Introduction
Osteoarthritis (OA) is a degenerative disease of
articular cartilage, which is characterized by local and
progressive destruction of articular cartilage, and osteophyte
formation in the subcartilage edges, and is often accompanied by
synovial inflammation. At present, the pathogenesis of articular
cartilage injury in OA remains to be fully elucidated. Studies in
previous decades have confirmed that oxidative stress is important
in the pathogenesis and progression of OA (1,2).
Oxidative stress is characterized by excessive reactive oxygen
species (ROS) production and reduced cellular antioxidant activity,
leading to membrane lipid peroxidation and mitochondrial
dysfunction (3–5). If oxidative insult persists,
programmed cell death is initiated, which can cause apoptosis of
chondrocytes and extracellular matrix degradation (1,2).
Oxidative stress is primarily caused by the
excessive accumulation of ROS, which include hydrogen peroxide
(H2O2), superoxide anions and hydroxyl
radicals. H2O2 is a major component of ROS
and is used extensively as an inducer in oxidative stress models
(5–9). Therefore, treatment strategies with
the potential to prevent oxidative stress induced by
H2O2 are considered valuable for patients
with OA. Baicalin is a flavonoid glycoside extracted from a type of
traditional Chinese medicine, Scutellaria baicalensis, and
its chemical structure has been determined (10). Baicalin has been shown to possess
antibacterial, anti-inflammatory and anti-apoptotic properties, and
is widely used in the treatment of infectious and inflammatory
diseases (8,11–14).
The aim of the present study was to investigate the
potential protective effects of baicalin treatment on
H2O2-induced oxidative stress in endplate
chondrocytes in vitro, particularly its effect on cell
apoptosis.
Materials and methods
Isolation and culture of primary rat
endplate chondrocytes
A total of 40 4-week-old male Sprague-Dawley rats
weighing 160–200 g were obtained from Shanghai SLAC Laboratory
Animal Co., Ltd. (Shanghai, China) and housed at a temperature of
26°C, 40–60% humidity, standard diet and a 14-h light/10-h dark
cycle. Rat endplate chondrocytes were obtained following cervical
dislocation after anesthesia according to a previous study
(15,16). In brief, the region of the lumbar
posterior midline was shaved and then covered with sterile drapes.
The rats were placed in a prone position on the operating table.
Cartilage of the L3-L5 endplates was carefully removed from the
vertebrae and placed into a small beaker with high-glucose (4.5
g/l; 25 mM) Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA). Sterile ophthalmic
scissors were used to cut the tissues into 1-mm3
sections, and these were transferred to a 50-ml sterile test tube.
The tissues were digested with 0.25% trypsin (Hyclone; GE
Healthcare Life Sciences; Logan, UT, USA) at 37°C for 30 min,
followed by the addition of 0.2% collagenase (Sigma; Merck KGaA;
Darmstadt, Germany) at 37°C for 2 h. Following centrifugation (400
× g for 10 min at room temperature), the supernatant was discarded
and the tissue suspension was washed three times with DMEM with 10%
heat-inactivated fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.). The released cells were cultured in DMEM with
10% FBS, 100 U/ml penicillin and 100 µg/ml streptomycin (both from
Gibco; Thermo Fisher Scientific, Inc.). The cells were maintained
at 37°C in a humidified atmosphere of 5% CO2, and used
at passage 4–6 for all experiments. The present study was approved
by the Ethics Committee of Shanghai East Hospital Affiliated to
Tongji University (Shanghai, China).
Cell viability of endplate
chondrocytes treated with H2O2
The endplate chondrocytes were grown in 96-well
plates (BD Biosciences, Franklin Lakes, NJ, USA) at a density of
~1×105/ml (200 µl/well) in DMEM supplemented with 10%
FBS, 100 U/ml penicillin and 100 µg/ml streptomycin. After 24 h,
the cells were washed with PBS buffer and incubated in serum-free
medium at 37°C for 12 h. The cells were then washed again and
incubated with medium containing different concentrations (0.1, 0.5
and 1.0 mM) of H2O2 (Lingfeng Chemicals Co.,
Ltd., Shanghai, China) at 37°C for 4 h, respectively. Untreated
cells were referred to as the normal control cells. Cell viability
was determined using a CCK-8 assay (Dojindo Molecular Technologies,
Inc., Kumamoto, Japan). The CCK-8 reagent was diluted 10-fold with
DMEM prior to being added (100 ml) to each well. After 2.5 h at
37°C, sample optical density (OD) values were read at 450 nm using
a multimode microplate reader (FlexStation 3; Molecular Devices
LLC, Sunnyvale, CA, USA). The OD450 is proportional to the degree
of cell viability. Data are shown as the mean of at least three
independent experiments.
Annexin V/PI assay of endplate
chondrocytes treated with H2O2
The apoptosis of endplate chondrocytes induced by
H2O2 was measured using a commercial Annexin
V/PI assay kit (Nanjing KeyGen Biotech Co., Ltd., Nanjing, China)
according to the manufacturer's protocol. Briefly, the endplate
chondrocytes were cultured in 6-well plates (BD Biosciences) at a
density of ~1.0×105/ml (2 ml/well) and incubated in DMEM
supplemented with 10% FBS, 100 U/ml penicillin and 100 µg/ml
streptomycin. After 24 h, the cells were washed and incubated in
serum-free medium for 12 h. The cells were then washed again and
incubated in medium with various concentrations of
H2O2 (0.1, 0.5 and 1.0 mM) at 37°C for 4 h.
Following incubation, the cells were trypsinized and washed with
PBS. Following centrifugation at 100 × g for 5 min at room
temperature, the cells were resuspended in 500 µl binding buffer.
