Introduction
Atherosclerosis is a chronic inflammatory disease
that contributes to cardiovascular disease, a leading cause of
mortality (1). It is characterized
by excessive lipid deposition in the arterial wall and
atherosclerotic plaque formation (2,3). It
causes insufficient blood supply to coronary arteries and
myocardial ischemia, which are the pathological basis of coronary
heart disease (4). The etiology of
atherosclerosis is multifactorial and includes diabetes mellitus,
hypertension, smoking, dyslipidemia and genetic factors (5,6).
Atherosclerotic lesion formation starts with vascular endothelial
cell injury, lipid deposition and abnormal proliferation of
vascular smooth muscle cells (VSMCs) (7). During atherosclerosis progression,
proliferation and migration of VSMCs from the media to intima
contribute to intimal hyperplasia and vascular stenosis (8).
Adipose differentiation-related protein (ADRP), also
termed perilipin 2 or adipophilin, is expressed in the majority of
cell types and is commonly used as a lipid droplet marker (9). McIntosh et al (10) reported that ADRP interacts with
lipids on the lipid droplet surface and regulates the levels of key
enzymes and lipids that have critical roles in maintaining the
structure and function of lipid droplets. ADRP is highly expressed
in atherosclerotic plaques in humans (11). Xu et al (12) previously demonstrated that
elevation of ADRP expression in human atherosclerotic plaques is
closely associated with the instability of plaques, and Paul et
al (13) demonstrated that
ADRP deficiency protects apolipoprotein E (apoE)−/− mice
against atherosclerosis by suppressing foam cell formation, which
is a hallmark of atherosclerosis. However, the role of ADRP in the
proliferation and migration of human VSMCs induced by
platelet-derived growth factor (PDGF) in vitro and the
effect of ADRP knockdown on neointima formation in vivo
remains to be established.
Mitogen-activated protein kinases (MAPKs) are
expressed in all types of cells (14). The MAPK family consists of multiple
kinases, including p38 MAPK, c-Jun N-terminal kinase and
extracellular signal-regulated kinase (ERK) (15). Protein kinase B (Akt) is a
serine/threonine protein kinase, and Akt signaling is involved in
cell proliferation, migration and tumor metastasis (16,17).
Previous evidence has demonstrated that ERK and Akt signaling
pathways participate in the regulation of VSMC proliferation and
migration (16,18–20).
In the present study, ADRP knockdown was performed
in primary human aortic VSMCs using small interfering (si)RNA. The
cells were treated with PDGF following siRNA transfection.
Subsequently, proliferation and migration of VSMCs were measured
and the underlying mechanism was investigated. In addition, the
present study investigated the effect of ADRP knockdown on
atherosclerosis in apoE−/− mice.
Materials and methods
Cell culture
Primary human aortic VSMCs were purchased from CHI
Scientific, Ltd. (Jiangyin, China) and cultured in Dulbecco's
Modified Eagle's Medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) containing 15% fetal bovine serum (FBS;
Hyclone; GE Healthcare Life Sciences, Logan, UT, USA) at 37°C in a
humidified atmosphere with 5% CO2.
Transfection of siRNA
siRNA targeting ADRP and non-targeting siRNA (siCtr)
were synthesized by Shanghai GenePharma Co., Ltd. (Shanghai,
China). The sequences were as follows: siADRP,
5′-GAGACUGCCUAUUCUGAAUTT-3′; and siCtr,
5′-UUCUCCGAACGUGUCACGUTT-3′. VSMCs were seeded into 6-well plates
(4×105 cells/well). After culturing for 23 h at 37°C, the medium
was replaced with serum-free DMEM and the cells were cultured for 1
h at 37°C. Subsequently, Lipofectamine® 2000
Transfection reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
was used to transfect 100 pmol siADRP or siCtr into VSMCs. After 4
h, the culture medium was replaced by DMEM containing 15% FBS and
cultured at 37°C.
Cell viability
Cells were seeded into 96-well plates at a density
of 3×103 cells/well, cultured at 37°C for 24 h and
subsequently transfected with the siRNA as described. Twenty-four
hours post-transfection, the cells were incubated with PDGF (10
ng/ml; PeproTech, Inc., Rocky Hill, NJ, USA) at 37°C for 24, 48 and
72 h. The cells in the control group were cultured at 37°C without
any treatment. Subsequently, 5 mg/ml MTT reagent (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) was added into each well in 96-well
plates and incubated for 4 h at 37°C. To dissolve the formazan
crystals, dimethylsulfoxide (Sigma-Aldrich; Merck KGaA) was added
after removing the culture medium and the absorbance at 490 nm was
measured using a microplate reader (BioTek Instruments, Inc.,
Winooski, VT, USA).
Flow cytometry analysis
After stimulating with 10 ng/ml PDGF for 48 h at
37°C, the siRNA-transfected cells were fixed and permeabilized with
70% ethanol for 2 h at 4°C and harvested by centrifugation at 550 ×
g for 5 min. For cell cycle analysis, the cells (1–5×106
cells) were incubated with RNase A at 37°C for 30 min in a water
bath and subsequently stained with propidium iodide staining
solution (Wanleibio, Shenyang, China) at 4°C for 30 min in the
dark. Cell cycle distribution was analyzed by a BD Accuri C6 flow
cytometer using Accuri C6 software (both from BD Biosciences, San
Jose, CA, USA).
Wound healing assay
Cells were treated with PDGF for 48 h after siRNA
transfection. Subsequently, the cells were cultured to ~90%
confluence in serum-free DMEM containing 1 µg/ml mitomycin C
(Sigma-Aldrich; Merck KGaA) at 37°C for 1 h before they were
subjected to wound healing assay. Briefly, a 200 µl pipette tip was
used to scratch the cell layers. Subsequently, the wound was washed
with serum-free medium and further cultured in serum-free medium
for 0, 12 and 24 h at 37°C. The cells were imaged under an inverted
microscope (magnification, ×100) (Motic Incorporation, Ltd.,
Causeway Bay, Hong Kong).
Migration assay
Cell migration ability was determined by Transwell
migration assay. Briefly, cells in serum-free medium
(1×104 cells) were added into each well in the upper
chamber and DMEM containing 20% FBS was added into lower chamber.
Subsequently, the chamber (Corning Incorporated, Corning, NY, USA)
was cultured for 24 h at 37°C. After washing with PBS, the
non-migrated cells were removed by cotton swabs and the cells on
the lower membrane were fixed with 4% paraformaldehyde (Sinopharm
Chemical Reagent Co., Ltd., Shanghai, China) for 20 min at room
temperature, stained with crystal violet (Amresco, LLC, Solon, OH,
USA) and imaged under an inverted microscope (magnification, ×200)
(Motic Incorporation, Ltd.). The average numbers of migrated cells
in five fields of view were calculated.
