Introduction
Amyotrophic lateral sclerosis (ALS) is a
neurodegenerative disease, characterized by the loss of motor
neurons in the motor cortex, nucleus of the brainstem and anterior
horn of the spinal cord (1). The
mechanism of the degeneration of motor neurons in ALS is not well
defined and there is no effective therapy that has the ability to
block ALS progression. The majority of ALS patients only survive
3–5 years following diagnosis (1).
Accumulating evidence suggests that iron homeostasis
is disordered in ALS. Jeong et al (2) demonstrated that iron accumulation in
the spinal cord of super oxide dismutase (SOD1)G37R
transgenic mice at 12 months of age and the iron influx proteins
[transferrin receptor (TfR) and divalent metal-ion transporter 1
(DMT1)], increased significantly compared with wild-type control
mice. Hadzhieva et al (3)
observed an increased total iron content in G93A-SOD1 SH-SY5Y
neuroblastoma cells, and mRNA expression of TfR and DMT1 was
increased in G93A-SOD1 cells. In ALS patients, the level of iron in
the cerebrospinal fluid was higher than the levels in control
subjects (4). In addition, the
serum ferritin level was demonstrated to be elevated in patients
with ALS (5). Langkammer et
al (6) identified increased
iron deposition in localized regions along the corticospinal tract
in ALS patients (6). Corroborating
these findings, it was previously reported that phase shift values
were significantly higher in the motor cortex of patients with ALS
using susceptibility weighted imaging, indicating increased iron
levels in this area (7).
Additionally, pathological studies have demonstrated increased iron
accumulation in the motor cortex of patients with ALS (8).
As it appears that iron levels are affected in ALS,
it is possible that drugs regulating iron metabolism may be useful
for treatment of ALS. Treatment of SOD1G37R transgenic
mice with an iron chelator extended the life span by 5 weeks
(2). It was also previously
reported that epigallocatechin-3-gallate (EGCG) may act as an iron
chelator to treat neurodegenerative diseases (9,10).
EGCG is the major constituent of green tea polyphenols, accounting
for >10% of its composition. EGCG is a natural anticancer agent,
and also demonstrated potential neuroprotective functions (11). In previous studies, EGCG has
exhibited multifunctional therapeutic effects in a mouse model of
ALS (12,13). Thus, the effects of EGCG on iron
metabolism were investigated in an ALS model.
Materials and methods
Materials
Threo-hydroxyaspartate (THA) was purchased from
Sigma-Aldrich; Merck KGaA (catalog no. H2775; Darmstadt, Germany).
The antibody against neurofilament (SMI-32) was purchased from
Covance, Inc. (catalog no. SMI-32R; Princeton, NJ, USA) and the
antibody against TfR was purchased from Invitrogen; Thermo Fisher
Scientific, Inc. (catalog no. 13-6800; Waltham, MA, USA).
Antibodies against DMT1 (catalog no. sc-30120) and β-actin (catalog
no. sc-47778) were purchased from Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA).
Organotypic culture of rat spinal
cord
Organotypic spinal cord cultures were prepared as
described previously (14–16). Briefly, lumbar spinal cords were
removed from 6 to 8-day-old male and female Sprague Dawley rats
(Animal Center of Hebei Medical University, Shijiazhuang, China)
under sterile conditions and sectioned transversely at 350 µm
intervals using a tissue chopper (Mickle Laboratory Engineering
Co., Ltd., Guildford, UK). The rats were housed in clear plastic
cages with sawdust bedding at standard room temperature under a 12
h light/dark cycle. All rats had free access to food and water.
