Introduction
Idiopathic normal pressure hydrocephalus (iNPH) is
caused by the accumulation of cerebrospinal fluid (CSF) and it
leads to gait disturbance, urinary incontinence, and dementia
(1). Bradley et al
suggested that benign external hydrocephalus in infancy could
exacerbate in late adulthood and might lead to iNPH (1,2).
Other groups have reported the existence of genetic predisposition
to the disease (3,4). However, the precise mechanism of
onset of iNPH is still unknown. However, unlike other dementias,
iNPH is treatable by shunt operation, which relieves the
accumulated CSF (1).
In Japan, approximately 5% of dementia patients are
diagnosed with iNPH. However, some symptoms of iNPH are similar to
those of Alzheimer's disease (AD) or other dementias. Therefore, it
is often difficult to distinguish iNPH from other dementias.
Identifying biomarkers for iNPH is important to diagnose latent
iNPH patients among dementia patients.
The objective of this study was to identify
potential biomarkers for iNPH. To this purpose, we conducted a
proteomic analysis of CSF of iNPH and AD patients using
high-performance liquid chromatography (HPLC) coupled with
electrospray ionization quadrupole time-of-flight mass spectrometry
(LC-TOF-MS). We identified protein tyrosine phosphatase receptor
type Q (PTPRQ) as a potential biomarker for iNPH.
PTPRQ is known as a gene causing hearing
loss, deafness autosomal recessive 84A (DFNB84A) (5,6).
Goodyear et al reported that PTPRQ null mice exhibit
loss and fusion of cochlear stereocilia, resulting in hearing loss
(7). Nayak et al indicated
that the extracellular region of PTPRQ in the vestibular end organ
is modified with dermatan sulfate. Depending on the location of the
dermatan sulfate on PTPRQ, neighboring stereocilia adhere to or
repel each other, thus maintaining a proper distance between the
cilia (8).
In connection with these reports, patients with
hydrocephalus-associated hearing loss are reported, and there are
some reports on hearing loss related to shunt surgery (9–11).
These reports suggest the relevance among iNPH, auditory capacity
and PTPRQ. Therefore, in this study, we attempted to investigate
the characteristics and concentration of PTPRQ in the CSF and the
relation to auditory capacity.
Materials and methods
Subjects
Subjects with different diagnoses including healthy
control, AD, iNPH and shunt non-responder (SNR) patients were
examined. Subject characteristics are summarized in Table I. iNPH diagnosis was made according
to the ‘Guidelines for management of idiopathic normal pressure
hydrocephalus: Second edition’ (12). Shunt surgeries were done for
probable iNPH patients and determined to be effective in case of 1
point or more improvement on the modified Rankin scale, or 2 points
or more on the Japanese iNPH grading scale (13). AD diagnosis was made according to
the published ‘Research criteria for the diagnosis of Alzheimer's
disease: Revising the NINCDS-ADRADA criteria’ (14).
 | Table I.Subject characteristics. |
Table I.
Subject characteristics.
|
| LC-TOF-MS | ELISAa |
|---|
|
|
|
|
|---|
|
| AD | iNPHb | Control | AD | iNPHc | SNR |
|---|
| Subject no. | 7 | 8 | 12 | 48 | 36 | 8 |
| Mean age (SD) | 71.3 (7.7) | 75.3 (4.5) | 71.2 (9.6) | 75.5 (8.0) | 79.6 (5.1) | 78.9 (3.0) |
| Age range | 62–84 | 70–84 | 52–85 | 54–87 | 67–88 | 76–84 |
| Male/female | 4/3 | 5/3 | 3/9 | 20/28 | 16/20 | 3/5 |
Bioresources were largely obtained from the BioBanks
of the National Center for Geriatrics and Gerontology (NCGG, Aichi,
Japan) and the National Center of Neurology and Psychiatry (NCNP,
Tokyo, Japan) and partly donated by the Department of Biological
Regulation, School of Medicine, Tottori University (Tottori,
Japan). All samples were obtained with written informed consent
from the patients before sampling between 2011 and 2015. This study
was reviewed and approved by the Ethics Committees of the Institute
and BioBanks.
Collection of CSF
CSF was collected by lumber puncture and centrifuged
for 10 min at 800 × g at 4°C. The supernatant was aliquoted into
low protein-binding tubes and immediately frozen in liquid nitrogen
and stored at −80°C until use.
Proteomic analysis by mass
spectrometry
Four hundred microliters of CSF was concentrated by
centrifugation using Ultra-4 Centrifugal Filters Ultracel-3K (Merck
Millipore Japan, Tokyo, Japan) for 1 h at 4,000 × g at 4°C.
Approximately 100 µl of concentrated CSF was filtered through an
Ultrafree-MC GV 0.22-µm spin column (Merck Millipore Japan) for 2
min at 16,000 × g at 4°C to remove debris. CSF proteins were
separated into high- and low-abundance proteins by HPLC using Human
14 Multiple Affinity Removal System Column (Agilent Technologies,
Inc., Tokyo, Japan) according to the manufacturer's protocol. The
low-abundance protein fraction was concentrated again and stored at
−80°C until analysis.