Gelatin zymography
The activities of matrix metalloproteinase (MMP)-2
and MMP-9 were examined by gelatin zymography. At 24 h after
transfection, the cells were treated with PDGF for 24 h at 37°C.
The conditioned medium was collected and the protein concentration
was quantified using a Bicinchoninic Acid assay kit (Wanleibio).
Proteins (30 µg per lane) were subsequently separated by 10%
SDS-PAGE containing gelatin (Sigma-Aldrich; Merck KGaA). The gels
were washed twice for 40 min each in 2.5% Triton X-100 to remove
the SDS, incubated with developing buffer for 40 h at 37°C and
stained for 3 h at room temperature with 0.05% Coomassie Brilliant
Blue R-250 (Amresco, LLC). After three destaining washes in a
mixture of methanol and acetic acid (methanol + acetic acid: 30 +
10%, 30 min; 20 + 10%, 1 h; 10 + 5%, 2 h; respectively) at room
temperature, the gels were imaged under a Gel Imaging Analysis
system (Beijing Liuyi Biotechnology Co., Ltd., Beijing, China) and
the band intensities were quantified using Gel-Pro Analyzer
software version 4.0 (Media Cybernetics, Inc., Rockville, MD,
USA).
Animals
Male apoE−/− mice (6-weeks-old; weight,
20 g; n=24) were purchased from Beijing Vital River Laboratory
Animal Technology Co., Ltd. (Beijing, China). The mice were housed
in cages (temperature, 22°C) with a 12-h light/dark cycle and
allowed ad libitum access to food and water. The protocols
of animal experiments were performed following the National
Institutes of Health Laboratory Animal Care and Use Guidelines
(21) and approved by the Animal
Ethics Committee at Jilin University.
Animal model of atherosclerosis
ApoE−/− mice were randomized into the
following four groups: Control, Model, siCtr and siADRP.
ApoE−/− mice were fed a high-fat diet containing 0.15%
cholesterol and 21% fat for 12 weeks to establish the animal model
of atherosclerosis. One week after establishment of the mouse model
of atherosclerosis, the mice were injected with saline or
siCtr/siADRP (5 mg/kg) via the tail vein twice a week for 3 weeks
(n=6 per group). ApoE−/− mice fed a standard chow diet
served as the control. Following animal experiments, all mice were
euthanized by sodium pentobarbital (100 mg/kg body weight; LookChem
Co., Ltd., Hangzhou, China) and underwent thoracotomy and the
thoracic aortas were exposed and excised. Some aortas were fixed in
4% paraformaldehyde for histological evaluation and some aortas
were snap-frozen in liquid nitrogen and stored at −70°C for further
analyses.
Hematoxylin and eosin (H&E)
staining
The paraformaldehyde-fixed aortas (for 48 h, at room
temperature) were dehydrated in gradient ethanol, embedded in
paraffin and cut into 5-µm sections. The sections were stained with
H&E and imaged under an Olympus DP73 microscope (Olympus
Corporation, Tokyo, Japan). The neointima and media areas were
measured and the ratio of neointima/media area was calculated as
described previously (22).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from mouse aortas using a
Total RNA Extraction kit (Tiangen Biotech Co., Ltd., Beijing,
China) and cDNA was synthesized using Super M-MLV Reverse
Transcriptase (BioTeke Corporation, Beijing, China) according to
the protocol described below. The reaction mixture contained 1 µg
RNA, 2 µl dNTP (BioTeke Corporation), 1 µl random primer (Sangon
Biotech Co., Ltd., Shanghai, China), 1 µl oligo (dT)15 (Tiangen
Biotech Co., Ltd.) and ddH2O was added to a final volume
of 14.5 µl. The reaction was denatured in a 70°C water bath for 5
min, and incubated in an ice bath for 2 min. To perform the RT
reaction, 4 µl 5X buffer, 0.5 µl RNasin and 1 µl super M-MLV
reverse transcriptase (200 U) was added into the reaction mixture
and incubated at 25°C for 10 min, 42°C for 50 min and 95°C for 5
min. The mRNA expression level of ADRP was evaluated using a
Real-Time Quantitative PCR system (Bioneer Corporation, Daejeon,
Korea) and SYBR-Green I (Beijing Solarbio Science & Technology
Co., Ltd., Beijing, China). qPCR conditions consisted of 10 min at
95°C, followed by 10 sec at 95°C, 20 sec at 60°C and 30 sec at 72°C
(40 cycles). The relative gene expression was assessed using the
2−ΔΔCq method to normalize expression to β-actin
(23). Each experiment was
repeated three times. The primer sequences were as follows: ADRP,
5′-AAGAGCCAGGAGACCATT-3′ (forward) and 5′-CCACCCACGAGACATAGA-3′
(reverse); and β-actin, 5′-TTTTCCAGCCTTCCTTCTTGGGTAT-3′ (forward)
and 5′-CTGTGTTGGCATAGAGGTCTTTACG-3′ (reverse).
Western blotting
Primary human aortic VMSCs and mice aortic tissues
were treated with lysis buffer (Wanleibio) containing 1% PMSF on
ice for 5 min to obtain the protein extracts (10,005 × g, 10 min,
4°C) and a BCA assay kit (Wanleibio) was used to determine the
protein concentration according to the manufacturer's protocol.
Total proteins were boiled with loading buffer, subjected to 8 or
10% SDS-PAGE (40 µg per lane) and transferred onto polyvinylidene
fluoride membranes (EMD Millipore, Billerica, MA, USA). After
blocking with 1% bovine serum albumin (Amresco, LLC) or 5% non-fat
milk for 1 h at room temperature with gentle shaking, the membranes
were incubated with rabbit anti-human polyclonal antibodies against
proliferating cell nuclear antigen (PCNA; cat. no. bs-2006R; 1:500;
BIOSS, Beijing, China), MMP-2 (cat. no. BA0569; 1:400; Wuhan Boster
Biological Technology, Ltd., Wuhan, China), MMP-9 (cat. no. BA0573;
1:400; Wuhan Boster Biological Technology, Ltd.), ERK (cat. no.
bs-2637R; 1:500; BIOSS), phosphorylated (p)-ERK (cat. no. bs-1522R;
1:500; BIOSS), Akt (cat. no. sc-8312; 1:500; Santa Cruz
Biotechnology, Inc., Dallas, Texas, USA), p-Akt (cat. no.
sc-135651; 1:500; Santa Cruz Biotechnology, Inc.), ADRP (cat. no.
bs-1164R; 1:500; BIOSS) and β-actin (cat. no. sc-47778; 1:1,000;
Santa Cruz Biotechnology, Inc.) at 4°C overnight, followed by
incubation with a horseradish peroxidase-conjugated goat
anti-rabbit/mouse secondary antibody (cat. no. WLA023/WLA024;
1:5,000; Wanleibio) at 37°C for 45 min. β-actin served as an
internal control. The protein bands were visualized using Enhanced
Chemilumiescence reagent (Wanleibio) and quantified by Gel-Pro
Analyzer software version 4.0 (Media Cybernetics, Inc., Rockville,
MD, USA). Each experiment was repeated three times.
