Introduction
Ischemic osteonecrosis of the femoral head commonly
leads to the destruction of the hip joint and is a progressive,
debilitating disease (1). Multiple
factors may result in the occurrence of the disease, including
fracture and dislocation, glucocorticoid and radiation therapy,
alcoholism, connective tissue disease, or infection (2). The key elements in the pathogenesis
of ischemic necrosis of the femoral head are vascular disruption
and defective bone remodeling. Vascular disruption normally induces
cell ischemia by hypoxia. The hypoxic condition normally induces
apoptosis in osteoblasts, vascular endothelial cells and smooth
muscle cells by triggering mitochondrial permeability. A previous
study demonstrated that hypoxia activated a mitochondrial-dependent
classic apoptosis pathway, which increased cytochrome c
release and the subsequent cleaving and activation of caspase 9 and
other effector caspases (3).
Sirtuins (SIRTs), which were first identified in
yeast, are nicotinamide adenosine dinucleotide (NAD)-dependent
deacetylases. Currently, seven SIRTs have been identified and are
expressed in mammalian cells. SIRT1, SIRT6 and SIRT7 are localized
in the nucleus, SIRT2 in the cytoplasm and SIRT3, SIRT4 and SIRT5
are expressed in the mitochondria (4,5).
SIRT1 affects a series of biological functions, including DNA
repair, tumor suppression, energy metabolism and mitochondrial
permeability. SIRT1 controls longevity and delays senility in lower
organisms and mammals (6).
Previous studies have demonstrated that SIRT1 has vital inhibitory
effects in chondrocytes and on cardiac myocyte apoptosis in
vascular endothelial cells and mesenchymal stem cells (7–9).
These effects are primarily mediated by deacetylating substrates,
including important proteins, transcription factors or co-factors,
including p53, protein kinase B (Akt), the Forkhead box O (FoxO)
family and β-catenin, a co-activator of canonical Wnt signaling.
The deacetylation of these proteins by SIRT1 inhibits the apoptosis
process in mammalian cells (10–12).
Therefore, it is possible that SIRT1 serves an important role in
cell defence and survival through interaction with
apoptosis-inducible nuclear proteins. Due to the protective effects
of SIRT1 in cells, SIRT1 is additionally a key regulator of aging
and apoptosis in osteoblasts. Osteoblasts, which are the primary
functional cells for bone formation, are responsible for the
synthesis, secretion and mineralization of bone matrix. Any factor
that suppresses the generation and differentiation or promotes the
apoptosis of osteoblasts may reduce bone formation and result in an
imbalance in bone remodeling, leading to bone loss. Thus
hypoxia-induced apoptosis and necrosis interfere with the bone
remodelling process. To the best of our knowledge, no studies have
reported the direct role of SIRT1 in hypoxia-induced apoptosis of
osteoblasts.
The present study investigated the protective effect
of SIRT1 in the survival of MC3T3-E1 osteoblast cells against a
hypoxic stimulus, and attempted to identify the associated
underlying mechanism. The results of the present study demonstrated
that SIRT1 deacetylases the substrates of phosphoinositide 3-kinase
(PI3K)/Akt and nuclear factor (NF)-κB in downstream anti-apoptosis
pathways and serves an important role in mediating the
anti-apoptotic effects in MC3T3-E1 cells.
Materials and methods
Cell culture and transfection
MC3T3-E1 cells were purchased from American Type
Culture Collection (Manassas, VA, USA) and were plated in 6-well
plates at a density of 1.5×106 cells/well in Minimum
Essential α-Medium (αMEM; Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS),
10 units/ml penicillin and 10 µg/ml streptomycin (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany). For the SIRT1 silencing
experiment, transfection of MC3T3-E1 cells was performed by using
small interference (si)RNA. The siRNA sequences that were used were
as follows: si-SIRT1 sense, 5′-GAGACUGCGAUGUCAUAAUTT-3′ and
antisense, 5′-AUUAUGACAUCGCAGUCUCTT-3′; and scrambled siRNA sense,
5′-CGCUCCGAACGUGCUACGUTT-3′ and antisense,
5′-ACGGUACACGUUCAAAGAATT-3′. siRNAs were purchased from GE
Dharmacon (Lafayette, CO, USA). Transient transfection was
performed using Lipofectamine 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) at 37°C for 6 h using a final concentration of
siRNAs of 5 nM, according to the manufacturer's protocol). For the
SIRT1 overexpression experiment, MC3T3-E1 cells were transfected
for 8 h at 37°C with 1×109 particle forming units of
SIRT1 adenovirus, which was obtained from Professor B.H. Park
(Department of Biochemistry, Chonbuk National University, Jeonju,
Republic of Korea). Viruses were purified from the supernatants of
293 cell cultures by cesium chloride density gradient
centrifugation at 70,000 × g at 4°C for 3 h. As a control, the
pAd/CMV/V5-GW/lacZ vector (Invitrogen; Thermo Fisher Scientific,
Inc.) was used to produce LacZ-bearing adenovirus. MC3T3-E1 cells
were incubated in α-MEM medium without antibiotics for 24 h prior
to transfection to enhance the transfection efficiency.
Establishment of hypoxic culture
conditions
MC3T3-E1 cells were incubated at 37°C in anaerobic
jars (Oxoid; Thermo Fisher Scientific, Inc.) with oxygen-absorbing
packs (AnaeroGen; Oxoid; Thermo Fisher Scientific, Inc.). Prior to
transferring to the hypoxic chamber, the cells were moved to a
glucose-free medium. Within 0.5 h, O2 had decreased to less than
0.1%.