Proteomic analysis was conducted with LC-TOF-MS
(QTOF Ultima; Nihon Waters K.K., Tokyo, Japan) as previously
described (15–17). Briefly, samples were
trypsin-digested and MS peaks were detected with LC-TOF-MS,
followed by normalization and quantification with the 2DICAL
software (Mitsui Knowledge Industry Co., Ltd., Tokyo, Japan)
(18).
Enzyme-linked immunosorbent assay
(ELISA)
An ELISA kit for PTPRQ quantification was purchased
from Cloud-Clone Corp. (Houston, TX, USA) and used according to the
manufacturer's protocol. One hundred microliters of CSF was added
per well. After quantification, the outliers more than median ± 3 ×
SD were excluded from analysis.
Western blotting
Human total protein lysates of kidneys, brain,
diencephalon (DiE), and cerebral meninges (CM) were purchased from
Cosmo Bio Co., Ltd. (Tokyo, Japan). Equal amounts of samples were
mixed with Laemmli sample buffer (final volume containing 5%
β-mercaptoethanol), boiled for 3 min, and separated on a 7.5%
Mini-Protean TGX precast gel (Bio-Rad Laboratories, Tokyo, Japan).
The separated proteins were transferred to a PVDF membrane using a
Trans-Blot Turbo Transfer system (Bio-Rad Laboratories). The
membrane was incubated with 5% skim milk in Tris-buffered saline
with Tween-20 (TBST) at room temperature for 1 h and then with
rabbit polyclonal anti-PTPRQ antibody recognizing the extracellular
domain (cat. no. PAD603Hu01, immunized with amino acids 36 to 294,
1:400 dilution; Cloud-Clone Corp.) or the intracellular domain
(cat. no. sc-368569, immunized with amino acids 2208 to 2299,
1:1,000 dilution; Santa Cruz Biotechnology, Inc., Dallas, TX, USA)
of PTPRQ in Can Get Signal 1 enhancer solution (Toyobo Co., Ltd.,
Osaka, Japan) at 4°C, overnight. After washing thrice with TBST,
the membrane was incubated with HRP-linked anti-rabbit IgG (cat.
no. 7074; Cell Signaling Technology Japan, K. K., Tokyo, Japan)
diluted to 1:2,000 in Can Get Signal 2 (Toyobo Co., Ltd.) and
incubated at room temperature for 1 h. After four washes with TBST,
signal was detected with ImmunoStar LD (Wako Pure Chemical
Industries, Ltd., Osaka, Japan) and an ImageQuant LAS-4000 imager
(Fujitsu Ltd., Tokyo, Japan). The images were processed using
MultiGauge software v3.2 (FujiFilm Corp., Tokyo, Japan). To improve
visibility, we adjusted the exposure, contrast, and angle of the
images.
Immunohistochemistry of the mouse
brain tissue
The brain tissues were dissected from a male C57BL/6
mouse (8-week-old), fixed in 10% neutral formalin for 24 h,
embedded in paraffin, and cut into 2-µm sections. Sections were
dewaxed using xylene, rehydrated in a graded series of alcohol to
water, and subjected to antigen retrieval using retrieval solution,
pH 6.0 (Nichirei Biosciences, Inc., Tokyo, Japan) in a pressure
cooker for 10 min. Endogenous peroxidase was blocked by incubation
of the brain sections with 3% hydrogen peroxide for 10 min.
Sections were subsequently blocked with 3% bovine serum albumin
(Wako Pure Chemical Industries, Ltd.) for 30 min to prevent
non-specific antibody binding.
The sections were incubated with rabbit polyclonal
IgG anti-PTPRQ (immunized with amino acids 2208 to 2299, 1:50
dilution; Santa Cruz Biotechnology, Inc.) for 60 min, followed by
incubation with the secondary anti-rabbit IgG antibody conjugated
to horseradish peroxidase (HRP) (Nichirei Biosciences, Inc.) for 60
min. Following each treatment, the slides were washed using
phosphate-buffered saline (PBS; 3×5 min washes). Finally, they were
stained using DAB. Sections were counterstained using Mayer's
hematoxylin, dehydrated, cleared, and mounted.
Stained sections were analyzed under the microscope
BX-53 (Olympus Corporation, Tokyo, Japan), and representative
photographs were acquired using a digital camera DP21 (Olympus
Corporation).
Hearing evaluation
The correlation between the amount of PTPRQ in the
CSF and auditory capacity was investigated in 26 patients with iNPH
who had undergone pure tone audiometry before the shunt operation.
The patients included 8 males and 18 females (mean age, 79.9±3.6;
range, 72 to 88; median PTPRQ concentration, 0.70±0.5; range, 0.24
to 2.0 ng/ml). The average threshold of right and left ear at
frequencies of 125, 500, 1,000, 2,000, 4,000, and 8,000 Hz was
determined. Samples from all these patients were included in the
ELISA.