The suspensions were transferred into 1.5-ml tubes, and 5 µl of
Annexin V and 5 µl of PI solution were added. The cells were
incubated in the dark at room temperature for 20 min, and flow
cytometric analysis was performed using a BD FACSAria II flow
cytometer (BD Bioscience). Data were analyzed using BD FACSDiva
software version 6.1.3 (BD Bioscience), and Flowjo version 7.6.1
(FlowJo LLC, Ashland, OR, USA). The percentage apoptosis was
determined as the percentage of Annexin V-positive/PI-negative
cells.
Cell viability of endplate
chondrocytes treated with baicalin
The endplate chondrocytes were incubated with 0.5 mM
H2O2 at 37°C and 5% CO2 for 4 h,
with baicalin added at different treatment time-points (−1, 0, 1
and 2 h). Untreated cells and cells treated with
H2O2 only were referred as the control
groups. The viability of the endplate chondrocytes was determined
using a CCK-8 assay, as described above. In addition to determining
the optimal incubation duration, the optimal concentration of
baicalin on endplate chondrocytes was investigated. The cells were
pretreated with baicalin at various concentrations (50, 100 and 200
µmol/l) for 1 h, and were incubated with 0.5 mM
H2O2 at 37°C for 4 h, respectively. Untreated
cells and cells treated with H2O2 only were
referred as the control groups. As pretreatment of endplate
chondrocytes with 100 µmol/l baicalin was optimal for inhibiting
the oxidative stress induced by H2O2, the
cell viability was compared between the
H2O2-only group and the groups treated with
H2O2 at different time-points (4, 8, 12 and
24 h). Untreated cells were referred to as the control group. All
the above data are shown as the mean of at least three independent
experiments.
Annexin V/PI assay of endplate
chondrocytes pretreated with baicalin
The cells were pretreated with 100 µmol/l for 1 h,
and then incubated in medium with 0.5 mM H2O2
at 37°C for 4 h. Based on the previously described method, the
inhibitory effect of baicalin on the apoptosis of endplate
chondrocytes induced by H2O2 was measured
using an Annexin V/PI assay kit.
Western blot analysis
The protein expression of endplate chondrocytes was
evaluated in the groups of chondrocytes treated with 0.5 mM
H2O2 only for 4 h, with 0.5 mM
H2O2 for 4 h+100 µmol/l baicalin pretreatment
for 1 h, and in normal control group. For the extraction of
proteins, the cells were placed in RIPA lysis buffer (Beyotime
Institute of Biotechnology, Haimen, China) for 15 min at 4°C and
centrifuged at 1,300 × g for 30 min at 4°C. The protein
concentrations were determined using a NanoDrop instrument, and 40
µg of protein from each sample were run on a 15% SDS-PAGE gels. The
separated proteins were transferred onto PVDF membranes. Following
blocking with 5% nonfat dry milk in double-distilled water at room
temperature for 1 h, the membranes were washed three times with TBS
containing 0.1% Tween-20 (TBS-T) and incubated overnight at 4°C
with primary mouse monoclonal anti-PARP (ab203467; 1:500),
anti-B-cell lymphoma-2-associated X protein (Bax, ab32503;
1:1,000), anti-pro-caspase-3 (ab13847; 1:500), or anti-GAPDH
antibodies (ab8245; 1:500) (all from Abcam, Cambridge, MA, USA).
The membranes were then washed three times with TBS-T, followed by
1 h incubation at room temperature in a 1:1,000 dilution of goat
anti-mouse/rabbit HRP antibody (715-035-150/415-035-166; Jackson
ImmunoResearch Laboratories, Inc., West Grove, PA, USA). Following
incubation, the membranes were washed three times with TBS-T.
Immune complexes were visualized using enhanced chemiluminescence
followed by exposure to a Tanon 5200 instrument (Tanon Science and
Technology Co., Ltd., Shanghai, China). Data were normalized to the
GAPDH content of the same sample.
Measurement of oxidative activity
The oxidative activities were measured at 6, 12 and
24 h in the 0.5 mM H2O2, 0.5 mM
H2O2+100 µmol/l baicalin pretreatment for 1
h, and normal control groups. The concentrations of malondialdehyde
(MDA), superoxide dismutase (SOD) and nitric oxide (NO) were
assessed using dedicated kits (Nanjing KeyGen Biotech Co., Ltd.)
according to the manufacturer's protocols.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
The mRNA expression levels of endothelial nitric
oxide synthase (eNOS) were determined in the groups treated with
0.5 mM H2O2 only for 4 h, 0.5 mM
H2O2 for 4 h+100 µmol/l baicalin pretreatment
for 1 h, and the normal control group. Total cellular RNA was
extracted from the endplate chondrocytes using TRIzol (Invitrogen;
Thermo Fisher Scientific, Inc.). OLIGO was used as the reverse
transcription primer, and the total RNA was extracted from the
cells of each group as the template, which was then reverse
transcribed into cDNA under the RT enzyme. The reaction system
included 500 ng RNA template, 5X buffer 2 µl, PrimeScript RT enzyme
mix I 0.5 µl, 10 µmol/l 50 M OLIGO 0.5 µl and 100 µM random 6 mers
0.5 µl. DEPC water was added up to 10 µl at 37°C for 15 min. PCR
amplification was performed in a 20 µl reaction volume. The primer
sequences were as follows: eNOS forward,
5′-CCAGCTAGCCAAAGTCACCAT-3′ and reverse,
5′-GTCTCGGAGCCATACAGGATT-3′; GAPDH forward,
5′-CGGAGTCAACGGATTTGGTCGTAT-3′ and reverse,
5′-AGCCTTCTCCATGGTGGTGAAGAC-3′. Triplicate reactions were run with
a volume of 20 µl, containing 2 µl cDNA, 10 µl 2X SYBR-Green mix, 6
µl ddH2O, 1 µl PCR forward primer (10 µM) and 1 µl PCR
reverse primer (10 µM). Following an initial denaturation at 95°C
for 5 min, the PCR conditions were as follows: 35 cycles of
denaturation at 95°C for 30 sec, annealing at 55°C for 30 sec, and
extension at 72°C for 30 sec. The 2−ΔΔCq (quantification
cycle) method (17) was used to
calculate the mRNA expression levels of eNOS in each sample. Data
were normalized to the GAPDH content of the same sample.