Statistical analysis
Data are presented as the mean ± standard deviation.
Statistical analyses were performed using one-way analysis of
variance followed by Bonferroni's post hoc test using GraphPad
Prism software version 5.0 (GraphPad Software, Inc., La Jolla, CA,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
ADRP knockdown attenuates PDGF-induced
proliferation of human VSMCs
VSMCs were transfected with or without siCtr/siADRP
and treated with 10 ng/ml PDGF. Cell viability was examined by MTT
assay. The results demonstrated that PDGF increased VSMC viability
at 24, 48 and 72 h, compared with the control group (Fig. 1A). However, ADRP knockdown
significantly inhibited the increases in VSMC viability induced by
PDGF incubation at 72 h. Furthermore, the present study analyzed
the cell cycle distribution in VSMCs. It was observed that the
G1-phase cell population in the PDGF group was significantly lower
compared with the control group (Fig.
1B). The cell percentage at S-phase in the PDGF group was
significantly higher compared with the control group. However, in
the presence of PDGF, siADRP significantly increased G1-phase cell
population and decreased the cell population at S-phase and
G2-phase (Fig. 1B). The expression
level of proliferation marker PCNA was confirmed by western
blotting (Fig. 1C). PDGF
significantly increased PCNA protein expression levels compared
with control cells, while ADRP knockdown attenuated PDGF-stimulated
elevation of PCNA expression (Fig.
1C).
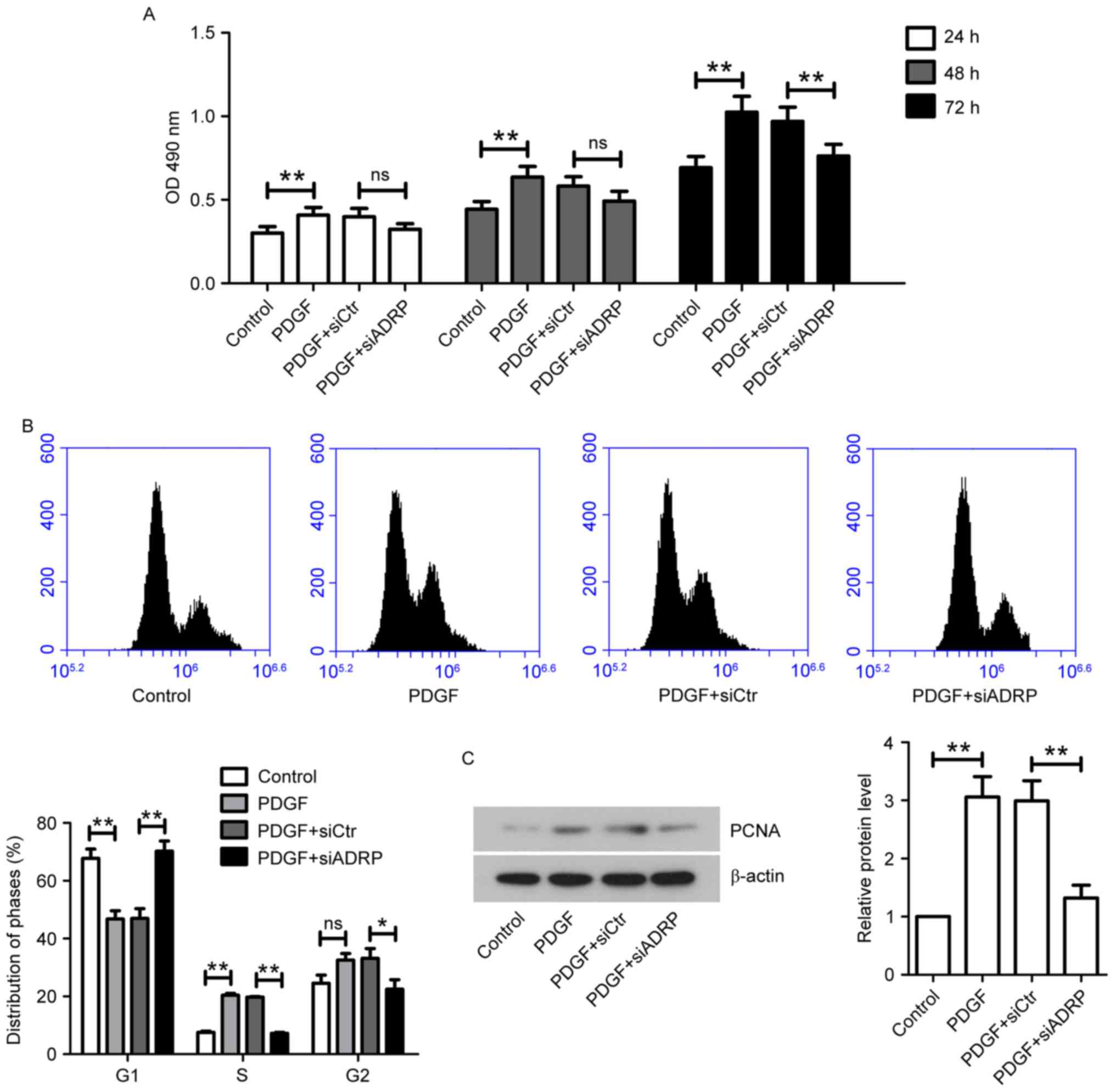 | Figure 1.Effect of ADRP knockdown on VSMC
viability and cell cycle progression induced by PDGF. (A) At 24 h
after transfection with siCtr or siADRP, VSMCs were stimulated with
10 ng/ml PDGF. Cell viability was measured by MTT assay at 24, 48
and 72 h. The absorbance at 490 nm was measured. VSMCs were
incubated for 48 h with PDGF at 24 h post-transfection. Cell cycle
distribution was analyzed by (B) flow cytometry and PCNA protein
expression was quantified by (C) western blotting. β-actin served
as an internal control. Data are presented as the mean ± standard
deviation. *P<0.05 and **P<0.01. ADRP, adipose
differentiation-related protein; VSMCs, vascular smooth muscle
cells; PDGF, platelet-derived growth factor; siRNA, small
interfering RNA; siCtr, control siRNA; siADRP, siRNA targeting
ADRP; PCNA, proliferating cell nuclear antigen; OD, optical
density. |
ADRP knockdown inhibits
PDGF-stimulated migration of VSMCs
The present study further examined the effect of
ADRP knockdown on cell migration by performing a wound healing
assay. As presented in Fig. 2A,
PDGF significantly promoted wound closure at 12 and 24 h after the
scratch, whereas knockdown of ADRP exhibited an inhibitory effect
on PDGF-induced migration of VSMCs. This inhibitory effect on cell
migration was further confirmed by the Transwell migration assay
(Fig. 2B). MMP-2 and MMP-9 are
critical for tissue remodeling and cell migration (24). Therefore, the effect of ADRP
knockdown on MMP-2 and MMP-9 in cells exposed to PDGF was examined
using western blotting (Fig. 2C)
and gelatin zymography (Fig. 2D).