Analysis of cell viability
An MTT assay was used to estimate cell viability
(12,13). Briefly, cells were plated at a
density of 1×104 cells/well in α-MEM medium supplemented
with 10% FBS and antibiotics in 96-well plates. Following exposure
to hypoxia for 24 h, MTT solution (Sigma-Aldrich; Merck KGaA) was
added in plates at a final concentration of 0.5 mg/ml for 3 h at
37°C. Following removal of the medium, 100 µl dimethyl sulfoxide
solution was added to dissolve the formazan crystals. The
absorbance at 570 nm was detected with a microplate reader (Synergy
4 Hybrid Multi-Mode; BioTek Instruments, Inc., Winooski, VT,
USA).
Detection of caspase 3 and 7
activity
Caspase 3 and 7 activity was detected using a
luminescence-based Caspase-Glo® 3/7 Assay (Promega
Corporation, Madison, WI, USA) according to the manufacturer's
protocol. Cells were cultured in hypoxia for 24 h and Caspase-Glo
3/7 reagent was added to each well in a 1:1 ratio according to the
manufacturer's protocol. Following agitation for 10 min on a plate
shaker at room temperature, 90% of the lysate volume was
transferred to a 96-well solid-white plate. Cell lysates were
analyzed with a microplate reader (Synergy 4 Hybrid Multi-Mode;
BioTek Instruments, Inc.).
Detection of apoptosis
Hypoxia-induced cell apoptosis was determined using
the Cell-APOPercentage™ Apoptosis Assay (cat. no. GB2356929;
Biocolor Ltd., Carrickfergus, Northern Ireland). Digital images of
APOPercentage dye-labeled cells, appearing bright pink against a
white background under a light microscope, were used to detect
apoptotic cells. To quantify the level of apoptosis, cells were
incubated with a 1:100 dilution of the dye for 5 min at 37°C
following culture under each experimental condition. Cells were
then washed with PBS and the dye within the labeled cells was
released into the supplied dye-release reagent. The contents of
each well (250 µl) were transferred to a 96-well flat-bottomed
plate, and absorbance was read at 550 nm with a microplate
reader.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
The relative levels of SIRT1, tumor necrosis factor
receptor-associated factor 2 (TRAF2) and cellular inhibitor of
apoptosis 2 (cIAP2) mRNA were measured using RT-qPCR, as previously
described (13). Total RNA was
extracted from cells at a density of 1.5×106 cells/well,
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). RNA was precipitated with RNase-free chloroform
and isopropanol (Sigma-Aldrich; Merck KGaA) and dissolved in
diethyl pyrocarbonate-treated distilled water (Promega
Corporation). Total RNA (1 µg) free of genomic DNA contamination
was reverse-transcribed into cDNA using the random hexamer primer
provided in the RevertAid First Strand cDNA Synthesis kit (cat. no.
K1612; Applied Biosystems; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol in a 20 µl reaction
volume. The temperature protocol was as follows: At 25°C for 5 min,
at 37°C for 60 min and at 70°C for 5 min. Specific primers for each
gene were designed using the Primer Express software version 3.0
(Applied Biosystems; Thermo Fisher Scientific, Inc.). Primer
sequences used in the present study were as follows: SIRT1 forward,
5′-AAGTTGACTGTGAAGCTGTACG-3′ and reverse,
5′-TGCTACTGGTCTTACTTTGAGGG-3′; TRAF2 forward,
5′-TCCCTGGAGTTGCTACAGC-3′ and reverse, 5′-AGGCGGAGCACAGGTACTT-3′;
cIAP2 forward, 5′-TTTCCGTGGCTCTTATTCAAACT-3′ and reverse,
5′-GCACAGTGGTAGGAACTTCTCAT-3′; and GAPDH forward,
5′-TGTGGGCATCAATGGATTTGG-3′ and reverse,
5′-ACACCATGTATTCCGGGTCAAT-3′. qPCR was performed using the ABI
Prism 7900HT Sequence Detection System (Applied Biosystems; Thermo
Fisher Scientific, Inc.). PCR reactions were carried out in a final
volume of 25 µl, containing 1 µl forward and 1 µl reverse primers
(10 µM), 0.5 µl cDNA (500 nM), 0.5 µl dNTP Mix (200 µM), and 1 U
(0.2 µl) GoTaq® DNA polymerase in 1X Green
GoTaq® Reaction Buffer (Promega Corporation).
Thermocycling conditions were as follows: Initial denaturation at
95°C for 2 min, followed by 40 cycles of denaturation at 94°C for
30 sec, annealing for 45 sec at an optimised temperature for each
primer pair, and extension at 72°C for 90 sec, according to the
manufacturer's protocol. Relative gene expression was quantified
according to the comparative Cq method (14) and normalized to GAPDH expression.
All reactions were performed in triplicate.
Western blot analysis
Cells (1.5×106) were homogenized and
lysed in lysis buffer (20 mmol/l HEPES, pH 7.2, 1% Triton X-100,
10% glycerol, 1 mmol/l phenylmethylsulfonyl fluoride, 10 µg/ml
leupeptin, and 10 µg/ml aprotinin; Sigma-Aldrich; Merck KGaA) on
ice for 30 min. Protein concentrations were quantified using the
Bradford assay (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Equal amounts (20 µg) of extracted protein samples were separated
by 12% SDS-PAGE and transferred onto polyvinylidene difluoride
membranes. Following blocking in 5% skimmed milk at 4°C for 1 h,
the membranes were probed with the following primary antibodies at
4°C for 12 h: Anti-SIRT1 (1:1,000; cat. no. 8469; Cell Signaling
Technology, Inc., Danvers, MA, USA), anti-phosphorylated (p)-p65
(1:1,000; cat. no. 3033; Cell Signaling Technology, Inc.),
anti-acetylated-lysine (1:1,000; cat. no. 9441; Cell Signaling
Technology, Inc.), anti-β-actin (1:2,000; cat. no. sc-81178; Santa
Cruz Biotechnology, Inc., Dallas, TX, USA), anti-B-cell lymphoma 2
(Bcl-2; 1:1,000; cat. no. sc-492; Santa Cruz Biotechnology, Inc.),
anti-Bcl-2 associated X protein (Bax; 1:1,000; cat. no. sc-20067;
Santa Cruz Biotechnology, Inc.), anti-caspase 3 (1:1,000; cat. no.
sc-1225; Santa Cruz Biotechnology, Inc.), anti-caspase 9 (1:1,000;
cat. no. sc-133109; Santa Cruz Biotechnology, Inc.),
anti-cleaved-caspase 3 (1:1,000; cat. no. 9664; Cell Signaling
Technology, Inc.), anti-cleaved caspase 9 (1:1,000; cat. no. 9509;
Cell Signaling Technology, Inc.), anti-p-Akt (1:1,000; cat. no.