Statistical analyses
For proteomic analysis and ELISA, data were analyzed
using Welch's t-test in StatFlex version 6 (Artech Co., Ltd.,
Osaka, Japan). For hearing evaluation, the correlation between the
amount of PTPRQ in the CSF and the pure tone audiometry was
analyzed using a general linear model in the statistical analysis
system (SAS) version 9.3 (SAS Institute Japan, Tokyo, Japan). A
value of P<0.05 was considered to indicate a statistically
significant difference.
Results
The concentration of PTPRQ in CSF
We analyzed CSF samples from 7 AD and 8 iNPH
patients (Table I) with LC-TOF-MS.
We found 74 proteins showing a significantly different ion strength
between AD and iNPH samples (7 were higher and 67 were lower in
iNPH than in AD, data not shown). Among these, PTPRQ showed a
P-value <0.01 (Fig. 1A).
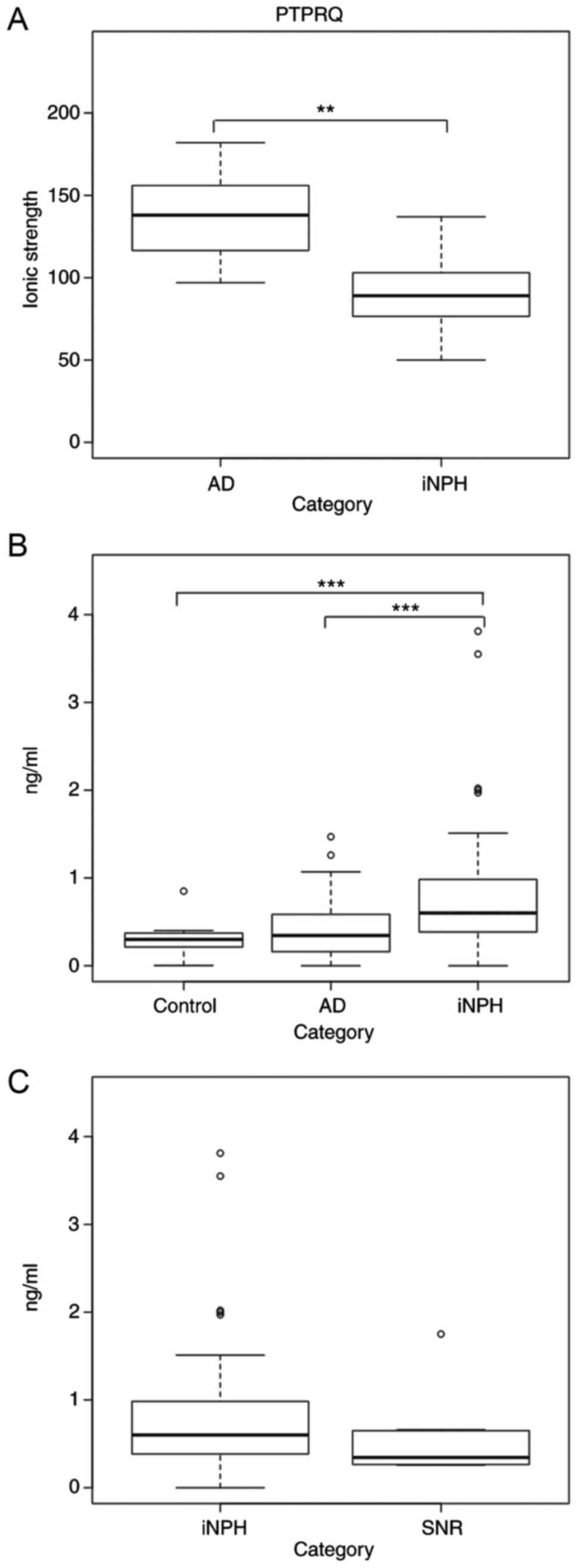 | Figure 1.PTPRQ concentration in CSF. (A) The
ionic strength of PTPRQ detected with LC-TOF-MS was higher in AD
than in iNPH patients. P=0.007 (Welch's t-test). (B) PTPRQ in CSF
as tested by ELISA. Welch's t-test was used to compare groups. The
P-value of control vs. iNPH was 0.0007 and that of AD vs. iNPH was
0.003. Median values of control, AD, and iNPH were 0.30, 0.35, and
0.68 ng/ml, respectively. (C) PTPRQ in CSF from iNPH and SNR are
indicated. Welch's t-test was performed and the P-value was 0.18.
The middle line in the box represents the median, while the upper
edge and lower edge represent the 1st quartile (1Q) and the 3rd
quartile (3Q), respectively. Short whiskers indicate the minimum
and maximum point of our data. The circles indicate outlier value.