The results indicated that PDGF increased the expression and
activities of MMP-2 and MMP-9 in VSMCs compared with control cells,
while ADRP knockdown significantly decreased PDGF-induced
upregulation of MMP-2 and MMP-9 levels and activities.
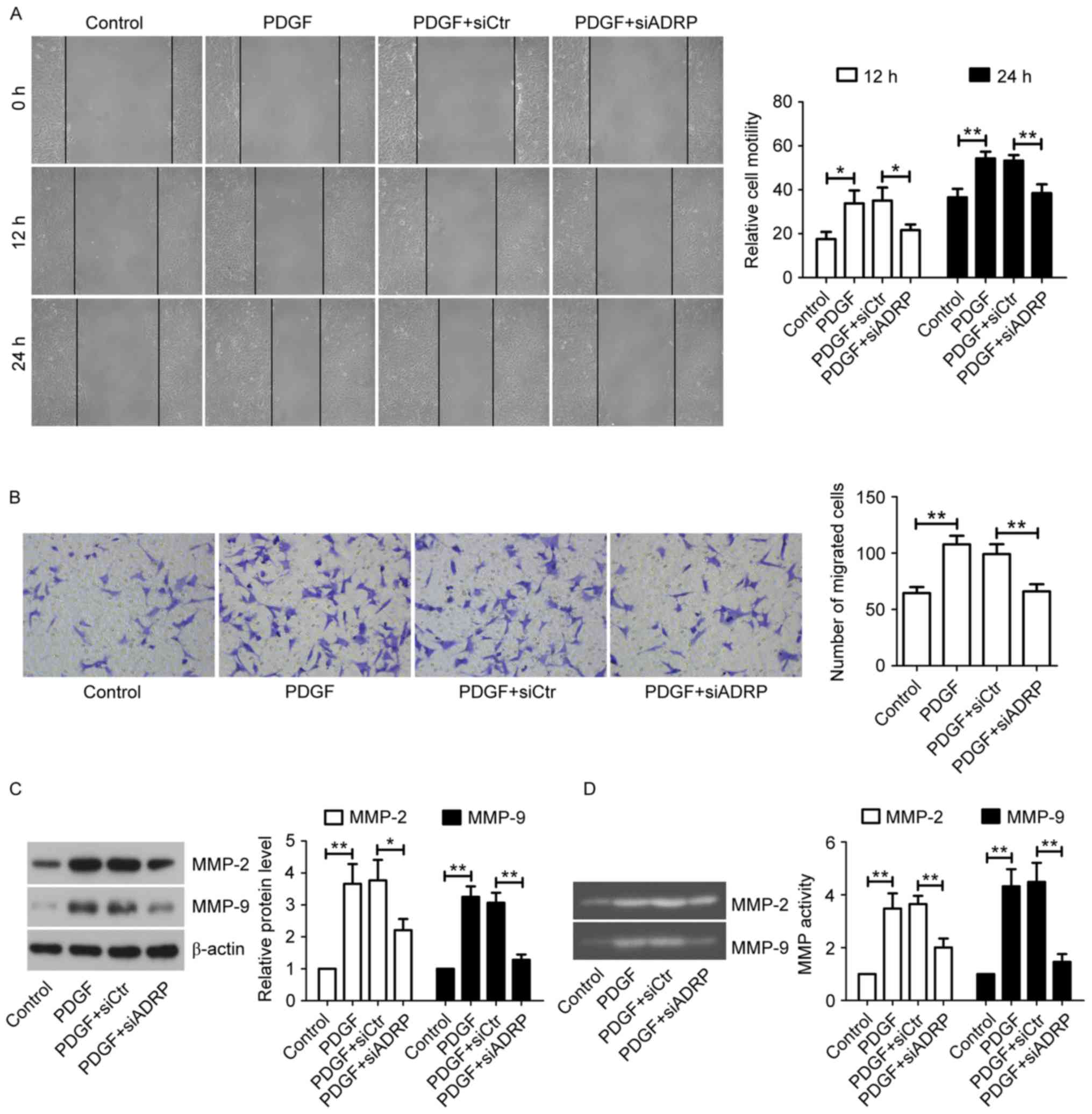 | Figure 2.Effect of ADRP knockdown on VSMC
migration induced by PDGF. (A) Cell migration was determined by
wound healing assay at 12 and 24 h (magnification, ×100). (B) Cell
migration ability was confirmed by Transwell migration assay
(magnification, ×200). VSMCs were incubated for 24 h with PDGF at
24 h post-transfection. The protein levels and activities of MMP-2
and MMP-9 were examined by (C) western blotting and (D) gelatin
zymography. Data are presented as the mean ± standard deviation.
*P<0.05 and **P<0.01, as indicated by brackets. ADRP, adipose
differentiation-related protein; VSMCs, vascular smooth muscle
cells; PDGF, platelet-derived growth factor; MMP, matrix
metalloproteinase; siRNA, small interfering RNA: siCtr, control
siRNA; siADRP, siRNA targeting ADRP. |
ADRP knockdown inhibits the ERK and
Akt signaling pathways
Accumulating evidence has demonstrated that the ERK
and Akt signaling pathways are involved in the proliferation and
migration of VSMCs (25,26). However, whether ADRP knockdown
inhibits VSMC proliferation and migration via the ERK and Akt
pathways remains to be established. Western blotting results
demonstrated that the ratios of p-ERK/ERK and p-Akt/Akt in VSMCs
were decreased by ADRP knockdown, despite the presence of PDGF
(Fig. 3).