4060; Cell Signaling Technology, Inc.), and anti-hypoxia inducible
factor-1α (HIF-1α; 1:2,000; cat. no. ab113642; Abcam, Cambridge,
MA, USA). Membranes were then incubated with the following
horseradish peroxidase-conjugated secondary antibodies; goat
anti-mouse immunoglobulin (Ig) G (1:5,000; cat. no. 31430; Thermo
Fisher Scientific, Inc.) and goat anti-rabbit IgG (1:5,000; cat.
no. 31460; Thermo Fisher Scientific, Inc.) at 4°C for 1 h. Protein
bands were visualized using enhanced chemiluminescence (Pierce;
Thermo Fisher Scientific, Inc.) on an Amersham Imager 600UV system
(GE Healthcare Life Sciences, Little Chalfont, UK).
Immunoprecipitation
Whole cell lysates were pre-cleared with Protein A/G
plus (Santa Cruz Biotechnology, Inc.) for 30 min at 4°C. Beads were
pelleted at 1,000 × g for 30 sec at room temperature, and
pre-cleared supernatants were incubated with 10–20 µg primary
antibodies against SIRT1 (1:50; cat. no. 8469; Cell Signaling
Technology, Inc.) and Akt (1:50; cat. no. sc-5298; Santa Cruz
Biotechnology, Inc.), using normal magnetic bead-conjugated IgG
(1:50; cat. no. 5873; Cell Signaling Technology, Inc.) as a
negative control. The reactions were kept on a rotating wheel at
4°C overnight. When agarose or a gel conjugate was unavailable,
lysates were incubated with primary antibody or an equivalent
amount of control IgG for 2 h at 4°C and then overnight along with
Protein A/G plus beads to collect the immune complexes. Beads were
collected by centrifugation at 16,000 × g at 4°C for 10 min, washed
several times with radioimmunoprecipitation assay lysis buffer
(Cell Signaling Technology, Inc.), washed once with PBS, and
re-suspended in SDS-PAGE sample loading buffer (Geno Technology,
Inc., St. Louis, MO, USA). Immune complexes and 75–100 µg input
proteins were resolved by 12% SDS-PAGE.
Statistical analysis
Data are expressed as the mean ± standard error of
the mean of at least 3 independent experiments. The statistical
significance of the differences between groups was assessed using
