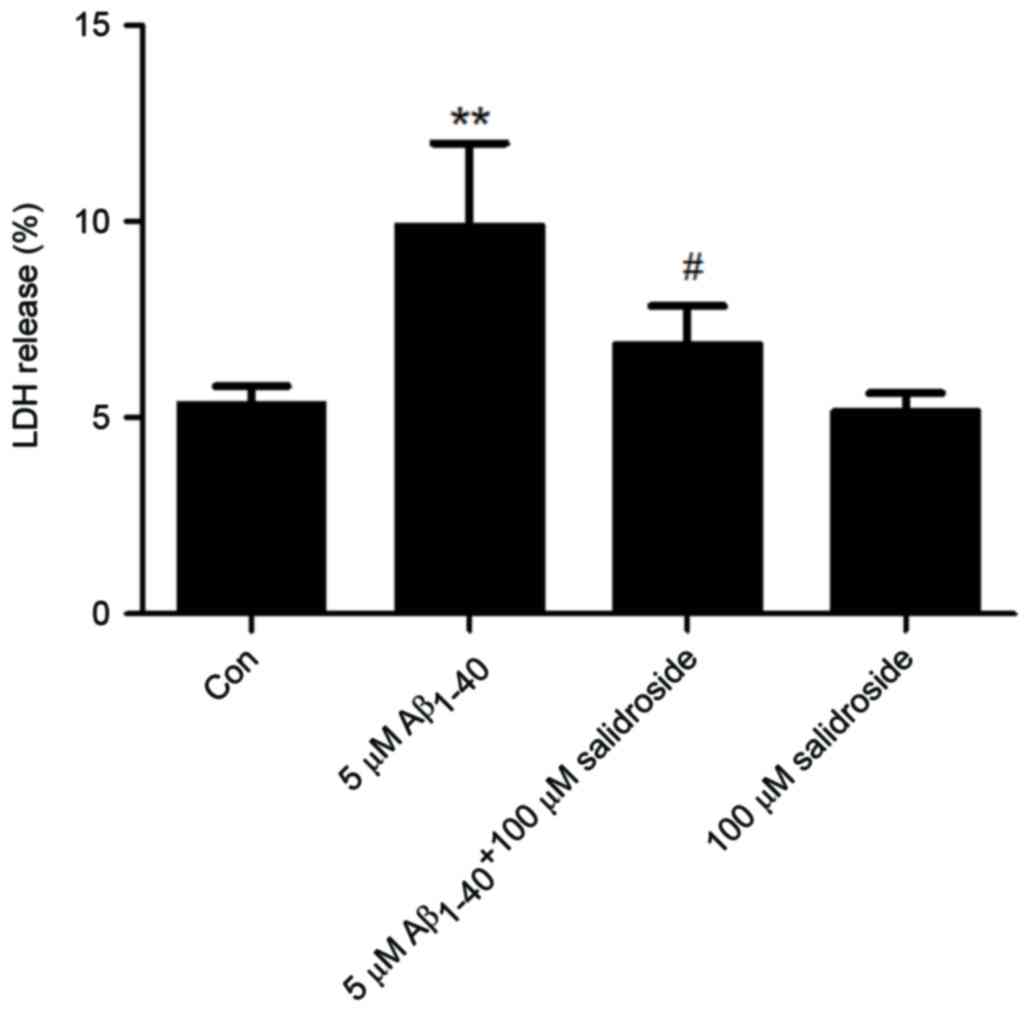
Salidroside reduces the LDH level of
Aβ1–40-treated cells
The LDH level of a culture supernatant indirectly
reflects the effects of Aβ1–40 and salidroside on cells. As shown
in Fig. 2, the Aβ1–40 group
displayed a significantly increased LDH level compared with that of
the normal control group (P<0.01). By contrast, the LDH level
was markedly reduced in the Aβ1–40 + salidroside group compared
with the Aβ1–40 group (P<0.05). The results demonstrated that
salidroside reduced the Aβ1-40-induced cellular damage.