The potential outliers lie outside the range of 1Q-(IQRx1.5) to
3Q+(IQRx1.5). IQR stands for interquartile range. **P<0.01,
***P<0.005. PTPRQ, protein tyrosine phosphatase receptor type Q;
AD, Alzheimer's disease; iNPH, idiopathic normal pressure
hydrocephalus; SNR, shunt non-responder; CSF, cerebrospinal fluid;
LC-TOF-MS, liquid chromatography coupled with electrospray
ionization quadrupole time-of-flight mass spectrometry; ELISA,
enzyme-linked immunosorbent assay. |
To validate the concentration of PTPRQ, we conducted
an ELISA using the same and additional samples of AD and iNPH
patients. Additionally, we tested 12 healthy control and 8 SNR
samples to evaluate the normal range of the PTPRQ concentration in
the CSF and whether PTPRQ could distinguish SNRs among iNPH. The
results confirmed that the PTPRQ level was significantly higher in
iNPH than in healthy control and AD subjects (Fig. 1B). Furthermore, the median PTPRQ
concentration in the CSF of SNRs was approximately 2 times lower
than that in iNPH, albeit not significantly (Fig. 1C, P=0.18).
Qualitative analysis of PTPRQ by
western blotting
In general, the ionic strength as determined by mass
spectrometry is correlated with the relative abundance of the
protein in the analyte. However, the abundances of PTPRQ estimated
with mass spectrometry and ELISA were contradictory (Fig. 1A and B). In mass spectrometry,
proteins are identified by quantifying the targets' molecular
weights (19). Therefore, proteins
having complex post-translational modifications occasionally fail
to be detected. Thus, we presumed that PTPRQ in iNPH would have
post-translational modifications different from that in AD. To
verify this possibility, we performed western blotting with CSF
samples from each of the diagnostic categories.
No obvious difference in PTPRQ band size was
observed between iNPH patients and others (Fig. 2). This result indicated that PTPRQ
in iNPH does not have a large difference in molecular weight as
compared to the other diagnostic categories, or at least it was not
detectable by western blotting. However, minor post-translational
modifications could not be excluded.
Validation of PTPRQ expression in the
brain tissues
Next, to confirm the expression of PTPRQ in the
brain, we performed western blotting with total protein lysates
from normal human tissues. Kidney lysate was used as a positive
control (20). Further, we
selected DiE and CM as well as whole-brain tissue lysates, which
are all in contact with CSF, for analysis.
In kidney lysate, three main bands were observed