Sections were quickly transferred to sterile Gey's balanced salt
solution containing glucose (6.4 mg/ml) and separated from one
another at room temperature. The tissue slices were placed on the
surface of 30 mm Millipore Millicell-CM membranes (EMD Millipore,
Billerica, MA, USA), five slices/membrane, and each membrane was
then placed in a 33 mm culture well containing 1 ml medium, which
consisted of 50% (v/v) minimal essential medium (Gibco; Thermo
Fisher Scientific, Inc.) with 25 mM HEPES, 25% (v/v)
heat-inactivated horse serum (Gibco; Thermo Fisher Scientific,
Inc.) (56°C for 30 min), and 25% (v/v) Hanks' balanced salt
solution (supplemented with 25.6 mg/ml D-glucose and 2 mM
glutamine). The cultures were maintained at 37°C in a humidified
incubator with 5% CO2 for up to 4 weeks. Culture medium,
along with test chemicals, was changed twice per week, unless
specified otherwise. THA was dissolved in water. EGCG was dissolved
in dimethyl sulfoxide (DMSO), and the final concentration of DMSO
was 0.1% in culture medium. The animal experimental protocol was
approved by the Animal Care Committee of Linyi People's Hospital
(Linyi, China).
Treatments on organotypic
cultures
Unless otherwise stated, the spinal cord explants
were initially cultured in vitro for 7 days. On day 7, the
explants were pretreated with 5 µM EGCG for 48 h, and then treated
with the combination of 5 µM EGCG and 100 µM THA for 3 weeks. The
concentration of 5 µM was identified as the effective dose of EGCG
on the spinal cord explants in the authors' preliminary experiments
(16). The culture medium along
with test chemicals was replaced twice per week. At the end of the
3 week treatment, the explants were harvested for analysis.
Immunohistochemical staining
Immunohistochemical staining was used to visualize
motor neurons in spinal cord explants. The explants, at the end of
an experimental treatment, were fixed with 4% paraformaldehyde in
0.1 M PBS (pH 7.4) for 40 min at room temperature, rinsed three
times with 0.1 M PBS, and stored in 0.1 M PBS at 4°C before use.
The explants were washed in TBS for 30 min and then treated with
10% horse serum (Gibco; Thermo Fisher Scientific, Inc.) for 1 h at
room temperature. The explants were subsequently incubated with the
anti-neurofilament antibody (SMI-32: 1:1,000) overnight at 4°C,
followed by washing with TBS and Tween-20 three times and incubated
with a biotinylated secondary antibody (1:1,000; catalog no.
SP-9002; ZSGB-BIO, Beijing, China) for 1 h at room temperature. The
explants were further washed and then incubated with a horseradish
peroxidase-conjugated ABC staining solution (ZSGB-BIO). The
explants were finally mounted on glass slides, and motor neurons in
the ventral horns, which were stained dark brown, were counted
under a light microscope. This is similar to the previously
reported methods (15,16).
Measurement of malondialdehyde
(MDA)
MDA is one of the most important degradation
products of lipid peroxidation. It reacts with thiobarbituric acid
to produce a product that can be sensitively measured
spectroscopically. Following the manufacturer's instructions,
enzyme activity was determined by monitoring the change in
absorbance at 532 nm using a cell malondialdehyde assay kit from
Nanjing Jiancheng Biongineering Institute (Nanjing, China).
Measurement of the total iron
The total iron content in spinal cord tissue was
determined, as described previously (15). The specimens were torrefied at
110°C for 4 h and then examined using a graphite furnace atomic
absorption spectrophotometer (AAnalyst 100; PerkinElmer, Inc.,
Waltham, MA, USA) by a solid sampling system (SSA 61Z; Analytik
Jena AG, Jena, Germany). Absorbance was read at 248.8 nm, and iron
content was calculated using a calibration curve prepared with pure
iron.
Western blot analysis
Spinal cord explants were processed at the end of an
experimental treatment to prepare whole tissue extracts, using a
tissue extraction reagents kit from Applygen Technologies, Inc.
(Beijing, China). The extraction of protein was quantified using a
bicinchoninic acid assay. A total of 60 µg of extracted protein was
resolved by 10% SDS-PAGE and transferred to polyvinylidene
difluoride membranes. The membranes were blocked with 5% skimmed
milk for 1 h at room temperature, and then probed with specific
primary antibodies [mouse monoclonal anti-TfR (catalog no.