one-way analysis of variance followed by a post hoc Fisher's
protected least significant difference test for multiple
comparisons. Statistical analysis was performed using GraphPad
Prism software version 5.02 (GraphPad Software, Inc., La Jolla, CA,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Abnormal SIRT1 expression in
hypoxia-induced apoptosis in MC3T3-E1 cells
RT-qPCR and western blot analysis were used to
detect the expression level of SIRT1 in hypoxia-induced apoptosis
in MC3T3-E1 cells. The differences in mRNA expression were compared
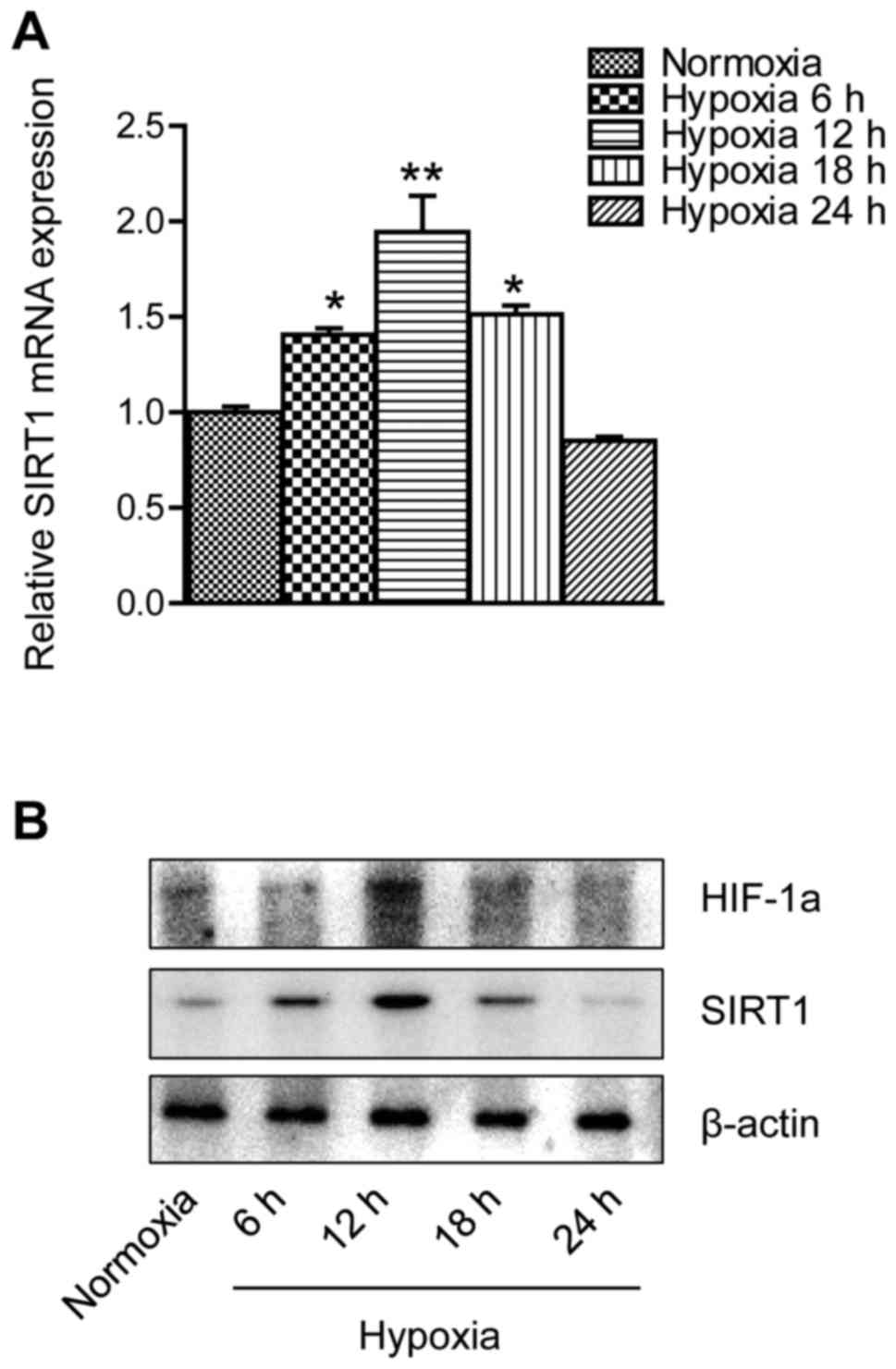
with RT-qPCR at different times under hypoxia. As demonstrated in
Fig. 1A, SIRT1 mRNA appeared to be
elevated in the first 12 h of the experiment but was markedly
downregulated in the later phase. In Fig. 1B, western blot analysis revealed a
pattern of SIRT1 protein expression levels similar to the PCR data.
In addition, HIF-1a protein expression appeared to be upregulated
in the first 12 h of the experiment, whereas it decreased in the
later phase.
SIRT1 interference affects
hypoxia-induced apoptosis in MC3T3-E1 cells
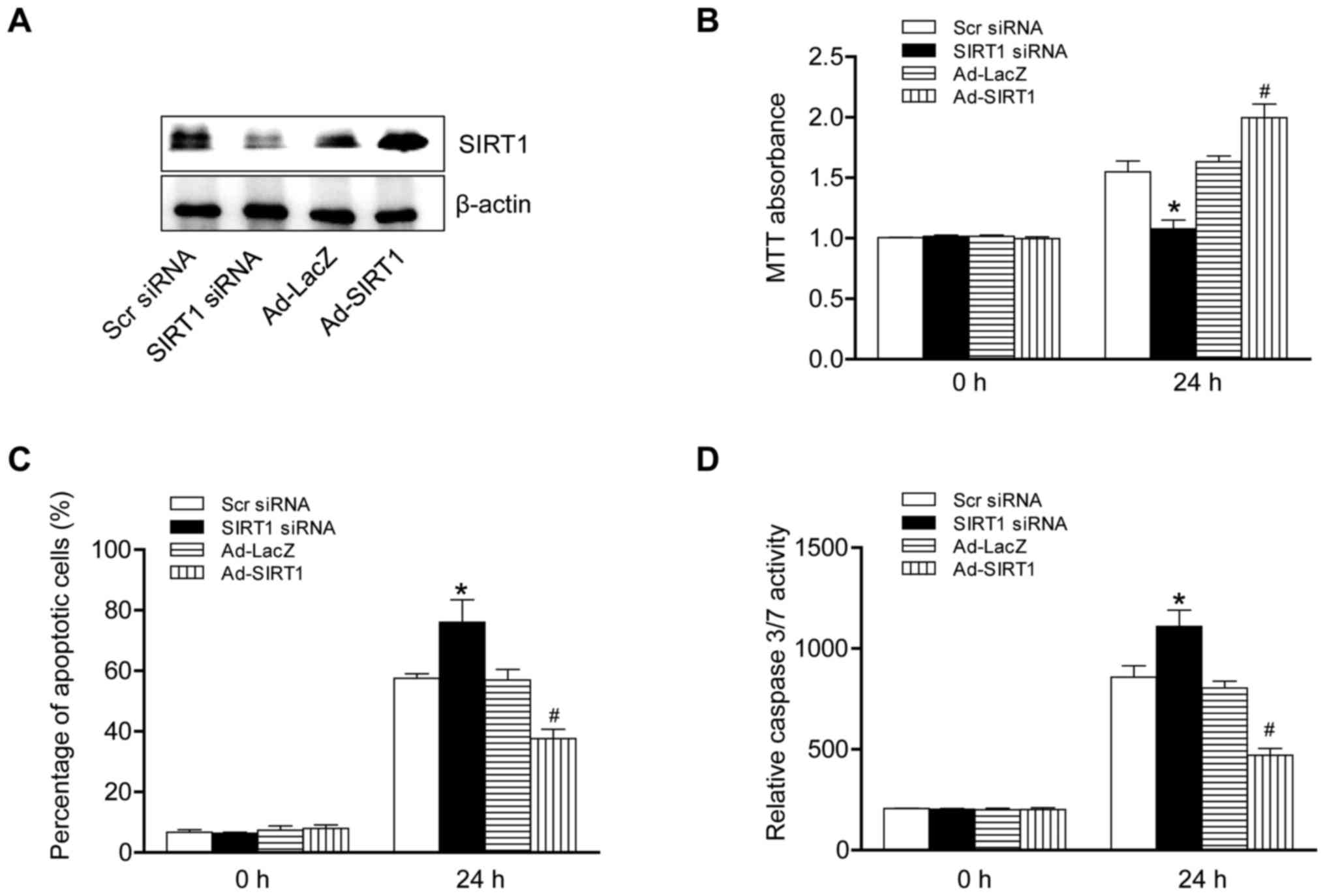
To investigate the role of SIRT1 in hypoxia-induced
apoptosis in MC3T3-E1 cells, the protein levels of SIRT1 were
altered. SIRT1 was overexpressed by using an adenovirus and
knocked-down by si-RNA. The western blotting data demonstrated that
the protein level of SIRT1 was successfully altered by knock-down
or overexpression (Fig. 2A). The
data demonstrated that SIRT1 overexpression in cells significantly
increased cell viability and markedly suppressed the pace of
apoptosis. The activity of the caspase 3/7 pathway was nullified by
SIRT1 overexpression which interrupted the process of apoptosis
response to hypoxia. By contrast, SIRT1 knockdown cells
demonstrated the opposite phenotype (Fig. 2B-D). Taken together, these results
suggested that SIRT1 served a protective effect in hypoxia-induced
apoptosis in MC3T3-E1 cells.