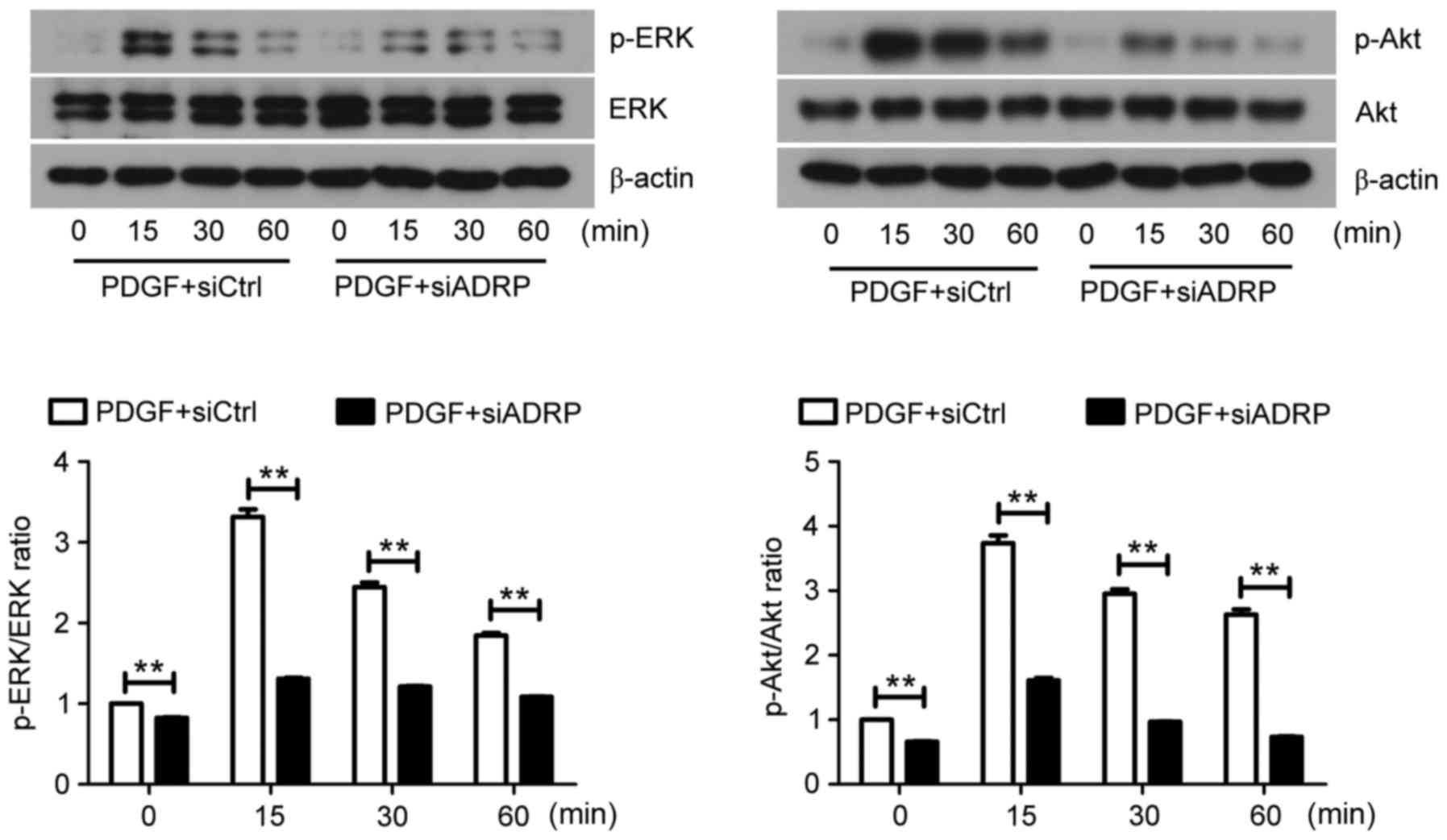 | Figure 3.Effect of ADRP knockdown on the ERK
and Akt signaling pathways. At 48 h after siRNA transfection, VSMCs
were exposed to PDGF for 0, 15, 30 and 60 min. The protein
expression of ERK, p-ERK, Akt and p-Akt was subsequently examined
by western blotting. Representative images are presented. Data are
presented as the mean ± standard deviation. **P<0.01, as
indicated by brackets. ADRP, adipose differentiate-related protein;
ERK, extracellular signal-regulated kinase; siRNA, small
interfering RNA; VSMCs, vascular smooth muscle cells; PDGF,
platelet-derived growth factor; p-, phosphorylated; siCtrl, control
siRNA; siADRP, siRNA targeting ADRP. |
ADRP knockdown attenuates neointima
formation in a mouse model of atherosclerosis
To investigate the effect of ADRP on neointima
formation in vivo, animal models of atherosclerosis were
administrated with siCtr or siADRP twice a week for 3 weeks, and
aortas were obtained. The mRNA and protein expression of ADRP in
the aortas was examined at the end of the animal experiments. The
results demonstrated that ADRP mRNA and protein expression was
significantly increased in model mice compared with controls
(Fig. 4A and B). However, elevated
ADRP expression was significantly reduced by siADRP injection, as
indicated by RT-qPCR and western blotting. In addition, neointima
formation was assessed using H&E staining. The results
indicated that the model mice exhibited a reduced lumen area and
increased neointima area, and neointima/media area ratio, when
compared with control mice (Fig.
4C). However, the lumen area was significantly increased and
the ratio of neointima/media area was significantly decreased in
the aortas of the siADRP-treated mice compared with siCtr-treated
mice (Fig. 4C).