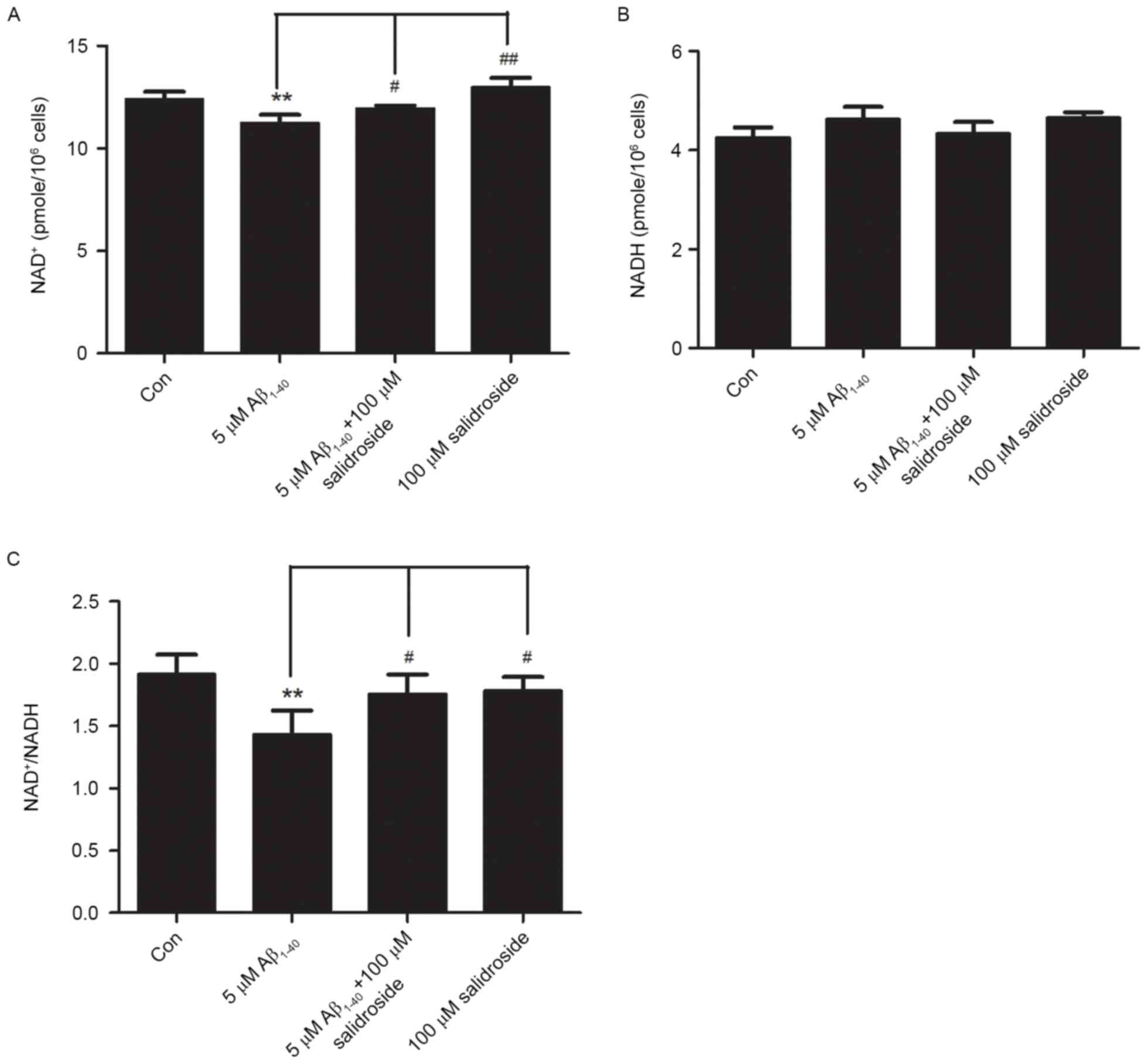
Salidroside increases the
NAD+ level and NAD+/NADH ratio of
Aβ1–40-treated cells
NAD+ and NADH are coenzymes that are
necessary for cellular energy metabolism. Under normal
physiological conditions, the NAD+/NADH ratio reflects
the cellular redox status and is in a constant state of dynamic
equilibrium. Disorders of cellular energy metabolism are
characterized by changes in NAD+ and NADH levels, and an
imbalance in the NAD+/NADH ratio (22). In the present study, the Aβ1–40
group exhibited a decreased level of NAD+ compared with
that of the normal control group (P<0.01; Fig. 3A). However, the NADH level did not
change significantly in the Aβ1–40 group compared with the normal
control group (Fig. 3B). As a
result, the NAD+/NADH ratio was reduced in the Aβ1–40
group (P<0.01; Fig. 3C). No
significant difference was identified in the NADH level between the
Aβ1–40 + salidroside group and the Aβ1–40 group (Fig. 3B). By contrast, the Aβ1–40 +
salidroside group demonstrated a significant increase in the
NAD+ level (P<0.05; Fig.