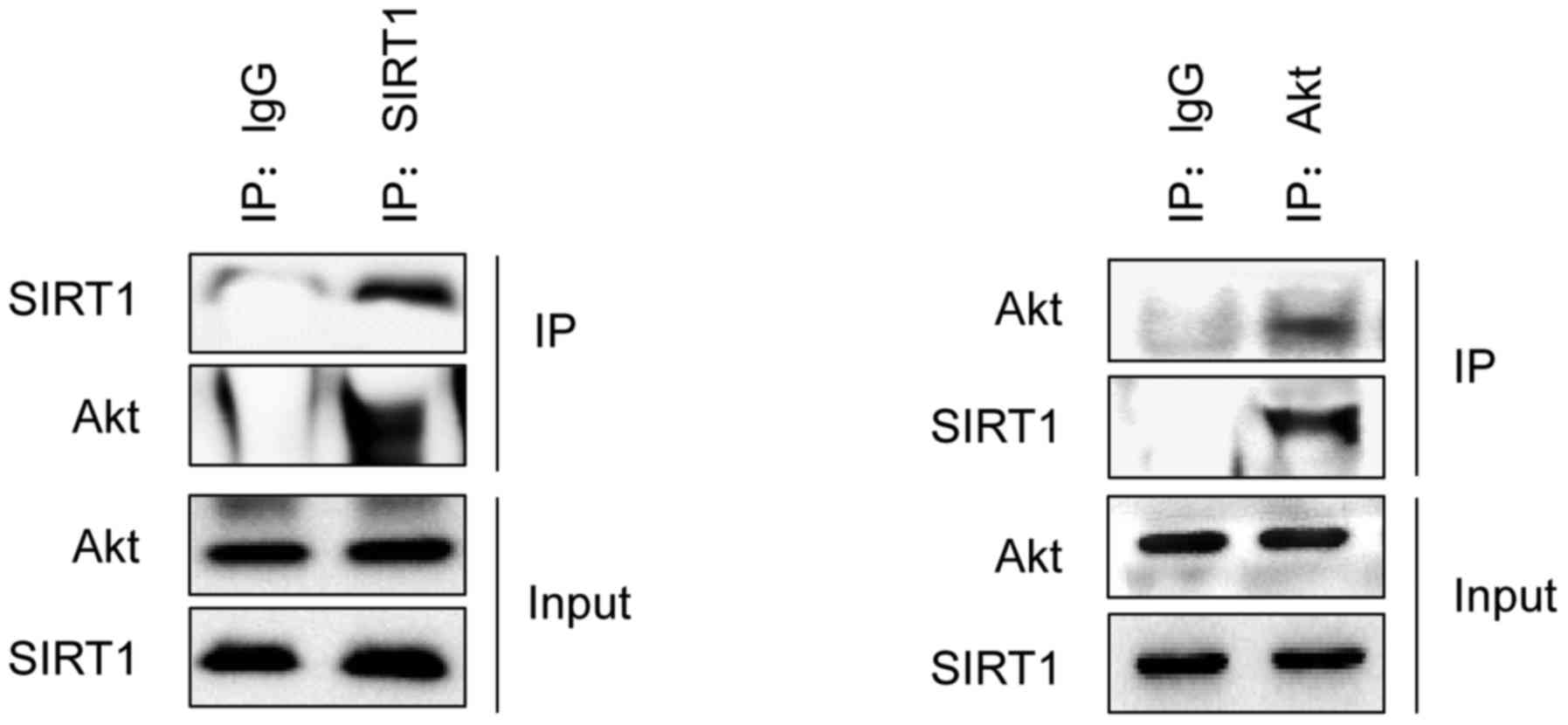
SIRT1 interacts with Akt and
associated binding proteins
To assess the mechanism of SIRT1 under hypoxia, a
search was conducted for its partner proteins. Immunoprecipitation
was performed to detect the co-expression of partner proteins.
SIRT1 was immunoprecipitated from osteoblast lysates and partner
proteins that were pulled down in the precipitate identified
(Fig. 3). It was identified that
Akt, a key protein in the apoptosis pathway, successfully
interacted with SIRT1. The Akt protein pulled down with SIRT1 but
not with the IgG negative control. To obtain further evidence for a
SIRT1/Akt interaction, an inverse experiment was performed in which
SIRT1 was intended to be pulled down by Akt. Lysine is normally an
interaction residue for SIRT1, so it may also interact with Akt. To
confirm this hypothesis, another immunoprecipitation was performed
to demonstrate that Akt interacted with lysine; the resulting beads
were analyzed by western blotting. These results demonstrated that
Akt physically binds to SIRT1 and that lysine is capable of
interacting with Akt.
SIRT1 interference of caspase
expression eliminates hypoxia-induced apoptosis
During the apoptosis process, two signaling pathways
are activated, the canonical and non-canonical. Previous studies
have demonstrated that cytochrome c, apoptotic protease
activating factor 1, and pro-caspase 9 released from the
mitochondria interact with each other to form the apoptosome that
drives the activation of caspase 3 (15,16).