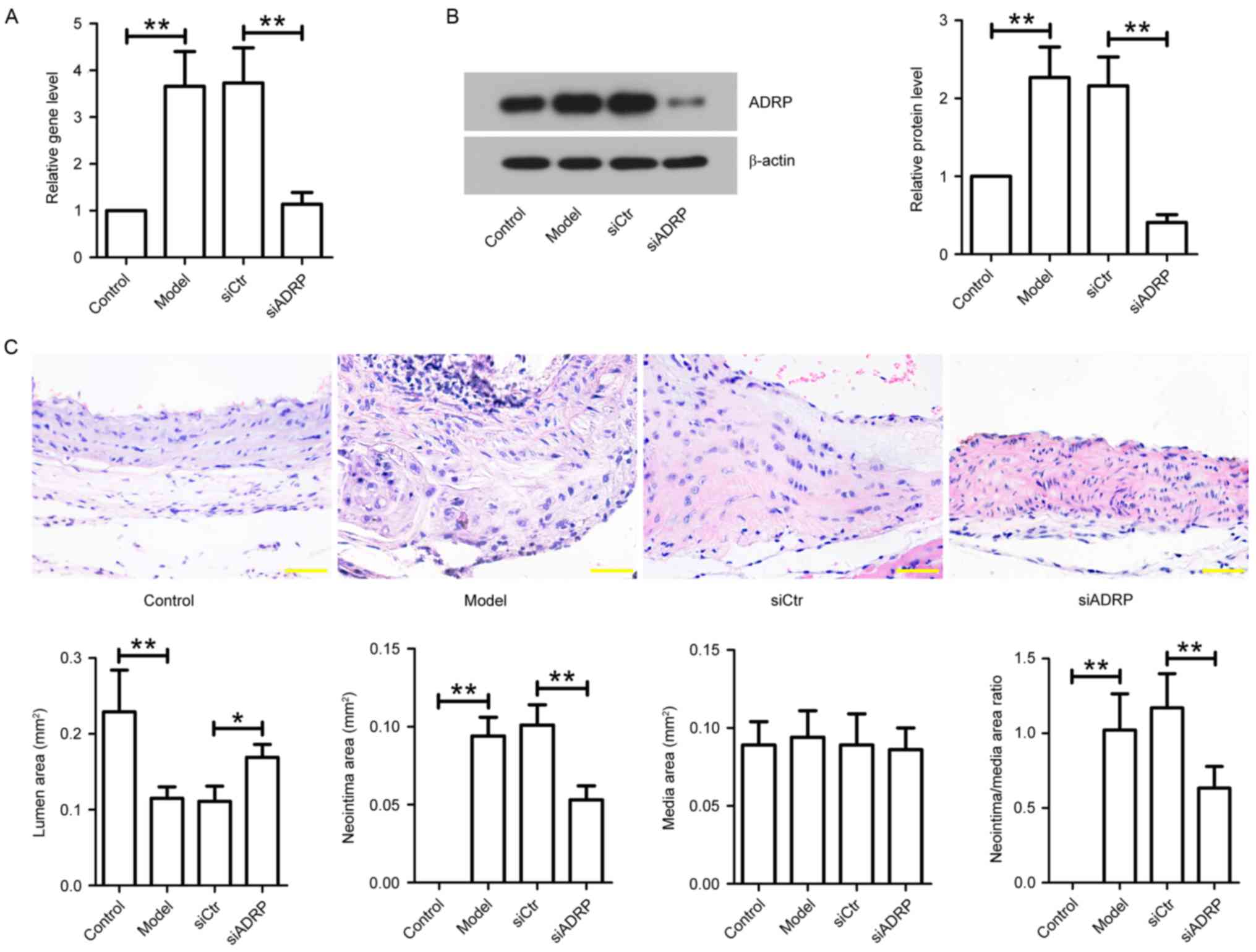 | Figure 4.Effect of ADRP knockdown on neointima
formation in apoE−/−mice. After establishing an in
vivo model of atherosclerosis, the mice were injected with
siCtr or siADRP twice a week. After 3 weeks of siRNA injection,
aortas were obtained. (A) ADRP mRNA levels were examined by reverse
transcription-quantitative polymerase chain reaction. (B) ADRP
protein levels were determined by western blotting. β-actin served
as an internal control. (C) Aortas were subjected to hematoxylin
and eosin staining, and lumen area, neointima area, media area and
neointima/media area ratios were measured to evaluate neointima
formation in mice. Scale bar, 50 µm. Data are presented as the mean
± standard deviation. *P<0.05 and **P<0.01, as indicated by
brackets. ADRP, adipose differentiation-related protein; apoE,
apolipoprotein E; siRNA, small interfering RNA; siCtr, control
siRNA; siADRP, siRNA targeting ADRP. |
Discussion
In the present study, ADRP was silenced in VSMCs
using siRNA. The results demonstrated that ADRP knockdown inhibited
PDGF-induced proliferation and migration of VSMCs by inhibiting the
ERK and Akt signaling pathways. In addition, apoE−/−
mice were used to establish an animal model of atherosclerosis in
order to examine the effect of ADRP knockdown on atherosclerosis
in vivo. The results indicated that ADRP knockdown inhibited
neointima formation in mice.
Abnormal proliferation of VSMCs is key to the
development of atherosclerosis (27). It has been widely demonstrated that
multiple growth factors and cytokines, including PDGF, contribute
to the abnormal proliferation of VSMCs (28,29).
ADRP is a major lipid-droplet protein. It regulates the formation
of foam cells from macrophages and has an important role in the
development of atherosclerosis (12,30).
However, to the best of our knowledge, no previous studies have
investigated the effect of ADRP knockdown on the proliferation of
VSMCs induced by PDGF. The present study demonstrated that ADRP
knockdown inhibited PDGF-induced growth and proliferation of VSMCs,
and that this anti-proliferative effect was associated with
increased cell cycle arrest at G1 phase. Cell proliferation is
controlled by cell cycle progression, which is regulated by the
complexes of cyclins and cyclin-dependent kinases (31). PCNA is a cell cycle protein in the
nucleus and functions in DNA synthesis (32). PCNA is commonly used as a marker to
assess the state of cell proliferation (33). The results of the current study
demonstrated that ADRP silencing decreased the expression level of
PCNA in PDGF-treated VSMCs. These results indicated that the
inhibitory effect of ADRP knockdown on cell proliferation may be
due to cell cycle arrest at G1 phase and the downregulation of PCNA
expression.
A previous study demonstrated that migration of
VSMCs is also involved in the progression of cardiovascular
diseases, such as atherosclerosis (34). The proliferative and migratory
activities can be enhanced by growth promoters such as PDGF
(35). However, the effect of ADRP
knockdown on PDGF-treated VSMCs remains unknown. The current study
also assessed the migration of VSMCs using wound healing and
Transwell migration assays. The results demonstrated that
PDGF-stimulated VSMC migration was inhibited by knockdown of ADRP.
MMPs belong to the zinc-dependent proteinase family (36). During VSMC migration, MMPs are
secreted to degrade extracellular matrix proteins and the vascular
basement membrane (35,37). A recent study reported that MMP-2
and MMP-9 are associated with the pathogenesis of atherosclerosis,
and the levels of MMPs are upregulated in atherosclerotic plaques
(34). In the present study, ADRP
knockdown significantly decreased MMP-2 and MMP-9 levels and
activities in PDGF-stimulated VSMCs. The results indicated that
ADRP silencing suppresses VSMC migration by regulating the
expression and activities of MMP-2 and MMP-9.