(Fig. 3A and B), likely
corresponding to the splicing variants depicted in Fig. 3C. Band 2 (~250 kDa) corresponds to
the canonical isoform of PTPRQ (Uniprot ID Q9UMZ2), bands 1
(>250 kDa) and 3 (~180 kDa) are likely isoforms with different
numbers of the fibronectin 3 domain (FNIII) (5,20).
Band 4 (<150 kDa) was not observed in kidney lysate, but showed
signal in whole-brain, DiE and CM. In these tissue lysates, all
bands were detected at almost the same position by antibodies both
detecting the extracellular (Fig.
3A) and the intracellular domain of PTPRQ (Fig. 3B). Band densities were stronger in
CM than in brain and DiE samples, suggesting that PTPRQ is
preferentially expressed in the CM. In CSF, the estimated molecular
weight of the PTPRQ was approximately 170 kDa when antibody for the
extracellular domain of PTPRQ was used (Figs. 2 and 3A), whereas this band disappeared when
antibody for the intracellular domain was used (Fig. 3B). These results suggested that
PTPRQ in the CSF consists of part of the extracellular domain and
is derived from the brain tissues.
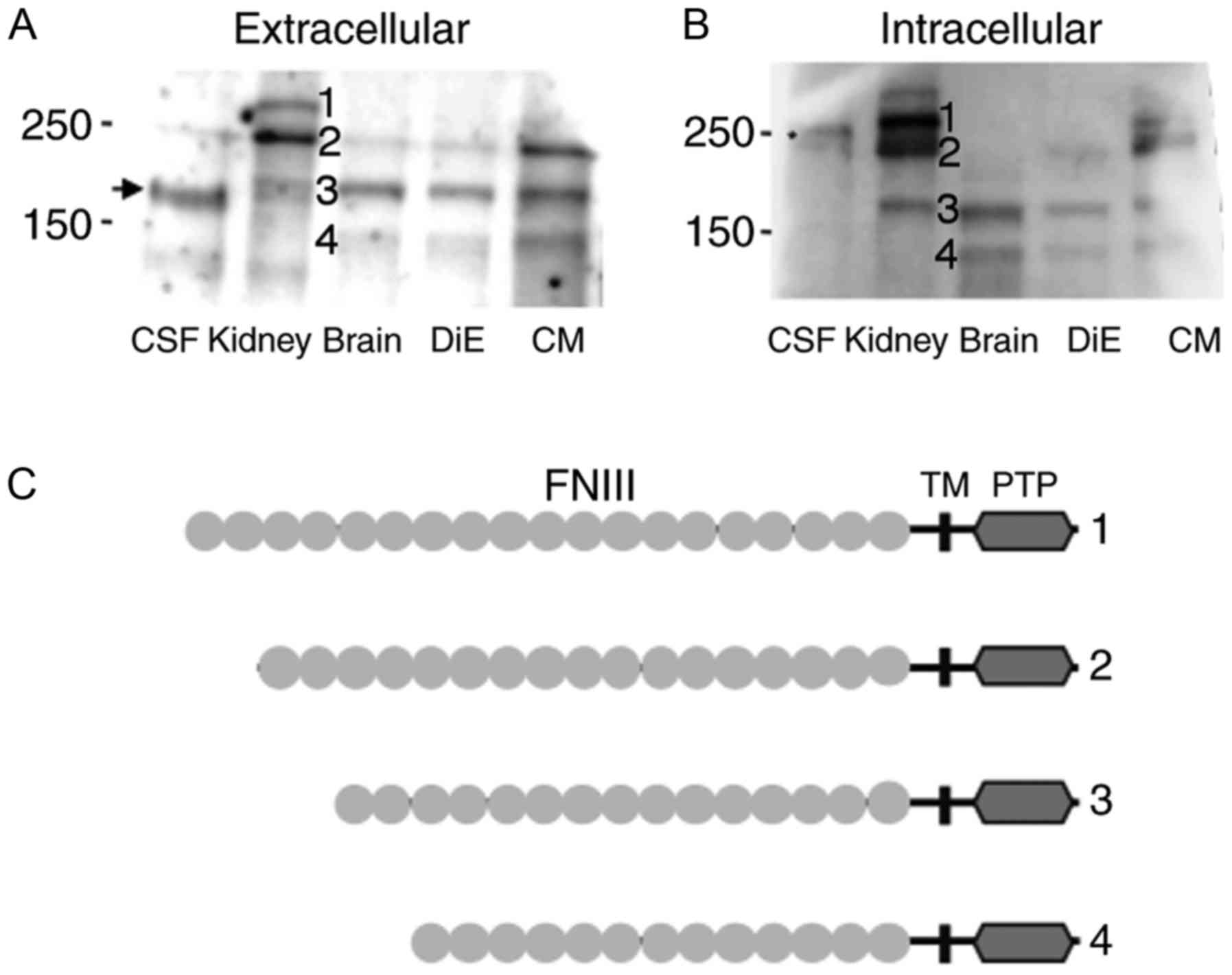 | Figure 3.Western blotting of PTPRQ in the CSF
and indicated tissue lysates. (A and B) Five microliters of CSF
from iNPH patients and 25 µg of total protein lysates from human
kidney (positive control), brain, DiE, and CM tissues from healthy
adults were analyzed. Western blots using antibodies recognizing
(A) the extracellular and (B) the intracellular domain of PTPRQ are
shown. (C) Schematic representations of the known (1,2,3) and
predicted (4) isoforms of PTPRQ.
Numbers correspond to the band numbers indicated on the western
blot. PTPRQ, protein tyrosine phosphatase receptor type Q; CSF,
cerebrospinal fluid; iNPH, idiopathic normal pressure
hydrocephalus; DiE, diencephalon; CM, cerebral membrane; FNIII,
fibronectin 3 domain; TM, transmembrane; PTP, protein tyrosine
phosphatase. The asterisk indicates a non-specific band near 250
kDa. |
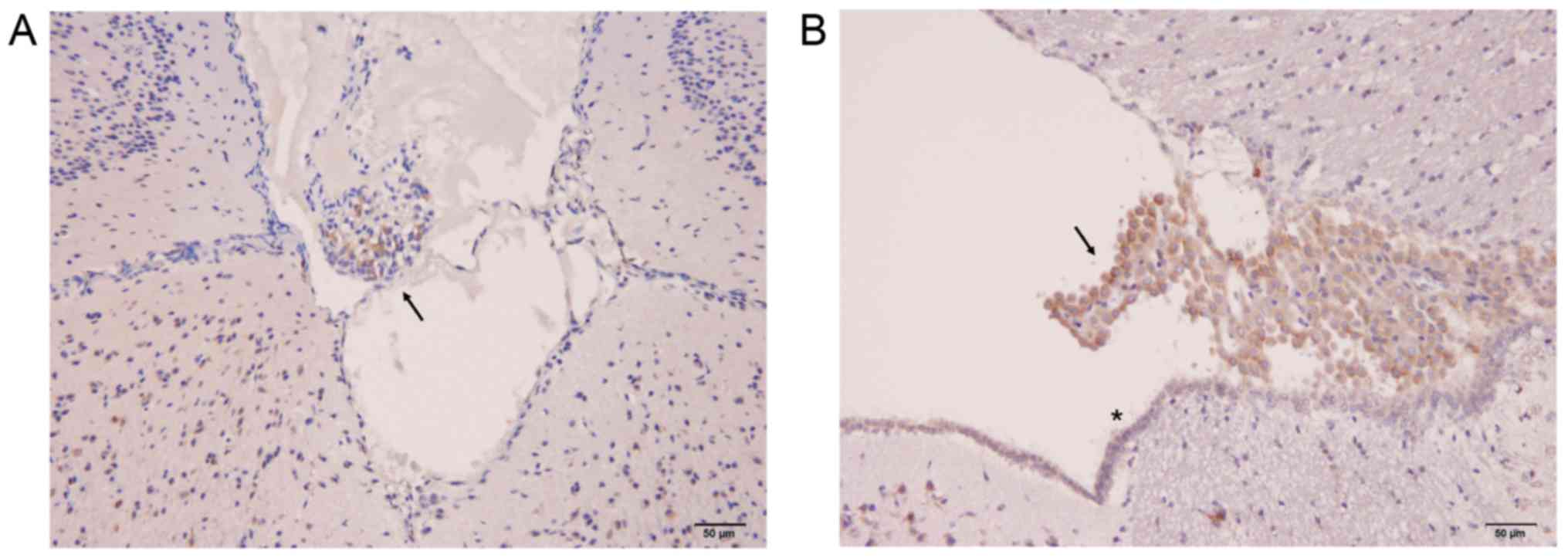
Further, to examine the localization of PTPRQ in the
brain, immunohistochemistry was performed using anti-PTPRQ
antibody. The CM with arachnoid villi and DiE with third ventricle,
which include choroid plexus, were chosen from the mouse. As a
result, arachnoid villi and choroid plexus were found to be stained
with anti-PTPRQ antibody (Fig. 4,
arrows). Furthermore, the surface area of the third ventricle,
where the ependymal cells along with cilia structure is observed
and directly contacts the CSF, was immunostained with anti-PTPRQ
antibody (Fig. 4B, asterisk).