Due to the crucial role of caspases 3 and 9 in the apoptotic
process, their protein expression was detected by western blotting.
The hypoxia treatment potentiated apoptosis and the active forms of
caspases 3 and 9 were increased. A high level of cleaved caspases 3
and 9 were detected. SIRT1 silencing treatment further increased
the amount of cleaved caspases 3 or 9. In contrast, SIRT1
overexpression eliminated the expression of active forms of
caspases under hypoxia (Fig. 4A).
These findings suggest that SIRT1 may exert its regulatory effect
on MC3T3-E1 cell apoptosis by regulating the ratio of pro-caspase
9/caspase 9 and pro-caspase 3/caspase 3.
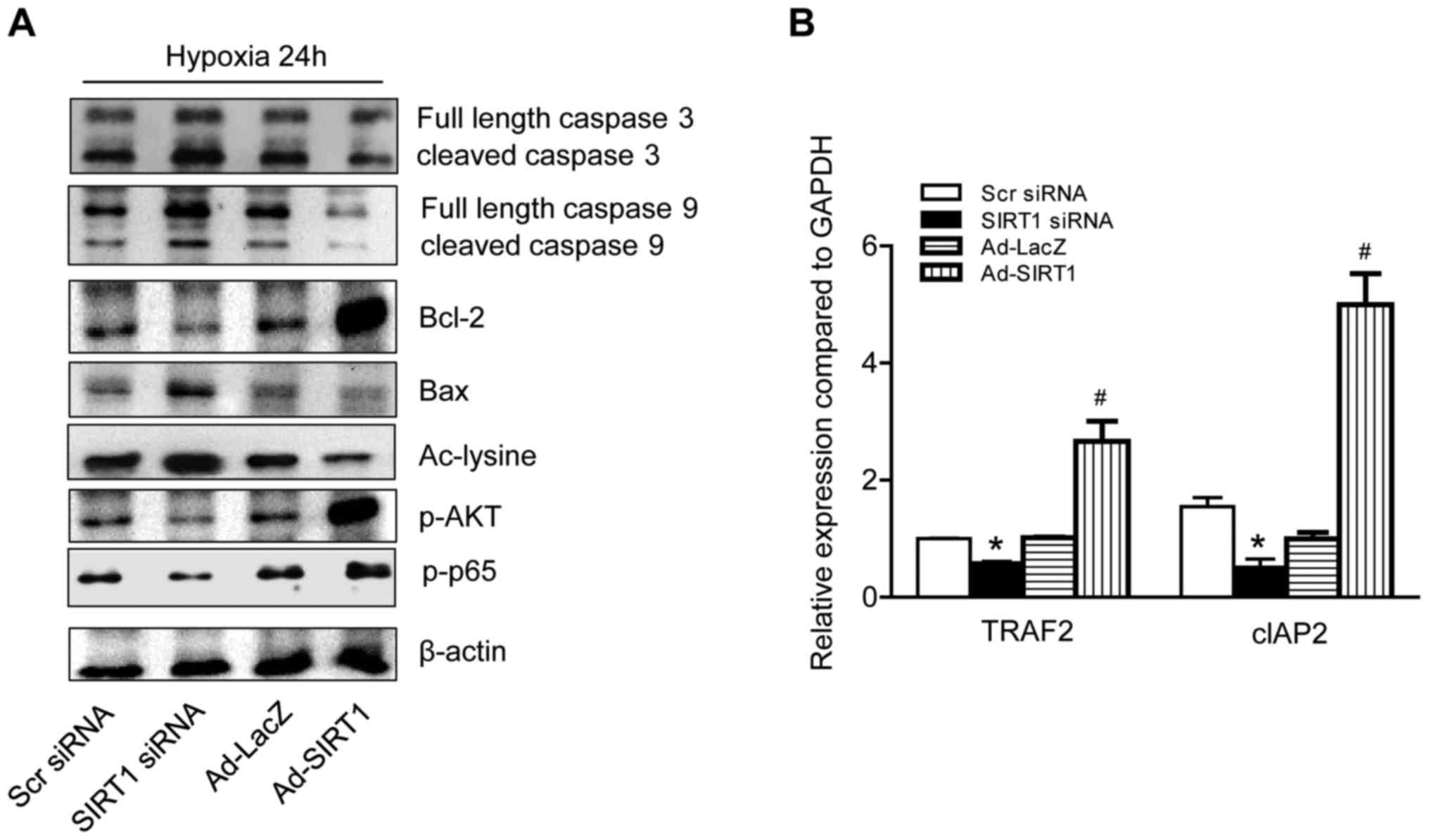 | Figure 4.SIRT1 modifies apoptosis signaling
pathway in hypoxic conditions. (A) MC3T3-E1 cells were transfected
with SIRT1 and SIRT1 siRNA. Western blot analysis was performed to
detect the total amount of Bax, Bcl-2, p-AKT, p-p65 and caspase 3
and 9 expression in cells. (B) MC3T3-E1 cells were treated as
demonstrated in (A), and then were exposed to hypoxia for 24 h. The
group without hypoxia treatment was taken as the control group.
Reverse transcription-quantitative polymerase chain reaction was
performed to detect the transfactors TRAF2 and cIAP2 in the hypoxia
cells. *P<0.05 vs. Scr siRNA group. #P<0.05 vs.
