Introduction
Temporal lobe epilepsy (TLE) is characterized by the
recurrence of seizure activity; however, the impact of frequent
seizure activity on the neuronal networks of the brain is still not
completely understood. Likewise, it is unclear why hippocampal
newborn neurons after status epilepticus (SE) exhibit
hyperexcitability in the hippocampal network and then influence
ongoing epilepsy-related processes.
Seizure-induced increases in hippocampal
neurogenesis have been observed in many TLE models, which are
thought to contribute to abnormal brain activity (1). The newly-generated neurons originate
primarily from neural progenitor cells in the subgranular zone of
the dentate gyrus (DG) of hippocampus after epilepsy. They
eventually integrate into hippocampal circuitry and may contribute
to hippocampal network plasticity after the initial insult
(2). These newly-generated neurons
have immature synapses (3), which
can receive synaptic input and transmit action potential output
(4), and therefore become the
center of spontaneous recurrent seizures (SRS).
5-bromo-2′-deoxyuridine (BrdU), an S-phase-specific marker, allows
the detection of newly formed cells by incorporation into DNA
(5). Because BrdU application is
mainly conducted by intraperitoneal injection, epilepsy or ischemia
which disturb or disrupt the function of the blood-brain barrier,
might induce increases in the numbers of BrdU-labeled cells which
is independent of changes in proliferation due to the altered BrdU
availability in the brain (6,7).
Doublecortin (DCX), a protein associated with cell migration, which
express in late type-3 progenitor cells in the process of neural
stem cell proliferation and differentiation (8). DCX-positive cells are thought to be a
migratory state as DCX plays a pivotal role during neural
development where it is involved in radial glia-mediated neuronal
migration and in hippocampal development (9). DCX-positive cells represent neural
progenitor cell migration during hippocampal neurogenesis and the
expression of DCX in newly-generated cells was increased after
epilepsy stimuli (10). Thus, DCX
marker was used in our study to identify the developmental stage of
migration and differentiation of newborn neurons in
hippocampus.
Axon initial segment (AIS) is an excitable neuronal
domain positioned between the axonal and somato-dendritic
compartment, which is involved in action potential initiation and
fine-tunes the excitability of neurons (11). In recent years, our understanding
of the molecular structure of AIS, its structural and functional
plasticity related to the excitability of neurons, has seen major
advances (12,13). Ankyrin G (AnkG) protein, a vital
scaffolding molecule in AIS has been accepted and widely used as a
biomarker to reflect the molecular structure of AIS (14). It is now widely accepted that the
structural properties of AIS, such as length or location relative
to the soma, change in an activity-dependent manner (15). This structural plasticity of AIS is
known to be crucial for homeostatic control of neuronal
excitability. The impact of AIS on neuronal excitability is
dependent on the composition and characteristics of ion channels in
the domain (16). In cultured
hippocampus neurons, the location of AIS changes during the
depolarization, but the movement of AIS can be prevented by the
T/L-type calcium currents blocker Mibefradil (17). However, we still have little
knowledge about how AIS plasticity in hippocampal newly-generated
neurons altered after SE and its role in recurrent seizures. So
far, the effects of calcium-current blocker Mibefradil on AIS
plasticity and neuronal excitability in newly-generated neurons
after epilepsy in vivo have not been determined.
In the present study, we first established
pilocarpine-induced epileptic rats to assess hippocampal
neurogenesis after pilocarpine-induced TLE and then explore AIS
plasticity in hippocampal newly-generated neurons using
immunofluorescent labeling of DCX and AnkG. Because voltage-gated
calcium channels in AIS is essential for neuronal excitability, we
suppose that used calcium channels blocker Mibefradil treatment
could have effects on changes of AIS plasticity in hippocampal
neurogenesis and neuronal excitability. Simultaneously, we employed
electroencephalography (EEG) to detect the long-term consequences
after treatment in pilocarpine-induced TLE rats. The ultimate goal
of this study is to provide experimental evidence for AIS
plasticity in hippocampal neurogenesis in the management of
epileptic patients.
Materials and methods
Animals and pilocarpine-induced
epilepsy models
Healthy, young male Sprague-Dawley (SD) rats (aged
6–8 weeks, 200–250 g, n=72) were used in this study. Rats were
randomly assigned as control rats, rats subjected to pilocarpine,
post-SE rats treated with Mibefradil and treated with Mibefradil
only. Rats were injected intraperitoneally (i.p.) with lithium
chloride (127 mg/kg; Sigma-Aldrich, St. Louis, MO, USA) and
pilocarpine (25 mg/kg; Sigma-Aldrich). Racine's classification was
used to evaluate seizure severity (18). After SE, all rats were administered
with chloral hydrate injection (3 ml/kg, i.p.; Tonghua Dongbao
Pharmaceutical Co., Ltd., Tonghua, China) to stop behavioral
seizures. The matched control rats were injected i.p. with the same
amount of normal saline. All procedures involving animals were
performed according to protocols approved by the Institutional
Animal Care and Use Committee of Central South University. The
acute stage and chronic stage are period from 24 h to 7 days, 50–60
days post-SE, respectively. In our study, day 7 and day 60 were
used to represent the corresponding acute and chronic stages. The
same amounts of experimental and control rats were sacrificed at
different time points, respectively, at day 7 and day 60 (n=9 in
each group at each time point).
Implantation of the mini drug delivery
system
Within 6 h after SE onset, the rats were
anesthetized and fixed into the stereotaxic apparatus. Each
mini-osmotic pump (ALZET® micro-osmotic pump, 1007D;
Durect Corporation, Cupertino, CA, USA) was attached to a cannula
and placed in a subcutaneous pocket. Subsequently, the cannula was
implanted into the lateral hippocampus (positions: −4.0 mm
posterior to bregma, 3.0 mm bilateral to the midline, and 3.1 mm
under the skull surface) with infusion of Mibefradil (1.5 mg/ml in
0.1 M PBS; Sigma-Aldrich) for seven consecutive days (19,20).
The cannula was secured with dental acrylic cement.