13-6800), 1:500; rabbit polyclonal anti-DMT1 (catalog no.
sc-30120), 1:200; mouse monoclonal anti-β-actin (catalog no.
sc-47778), 1:500)] overnight at 4°C, followed with secondary
antibodies [goat anti-mouse IgG-horseradish peroxidase (HRP),
1:10,000, catalog no. sc-2005; goat anti-rabbit IgG-HRP, 1:3,000,
catalog no. sc-2004 (Santa Cruz Biotechnology, Inc.)] for 1 h at
room temperature, and detection was performed with an enhanced
chemiluminescence substrate (Beyotime Institute of Biotechnology,
Haimen, China). The data were obtained by measuring the density of
target protein banding to the density of corresponding β-actin
bands, and Quantity One software version 4.6.7 (Bio-Rad
Laboratories lnc., Hercules, CA, USA) was used for
quantification.
Statistical analysis
Results are expressed as the mean ± standard
deviation. Statistical analyses were performed using the SAS system
(SAS Institute Inc., Cary, NC, USA) for Microsoft Windows version 8
(Microsoft Corporation, Redmond, WA, USA). A one-way analysis of
variance followed by the Student-Newman-Keuls test was applied to
analyze the data. P<0.05 was considered to indicate a
statistically significant difference.
Results
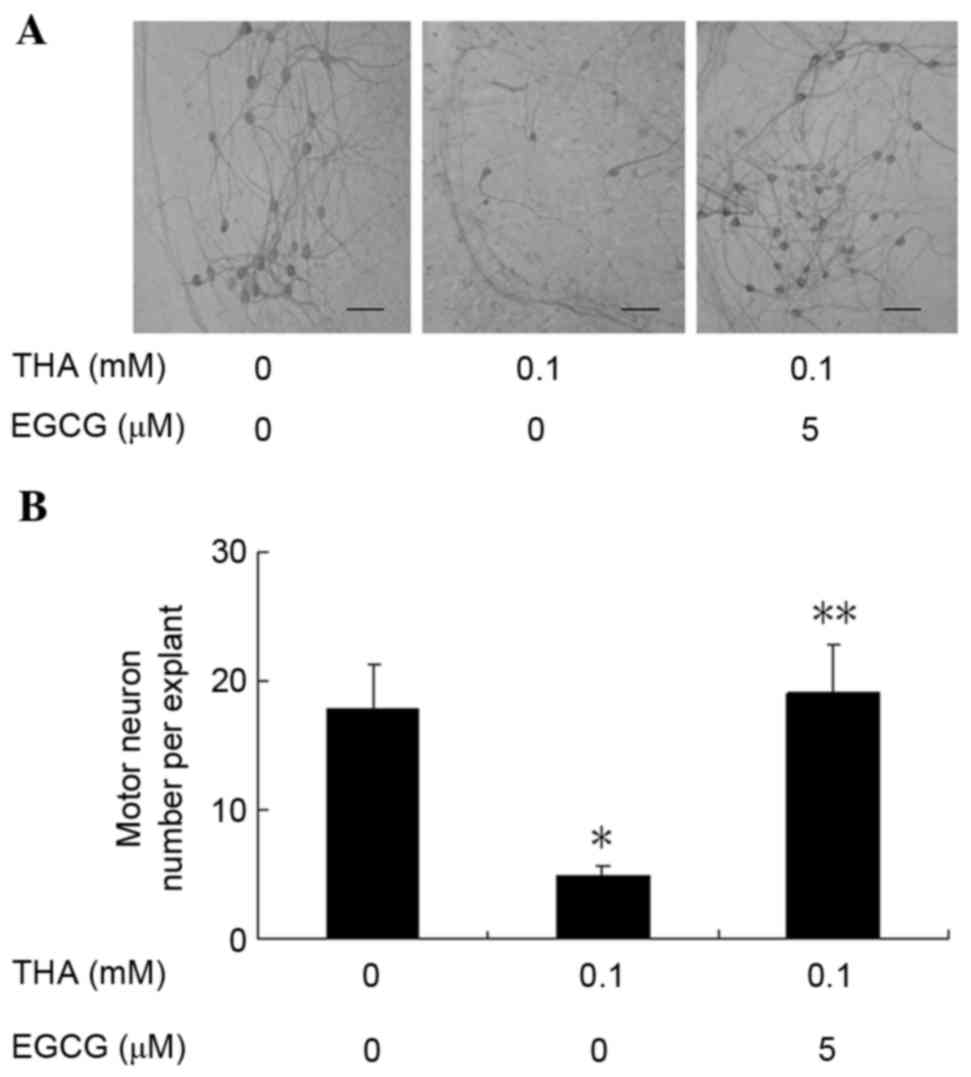
Effect of EGCG on motor neurons
After 1 week in culture, rat lumbar spinal cord
explants were pretreated with EGCG at 5 µM for 48 h and then
treated with a combination of EGCG (at the same concentration as
pre-treatments) and THA at 100 µM for 3 weeks. At the end of the 3
week treatment, the explants were harvested and immunostained with
the anti-neurofilament antibody, SMI-32, for the visualization and
counting of motor neurons. Representative images of ventral horn
neurons of explants treated with vehicle, 100 µM THA, and 5 µM EGCG
plus 100 µM THA are presented in Fig.