Statistical analysis
All data are expressed as the mean ± standard
deviation. Statistical analysis was performed using Student's
t-test and one-way analysis of variance using SPSS version 18.0
software (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered
to indicate a statistically significant difference.
Results
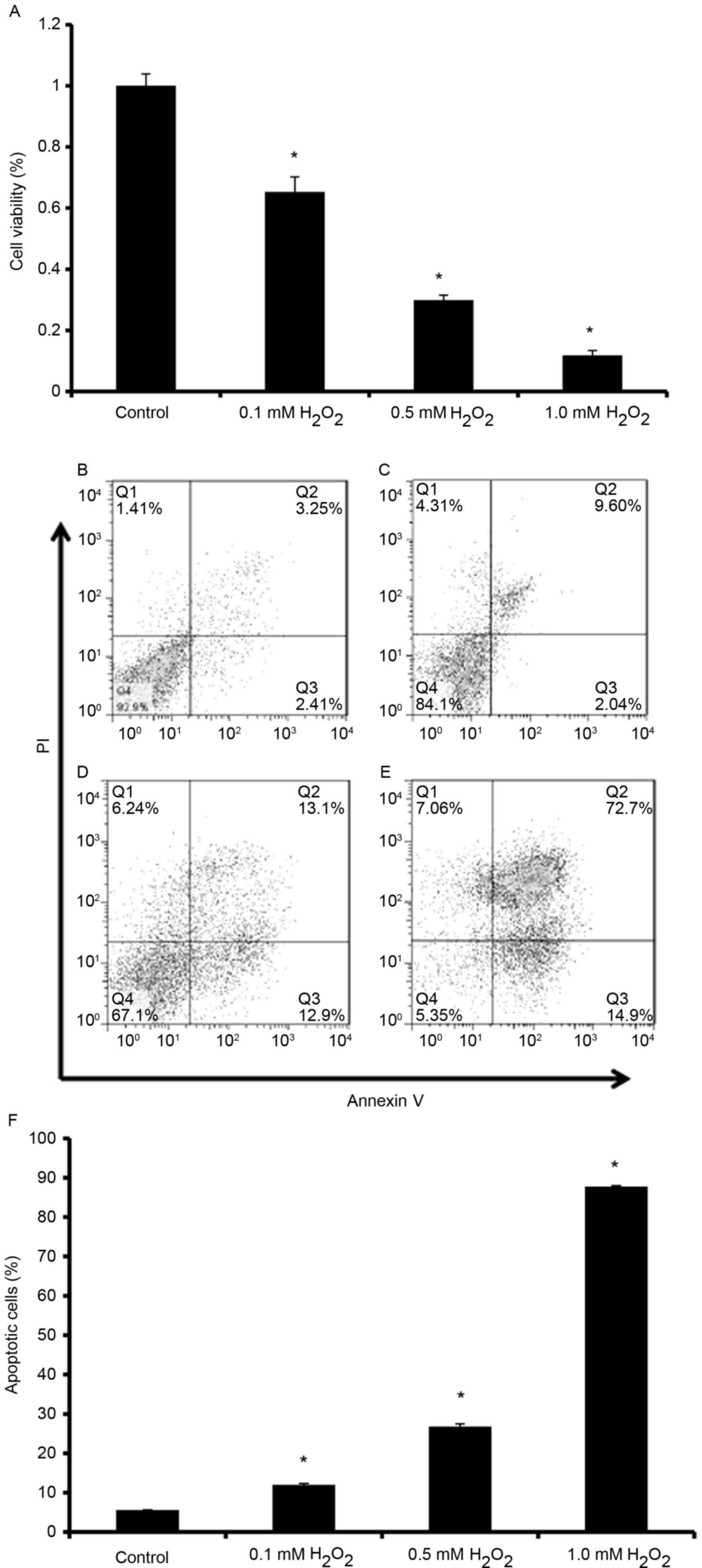
H2O2 induces
apoptotic cell death of endplate chondrocytes
To characterize the effects of
H2O2 on the induction of cell death of
endplate chondrocytes, cell viability and apoptotic rates were
detected. As H2O2 concentration increased,
the cell viability gradually decreased (Fig. 1A; P<0.05), and apoptotic cell
death was significantly increased (Fig. 1B-F; P<0.05). On the basis of
these experiments on cell viability and the Annexin V/PI staining
assay, 0.5 mM H2O2 was selected as a model
dose for the following experiments.