3A) and the NAD+/NADH ratio (P<0.05; Fig. 3C).
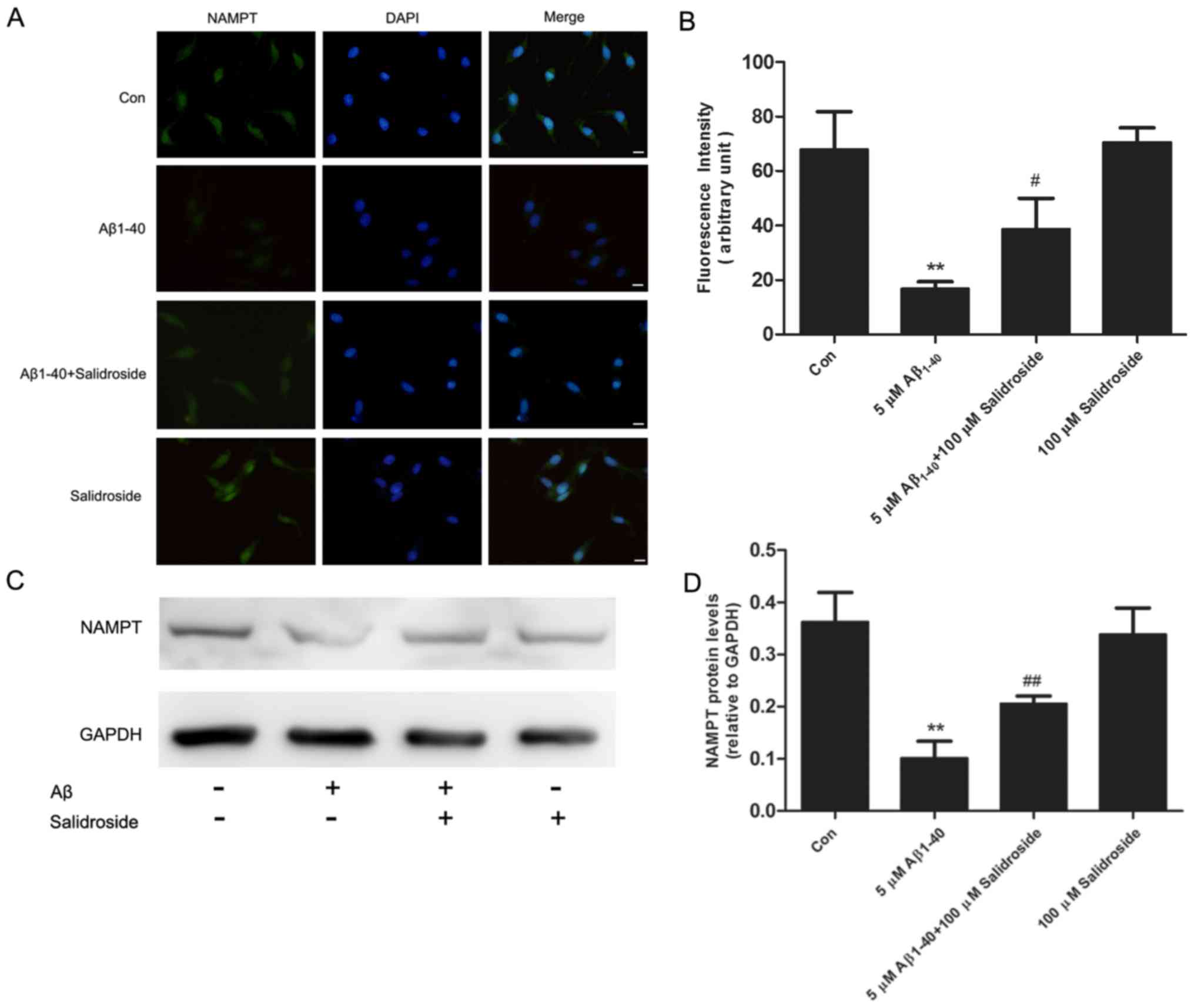
Salidroside upregulates the expression
level of NAMPT
NAMPT is the rate-limiting enzyme in the salvage
pathway of NAD+ biosynthesis and is important in
regulating the homeostasis of energy metabolism. The results of an
immunofluorescence assay demonstrated that the NAMPT-derived
fluorescence signal was reduced following 24 h of Aβ1–40 treatment,
whereas salidroside treatment increased the NAMPT fluorescence
signal (Fig. 4A and B). In
addition, western blot analysis demonstrated that NAMPT expression
was decreased in the Aβ1–40 group compared with the normal control
group. Salidroside treatment significantly increased the expression
level of NAMPT (Fig. 4C and D).
Thus, the results indicate that salidroside regulates NAMPT protein
expression.
Discussion
AD is the most common cause of dementia in the
elderly, but the exact pathogenetic mechanisms underlying AD remain
uncertain. In previous studies, one extensively investigated
mechanism is Aβ-mediated neurotoxicity; the aggregates of Aβ
peptides exhibit cytotoxicity in vivo and in vitro
(23). Aβ1–40 and
Aβ1–42 are two major forms of Aβ, although
Aβ1–42 is markedly more prone to aggregation and more
toxic to neurons than Aβ1–40; however, Aβ1–40
was the predominant isoform of the most abundant cleaved form of
APP (~90%) and exerts a toxic effect on neurons in the AD brain
(24). In addition, the deposition
of Aβ1–40 is required for the development of mature
amyloid plaques from the initial deposition of Aβ1–42,
which was regarded as an early pathological process of AD, and
Aβ1–40 is often used to establish the model of AD in
vivo and in vitro (25). Injecting 5 mg/ml Aβ1–40
caused hippocampal neuronal loss or functional impairment and is
thus considered as an indication of energy metabolism (26). As Aβ1–40 possesses
neuronal toxicity and is capable of inducing neuronal