1A. All motor neurons in both ventral horns of each explant
were counted (10 explants/group). The control group averaged
17.8±3.42 motor neurons per explant, but only 4.8±0.77 motor
neurons survived per explant following THA treatment (Fig. 1B). This finding is similar to those
reported previously, and is significantly decreased when compared
with the control group (16). The
number of motor neurons (19±3.80) per explant was even higher in
explants treated with 5 µM EGCG plus 100 µM THA when compared with
the control group (Fig. 1B),
demonstrating that EGCG completely blocked THA-induced motor neuron
death.
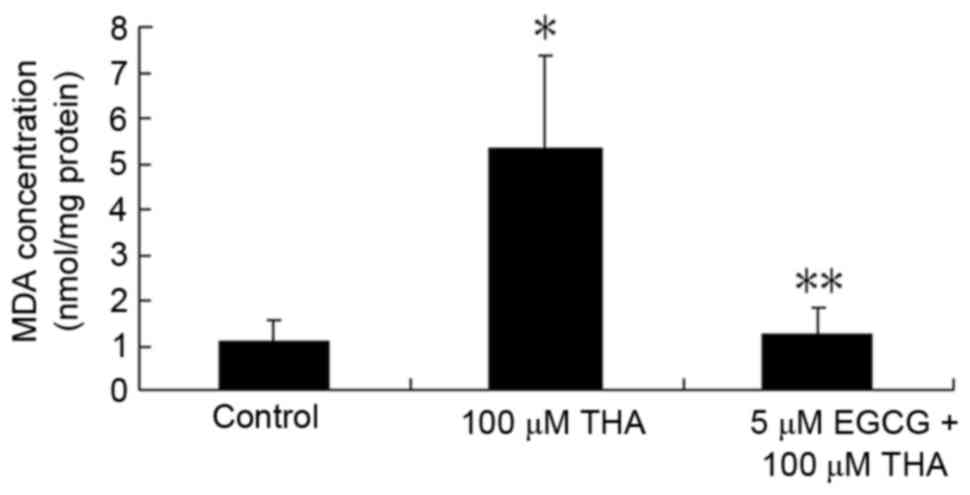
Effect of EGCG on lipid
peroxidation
Levels of lipid peroxides were measured in tissue
using MDA as a marker of lipid peroxidation. At the end of the 3
week treatment, the spinal cord explants were harvested for
measurement of MDA. The concentration of MDA was 1.07±0.45 nmol/mg
protein in the control explants, while THA treatment significantly
increased the levels of tissue MDA (5.35±2.05 nmol/mg protein).
EGCG prevented the effects of THA and decreased the MDA to
1.25±0.54 nmol/mg protein, which was significantly lower than the
THA treated group (Fig. 2).