Behavioral recording
On day 60 post-SE, we recorded SRSs by using video
monitoring for 14 days from 8:00-10:00 a.m. to 4:00-6:00 p.m. each
day. The latency period (days), seizure frequency by cumulative
analysis over time (per day) and seizure duration (sec) were
recorded. During this period, animals had free access to food and
water as normal and no rats died. After behavioral observation, all
the rats were sacrificed under anesthesia for the next
experiments.
Implantation of EEG recording
electrodes and EEG analysis
In the chronic stage of SE, animals were
anesthetized using 10% chloral hydrate (3 ml/kg, i.p.), then fixed
into the stereotaxic apparatus. Electrodes, consisting of a
stainless steel wire were implanted into the positions: 2.5 mm
anterior to bregma, 2.5 mm bilateral to midline, and 2.5 mm
posterior to lambda and fixed with dental cement. After recovery,
spontaneous EEG seizures were recorded for 2 h to quantify the
frequency and mean duration during an EEG recording session. Data
were collected and analyzed using an acquisition system (Physical
Signal Recorder RM6240; Chengdu Instrument Factory, Chengdu,
China). The EEG data were analyzed by an experienced
neurologist.
Immunofluorescence staining
The rats were deeply anesthetized and transcardially
perfused with 4% paraformaldehyde. The brains were excised and
postfixed, then cut using a cryotome into serial coronal sections
(25 µm thick) (Leica CM 1850; Leica, Wetzler, Germany). Sections
were incubated in a blocking solution (10% normal donkey serum,
0.3% Triton X-100 in PBS, 2.5% bovine serum albumin in PBS) at room
temperature for 2 h. For DCX staining, the sections were washed and
incubated for 12 h at 4°C with goat anti-DCX antibody (diluted
1:125; Santa Cruz Biotechnology, Inc., Santa Cruz, CA) followed by
washing and incubation for 2 h in secondary antibody (diluted
1:1,000; Alexa Fluor 555 donkey anti-goat IgG; Invitrogen Life
Technologies, Carlsbad, CA, USA). For AnkG/DCX double staining,
sections were incubated in mouse anti-AnkG antibody (diluted 1:50;
Merck Millipore, Darmstadt, Germany) and goat anti-DCX antibody
(diluted 1:125; Santa Cruz Biotechnology, Inc.) at 4°C for 48 h.
Sections were rinsed and incubated with Alexa Fluor 488 donkey
anti-mouse IgG + Alexa Fluor 555 donkey anti-goat IgG (all diluted
1:1,000; Invitrogen Life Technologies). The sections were then
washed and mounted with Vectashield mounting media.
Quantification
Immunopositive cells were counted by an experimenter
blinded to the treatment conditions. The DG was operationally
defined as a region extending from the interface of the hilus and
granule cell layer to a width of approximately two cell bodies into
hilus. The immunofluorescence images of DCX-positive cells in DG
were acquired with a 20X objective (DM5000B light microscope;
Leica). For quantification analysis, sections were coded and the
captured images were analyzed using the ImageJ software package
(National Institutes of Health, Bethesda, MD, USA). Five
microscopic fields from the hippocampal DG regions were selected to
calculate neuronal counts in each view at ×40 magnifications. The
number of DCX immunopositive cells from five sections was counted
and a mean cell count per section was obtained.
Dendrite length was detected using a confocal
microscope (Carl Zeiss AG, Oberkochen, Germany) from five coronal
sections per rat. Three-dimensional reconstructions of the
DCX-positive cells were performed from Z-series stacks of confocal
images. Each section was scanned as a 0.5 µm thick Z-stack and
included 30 Z sections that contained the entire region of
DCX-positive. DCX-labeled cells were then counted by analyzing each
Z section and the reconstructed intact dendrite length were
measured by reconstructed Z-sections projected with ImageJ
software. The apical dendrite lengths of the DCX-positive cells
were traced and analyzed. For AIS analysis, the lengths and
position of AIS were measured from five coronal sections per rat
with using ImageJ software package.
Statistics
All continuous variables were tested to confirm that
they fit a normal distribution before further analysis. For
analysis of DCX-positive cells and dendrite length, a 3-way ANOVAs
followed by Tukey multiple comparison tests (α=0.05) was used to
calculate the difference between any two groups of rats of
different stages. For analysis of AIS changes per microscopic view,
3-way ANOVAs followed by Tukey multiple comparison tests (α=0.05)
to was used to compare between-group difference in rats of
different stages. For analysis of EEG data, one-factor ANOVA
followed by Tukey multiple comparison tests (α=0.05) to was used to
compare between-group difference in rats. Results are presented
throughout as mean ± standard deviation. P<0.05 was considered
to indicate a statistically significant difference. three-way
ANOVAs were performed using SPSS 21.0 (SPSS, Inc, Chicago, IL,
USA); all other statistical analyses were performed using GraphPad
Prism 6.0 (GraphPad Software, Inc., La Jolla, CA, USA).