neurotoxicity, PC12 cells treated with Aβ1–40 were used
as the cell model of AD in the current study. The neuronal toxicity
of Aβ1–40 is reflected in its ability to cause energy
dysmetabolism, and induce cell damage and death (27,28).
A number of previous studies used PC12 cells to investigate the
molecular mechanisms underlying the development of degenerative
diseases of the nervous system (29–32).
Therefore, the present study employed the PC12 cells that were
damaged by Aβ1–40 exposure as the cell model of AD and
the significance of NAMPT in AD was evaluated.
The present study demonstrated that
Aβ1–40 treatment reduced the NAD+ expression
level. The underlying reason may be associated with decreased NAMPT
expression. As the rate-limiting enzyme in the NAD+
salvage pathway, NAMPT promotes the production of NAD+
(33). Previous studies
demonstrated that an elevated NAMPT level reduces cell death
(34,35), which is consistent with the results
of the present study, which indicate that salidroside increased
NAMPT expression and improved cell survival. In addition, the
present study identified that salidroside significantly reduced the
level of LDH released when compared with that in the
Aβ1–40 group. The results of the LDH assay were
consistent with those of the MTT assay, demonstrating that
salidroside treatment attenuated the Aβ1–40-induced PC12
cell injury and exerted a protective effect on PC12 cells.
The expression level of Aβ increases as AD
progresses, which causes mitochondrial dysfunction and energy
metabolism disorders. The elevated Aβ expression level represents
one of the reasons behind neuronal damage and apoptosis (36). NAD+ participates in
energy metabolism. Furthermore, loss of NAD+ homeostasis
leads to decreased sirtuin-1 (SIRT1) activity. Consequently,
SIRT1-mediated deacetylation of signaling molecules (including
transcription factors and enzymes) is reduced (37,38).