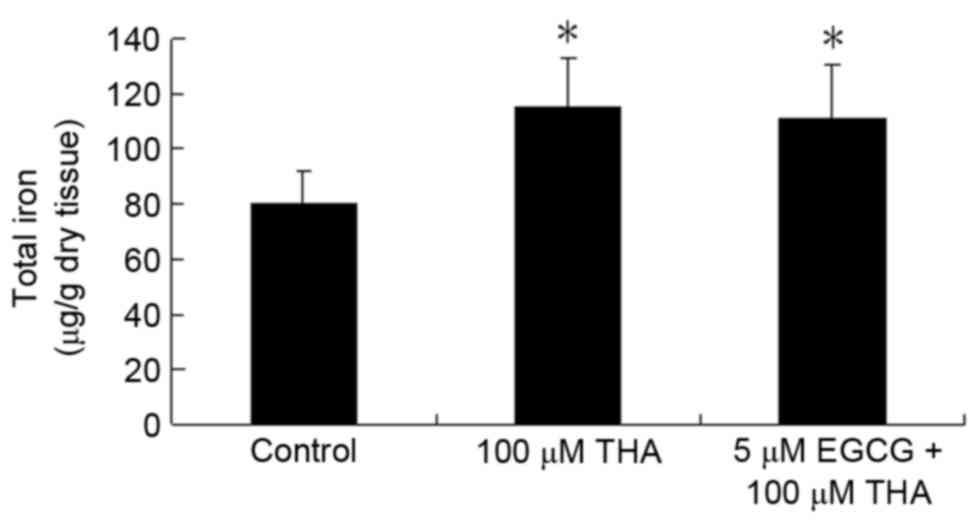
Effect of EGCG on total iron
The total iron content in the explants measured at
the end of 3 weeks of THA treatment was increased significantly
compared with the control group. However, EGCG did not prevent the
iron increase caused by THA. The total iron in the THA-treated
group was 115.27±18.00 µg/g dry tissue, and 111.06±19.35 µg/g dry
tissue in the group treated with EGCG plus THA. There was no
significant difference between these two experimental groups
(Fig. 3; P>0.05).
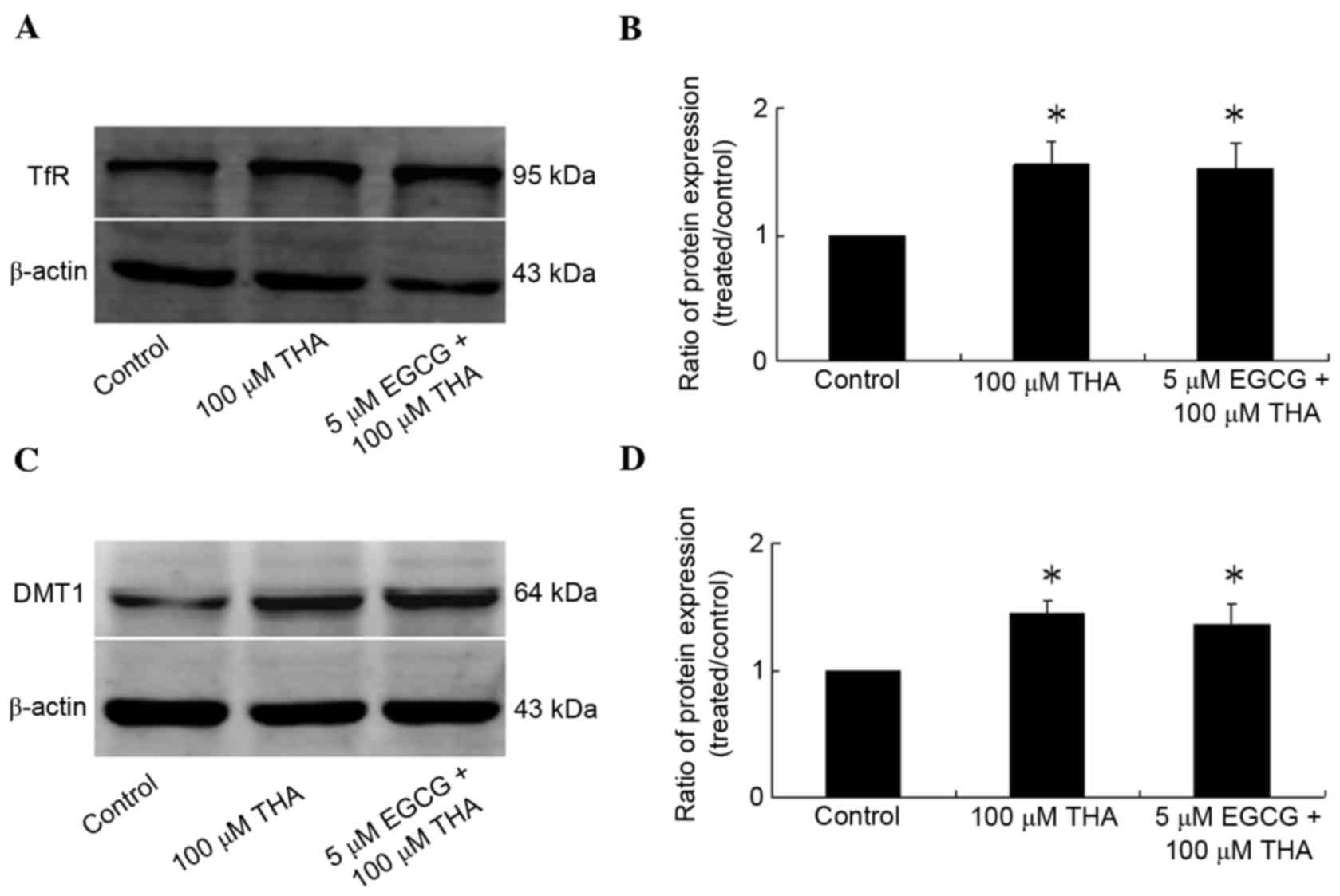
Effect of EGCG on iron metabolism
protein
Rat lumbar spinal cord explants were treated with
THA and EGCG for 3 weeks and harvested for measurement of
expression of iron metabolism proteins. The results demonstrated
that the expression levels of TfR and DMT1 increased significantly
following THA treatment compared with the control group (Fig. 4; TfR, P=0.0316; DMT1, P=0.0186).
TfR and DMT1 are key cellular iron uptake proteins; however there
were no differences in the expression of the TfR and DMT1 between
groups treated with EGCG+THA and those treated with THA only
(Fig. 4; P>0.05), thus EGCG had
no effect on the expression levels of TfR and DMT1 in spinal cord
explants.
Discussion
Iron catalyzes the formation of the highly reactive
hydroxyl radicals from hydrogen peroxide by the Fenton reaction
(H2O2+Fe2+→OH·+OH−+Fe3+)
and, therefore, it can potentiate the toxic effects of reactive
oxidative species (ROS) (17).
Previous studies indicate that oxidative stress is a major
contributory factor leading to chronic motor neuron death in ALS
(18). While iron is essential for
normal physiology, the presence of excessive amounts of iron is