Results
Proliferation of newborn neurons in
pilocarpine-induced epileptic rats
The birth of new neurons in DG area of hippocampus
progresses from progenitor cells expressed DCX to mature neurons
expressed the marker NeuN. Double immunofluorescence staining of
DCX and NeuN were used to evaluate the newborn neurons in
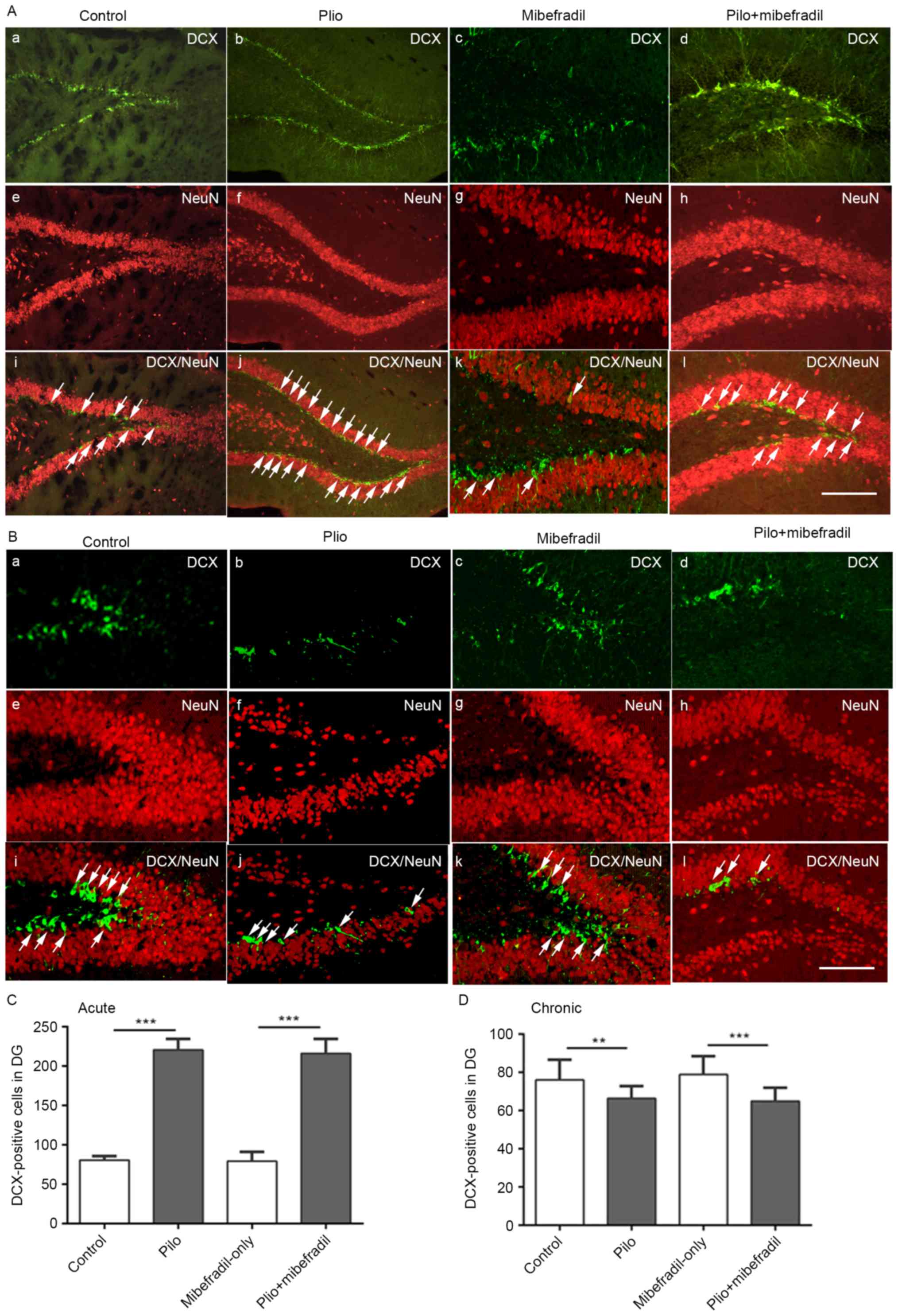
hippocampus of rats; typical images are showed in Fig. 1. Compared with controls, SE rats
showed significantly increased number of newborn neurons in the DG
of hippocampus on day 7 (ANOVA with post hoc Tukey's test,
DCX/NeuN-positive cells: 220±13.9 vs. 80.7±5.0, n=18 per group,
P<0.001). In epileptic rats on day 60, the number of newborn
neurons in the entire DG in SE rats appeared to decline compared to
that of control rats (ANOVA with post hoc Tukey's test,
DCX/NeuN-positive cells: 76.0±10.7 vs. 66.4±6.5, n=18 per group,
P=0.004).
To clarify the function of AIS plasticity of newborn
neurons in epileptogenesis, we used T/L-type calcium blocker
Mibefradil treatment using intrahippocampal infusion. Quantitative
analysis of the DCX staining was conducted to assess the
neurogenesis in hippocampus, as showed in Fig. 1. Compared with Mibefradil-only
rats, Mibefradil treatment after pilocarpine-induced epileptic rats
showed DCX-positive cells increase in both days 7 and 60 (ANOVA
with post hoc Tukey's test, 216.2±18.0 vs. 79.5±11.7 on day 7;
64.9±7.0 vs. 78.9±9.6 on day 60, all P<0.001). However, no
significant differences were observed for Mibefradil-only and
control groups (ANOVA with post hoc Tukey's test, 79.5±11.7 vs.
80.7±5.0 on day 7; 78.9±9.6 vs. 76.0±10.7 on day 60, n=18 per
group, all P>0.05), Mibefradil treatment after SE rats and
epileptic rats on both day 7 and day 60 (216.2±18.2 vs. 220±13.9 on
day 7, P=0.71; 64.9±7.0 vs. 66.4±6.5 on day 60, P=0.95; n=18 per
group).