These results suggest that PTPRQ in the CSF comes in contact with
the CSF from these structures.
Evaluation of hearing capability
Next, we investigated the potential correlation
between the amount of PTPRQ in the CSF and auditory capacity by
using a general linear model. We found no correlation between the
amount of PTPRQ and the average hearing threshold at each frequency
(Table II).
| Table II.Pure tone audiometry results of
patients. |
Table II.
Pure tone audiometry results of
patients.
| Frequency (Hz) | 125 | 250 | 500 | 1,000 | 2,000 | 4,000 | 8,000 |
|---|
| Mean ±
SDa | 36.9±14.5 | 41.3±17.7 | 41.9±16.0 | 41.8±15.6 | 59.0±16.6 | 57.4±19.4 | 71.0±19.8 |
| Correlation
factor | −0.158 | −0.035 | 0.036 | 0.101 | −0.008 | −0.006 | −0.027 |
| P-value | 0.441 | 0.865 | 0.863 | 0.623 | 0.969 | 0.975 | 0.897 |
Discussion
Here, we report PTPRQ as a novel potential biomarker
for iNPH. In the CSF of patients with iNPH, the PTPRQ concentration
was significantly higher than in healthy controls and patients with
AD. In addition, the median PTPRQ concentration in the CSF of SNRs
was two times lower than that of patients with iNPH, although the
difference was not significant.
As the abundances of PTPRQ estimated with mass
spectrometry and ELISA were contradictory (Fig. 1A and B), we presumed PTPRQ in iNPH
would have differential post-translational modifications compared
to AD and performed western blotting. We found no obvious
difference in PTPRQ size between iNPH patients and others (Fig. 2). However, there is a report
indicating that transferrin in CSF from iNPH patients have a unique
N-glycan modification (21),
suggesting that PTPRQ in CSF from iNPH patients also possibly have
unique modification. Unfortunately, PTPRQ in CSF is not abundant
compared to transferrin, it makes difficult to analyze PTPRQ
modifications at present. Further analyses with more accurate and
high sensitive methods are needed.
Another western blotting indicated that the PTPRQ in
the CSF consists of part of the extracellular domain of PTPRQ
expressed in brain tissues. One intense band, which was detected
only in the brain tissues (Fig. 3,
band 4), did not correspond to any previously reported isoform of
PTPRQ. However, PTPRQ isoforms have been studied only in kidney,
retina, and testes (20). Band 4
may be an isoform unique to the brain tissue; we hypothesize it to
be another FNIII-number variant. Additional immunohistochemistry of
the mouse CM and DiE using anti-PTPRQ antibody revealed that PTPRQ
is expressed at the site of CSF dynamics, arachnoid villi and
choroid plexus, where the CSF is resorbed and produced,
respectively (22). Furthermore,
the surface area of the third ventricle, where the ependymal cells
possessing the cilia structure, was immunostained with anti-PTPRQ
antibody; these cilia structures are considered to have an
important role in the CSF dynamics (23–25).
The physiological meaning of these findings remains
unclear, though we suspect that PTPRQ might play a role in
maintaining cilia structure in the brain tissues that are in
contact with the CSF, the same as in the ear organ (7,8). If
so, the increase of PTPRQ fragments in the CSF might reflect a
disruption of the ciliary structure in brain. Actually, some
reports have indicated that genetic mutations in ciliary proteins
produce congenital hydrocephalus (24,26–29).
Thus, a disruption of the ciliary structure in brain possibly be
one of the cause of iNPH. In the case of SNRs, the relatively low
concentration of PTPRQ in the CSF might indicate progressed ciliary
deficit, resulting in the exhaustion of PTPRQ protein.
Alternatively, the disease in these patients might have another
pathogenesis mechanism, without the increase of PTPRQ in the CSF.
To evaluate these hypotheses, brain autopsies from iNPH patients
are needed, though it is difficult to obtain at present.