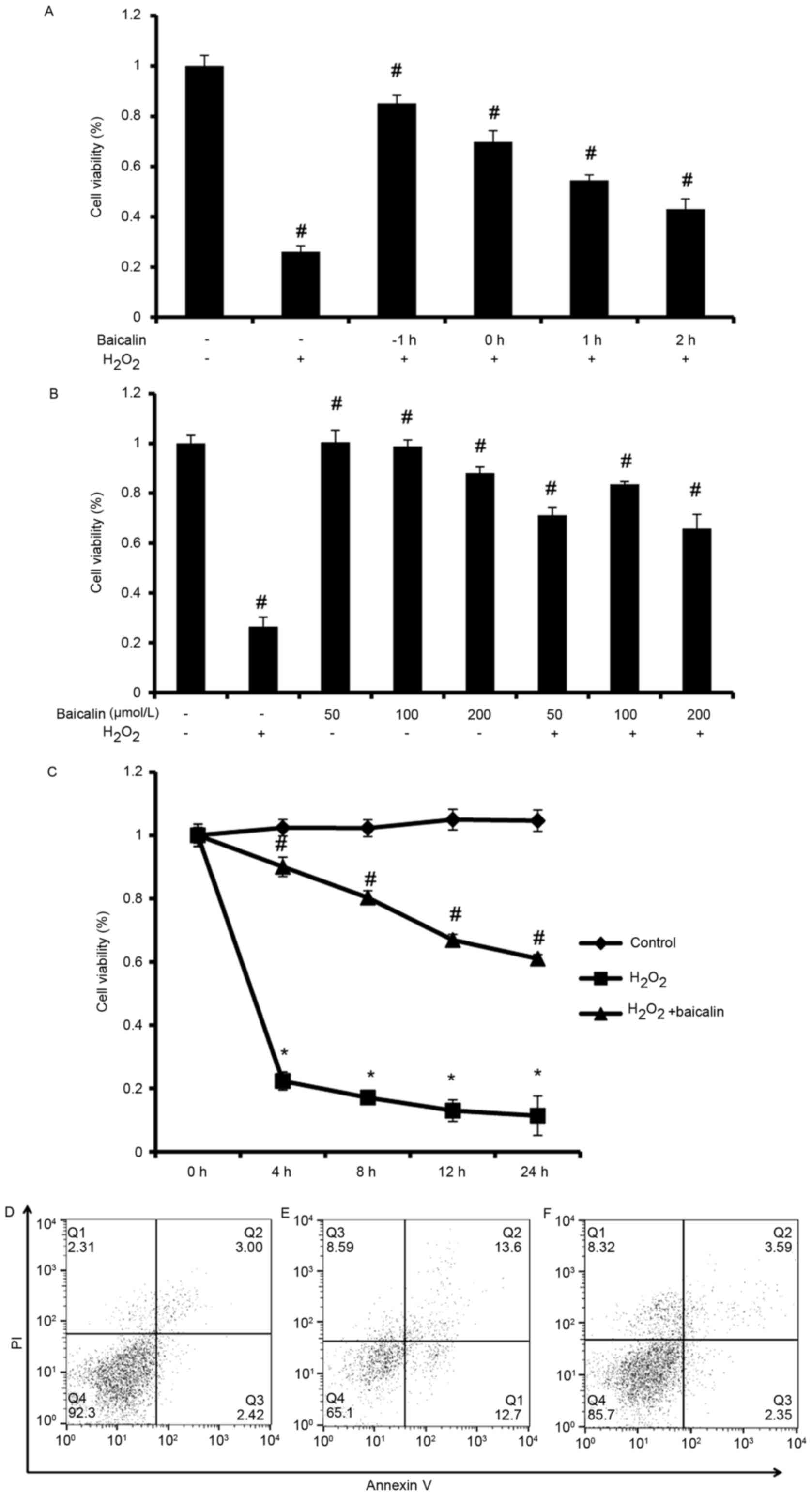
Baicalin inhibits apoptotic cell death
of endplate chondrocytes induced by H2O2
As shown in Fig.
2A, cell viability was highest when endplate chondrocytes were
pretreated with baicalin for 1 h (P<0.05). Subsequently, the
cell viability gradually decreased when the endplate chondrocytes
were treated with baicalin at or following the start of incubation
with H2O2. Without the coexistence of
H2O2, no significant differences in cell
viability were observed among the group pretreated with 50 µmol/l
baicalin, the group pretreated with 100 µmol/l baicalin and the
control group (P>0.05). However, when the pretreatment
concentration of Baicalin reached 200 µmol/l, the cell viability of
endplate chondrocytes was significantly lower, compared with that
in the control (P<0.05, Fig.
2B). Furthermore, in the cells incubated with 0.5 mM
H2O2 for 4 h with baicalin pretreatment for 1
h, the viability of cells in the group pretreated with 100 µmol/l
baicalin was significantly higher, compared with that in the other
two groups (P<0.05), which was considered the optimal dose for
application in the subsequent experiments (Fig. 2B). As shown in Fig. 2C, compared with the group treated
with H2O2 alone, cell death was significantly
decreased in the group treated with
H2O2+baicalin at 4, 8, 12 and 24 h
(P<0.05), which suggested that baicalin may inhibit the cell
apoptosis induced by H2O2. The results of the
Annexin V/PI staining assay (Fig.
2D-F) also showed that baicalin inhibited the cell apoptosis
induced by H2O2.