Dendrite development of newborn
neurons in pilocarpine-induced epileptic rats
Also, morphological features of DCX-immunolabeled
cells in the DG from epileptic rats were different from those
observed from controls. Compared to control rats, epileptic rats
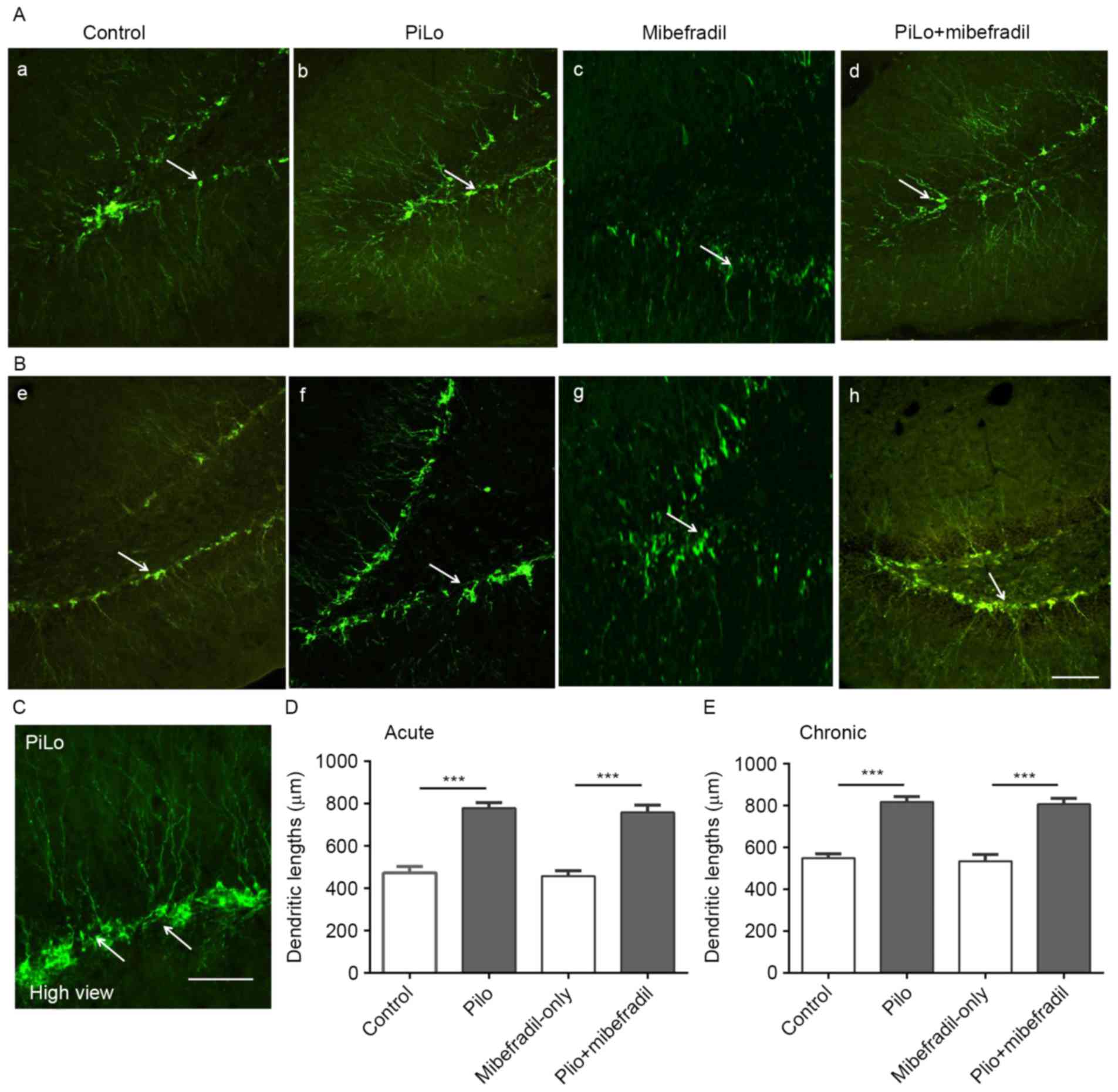
showed significantly increased apical dendrites lengths in newborn
neurons on both days 7 and 60 (ANOVA with post hoc Tukey's test,
472.7±30.5 vs. 779.7±25.7 µm on day 7; 548.6±20.6 vs. 817.1±24.9 µm
on day 60, n=18 per group, all P<0.001) (Fig. 2), suggesting that
pilocarpine-induced epileptic rats exhibit longer and extended
apical dendrites in hippocampal newborn neurons. Furthermore, the
increased hippocampal newborn neurons in acute stage post-SE in our
study are consistent with the report that acute seizures mainly
prompted the proliferation of cells expressing DCX (21). These results indicate that these
newborn neurons with longer, extended apical dendrites could
facilitate reception of more excessive, recurrent synaptic input
and then contribute to an increase in the excitability of
hippocampal network after seizures.
Compared with Mibefradil-only rats, Mibefradil
treatment rats showed significantly increased in the lengths of
apical dendrites in newborn neurons on both days 7 and 60 (ANOVA
with post hoc Tukey's test, 758.6±35.0 vs. 458.0±25.9 µm on day 7,
806.6±27.6 vs. 534.1±32.6 µm on day 60, n=18 per group all
P<0.001) (Fig. 2). However, no
significant differences in the apical dendrites lengths were found
in newborn neurons between the Mibefradil treatment rats and
epileptic rats (ANOVA with post hoc Tukey's test, 758.6±35.0 vs.
779.7±25.7 µm on day 7, P=0.12; 806.6±27.6 vs. 817.1±24.9 µm on day
60, P=0.61; n=18 per group), Mibefradil-only and control groups
(458.0±25.9 vs. 472.7±30.5 µm on day 7, P=0.40; 534.1±32.6 vs.
548.6±20.6 µm on day 60, P=0.33; n=18 per group). These results
indicate that Mibefradil had less effect on the newborn neurons
number and morphology in epileptic rats.