We hypothesized PTPRQ in cochlear stereocillia would
also be disrupted and cause hearing loss in iNPH patients. Thus, we
expected a correlation between the auditory capacity and the amount
of PTPRQ in the CSF of iNPH. However, no significant correlation
was observed (Table II). It is
speculated that PTPRQ disruption might be a locally restricted
event in the brain of iNPH patients and hydrocephalus-associated
hearing loss would be just a result of the pressure difference
between CSF and inner ear fluids (9,10,30).
Although, our analysis was relatively small, further validation in
lager sample size is required.
Usually, the diagnosis of iNPH is performed by CSF
tap test, which removes several 10-ml aliquots of the CSF through a
lumbar puncture. Therefore, the concentration of PTPRQ in the CSF
of almost all the possible cases of iNPH could be readily assessed.
We are expecting the accumulation of enough samples and the
large-scale analysis would reveal the usefulness of PTPRQ as a
marker for iNPH.
Acknowledgements
The present study was supported by the Program for
Promotion of Fundamental Studies in Health Sciences of the National
Institute of Biomedical Innovation of Japan (10–44) and The
Research Funding for Longevity Sciences (26–20).
We thank Dr Nobuyoshi Shimoda for helpful discussions. We wish to
acknowledge Dr Takeshi Tomonaga for helpful advices on manuscript.
We thank Editage (www.editage.jp) for English language editing.
Glossary
Abbreviations
Abbreviations:
|
AD
|
Alzheimer's disease
|
|
CM
|
cerebral meninges
|
|
CSF
|
cerebrospinal fluid
|
|
DFNB84A
|
deafness autosomal recessive 84A
|
|
DiE
|
diencephalon
|
|
FNIII
|
fibronectin 3 domain
|
|
iNPH
|
idiopathic normal pressure
hydrocephalus
|
|
PTP
|
protein tyrosine phosphatase
|
|
PTPRQ
|
protein tyrosine phosphatase receptor
type Q
|
|
SNR
|
shunt non-responder
|
|
TM
|
transmembrane
|
References
|
1
|
Adams RD, Fisher CM, Hakim S, Ojemann RG
and Sweet WH: Symptomatic occult hydrocephalus with normal
cerebrospinal-fluid pressure. New Engl J Med. 273:117–126. 1965.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bradley WG Jr, Bahl G and Alksne JF:
Idiopathic normal pressure hydrocephalus may be a ‘two hit’
disease: Benign external hydrocephalus in infancy followed by deep
white matter ischemia in late adulthood. J Magn Reson Imaging.
24:747–755. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cusimano MD, Rewilak D, Stuss DT,
Barrera-Martinez JC, Salehi F and Freedman M: Normal-pressure
hydrocephalus: Is there a genetic predisposition? Can J Neurol Sci.
38:274–281. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Takahashi Y, Kawanami T, Nagasawa H, Iseki
C, Hanyu H and Kato T: Familial normal pressure hydrocephalus (NPH)
with an autosomal-dominant inheritance: A novel subgroup of NPH. J
Neurol Sci. 308:149–151. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schraders M, Oostrik J, Huygen PL, Strom
TM, van Wijk E, Kunst HP, Hoefsloot LH, Cremers CW, Admiraal RJ and
Kremer H: Mutations in PTPRQ are a cause of autosomal-recessive
nonsyndromic hearing impairment DFNB84 and associated with
vestibular dysfunction. Am J Hum Genet. 86:604–610. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shahin H, Rahil M, Abu Rayan A, Avraham
KB, King MC, Kanaan M and Walsh T: Nonsense mutation of the
stereociliar membrane protein gene PTPRQ in human hearing loss
DFNB84. J Med Genet. 47:643–645. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Goodyear RJ, Jones SM, Sharifi L, Forge A
and Richardson GP: Hair-bundle defects and loss of function in the
vestibular end organs of mice lacking the receptor-like inositol
lipid phosphatase, PTPRQ. J Neurosci. 32:2762–2772. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nayak G, Goodyear RJ, Legan PK, Noda M and
Richardson GP: Evidence for multiple, developmentally regulated
isoforms of Ptprq on hair cells of the inner ear. Dev Neurobiol.
71:129–141. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dixon JF and Jones RO:
Hydrocephalus-associated hearing loss and resolution after
ventriculostomy. Otolaryngol Head Neck Surg. 146:1037–1039. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sammons VJ, Jacobson E and Lawson J:
Resolution of hydrocephalus-associated sensorineural hearing loss
after insertion of ventriculoperitoneal shunt. J Neurosurg Pediatr.
4:394–396. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
van Veelen-Vincent ML, Delwel EJ, Teeuw R,
Kurt E, de Jong DA, Brocaar MP, Pauw BK, Avezaat CJ and van Zanten
BG: Analysis of hearing loss after shunt placement in patients with
normal-pressure hydrocephalus. J Neurosurg. 95:432–434. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mori E, Ishikawa M, Kato T, Kazui H,
Miyake H, Miyajima M, Nakajima M, Hashimoto M, Kuriyama N, Tokuda
T, et al: Guidelines for management of idiopathic normal pressure
hydrocephalus: Second edition. Neurol Med Chir (Tokyo). 52:775–809.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kubo Y, Kazui H, Yoshida T, Kito Y, Kimura
N, Tokunaga H, Ogino A, Miyake H, Ishikawa M and Takeda M:
Validation of grading scale for evaluating symptoms of idiopathic
normal-pressure hydrocephalus. Dement Geriatr Cogn Disord.