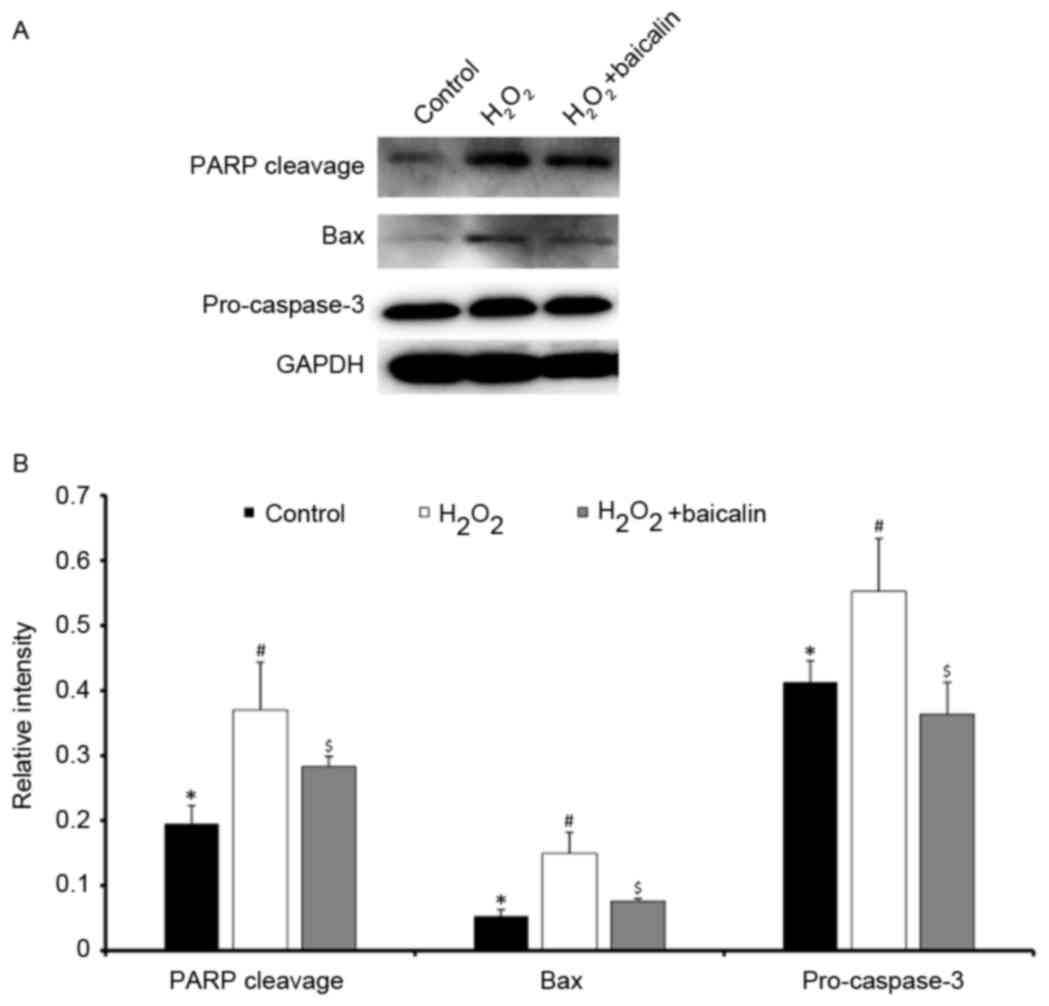
Baicalin inhibits the activation of
the apoptotic cell death pathway induced by
H2O2
Classic apoptotic cell death is induced through a
pathway, which involves the cleavage of PARP and pro-caspase-3, and
the activation of Bax. As shown in Fig. 3A and B, the protein expression of
cleaved PARP, Bax and pro-caspase-3 proteins were determined
following treatment of the endplate chondrocytes with 0.5 mM
H2O2 for 4 h, which were significantly
higher, compared with that in the normal control (P<0.05). By
contrast, baicalin pretreatment downregulated the protein
expression levels of cleaved PARP, Bax and pro-caspase-3
(P<0.05). The quantified the results of western blot analysis
are shown in Fig. 3B.
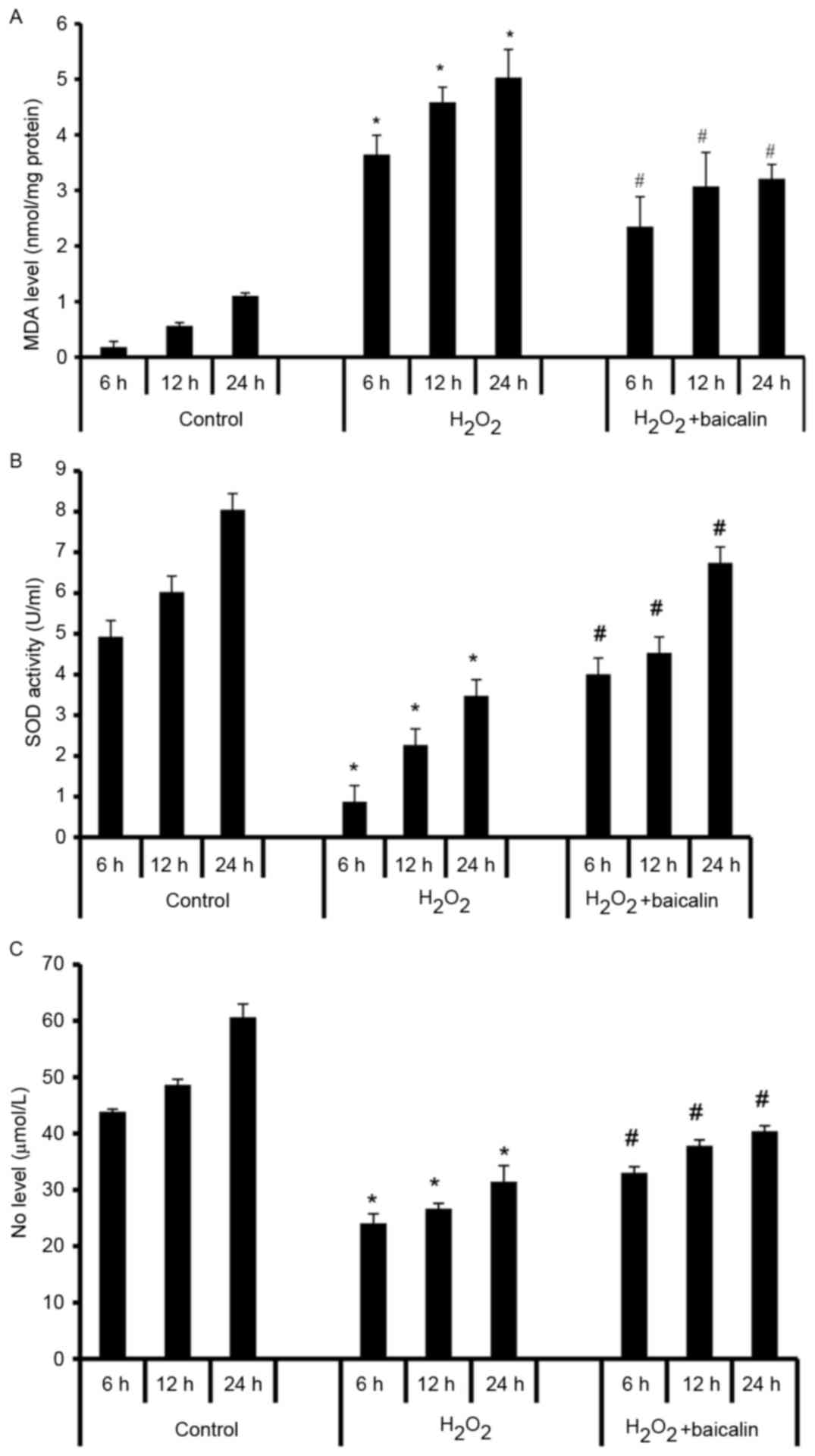
Baicalin decreases the oxidative
activity in endplate chondrocytes induced by
H2O2
As a biomarker of oxidative stress, the levels of
MDA in endplate chondrocytes were increased by
H2O2 treatment at 6, 12 and 24 h (P<0.05),
and this was reversed by baicalin at all time-points (P<0.05;
Fig. 4A). As an enzyme, which
regulates oxidative stress, the levels of SOD were significantly
decreased by exposure to H2O2 (P<0.05),
however, pretreatment with baicalin for 1 h effectively elevated
levels of SOD in endplate chondrocytes at the various time-points
(P<0.05; Fig. 4B). The effects
of baicalin on oxidative activity were further verified by
assessing levels of NO, a free radical signaling mediator. The