AIS plasticity of hippocampal newborn
neurons in pilocarpine-induced epileptic rats
Double-immunofluorescence method with AnkG and DCX
was used to reveal structural plasticity of the AIS in newborn
neurons in epileptic rats; representative images are showed in
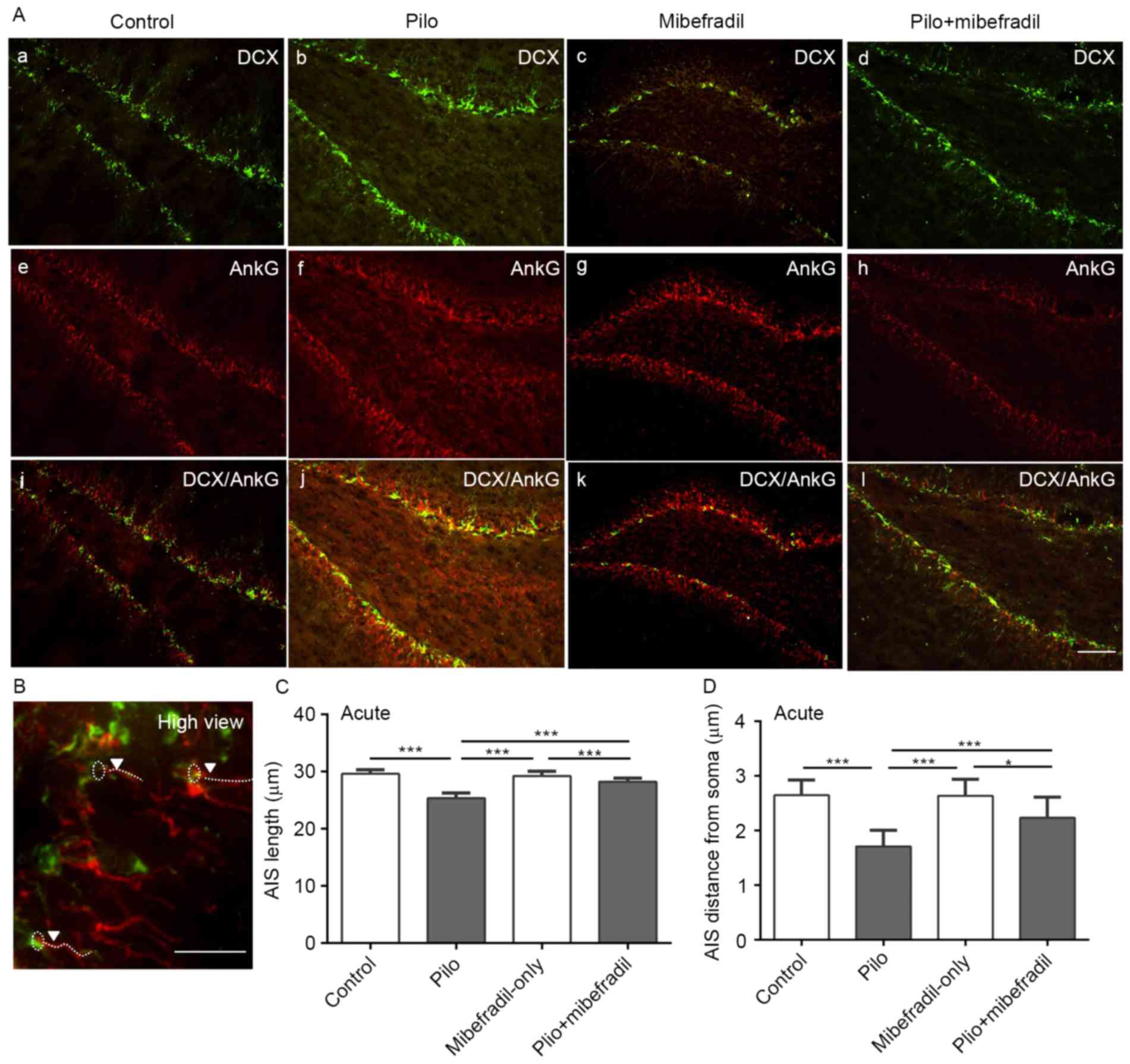
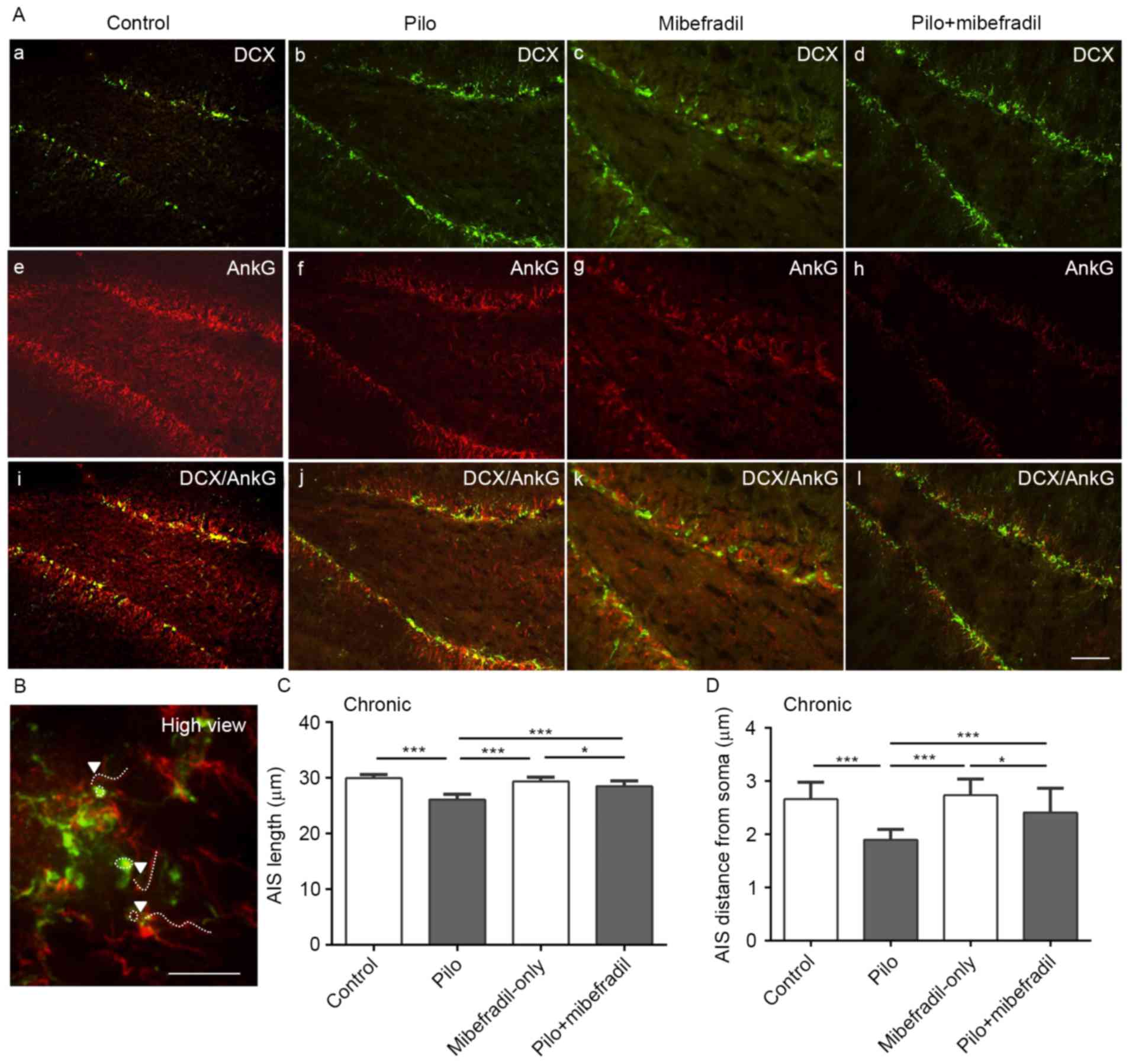
Figs. 3 and 4. Compared with the control rats, rats
subjected to pilocarpine showed reduced lengths of AIS in newborn
neurons of hippocampus on both days 7 and 60 (ANOVA with post hoc
Tukey's test, 29.6±0.7 vs. 25.3±0.9 µm on day 7; 29.9±0.7 vs.