Peroxisome proliferator-activated receptor γ coactivator 1-α
(PGC-1α) is a coactivator of the nuclear hormone receptor
peroxisome proliferator-activated receptor γ, which also undergoes
SIRT1-mediated deacetylation and participates in mitochondrial
biosynthesis and energy homeostasis (39). The decreased SIRT1 activity in the
brain affects the function of the downstream protein PGC-1α in
energy synthesis, resulting in a brain energy crisis (40). As NAMPT is important in
NAD+ synthesis, the present study investigated whether
salidroside exerts its regulatory effect on energy metabolism
pathways via NAMPT. Namely, salidroside attenuates the neuronal
toxicity of Aβ by further regulating NAD+/NADH levels.
Studies have demonstrated that the expression level of NAMPT
decreases gradually with age, with the decrease in NAMPT level
particularly evident in the brain of AD mice compared with aged
healthy mice. Such a decrease exacerbates neuronal apoptosis
(34,41,42).
By contrast, increased NAMPT expression levels in the hippocampus
and cortex improve cognitive function (35). Therefore, novel treatment
strategies for AD may involve the administration of salidroside to
regulate NAMPT expression and reduce the severity of AD.
The present study demonstrated that salidroside
exerted a protective effect on PC12 cells damaged by
Aβ1–40 exposure. An AD model was established by
Aβ1–40 treatment. The viability of PC12 cells decreased
and the level of LDH release significantly increased. In addition,
the protein expression of NAMPT, the level of NAD+ and
the ratio of NAD+/NADH were decreased. The
above-mentioned effects of Aβ1–40 were attenuated
following salidroside treatment. In addition, the present study
demonstrated that the decrease in NAMPT/NAD+ levels may
represent one of the molecular pathological mechanisms of AD.
In conclusion, the present study demonstrated that
salidroside exerted a protective effect on PC12 damaged by
Aβ1–40 treatment. The protective effect of salidroside
may reside in its regulatory effect in energy metabolism, such as
the increases in NAMPT expression levels and NAD+
production. Therefore, NAMPT and NAD+ are considered to
be key factors in AD, as well as other neurodegenerative diseases.
Thus, regulation of NAMPT and NAD+ may serve as novel
therapeutic strategies for the treatment of AD.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 81503626)
and the Shanghai Health Bureau Youth Fund (grant nos. 201540254 and
20134331).
References
|
1
|
Van Cauwenberghe C, Van Broeckhoven C and
Sleegers K: The genetic landscape of Alzheimer disease: Clinical
implications and perspectives. Genet Med. 18:421–430. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Scheltens P, Blennow K, Breteler MM, de
Strooper B, Frisoni GB, Salloway S and Van der Flier WM:
Alzheimer's disease. Lancet. 388:505–517. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Perl DP: Neuropathology of Alzheimer's
disease. Mt Sinai J Med. 77:32–42. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Serrano-Pozo A, Frosch MP, Masliah E and
Hyman BT: Neuropathological alterations in Alzheimer disease. Cold
Spring Harb Perspect Med. 1:a0061892011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sadigh-Eteghad S, Sabermarouf B, Majdi A,
Talebi M, Farhoudi M and Mahmoudi J: Amyloid-beta: A crucial factor
in Alzheimer's disease. Med Princ Pract. 24:1–10. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yin F, Boveris A and Cadenas E:
Mitochondrial energy metabolism and redox signaling in brain aging
and neurodegeneration. Antioxid Redox Signal. 20:353–371. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang R, Li JJ, Diao S, Kwak YD, Liu L, Zhi
L, Büeler H, Bhat NR, Williams RW, Park EA and Liao FF: Metabolic
stress modulates Alzheimer's β-secretase gene transcription via
SIRT1-PPARγ-PGC-1 in neurons. Cell Metab. 17:685–694. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu MF, Yin JH, Hwang CS, Tang CM and Yang
DI: NAD attenuates oxidative DNA damages induced by amyloid
beta-peptide in primary rat cortical neurons. Free Radic Res.