PDGF binds with its receptor and subsequently
activates various intracellular signaling pathways, including
signal transducer and activator of transcription, ERK/MAPK and
phosphatidylinositol 3-kinase/Akt, which contribute to VSMC
proliferation and migration (27,38).
However, whether ADRP regulates the proliferation and migration of
VSMCs via the ERK and Akt pathways remains unclear. After silencing
ADRP expression using siADRP in the current study, it was observed
that ADRP knockdown inhibited the phosphorylation of ERK and Akt in
PDGF-treated VSMCs. These results indicate that ADRP silencing may
suppress proliferation and migration of VSMCs by inhibiting ERK and
Akt signaling pathways.
Apolipoprotein E (apoE) is a primary apolipoprotein
and functions in maintaining cholesterol balance and lipid
metabolism (39).
ApoE−/− mice are susceptible to atherosclerosis and have
been widely used to induce atherosclerosis (40). Previous studies have reported that
ADRP overexpression induces foam cell formation, and ADRP
deficiency attenuates atherogenesis by impairing the formation of
foam cells (13,41). In the present study, siADRP
administration significantly decreased ADRP mRNA and protein
expression levels in aortas of apoeE−/− mice. The
current study subsequently assessed the effect of ADRP knockdown on
neointima formation in apoE−/− mice. Neointima
formation, characterized by thickening of intimal layer, is
involved in various diseases, including atherosclerosis and
angioplasty (42). The results
demonstrated that ADRP silencing significantly increased the lumen
area, and decreased the neointima area and neointima/media area
ratio in vascular tissues, as determined by H&E staining.
In conclusion, the present study demonstrated that
ADRP knockdown significantly inhibited PDGF-induced proliferation
and migration of VSMCs and arrested the cell cycle at G1 phase,
accompanied with inhibition of the ERK and Akt signaling pathways.
In the in vivo study, the results indicated that knockdown
of ADRP suppressed neointima formation in apoE−/− mice.
Therefore, ADRP may represent a promising target for the treatment
of atherosclerosis.
References
|
1
|
Cheng HY, Gaddis DE, Wu R, McSkimming C,
Haynes LD, Taylor AM, McNamara CA, Sorci-Thomas M and Hedrick CC:
Loss of ABCG1 influences regulatory T cell differentiation and
atherosclerosis. J Clin Invest. 126:3236–3246. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mathur P, Ding Z, Saldeen T and Mehta JL:
Tocopherols in the prevention and treatment of atherosclerosis and
related cardiovascular disease. Clin Cardiol. 38:570–576. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Su G, Sun G, Liu H, Shu L, Zhang J, Guo L,
Huang C and Xu J: Niacin suppresses progression of atherosclerosis
by inhibiting vascular inflammation and apoptosis of vascular
smooth muscle cells. Med Sci Monit. 21:4081–4089. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li T, Li D, Xu H, Zhang H, Tang D and Cao
H: Wen-Xin Decoction ameliorates vascular endothelium dysfunction
via the PI3K/AKT/eNOS pathway in experimental atherosclerosis in
rats. BMC Complement Altern Med. 16:272016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Abu-Fanne R, Maraga E, Abd-Elrahman I,
Hankin A, Blum G, Abdeen S, Hijazi N, Cines DB and Higazi AA:
α-Defensins induce a post-translational modification of low density
lipoprotein LDL that promotes atherosclerosis at normal levels of
plasma cholesterol. J Biol Chem. 291:2777–2786. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim HJ: Role of nucleotide-binding and
oligomerization domain 2 protein (NOD2) in the development of
atherosclerosis. Korean J Physiol Pharmacol. 19:479–484. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhai H, Chen QJ, Gao XM, Ma YT, Chen BD,
Yu ZX, Li XM, Liu F, Xiang Y, Xie J and Yang YN: Inhibition of the
NF-κB pathway by R65 ribozyme gene via adeno-associated virus
serotype 9 ameliorated oxidized LDL induced human umbilical vein
endothelial cell injury. Int J Clin Exp Pathol. 8:9912–9921.
2015.PubMed/NCBI
|
|
8
|
Zhou Y, Zhang MJ, Li BH, Chen L, Pi Y, Yin
YW, Long CY, Wang X, Sun MJ, Chen X, et al: PPARγ inhibits VSMC
proliferation and migration via attenuating oxidative stress
through upregulating UCP2. PLoS One. 11:e01547202016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu F, Wang C, Zhang L, Xu Y, Jang L, Gu
Y, Cao X, Zhao X, Ye J and Li Q: Metformin prevents hepatic
steatosis by regulating the expression of adipose
differentiation-related protein. Int J Mol Med. 33:51–58.
2014.PubMed/NCBI
|
|
10
|
McIntosh AL, Senthivinayagam S, Moon KC,
Gupta S, Lwande JS, Murphy CC, Storey SM and Atshaves BP: Direct
interaction of Plin2 with lipids on the surface of lipid droplets:
A live cell FRET analysis. Am J Physiol Cell Physiol.
303:C728–C742. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Magne J, Aminoff A, Sundelin J Perman,
Mannila MN, Gustafsson P, Hultenby K, Wernerson A, Bauer G,
Listenberger L, Neville MJ, et al: The minor allele of the missense
polymorphism Ser251Pro in perilipin 2 (PLIN2) disrupts an α-helix,
affects lipolysis, and is associated with reduced plasma
triglyceride concentration in humans. FASEB J. 27:3090–3099. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu B, Zhao H, Wang S, Sun X and Qin X:
Increased ADRP expression in human atherosclerotic lesions
correlates with plaque instability. Int J Clin Exp Med.
8:5414–5421. 2015.PubMed/NCBI
|
|
13
|
Paul A, Chang BH, Li L, Yechoor VK and
Chan L: Deficiency of adipose differentiation-related protein
impairs foam cell formation and protects against atherosclerosis.
Circ Res. 102:1492–1501. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wei L, Li Y and Suo Z: TSPAN8 promotes
gastric cancer growth and metastasis via ERK MAPK pathway. Int J
Clin Exp Med. 8:8599–8607. 2015.PubMed/NCBI
|
|
15
|
Park SL, Hwang B, Lee SY, Kim WT, Choi YH,
Chang YC, Kim WJ and Moon SK: p21WAF1 Is required for
interleukin-16-induced migration and invasion of vascular smooth
muscle cells via the p38MAPK/Sp-1/MMP-9 pathway. PLoS One.
10:e01421532015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xu T, Zhu H, Li D, Lang Y, Cao L, Liu Y,
Wu W and Chen D: Luteolin inhibits angiotensin II-stimulated VSMC
proliferation and migration through downregulation of Akt
phosphorylation. Evid Based Complement Alternat Med.