26.1±1.0 µm on day 60, n=18 per group, all P<0.001). To further
analyze the position of AIS in newborn neuron, we measured the
distance of AIS from the soma. As showed in Figs. 3A and 4A, the distance of AIS from the soma was
significantly decreased in epileptic rats compared to the control
rats in both days 7 and 60 (ANOVA with post hoc Tukey's test,
2.6±0.3 vs. 1.7±0.3 µm on day 7; 2.6±0.3 vs. 1.9±0.2 µm on day 60,
n=18 per group, all P<0.001). All results indicate that the
plasticity of AIS, reflecting on the decreases in the length of AIS
and shortening of the position of AIS from soma during both acute
and chronic status epileptics, is probably triggered by the altered
environment induced by epileptic seizures.
Interestingly, Mibefradil treatment resulted in
increases in the lengths of AIS in newborn neurons compared with
the epileptic controls on both post-SE days 7 and 60, (ANOVA with
post hoc Tukey's test, 28.3±0.59 vs. 25.3±0.95 µm on day 7;
28.6±0.93 vs. 26.1±0.95 µm on day 60, n=18, all P<0.001).
Comparisons of AIS location from the soma in newborn neurons showed
greater distance increases in the epileptic rats treated with
Mibefradil on both post-SE days 7 and 60 than epileptic rats (ANOVA
with post hoc Tukey's test, 2.24±0.38 vs. 1.71±0.30 µm on day 7;
2.41±0.46 vs. 1.90±0.19 µm on day 60, n=18 per group, all
P<0.001) (Figs. 3A and 4). However, Mibefradil treatment showed
significantly differences in both length and distance of AIS in
newborn neurons compared with Mibefradil-only group (length:
29.3±0.79 vs. 28.3±0.59 µm on day 7, P=0.003; 29.3±0.80 vs.
28.6±0.93 µm on day 60, P=0.02; distance: 2.64±0.30 vs. 2.24±0.38
µm on day 7, P<0.001; 2.74±0.31 vs. 2.41±0.46 µm on day 60,
P=0.02; n=18 per group), suggesting of no notable AIS changes in
Mibefradil-only rats. These results indicated that Mibefradil
treatment can effectively reverse the movement of AIS post-SE,
though the changes in aberrant neurogenesis and dendrites were not
seen.
The effect of mibefradil on behavioral
outcomes and spontaneous EEG seizures in chronic TLE
In the present study, pilocarpine-induced seizures
were characterized by brief tonic-clonic episodes with forelimb
automatisms with rearing and falling, which is similar to the
descriptions from previous reports (22). No seizures were observed in control
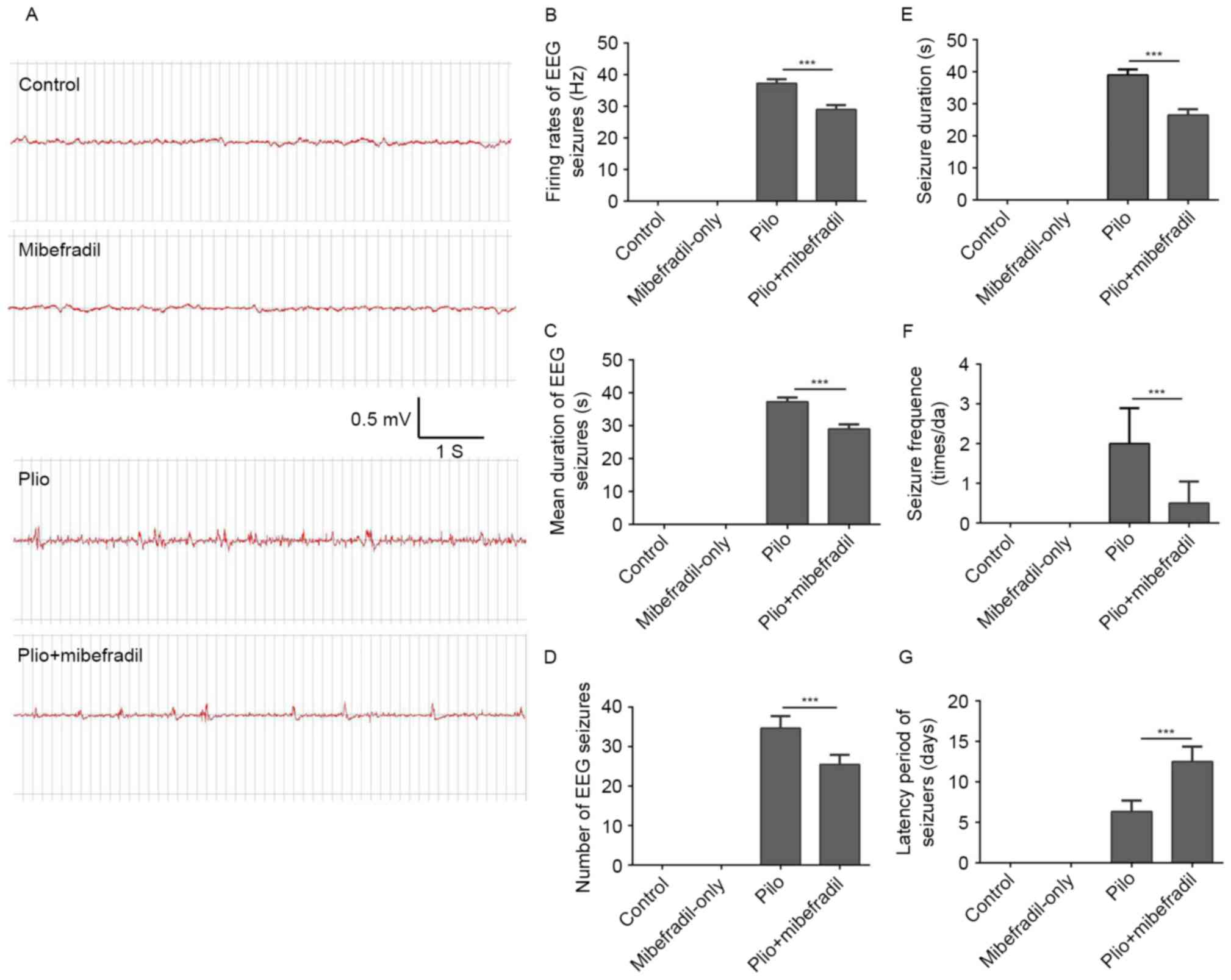
and Mibefradil-only groups. Data in Fig. 5E concerning the behaviors showed
that Mibefradil treatment substantially shortened the mean duration
of spontaneous seizures compared with the pilocarpine group (ANOVA
with post hoc Tukey's test, 26.5±1.9 vs. 39.0±1.8 s, n=3 per group,
P<0.001). In addition, the average latency periods were longer
and the frequency of spontaneous seizures was significantly reduced
in epileptic rats with Mibefradil treatment compared with the
epileptic controls (ANOVA with post hoc Tukey's test, 12.5±1.9 vs.