25:37–45. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dubois B, Feldman HH, Jacova C, Dekosky
ST, Barberger-Gateau P, Cummings J, Delacourte A, Galasko D,
Gauthier S, Jicha G, et al: Research criteria for the diagnosis of
Alzheimer's disease: Revising the NINCDS-ADRDA criteria. Lancet
Neurol. 6:734–746. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Matsubara J, Ono M, Negishi A, Ueno H,
Okusaka T, Furuse J, Furuta K, Sugiyama E, Saito Y, Kaniwa N, et
al: Identification of a predictive biomarker for hematologic
toxicities of gemcitabine. J Clin Oncol. 27:2261–2268. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ono M, Matsubara J, Honda K, Sakuma T,
Hashiguchi T, Nose H, Nakamori S, Okusaka T, Kosuge T, Sata N, et
al: Prolyl 4-hydroxylation of alpha-fibrinogen: A novel protein
modification revealed by plasma proteomics. J Biol Chem.
284:29041–29049. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Takakura M, Yokomizo A, Tanaka Y,
Kobayashi M, Jung G, Banno M, Sakuma T, Imada K, Oda Y, Kamita M,
et al: Carbonic anhydrase I as a new plasma biomarker for prostate
cancer. ISRN Oncol. 2012:7681902012.PubMed/NCBI
|
|
18
|
Ono M, Shitashige M, Honda K, Isobe T,
Kuwabara H, Matsuzuki H, Hirohashi S and Yamada T: Label-free
quantitative proteomics using large peptide data sets generated by
nanoflow liquid chromatography and mass spectrometry. Mol Cell
Proteomics. 5:1338–1347. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mann M, Højrup P and Roepstorff P: Use of
mass spectrometric molecular weight information to identify
proteins in sequence databases. Biol Mass Spectrom. 22:338–345.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Seifert RA, Coats SA, Oganesian A, Wright
MB, Dishmon M, Booth CJ, Johnson RJ, Alpers CE and Bowen-Pope DF:
PTPRQ is a novel phosphatidylinositol phosphatase that can be
expressed as a cytoplasmic protein or as a subcellularly localized
receptor-like protein. Exp Cell Res. 287:374–386. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Futakawa S, Nara K, Miyajima M, Kuno A,
Ito H, Kaji H, Shirotani K, Honda T, Tohyama Y, Hoshi K, et al: A
unique N-glycan on human transferrin in CSF: A possible biomarker
for iNPH. Neurobiol Aging. 33:1807–1815. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
McComb JG: Recent research into the nature
of cerebrospinal fluid formation and absorption. J Neurosurg.
59:369–383. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Narita K and Takeda S: Cilia in the
choroid plexus: Their roles in hydrocephalus and beyond. Front Cell
Neurosci. 9:392015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Banizs B, Pike MM, Millican CL, Ferguson
WB, Komlosi P, Sheetz J, Bell PD, Schwiebert EM and Yoder BK:
Dysfunctional cilia lead to altered ependyma and choroid plexus
function, and result in the formation of hydrocephalus.
Development. 132:5329–5339. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Siyahhan B, Knobloch V, de Zélicourt D,
Asgari M, Daners M Schmid, Poulikakos D and Kurtcuoglu V: Flow
induced by ependymal cilia dominates near-wall cerebrospinal fluid
dynamics in the lateral ventricles. J R Soc Interface.
11:201311892014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ibañez-Tallon I, Pagenstecher A, Fliegauf
M, Olbrich H, Kispert A, Ketelsen UP, North A, Heintz N and Omran
H: Dysfunction of axonemal dynein heavy chain Mdnah5 inhibits
ependymal flow and reveals a novel mechanism for hydrocephalus
formation. Hum Mol Genet. 13:2133–2141. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lechtreck KF, Delmotte P, Robinson ML,
Sanderson MJ and Witman GB: Mutations in Hydin impair ciliary
motility in mice. J Cell Biol. 180:633–643. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee L: Riding the wave of ependymal cilia:
Genetic susceptibility to hydrocephalus in primary ciliary
dyskinesia. J Neurosci Res. 91:1117–1132. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tissir F, Qu Y, Montcouquiol M, Zhou L,
Komatsu K, Shi D, Fujimori T, Labeau J, Tyteca D, Courtoy P, et al:
Lack of cadherins Celsr2 and Celsr3 impairs ependymal ciliogenesis,
leading to fatal hydrocephalus. Nat Neurosci. 13:700–707. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gordon AG: Endolymphatic hydrops and CSF
pressure. J Neurosurg. 60:1332–1334. 1984.PubMed/NCBI
|