Ad-LacZ group. The data are presented as the mean ± standard
deviation of three independent experiments. SIRT1, nicotinamide
adenosine dinucleotide-dependent deacetylase sirtuin-1; siRNA,
small interfering RNA; Bcl-2, B-cell lymphoma 2; Bax, Bcl-2
associated X, apoptosis regulator; p, phosphorylated; Akt, protein
kinase B; TRAF2, tumor necrosis factor receptor-associated factor
2; Scr, scrambled; Ad, adenovirus; Ad-Lacz, lacZ-bearing
adenovirus. |
SIRT1-activates protective signaling
pathways in hypoxia-induced apoptosis
The Bcl-2 family regulates the canonical pathway of
apoptosis by activation or suppression. Of the Bcl-2 family
members, Bax and Bcl-2 are recognized as two of the most important,
that exert either pro- or anti-apoptotic effects in cells (17). In response to hypoxic stress, Bax
and Bcl-2 protein levels were altered by the differing amounts of
SIRT1 present in the MC3T3-E1 cells (Fig. 4A). Bax was abundant in SIRT1
silenced MC3T3-E1 cells. In contrast, the Bax level was
significantly reduced in SIRT1 overexpressed cells. Bcl-2, the
anti-apoptosis protein, demonstrated the opposite phenotype and was
markedly increased in SIRT1 overexpression cells. Several proteins
or transcription factors were additionally detected using western
blotting or RT-qPCR (Fig. 4B).
Levels of Akt, p65, TRAF2, and cIAP2 were significantly lower in
SIRT1 silenced cells compared with SIRT1 overexpressed cells.
Together, these results suggested that SIRT1 attenuated the
apoptosis of MC3T3-E1 cells by regulating the expression levels of
various proteins.
Discussion
The present study to the best of our knowledge,
presents the first evidence that SIRT1 modulates hypoxia-induced
apoptosis in osteoblast cells through modulated caspase activation.
To date, SIRT1 has been reported to interact with certain nuclear
proteins, including p53 (18) or
fork head family proteins (11,19),
and modulate the functions of these proteins through deacetylation.
The present study demonstrated that SIRT1 may deacetylate Akt and
stimulate a protective effect through phosphatization, which
reduces caspase activation, and that SIRT1 inhibits hypoxia-induced
apoptosis in osteoblasts.
The POPercentage apoptosis assay kit is an efficient
assay for identifying cell apoptosis and providing direct
quantification. Using this assay, it was observed that SIRT1
overexpressed cells exhibited decreased levels of apoptosis. The
caspase 3 and 7 activity assay demonstrated that the canonical
pathway of apoptosis was involved in SIRT1 signaling. Caspases are
a family of protease enzymes serving essential roles in programmed
cell death (20). Caspase 9 is an
initiator caspase in the apoptosis pathway. The aspartic acid
specific protease caspase 9 has been linked to the mitochondrial
death pathway. It improves and activates the effector caspase,
caspase 3, which is a major effector caspase responsible for the
cleavage of cellular substrates during apoptosis (21). In the present study, it was
identified that hypoxia treatment resulted in a significant
increase in cleaved caspases 3 or 9. SIRT1 interference
significantly altered the amount of the cleaved caspase and
abrogated caspase-mediated apoptosis. These results suggested that
SIRT1 regulates the ratio of pro-caspase 9/caspase 9 and
pro-caspase 3/caspase 3, which in turn may affect the development
of MC3T3-E1 cell apoptosis.
SIRT1 is a NAD+-dependent deacetylase and
essential for cell survival, including the process of apoptosis.
The lysine residues on proteins acetylate or deacetylate and may
affect a number of protein functions, including transcriptional
activity, DNA binding, protein binding, protein stability and
translocation. Previously (22),
SIRT1 has been reported to promote cell survival rate through
mediating deacetylation of apoptosis-inducible nuclear proteins.
The phosphorylation of p53, FOXO and Ku70 can inhibit proapoptotic
signalling, as it decreases caspase 3/9 levels (22). Consistent with this finding, it was
hypothesized that SIRT1-mediated phosphorylation of Akt or nuclear
factor κB may also occur through the deacetylation of lysine
residues, thus leading to protein activation and the induction of
cell apoptosis. The western blotting data demonstrated that more
Akt and p65 protein was phosphorylated in SIRT1 overexpressed
cells. The activated proteins promoted transcription in the
nucleus, and upregulated expression of protective genes, including
TRAF2 and cIAPs (23).
These anti-apoptotic genes promote an alternation
between Bax and Bcl-2. Bcl-2 family members form hetero- or
homodimers and act as anti- or pro-apoptotic regulators that are
involved in a wide variety of cellular activities. As two typical
proteins of the Bcl family that restrain and promote apoptosis,
Bcl-2 and Bax serve key roles in regulating the effect of
mitochondrial membrane permeability, mitochondrial function and
cytochrome c release; pro-apoptotic members induce apoptosis
and anti-apoptotic members inhibit apoptosis (24). Therefore, the balance between pro-
and anti-apoptotic members appears to be of critical importance.
SIRT1 knockdown resulted in increased amounts of Bax, a
pro-apoptotic member of the Bcl family. In contrast, the amount of
Bcl-2, an anti-apoptotic member of the Bcl family, was markedly
reduced. These observations suggest that increased Bax coupled with
reduced Bcl-2 expression was, at least in part, one of the
mechanisms resulting in increased apoptosis by the knockdown of
SIRT1. The present study identified that SIRT1 may affect cell
survival by mediating Bax and Bcl-2 expression levels. The hypoxia
stimulus led to a significant increase of Bax. The amount of SIRT1
significantly affected the amount of Bax and Bcl-2 protein,
suggesting that the Bcl family balance serves a critical role in
the development of SIRT-mediated cellular apoptosis.
In summary, SIRT1 was identified as an important
regulator of cellular apoptosis. Its expression was markedly
reduced in hypoxia-induced apoptosis in MC3T3-E1 osteoblast cells.
SIRT1 interference significantly alters MC3T3-E1 cell viability and
affects the number of apoptotic cells and activity of caspases 3
and 7. Furthermore, SIRT1 expression may affect the activity of
PI3K/AKT and NF-κB and alter the Bcl family balance, which in turn
alters the development of MC3T3-E1 cell apoptosis. Therefore, SIRT1
acts as an important regulator of MC3T3-E1 cell apoptosis. SIRT1
intervention may potentially provide a novel strategy for the
treatment of ischemic necrosis of the femoral head by controlling
apoptosis.