6.3±1.4 days, n=3 per group, P<0.001) (Fig. 5F and G).
The spontaneous seizures measured by EEG recordings
showed that epileptic rats promoted the development of SRS with
characteristic spike and wave activity of EEG in the chronic phase
of TLE. No EEG seizures were observed in control rats and rats
receiving Mibefradil-only. Spontaneous EEG seizures were frequently
observed in rats subjected to pilocarpine; typical EEG images are
showed in Fig. 4A. During the 2 h
EEG recording, Mibefradil treatment rats showed significantly
reduced firing rates and shortened the mean duration of spontaneous
seizures compared with the pilocarpine rats (ANOVA with post hoc
Tukey's test, firing rates: 29.9±1.4 vs. 37.3±1.2 Hz, P<0.001;
duration: 2.4±0.19 vs. 5.4±0.19 sec, P<0.001; n=3 per group)
(Fig. 5B and C). In addition,
compared with the epileptic rats, epileptic rats with Mibefradil
treatment showed reduced frequency of spontaneous EEG seizures
(ANOVA with post hoc Tukey's test, 25.5±2.4 vs. 34.7±3.1, n=3,
P<0.001) (Fig. 5D). All results
indicate that Mibefradil treatment rescued the AIS movement and
alleviated the spontaneous seizures in epileptic rats, suggesting
that Mibefradil treatment can fine-tune the neuron excitability and
firing behavior after epilepsy.
Discussion
In the present study, we investigated features of
hippocampal neurogenesis and AIS plasticity in newborn neurons in
both acute and chronic phases of the experimental epileptic rats.
The pilocarpine-induced rats of TLE resulted in increased
proliferation and abnormal dendrite development of newborn neurons.
More importantly, we found a significant decrease in AIS length and
distance in hippocampal neurogenesis in pilocarpine-induced
epileptic rats, with increased spontaneous seizures observed in
both behavioral and EEG recordings. After Mibefradil treatments,
AIS length and distance tend to increase in epileptic rats, but no
significant changes were observed in Mibefradil-only rats. By
comparing the EEG and behaviors in Mibefradil treatment rats and
epileptic rats, Mibefradil treatment rats were distinguished from
non-treatment rats with decreased seizure activity. These results
suggest that the AIS plasticity in hippocampal newly-generated
neurons is associated with hyperexcitability of newborn neurons in
pilocarpine-induced rats of TLE. To our knowledge, this is the
first study that details the changes of AIS movement in hippocampal
neurogenesis in epilepsy and explores their relationship with the
development of recurrent seizures.
Growing evidence has showed that epileptic animals
often experience the proliferation and differentiation of the
neural progenitor cells in the DG area of hippocampus, as well as
changes of neuronal excitability (23,24).
This dramatic increase in the production of new neurons in the DG
following SE can contribute to, rather than counteract, abnormal
brain activity (1,25). These newly-generated neurons
developed abnormal dendrites which can facilitate reception of more
excessive, recurrent synaptic input by aberrantly sprouted mossy
fibers, and then contribute to an increase in the excitability of
the hippocampal network after seizures (26,27).
Elucidation of this hyperexcitability of hippocampal network can
facilitate the SRS mechanism in TLE. In this study, epileptic rats
showed increased hippocampal neurogenesis and abnormal dendrite
development of newborn neurons in the acute stage of epileptic
models and subsequent reduction in the chronic stage of TLE. Also,
seizures were always observed in chronic stage of both epileptic
patients and animals. These results suggested that changes in
hippocampal newly-generated neurons may be closely related to the
increased neuronal excitability in the recurrence of seizure
activity in the TLE. Our results are consistent with early studies
showing abnormal hippocampal neurogenesis in epileptic rats
(28); however, the find that
newborn neurons plasticity and excitability has not been reported
previously.
Besides detailing changes in newborn neurons in
hippocampus, we also explored the changes of neuronal excitability
domain, AIS in hippocampal new-generated neurons. AIS is a
specialized domain of the axon proximal to the soma and can play a
major role in neuronal excitability (29,30).
It is widely accepted that the structural characteristics of the
AIS such as length and position relative to soma, can contribute to
homeostatic control of excitability (31). This structural plasticity of AIS
may regulate the excitability of a neuron in response to its
environment, either attenuating or enhancing a neuron's sensitivity
(32,33). Our study revealed TLE rats showed
decreased length and distance to the soma in AIS of hippocampal
newborn neurons in both acute and chronic stages post-SE. These
findings are consistent with previous study of AIS in motor-sensory
cortical neurons in malformations of cortical development and
epileptic rats (34). However, it
must be noted that AIS plasticity in this study was from
hippocampal newborn neurons in epileptic rats in which AIS length
and distance were notably decreased compared with controls. Thus,
it is required to learn the hippocampal newborn neurons when study
the mechanism of epilepsy.