also implicated in numerous pathological processes, including
neurodegenerative disorders (19,20).
THA-induced glutamate excitotoxicity in organotypic
spinal cord cultures is a widely used model of motor neuron
degeneration that has been used for the development of
neuroprotective treatments (21).
Glutamate excitotoxicity is thought to result from intracellular
calcium overload, leading to the generation of ROS (22). The present study demonstrated that
iron levels and the expression of TfR and DMT1 (key cellular iron
uptake proteins) increased significantly in spinal cord tissue
following THA treatment (15).
Therefore, the THA ALS model was used to study the effect of EGCG
on iron metabolism proteins.
In the current study, EGCG (5 µM) was demonstrated
to protect motor neurons in the organotypic culture of rat spinal
cord and decrease lipid peroxidation caused by THA. It appears that
EGCG may have a neuroprotective effect at low concentrations. In
addition, the total iron content and the expression of influx
proteins (TfR and DMT1) increased significantly in spinal cord
following 3 weeks of THA treatment. However, there were no
differences in total iron levels and the expression of influx
proteins (TfR and DMT1) between the groups treated with EGCG+THA
and treated with THA only. It appears that, at the dose used, EGCG
had no effect on the total iron and the expression of influx
proteins in the organotypic spinal cord culture model.
EGCG can protect neurons through many different
mechanisms: Scavenging free radicals, chelating transitional
metals, modulating the expression of cell survival/death genes and
activating phase II drug metabolizing enzymes (23,24).