2015:9317822015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu W, Bagaitkar J and Watabe K: Roles of
AKT signal in breast cancer. Front Biosci. 12:4011–4019. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang SW, Lim L, Ju S, Choi DH and Song H:
Effects of matrix metalloproteinase 13 on vascular smooth muscle
cells migration via Akt-ERK dependent pathway. Tissue Cell.
47:115–121. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yu X, Li Z, Chen G and Wu WK: MicroRNA-10b
induces vascular muscle cell proliferation through Akt pathway by
targeting TIP30. Curr Vasc Pharmacol. 13:679–686. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
He M, Xue ZM, Li J and Zhou BQ:
Breviscapine inhibits high glucose-induced proliferation and
migration of cultured vascular smooth muscle cells of rats via
suppressing the ERK1/2 MAPK signaling pathway. Acta Pharmacol Sin.
33:606–614. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Institute of Laboratory Animal Resources:
Guide for the Care and Use of Laboratory Animals. National Academy
Press; Washington, DC: 1996
|
|
22
|
Kumar A, Hoover JL, Simmons CA, Lindner V
and Shebuski RJ: Remodeling and neointimal formation in the carotid
artery of normal and P-selectin-deficient mice. Circulation.
96:4333–4342. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang Y, George J, Li Y, Olufade R and
Zhao X: Matrix metalloproteinase-9 expression is enhanced in renal
parietal epithelial cells of zucker diabetic fatty rats and is
induced by albumin in in vitro primary parietal cell culture. PLoS
One. 10:e01232762015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guo J, Li L, Wu YJ, Yan Y, Xu XN, Wang SB,
Yuan TY, Fang LH and Du GH: Inhibitory effects of Brazilin on the
vascular smooth muscle cell proliferation and migration induced by
PDGF-BB. Am J Chin Med. 41:1283–1296. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Park ES, Kang SI, Yoo KD, Lee MY, Yoo HS,
Hong JT, Shin HS, Kim B and Yun YP: Camptothecin inhibits
platelet-derived growth factor-BB-induced proliferation of rat
aortic vascular smooth muscle cells through inhibition of PI3K/Akt
signaling pathway. Exp Cell Res. 319:982–991. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li PC, Sheu MJ, Ma WF, Pan CH, Sheu JH and
Wu CH: Anti-restenotic roles of dihydroaustrasulfone alcohol
involved in inhibiting PDGF-BB-stimulated proliferation and
migration of vascular smooth muscle cells. Mar Drugs. 13:3046–3060.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Han JH, Lee SG, Jung SH, Lee JJ, Park HS,
Kim YH and Myung CS: Sesamin inhibits PDGF-Mediated proliferation
of vascular smooth muscle cells by upregulating p21 and p27. J
Agric Food Chem. 63:7317–7325. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yan G, Wang Q, Hu S, Wang D, Qiao Y, Ma G,
Tang C and Gu Y: Digoxin inhibits PDGF-BB-induced VSMC
proliferation and migration through an increase in ILK signaling
and attenuates neointima formation following carotid injury. Int J
Mol Med. 36:1001–1011. 2015.PubMed/NCBI
|
|
30
|
Wang J, Si Y, Wu C, Sun L, Ma Y, Ge A and
Li B: Lipopolysaccharide promotes lipid accumulation in human
adventitial fibroblasts via TLR4-NF-κB pathway. Lipids Health Dis.
11:1392012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gu Y, Chen X, Shang C, Singh K, Barzegar
M, Mahdavian E, Salvatore BA, Jiang S and Huang S: Fusarochromanone
induces G1 cell cycle arrest and apoptosis in COS7 and HEK293
cells. PLoS One. 9:e1126412014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Poosarla C, Ramesh M, Ramesh K, Gudiseva
S, Bala S and Sundar M: Proliferating cell nuclear antigen in
premalignancy and oral squamous cell carcinoma. J Clin Diagn Res.
9:ZC39–ZC41. 2015.PubMed/NCBI
|
|
33
|
Xie M, Bu P, Li F, Lan S, Wu H, Yuan L and
Wang Y: Neonatal bisphenol A exposure induces meiotic arrest and
apoptosis of spermatogenic cells. Oncotarget. 7:10606–10615. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shih MF, Pan KH and Cherng JY: Possible
mechanisms of Di(2-ethylhexyl) phthalate-induced MMP-2 and MMP-9
expression in A7r5 Rat vascular smooth muscle cells. Int J Mol Sci.
16:28800–28811. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rudijanto A: The role of vascular smooth
muscle cells on the pathogenesis of atherosclerosis. Acta Med
Indones. 39:86–93. 2007.PubMed/NCBI
|
|
36
|
Aroui S, Aouey B, Chtourou Y, Meunier AC,
Fetoui H and Kenani A: Naringin suppresses cell metastasis and the
expression of matrix metalloproteinases (MMP-2 and MMP-9) via the
inhibition of ERK-P38-JNK signaling pathway in human glioblastoma.
Chem Biol Interact. 244:195–203. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ge X, Chen S, Liu M, Liang T and Liu C:
Evodiamine attenuates PDGF-BB-Induced migration of rat vascular
smooth muscle cells through activating PPARγ. Int J Mol Sci.
16:28180–28193. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen S, Liu B, Kong D, Li S, Li C, Wang H
and Sun Y: Atorvastatin calcium inhibits phenotypic modulation of
PDGF-BB-induced VSMCs via down-regulation the Akt signaling
pathway. PLoS One. 10:e01225772015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhu XJ, Chen LH and Li JH: The effects of
aerobic exercise on plasma adiponectin level and
adiponectin-related protein expression in myocardial tissue of
ApoE(−/−) mice. J Sports Sci Med. 14:877–882. 2015.PubMed/NCBI
|
|
40
|
Getz GS and Reardon CA: ApoE knockout and
knockin mice: The history of their contribution to the
understanding of atherogenesis. J Lipid Res. 57:758–766. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Feng X, Yuan Y, Wang C, Feng J, Yuan Z,
Zhang X, Sui W, Hu P, Zheng P and Ye J: Autophagy involved in
lipopolysaccharide-induced foam cell formation is mediated by
adipose differentiation-related protein. Lipids Health Dis.
13:102014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Huang L, Zhang SM, Zhang P, Zhang XJ, Zhu
LH, Chen K, Gao L, Zhang Y, Kong XJ, Tian S, et al: Interferon
regulatory factor 7 protects against vascular smooth muscle cell
proliferation and neointima formation. J Am Heart Assoc.
3:e0013092014. View Article : Google Scholar : PubMed/NCBI
|