levels of NO were significantly decreased in the
H2O2-induced endplate chondrocytes
(P<0.05), and this decrease of NO was reversed by baicalin at 6,
12 and 24 h (P<0.05; Fig. 4C).
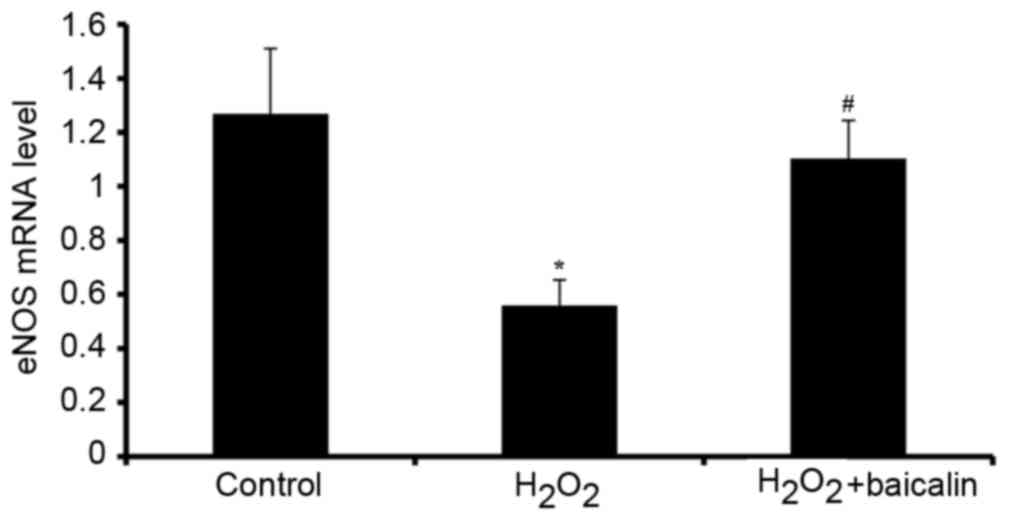
The mRNA levels of eNOS, an enzyme, which functions in catalyzing
the release of NO, were also investigated. The mRNA expression of
eNOS was downregulated by H2O2, and this was
reversed by baicalin (P<0.05; Fig.
5).
Discussion
In the present study, a model of 0.5 mM
H2O2-induced oxidative stress was
successfully induced in endplate chondrocytes, which was
characterized by reduced cell viability, increased intracellular
ROS and reduced intracellular antioxidant activity. These
pathophysiological processes led to apoptosis by activation of the
intrinsic apoptotic pathway. As a flavonoid glycoside extracted
from a type of traditional Chinese medicine, Scutellaria
baicalensis, baicalin has been reported to significantly
attenuate oxidative injury, and partially prevent apoptosis during
the oxidative stress reaction (8,10–14,18).
For example, Lin et al (8,11)
found that baicalin ameliorated H2O2-induced
cytotoxicity in a human renal proximal tubular epithelial cell line
(HK-2 cells), and showed effects against renal ischemia-reperfusion
injury through the inhibition of inflammation and apoptosis.
Therefore, the present study investigated the potential protective
effects of baicalin treatment on H2O2-induced
oxidative stress in endplate chondrocytes in vitro.