It is possible that EGCG may chelate free iron (Fe2+) by
removing it from the intracellular iron pool (24–26)
and potentially preventing the formation of highly reactive
hydroxyl radicals. The present study was unable to measure free
iron in the organotypic culture of rat spinal cord due to technical
limitations, however there are plans to follow up on the effect of
EGCG on cellular free iron in future investigations.
In conclusion, EGCG decreases oxidative stress and
protects motor neurons in an organotypic culture of rat spinal
cord, but, at the doses given, EGCG could not decrease the influx
of iron through regulation of iron metabolism proteins. The study
implied that EGCG may not block oxidative damage caused by iron via
chelating iron. Therefore, further studies are required to
investigate the mechanism of EGCG protection of motor neurons.
Acknowledgements
The present study was supported by grants from the
Shandong Provincial Natural Science Foundation of China (grant no.
ZR2010HM041) and the Shandong Provincial Postdoctoral Innovation
Foundation of China (grant no. 201102004).
Glossary
Abbreviations
Abbreviations:
|
EGCG
|
epigallocatechin-3-gallate
|
|
ALS
|
amyotrophic lateral sclerosis
|
|
TfR
|
transferrin receptor
|
|
DMT1
|
divalent metal-ion transporter 1
|
|
THA
|
threo-hydroxyaspartate
|
|
MDA
|
malondialdehyde
|
References
|
1
|
Zarei S, Carr K, Reiley L, Diaz K, Guerra
O, Altamirano PF, Pagani W, Lodin D, Orozco G and Chinea A: A
comprehensive review of amyotrophic lateral sclerosis. Surg Neurol
Int. 6:1712015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jeong SY, Rathore KI, Schulz K, Ponka P,
Arosio P and David S: Dysregulation of iron homeostasis in the CNS
contributes to disease progression in a mouse model of amyotrophic
lateral sclerosis. J Neurosci. 29:610–619. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hadzhieva M, Kirches E, Wilisch-Neumann A,
Pachow D, Wallesch M, Schoenfeld P, Paege I, Vielhaber S, Petri S,
Keilhoff G and Mawrin C: Dysregulation of iron protein expression
in the G93A model of amyotrophic lateral sclerosis. Neuroscience.
230:94–101. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hozumi I, Hasegawa T, Honda A, Ozawa K,
Hayashi Y, Hashimoto K, Yamada M, Koumura A, Sakurai T, Kimura A,
et al: Patterns of levels of biological metals in CSF differ among
neurodegenerative diseases. J Neurol Sci. 303:95–99. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Su XW, Clardy SL, Stephens HE, Simmons Z
and Connor JR: Serum ferritin is elevated in amyotrophic lateral
sclerosis patients. Amyotroph Lateral Scler Frontotemporal Degener.
16:102–107. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Langkammer C, Enzinger C, Quasthoff S,
Grafenauer P, Soellinger M, Fazekas F and Ropele S: Mapping of iron
deposition in conjunction with assessment of nerve fiber tract
integrity in amyotrophic lateral sclerosis. J Magn Reson Imaging.
31:1339–1345. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yu J, Qi F, Wang N, Gao P, Dai S, Lu Y, Su
Q, Du Y and Che F: Increased iron level in motor cortex of
amyotrophic lateral sclerosis patients: An in vivo MR study.