48:794–805. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ussher JR, Jaswal JS and Lopaschuk GD:
Pyridine nucleotide regulation of cardiac intermediary metabolism.
Circ Res. 111:628–641. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rongvaux A, Shea RJ, Mulks MH, Gigot D,
Urbain J, Leo O and Andris F: Pre-B-cell colony-enhancing factor,
whose expression is up-regulated in activated lymphocytes, is a
nicotinamide phosphoribosyltransferase, a cytosolic enzyme involved
in NAD biosynthesis. Eur J Immunol. 32:3225–3234. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ying W: NAD+/NADH and
NADP+/NADPH in cellular functions and cell death:
Regulation and biological consequences. Antioxid Redox Signal.
10:179–206. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Revollo JR, Grimm AA and Imai S: The
regulation of nicotinamide adenine dinucleotide biosynthesis by
Nampt/PBEF/visfatin in mammals. Curr Opin Gastroenterol.
23:164–170. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Villela D, Schlesinger D, Suemoto CK,
Grinberg LT and Rosenberg C: A microdeletion in Alzheimer's disease
disrupts NAMPT gene. J Genet. 93:535–537. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Imai S and Guarente L: NAD+ and
sirtuins in aging and disease. Trends Cell Biol. 24:464–471. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Herrera-Arozamena C, Martí-Marí O, Estrada
M, de la Fuente Revenga M and Rodríguez-Franco MI: Recent advances
in neurogenic small molecules as innovative treatments for
neurodegenerative diseases. Molecules. 21:pii: E1165. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qi Z, Qi S, Ling L, Lv J and Feng Z:
Salidroside attenuates inflammatory response via suppressing
JAK2-STAT3 pathway activation and preventing STAT3 transfer into
nucleus. Int Immunopharmacol. 35:265–271. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang ZR, Wang HF, Zuo TC, Guan LL and Dai
N: Salidroside alleviates oxidative stress in the liver with non-
alcoholic steatohepatitis in rats. BMC Pharmacol Toxicol.
17:162016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sun KX, Xia HW and Xia RL: Anticancer
effect of salidroside on colon cancer through inhibiting JAK2/STAT3
signaling pathway. Int J Clin Exp Pathol. 8:615–621.
2015.PubMed/NCBI
|
|
19
|
Zhang B, Wang Y, Li H, Xiong R, Zhao Z,
Chu X, Li Q, Sun S and Chen S: Neuroprotective effects of
salidroside through PI3K/Akt pathway activation in Alzheimer's
disease models. Drug Des Devel Ther. 10:1335–1343. 2016.PubMed/NCBI
|
|
20
|
Gao J, Zhou R, You X, Luo F, He H, Chang
X, Zhu L, Ding X and Yan T: Salidroside suppresses inflammation in
a D-galactose-induced rat model of Alzheimer's disease via
SIRT1/NF-κB pathway. Metab Brain Dis. 31:771–778. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang W, Peng M, Yang Y, Xiao Z, Song B
and Lin Z: Protective effects of salidroside on mitochondrial
functions against exertional heat stroke-induced organ damage in
the rat. Evid Based Complement Alternat Med. 2015:5045672015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang Y and Sauve AA: NAD+
metabolism: Bioenergetics, signaling and manipulation for therapy.