Initially, different time-points of induction with
baicalin incubation under H2O2 stimulation
were investigated, which showed that 1 h pretreatment with baicalin
exhibited optimal protective effects. Secondly, it was observed
that the potential protective effects of baicalin pretreatment
against oxidative stress-related injury in endplate chondrocytes
were dose-dependent. However, baicalin significantly decreased cell
viability when its concentration reached 200 µmol/l. Therefore, to
achieve optimal protective effects, 100 µmol/l baicalin was
selected as the optimal concentration for inhibiting oxidative
stress in endplate chondrocytes induced by
H2O2.
In the present study, the protective effects of
baicalin on oxidative stress of endplate chondrocytes induced by
H2O2 were confirmed through the analysis of
cell viability. The cell viability of endplate chondrocytes
stimulated by H2O2 was significantly lower,
compared with that in than the normal control, as detected by the
CCK-8 assay. Notably, a significant increase in cell viability was
observed following baicalin pretreatment for 1 h at the various
time-points. Baicalin was found to inhibit the activation of the
apoptotic cell death pathway triggered by
H2O2. Classic apoptotic cell death is induced
through a pathway involving the cleavage of PARP and pro-caspase-3,
and the activation of Bax (19–21).
The abnormally high expression levels of cleaved PARP, Bax and
pro-caspase-3 induced by H2O2 were
significantly reversed by pretreatment with baicalin. In addition,
baicalin suppressed oxidative activity in the endplate chondrocytes
induced by H2O2, via effectively reducing
levels of MDA, increasing levels of SOD, and elevating NO
activities (5,9,22).
eNOS, an enzyme which activates the expression of NO (23–25),
was also shown to be increased following pretreatment with
baicalin, which suggested that baicalin also inhibited the
apoptosis through upregulating the expression of eNOS.
However, the mechanism of baicalin-mediated
protection against oxidative stress remains to be fully elucidated.
Chen et al (18) reported
that baicalin inhibited oxidative-stress-induced apoptosis via
modulating the activation of extracellular signal-regulated kinases
and inducing the gene expression of heme oxygenase-1 in rat C6
glioma cells. Lin et al (8)
reported that the effect of baicalin was through the inhibition of
endoplasmic reticulum stress and the activation of nuclear factor
erythroid 2-related factor 2 signaling, which were important during
baicalin-mediated protection. The targets of baicalin also remain
to be fully elucidated. A previous study indicated that baicalin
activates AMP-activated protein kinase (AMPK) through the
Ca2+/calmodulin-dependent protein kinase kinase β
(CaMKKβ)-dependent pathway in HeLa and A549 cells (26). AMPK is important in cell physiology
and also affects the cell response to oxidative stress. However, at
present, no direct association between the renal protective effects
of baicalin and activation of the Ca2+/CaMKKβ-AMPK
pathway has been reported. Peroxisome proliferator-activated
receptor-γ (PPARγ) has also been suggested as a target of baicalin
(27). Baicalin was found to
activate PPARγ and suppress downstream nuclear factor-κB-mediated
inflammation in aged rat kidneys. In addition, studies have
suggested other potential targets, including the proteasome
(28), macrophages (29) and notch signaling (30). These possibilities were not
examined in the present study, and warrant further
investigation.
In conclusion, the results of the present study
suggested that baicalin pretreatment protected endplate
chondrocytes from the oxidative stress-related damage induced by
H2O2. The role of baicalin was predominantly
based on inhibiting the production of ROS, increasing intracellular
antioxidants and attenuating apoptosis. The results of the present
study require verification in future in vivo investigations,
however, the results provide further insight into the potential
benefits of baicalin for patients with oxidative stress-related
diseases, including OA.
Acknowledgements
This study was supported by the National Science
Foundation for Young Scholars of China (grant no. 81601675 to Dr
Yutao Pan).
References
|
1
|
Henrotin YE, Bruckner P and Pujol JP: The
role of reactive oxygen species in homeostasis and degradation of
cartilage. Osteoarthritis Cartilage. 11:747–755. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Henrotin Y, Kurz B and Aigner T: Oxygen
and reactive oxygen species in cartilage degradation: Friends or
foes? Osteoarthritis Cartilage. 13:643–654. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rocha M, Apostolova N, Hernandez-Mijares
A, Herance R and Victor VM: Oxidative stress and endothelial
dysfunction in cardiovascular disease: Mitochondria-targeted
therapeutics. Curr Med Chem. 17:3827–3841. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ying W and Xiong ZG: Oxidative stress and
NAD+ in ischemic brain injury: Current advances and future
perspectives. Curr Med Chem. 17:2152–2158. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mao CY, Lu HB, Kong N, Li JY, Liu M, Yang
CY and Yang P: Levocarnitine protects H9c2 rat cardiomyocytes from
H2O2-induced mitochondrial dysfunction and
apoptosis. Int J Med Sci. 11:1107–1115. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nogueira-Pedro A, Cesário TA, Dias CC,
Origassa CS, Eça LP, Paredes-Gamero EJ and Ferreira AT: Hydrogen
peroxide (H2O2) induces leukemic but not
normal hematopoietic cell death in a dose-dependent manner. Cancer
Cell Int. 13:1232013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pomari E, Stefanon B and Colitti M: Effect
of plant extracts on H2O2-induced
inflammatory gene expression in macrophages. J Inflamm Res.
7:103–112. 2014.PubMed/NCBI
|
|
8
|
Lin M, Li L, Zhang Y, Zheng L, Xu M, Rong
R and Zhu T: Baicalin ameliorates H2O2
induced cytotoxicity in HK-2 cells through the inhibition of ER
stress and the activation of Nrf2 signaling. Int J Mol Sci.