Acknowledgements
The present study was supported by the National
Research Foundation of Korea funded by the Korean government (grant
no. NRF-2015R1A2A2A01003762), the Fund of Biomedical Research
Institute of Chonbuk National University Hospital, and the Shandong
Provincial Natural Science Foundation, China (grant no.
ZR2014CQ037).
References
|
1
|
Bejar J, Peled E and Boss JH: Vasculature
deprivation-induced osteonecrosis of the rat femoral head as a
model for therapeutic trials. Theor Biol Med Model. 2:242005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kerachian MA, Harvey EJ, Cournoyer D, Chow
TY and Seguin C: Avascular necrosis of the femoral head: Vascular
hypotheses. Endothelium. 13:237–244. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pop C and Salvesen GS: Human caspases:
Activation, specificity, and regulation. J Biol Chem.
284:21777–21781. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Verdin E, Hirschey MD, Finley LW and
Haigis MC: Sirtuin regulation of mitochondria: Energy production,
apoptosis, and signaling. Trends Biochem Sci. 35:669–675. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Imai S and Guarente L: Ten years of
NAD-dependent SIR2 family deacetylases: Implications for metabolic
diseases. Trends Pharmacol Sci. 31:212–220. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Baur JA, Ungvari Z, Minor RK, Le Couteur
DG and de Cabo R: Are sirtuins viable targets for improving
healthspan and lifespan? Nat Rev Drug Discov. 11:443–461. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Simic P, Zainabadi K, Bell E, Sykes DB,
Saez B, Lotinun S, Baron R, Scadden D, Schipani E and Guarente L:
SIRT1 regulates differentiation of mesenchymal stem cells by
deacetylating β-catenin. EMBO Mol Med. 5:430–440. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Alcendor RR, Gao S, Zhai P, Zablocki D,
Holle E, Yu X, Tian B, Wagner T, Vatner SF and Sadoshima J: Sirt1
regulates aging and resistance to oxidative stress in the heart.
Circ Res. 100:1512–1521. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Takayama K, Ishida K, Matsushita T, Fujita
N, Hayashi S, Sasaki K, Tei K, Kubo S, Matsumoto T, Fujioka H, et
al: SIRT1 regulation of apoptosis of human chondrocytes. Arthritis
Rheum. 60:2731–2740. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vaziri H, Dessain SK, Eaton E Ng, Imai SI,
Frye RA, Pandita TK, Guarente L and Weinberg RA: hSIR2(SIRT1)
functions as an NAD-dependent p53 deacetylase. Cell. 107:149–159.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Motta MC, Divecha N, Lemieux M, Kamel C,
Chen D, Gu W, Bultsma Y, McBurney M and Guarente L: Mammalian SIRT1
represses forkhead transcription factors. Cell. 116:551–563. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lin CH, Lin CC, Ting WJ, Pai PY, Kuo CH,
Ho TJ, Kuo WW, Chang CH, Huang CY and Lin WT: Resveratrol enhanced
FOXO3 phosphorylation via synergetic activation of SIRT1 and
PI3K/Akt signaling to improve the effects of exercise in elderly
rat hearts. Age (Dordr). 36:97052014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang J, Koh HW, Zhou L, Bae UJ, Lee HS,
Bang IH, Ka SO, Oh SH, Bae EJ and Park BH: Sirtuin 2 aggravates
postischemic liver injury by deacetylating mitogen-activated
protein kinase phosphatase-1. Hepatology. 65:225–236. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fister S, Günthert AR, Aicher B, Paulini
KW, Emons G and Gründker C: GnRH-II antagonists induce apoptosis in
human endometrial, ovarian, and breast cancer cells via activation
of stress-induced MAPKs p38 and JNK and proapoptotic protein Bax.
Cancer Res. 69:6473–6481. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yuan S, Yu X, Asara JM, Heuser JE, Ludtke
SJ and Akey CW: The holo-apoptosome: Activation of procaspase-9 and
interactions with caspase-3. Structure. 19:1084–1096. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Birkinshaw RW and Czabotar PE: The BCL-2
family of proteins and mitochondrial outer membrane
permeabilisation. Semin Cell Dev Biol pii. S1084–S9521. 2017.
|
|
18
|
Luo J, Nikolaev AY, Imai S, Chen D, Su F,
Shiloh A, Guarente L and Gu W: Negative control of p53 by Sir2alpha
promotes cell survival under stress. Cell. 107:137–148. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Brunet A, Sweeney LB, Sturgill JF, Chua
KF, Greer PL, Lin Y, Tran H, Ross SE, Mostoslavsky R, Cohen HY, et
al: Stress-dependent regulation of FOXO transcription factors by
the SIRT1 deacetylase. Science. 303:2011–2015. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chipuk JE and Green DR: How do BCL-2
proteins induce mitochondrial outer membrane permeabilization?
Trends Cell Biol. 18:157–164. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang C and Youle RJ: The role of
mitochondria in apoptosis*. Annu Rev Genet. 43:95–118. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cohen HY, Miller C, Bitterman KJ, Wall NR,
Hekking B, Kessler B, Howitz KT, Gorospe M, de Cabo R and Sinclair
DA: Calorie restriction promotes mammalian cell survival by
inducing the SIRT1 deacetylase. Science. 305:390–392. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fu W, Liao X, Ruan J, Li X, Chen L, Wang
B, Wang K and Zhou J: Recombinant human erythropoietin
preconditioning attenuates liver ischemia reperfusion injury
through the phosphatidylinositol-3 kinase/AKT/endothelial nitric
oxide synthase pathway. J Surg Res. 183:876–884. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Leber B, Lin J and Andrews DW: Embedded
together: The life and death consequences of interaction of the
Bcl-2 family with membranes. Apoptosis. 12:897–911. 2007.
View Article : Google Scholar : PubMed/NCBI
|