Biochim Biophys Acta. 1864:1787–1800. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Selkoe DJ and Schenk D: Alzheimer's
disease: Molecular understanding predicts amyloid-based
therapeutics. Annu Rev Pharmacol Toxicol. 43:545–584. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cheng YF, Wang C, Lin HB, Li YF, Huang Y,
Xu JP and Zhang HT: Inhibition of phosphodiesterase-4 reverses
memory deficits produced by Aβ25–35 or Aβ1–40
peptide in rats. Psychopharmacology (Berl). 212:181–191. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Miguel-Hidalgo JJ and Cacabelos R:
Beta-amyloid(1–40)-induced neurodegeneration in the rat hippocampal
neurons of the CA1 subfield. Acta Neuropathol. 95:455–465. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yue T, Shanbin G, Ling M, Yuan W, Ying X
and Ping Z: Sevoflurane aggregates cognitive dysfunction and
hippocampal oxidative stress induced by β-amyloid in rats. Life
Sci. 143:194–201. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
He F, Cao YP, Che FY, Yang LH, Xiao SH and
Liu J: Inhibitory effects of edaravone in β-amyloid-induced
neurotoxicity in rats. Biomed Res Int. 2014:3703682014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Carrillo-Mora P, Luna R and Colín-Barenque
L: Amyloid beta: Multiple mechanisms of toxicity and only some
protective effects? Oxid Med Cell Longev. 2014:7953752014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Han XH, Cheng MN, Chen L, Fang H, Wang LJ,
Li XT and Qu ZQ: 7,8-Dihydroxyflavone protects PC12 cells against
6-hydroxydopamine-induced cell death through modulating PI3K/Akt
and JNK pathways. Neurosci Lett. 581:85–88. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Guo M, Li D, Shen H, Jin B, Ren Y, Li M
and Xing Y: Leptin-Sensitive JAK2 activation in the regulation of
Tau phosphorylation in PC12 cells. Neurosignals. 24:88–94. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen CL, Tsai WH, Chen CJ and Pan TM:
Centella asiatica extract protects against amyloid
β1-40-induced neurotoxicity in neuronal cells by activating the
antioxidative defence system. J Tradit Complement Med. 6:362–369.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Qian MC, Liu J, Yao JS, Wang WM, Yang JH,
Wei LL, Shen YD and Chen W: Caspase-8 mediates amyloid-β-induced
apoptosis in differentiated PC12 cells. J Mol Neurosci. 56:491–499.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jieyu H, Chao T, Mengjun L, Shalong W,
Xiaomei G, Jianfeng L and Zhihong L: Nampt/Visfatin/PBEF: A
functionally multi-faceted protein with a pivotal role in malignant
tumors. Curr Pharm Des. 18:6123–6132. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ghosh D, Levault KR and Brewer GJ:
Relative importance of redox buffers GSH and NAD(P)H in age-related
neurodegeneration and Alzheimer disease-like mouse neurons. Aging
Cell. 13:631–640. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Stein LR, Wozniak DF, Dearborn JT, Kubota
S, Apte RS, Izumi Y, Zorumski CF and Imai S: Expression of Nampt in
hippocampal and cortical excitatory neurons is critical for
cognitive function. J Neurosci. 34:5800–5815. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cabezas-Opazo FA, Vergara-Pulgar K, Pérez
MJ, Jara C, Osorio-Fuentealba C and Quintanilla RA: Mitochondrial
dysfunction contributes to the pathogenesis of Alzheimer's disease.
Oxid Med Cell Longev. 2015:5096542015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhu XH, Lu M, Lee BY, Ugurbil K and Chen
W: In vivo NAD assay reveals the intracellular NAD contents and
redox state in healthy human brain and their age dependences. Proc
Natl Acad Sci USA. 112:2876–2881. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang Y: Molecular links between caloric
restriction and Sir2/SIRT1 activation. Diabetes Metab J.
38:321–329. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rodgers JT, Lerin C, Gerhart-Hines Z and
Puigserver P: Metabolic adaptations through the PGC-1 alpha and
SIRT1 pathways. FEBS Lett. 582:46–53. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Katsouri L, Parr C, Bogdanovic N, Willem M
and Sastre M: PPARγ co-activator-1α (PGC-1α) reduces amyloid-β
generation through a PPARγ-dependent mechanism. J Alzheimers Dis.
25:151–162. 2011.PubMed/NCBI
|
|
41
|
Yoshino J, Mills KF, Yoon MJ and Imai S:
Nicotinamide mononucleotide, a key NAD(+) intermediate, treats the
pathophysiology of diet- and age-induced diabetes in mice. Cell
Metab. 14:528–536. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ghosh D, LeVault KR, Barnett AJ and Brewer
GJ: A reversible early oxidized redox state that precedes
macromolecular ROS damage in aging nontransgenic and 3xTg-AD mouse
neurons. J Neurosci. 32:5821–5832. 2012. View Article : Google Scholar : PubMed/NCBI
|