15:12507–12522. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen S, Tang Y, Qian Y, Chen R, Zhang L,
Wo L and Chai H: Allicin prevents
H2O2-induced apoptosis of HUVECs by
inhibiting an oxidative stress pathway. BMC Complement Altern Med.
14:3212014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cao Y, Mao X, Sun C, Zheng P, Gao J, Wang
X, Min D, Sun H, Xie N and Cai J: Baicalin attenuates global
cerebral ischemia/reperfusion injury in gerbils via anti-oxidative
and anti-apoptotic pathways. Brain Res Bull. 85:396–402. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lin M, Li L, Li L, Pokhrel G, Qi G, Rong R
and Zhu T: The protective effect of baicalin against renal
ischemia-reperfusion injury through inhibition of inflammation and
apoptosis. BMC Complement Altern Med. 14:192014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tang YJ, Zhou FW, Luo ZQ, Li XZ, Yan HM,
Wang MJ, Huang FR and Yue SJ: Multiple therapeutic effects of
adjunctive baicalin therapy in experimental bacterial meningitis.
Inflammation. 33:180–188. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhu J, Wang J, Sheng Y, Zou Y, Bo L, Wang
F, Lou J, Fan X, Bao R, Wu Y, et al: Baicalin improves survival in
a murine model of polymicrobial sepsis via suppressing inflammatory
response and lymphocyte apoptosis. PLoS One. 7:e355232012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xiping Z, Guanghua F, Jinxian H, Weihong
W, Rujun X, Wei Z, Jing Y, Qijun Y, Meijuan Y, Qing W and Lini F:
Baicalin protects thymus of rats with severe acute pancreatitis.
Inflammation. 33:157–165. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang YJ, Shi Q, Sun P, Zhou Q, Darowish M,
Li TF, Dong YF, Lu WW and Leong JC: Insulin-like growth factor-1
treatment prevents anti-Fas antibody-induced apoptosis in endplate
chondrocytes. Spine (Phila Pa 1976). 31:736–741. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yuan FL, Wang HR, Zhao MD, Yuan W, Cao L,
Duan PG, Jiang YQ, Li XL and Dong J: Ovarian cancer G
protein-coupled receptor 1 is involved in acid-induced apoptosis of
endplate chondrocytes in intervertebral discs. J Bone Miner Res.
29:67–77. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen YC, Chow JM, Lin CW, Wu CY and Shen
SC: Baicalein inhibition of oxidative-stress-induced apoptosis via
modulation of ERKs activation and induction of HO-1 gene expression
in rat glioma cells C6. Toxicol Appl Pharmacol. 216:263–273. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Boulares AH, Yakovlev AG, Ivanova V,
Stoica BA, Wang G, Iyer S and Smulson M: Role of poly(ADP-ribose)
polymerase (PARP) cleavage in apoptosis. Caspase 3-resistant PARP
mutant increases rates of apoptosis in transfected cells. J Biol
Chem. 274:22932–22940. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ruemmele FM, Dionne S, Qureshi I, Sarma
DS, Levy E and Seidman EG: Butyrate mediates Caco-2 cell apoptosis
via up-regulation of pro-apoptotic BAK and inducing caspase-3
mediated cleavage of poly-(ADP-ribose) polymerase (PARP). Cell
Death Differ. 6:729–735. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hoetelmans R, van Slooten HJ, Keijzer R,
Erkeland S, van de Velde CJ and Dierendonck JH: Bcl-2 and Bax
proteins are present in interphase nuclei of mammalian cells. Cell
Death Differ. 7:384–392. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liao JK, Shin WS, Lee WY and Clark SL:
Oxidized low-density lipoprotein decreases the expression of
endothelial nitric oxide synthase. J Biol Chem. 270:319–324. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Knowles RG and Moncada S: Nitric oxide
synthases in mammals. Biochem J. 298:249–258. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen CA, Wang TY, Varadharaj S, Reyes LA,
Hemann C, Talukder MA, Chen YR, Druhan LJ and Zweier JL:
S-glutathionylation uncouples eNOS and regulates its cellular and
vascular function. Nature. 468:1115–1118. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wu JR, Hsu JH, Dai ZK, Wu BN, Chen IJ,
Liou SF and Yeh JL: Activation of endothelial NO synthase by a
xanthine derivative ameliorates hypoxia-induced apoptosis in
endothelial progenitor cells. J Pharm Pharmacol. 68:810–818. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ma Y, Yang F, Wang Y, Du Z, Liu D, Guo H,
Shen J and Peng H: CaMKKβ is involved in AMP-activated protein
kinase activation by baicalin in LKB1 deficient cell lines. PLoS
One. 7:e479002012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lim HA, Lee EK, Kim JM, Park MH, Kim DH,
Choi YJ, Ha YM, Yoon JH, Choi JS, Yu BP and Chung HY: PPARγ
activation by baicalin suppresses NF-κB-mediated inflammation in
aged rat kidney. Biogerontology. 13:133–145. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wu YX, Sato E, Kimura W and Miura N:
Baicalin and scutellarin are proteasome inhibitors that
specifically target chymotrypsin-like catalytic activity. Phytother
Res. 27:1362–1367. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu LL, Gong LK, Wang H, Xiao Y, Wu XF,
Zhang YH, Xue X, Qi XM and Ren J: Baicalin inhibits macrophage
activation by lipopolysaccharide and protects mice from endotoxin
shock. Biochem Pharmacol. 75:914–922. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang AM, Ku HH, Liang YC, Chen YC, Hwu YM
and Yeh TS: The autonomous notch signal pathway is activated by
baicalin and baicalein but is suppressed by niclosamide in K562
cells. J Cell Biochem. 106:682–692. 2009. View Article : Google Scholar : PubMed/NCBI
|