Amyotroph Lateral Scler Frontotemporal Degener. 15:357–361. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kwan JY, Jeong SY, Van Gelderen P, Deng
HX, Quezado MM, Danielian LE, Butman JA, Chen L, Bayat E and
Russell J: Iron accumulation in deep cortical layers accounts for
MRI signal abnormalities in ALS: Correlating 7 tesla MRI and
pathology. PLoS One. 7:e352412012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Reznichenko L, Amit T, Zheng H,
Avramovich-Tirosh Y, Youdim MB, Weinreb O and Mandel S: Reduction
of iron-regulated amyloid precursor protein and beta-amyloid
peptide by (−)-epigallocatechin-3-gallate in cell cultures:
Implications for iron chelation in Alzheimer's disease. J
Neurochem. 97:527–536. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Weinreb O, Amit T and Youdim MB: The
application of proteomics for studying the neurorescue activity of
the polyphenol (−)-epigallocatechin-3-gallate. Arch Biochem
Biophys. 476:152–160. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mandel S, Weinreb O, Amit T and Youdim MB:
Cell signaling pathways in the neuroprotective actions of the green
tea polyphenol (−)-epigallocatechin-3-gallate: Implications for
neurodegenerative diseases. J Neurochem. 88:1555–1569. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Koh SH, Lee SM, Kim HY, Lee KY, Lee YJ,
Kim HT, Kim J, Kim MH, Hwang MS, Song C, et al: The effect of
epigallocatechin gallate on suppressing disease progression of ALS
model mice. Neurosci Lett. 395:103–107. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xu Z, Chen S, Li X, Luo G, Li L and Le W:
Neuroprotective effects of (−)-epigallocatechin-3-gallate in a
transgenic mouse model of amyotrophic lateral sclerosis. Neurochem
Res. 31:1263–1269. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rothstein JD, Jin L, Dykes-Hoberg M and
Kuncl RW: Chronic inhibition of glutamate uptake produces a model
of slow neurotoxicity. Proc Natl Acad Sci USA. 90:6591–6595. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu J, Guo Y, Sun M, Li B, Zhang Y and Li
C: Iron is a potential key mediator of glutamate excitotoxicity in
spinal cord motor neurons. Brain Res. 27:102–107. 2009. View Article : Google Scholar
|
|
16
|
Yu J, Jia Y, Guo Y, Chang G, Duan W, Sun
M, Li B and Li C: Epigallocatechin-3-gallate protects motor neurons
and regulates glutamate level. FEBS Lett. 584:2921–2925. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jomova K, Vondrakova D, Lawson M and Valko
M: Metals, oxidative stress and neurodegenerative disorders. Mol
Cell Biochem. 345:91–104. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Petri S, Korner S and Kiaei M: Nrf2/ARE
signaling pathway: Key mediator in oxidative stress and potential
therapeutic target in ALS. Neurol Res Int.
2012:8780302012.PubMed/NCBI
|
|
19
|
Hadzhieva M, Kirches E and Mawrin C:
Review: Iron metabolism and the role of iron in neurodegenerative
disorders. Neuropathol Appl Neurobiol. 40:240–257. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ward RJ, Dexter DT and Crichton RR:
Neurodegenerative diseases and therapeutic strategies using iron
chelators. J Trace Elem Med Biol. 31:267–273. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Silani V, Braga M, Ciammola A, Cardin V
and Scarlato G: Motor neurones in culture as a model to study ALS.
J Neurol. 247:(Suppl 1). SI28–SI36. 2000. View Article : Google Scholar
|
|
22
|
Carriedo SG, Sensi SL, Yin HZ and Weiss
JH: AMPA exposures induce mitochondrial Ca(2+) overload and ROS
generation in spinal motor neurons in vitro. J Neurosci.
20:240–250. 2000.PubMed/NCBI
|
|
23
|
Mandel S and Youdim MB: Catechin
polyphenols: Neurodegeneration and neuroprotection in
neurodegenerative diseases. Free Radic Biol Med. 37:304–317. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Weinreb O, Amit T, Mandel S and Youdim MB:
Neuroprotective molecular mechanisms of
(−)-epigallocatechin-3-gallate: A reflective outcome of its
antioxidant, iron chelating and neuritogenic properties. Genes
Nutr. 4:283–296. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mandel SA, Avramovich-Tirosh Y,
Reznichenko L, Zheng H, Weinreb O, Amit T and Youdim MB:
Multifunctional activities of green tea catechins in
neuroprotection. Modulation of cell survival genes, iron-dependent
oxidative stress and PKC signaling pathway. Neurosignals. 14:46–60.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mandel SA, Amit T, Kalfon L, Reznichenko
L, Weinreb O and Youdim MB: Cell signaling pathways and iron
chelation in the neurorestorative activity of green tea
polyphenols: Special reference to epigallocatechin gallate (EGCG).
J Alzheimers Dis. 15:211–222. 2008. View Article : Google Scholar : PubMed/NCBI
|