There are several causes explaining the changes of
AIS plasticity observed in epileptic rats. The initial decrease in
AIS length of hippocampal newborn neurons could be triggered by the
proteolysis of the AnkG introduced by calcium-dependent cysteine
protease calpain in the context of injury (35). Because the proteolysis of AIS
cytoskeletal proteins are rapid and irreversible (36), the AIS cytoskeleton disrupts as we
observed in epileptic rats in both acute and chronic stages. For
another, change in AIS location is related to the extent of its
isolation from the soma and a greater distance can increase the
isolation (29). When the AIS is
located proximally from the soma, the AIS becomes more excitable
because the charge dissipation decreases, making it easy to
depolarize the AIS above the action potential threshold in the
presence of the shunting conductance. Such a finding is consistent
with the reports by Harty et al (13), who showed that AIS is closer to the
soma in pyramidal neurons in deep layer 5 of somatosensory cortex
in amygdala kindling epileptic rats.
However, the changes of AIS plasticity in both
length and distance as we observed in epileptic rats can mutually
influence the neuronal excitability. For example, Baranauskas et
al examined the AIS plasticity in cortical pyramidal neurons,
finding that AIS length and location depend on the balance of its
depolarizing accessibility and electrical isolation from
somatodendritic conductance loads (37). Also, Gulledge and Bravo reported
when AIS was shorter or adjacent to the soma in the dentate granule
neuron, neuronal excitability was highest (38). Because ion channels in the AIS
arrangement play distinct roles in action potential initiation, AIS
plasticity may primarily through changes in AIS length to fine-tune
excitability at relatively slow timescales after stimuli (39). Our study also revealed that the
pilocapine-induced epileptic rats showed significantly decreased
AIS lengths but only within a few micrometers in AIS locations.
AIS is enriched with multiple types of ion channels,
including Na+, K+ and more important,
Ca+ channels. The Ca+ channels in AIS are essential for
action potential initiation (40);
thus it is practical to explore the calcium-current changes in AIS
using the calcium-current blocker. Consistent with our findings,
another study also showed a blocker Mibefradil could effectively
suppress the AIS movement in hippocampal neurons in vitro
(17), and imply that AIS
plasticity in hippocampal newly-generated neuron can be a target
excitable site for recurrent of epilepsy. However, our study also
revealed the epilepsy network and long-term seizure activity
attenuated in epileptic rats after Mibefradil treatment, supporting
the contribution of AIS to neuronal excitability in epilepsy. These
results suggested that AIS plasticity of newborn neurons could
increase neuronal excitability after pilocarpine-induced TLE rats,
which may contribute to chronic epilepsy; this is congruent between
in vitro and in vivo and provides the basis for
studying the AIS plasticity occurring in human using animals.
We acknowledge that this study has some limitations.
First, we did not perform electrophysiological studies on
hippocampal newborn neurons in vitro to further verify the
changes in AIS plasticity on newly-generated neurons. Further
studies will need to clarify the details of changes in ion channels
at the AIS such as using a novel method of axon-bleb patch-clamp
together with somatic nucleated patch recording from hippocampal
newborn neurons. Second, we cannot rule out the possibility that
the treatment of Mibefradil can also affect other types of neurons
other than newborn neurons. Moreover, not only the T/L-type calcium
blocker Mibefradil, but also the L-type calcium currents blocker
nifedipine can abolish activity-dependent AIS movement (17). It may require more specific
research methods to distinguish the difference between the two
types of calcium currents on AIS plasticity. Finally, it will be
helpful to attempt other genetic engineering methods to explore
possible related micro RNAs to further regulate the AIS expression
and movement in epileptic animal or cultured hippocampal newborn
neurons and then explore the pathophysiological mechanisms of the
AIS plasticity in epilepsy.
In conclusion, our study confirmed the aberrant
hippocampal neurogenesis together with prolonged plasticity of AIS
in hippocampal newborn neurons in pilocarpine-induced TLE rats.
Mibefradil treatment effectively suppressed the movement of AIS in
hippocampal newborn neurons which was accompanied by decreased
long-term seizure activity. These new findings have undoubtedly
provided experimental evidence that AIS plasticity in newborn
neurons play a key role in epilepsy and this might be a potential
therapeutic target to help control seizure activities.
Acknowledgements
This study was supported by National Natural Science
Foundation of China (nos. 81100967 and 81371435).
Glossary
Abbreviations
Abbreviations:
|
TLE
|
temporal lobe epilepsy
|
|
AIS
|
axon initial segment
|
|
SE
|
status epilepticus
|
|
DCX
|
doublecortin
|
|
AnkG
|
ankyrin G
|
|
DG
|
dentate gyrus
|
|
SRS
|
spontaneous recurrent seizures
|
References
|
1
|
Murphy BL, Hofacer RD, Faulkner CN, Loepke
AW and Danzer SC: Abnormalities of granule cell dendritic structure
are a prominent feature of the intrahippocampal kainic acid model
of epilepsy despite reduced postinjury neurogenesis. Epilepsia.
53:908–921. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Overstreet-Wadiche LS, Bromberg DA, Bensen
AL and Westbrook GL: Seizures accelerate functional integration of
adult-generated granule cells. J Neurosci. 26:4095–4103. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shapiro LA and Ribak CE: Newly born
dentate granule neurons after pilocarpine-induced epilepsy have
hilar basal dendrites with immature synapses. Epilepsy Res.
69:53–66. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang LP, Kempermann G and Kettenmann H: A
subpopulation of precursor cells in the mouse dentate gyrus
receives synaptic GABAergic input. Mol Cell Neurosci. 29:181–189.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bonzano S and De Marchis S: Detecting
neuronal differentiation markers in newborn cells of the adult
brain. Methods Mol Biol. 1560:163–177. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Erdő F, Denes L and de Lange E:
Age-associated physiological and pathological changes at the
blood-brain barrier: A review. J Cereb Blood Flow Metab. 37:4–24.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
von Bohlen und Halbach O:
Immunohistological markers for proliferative events, gliogenesis,
and neurogenesis within the adult hippocampus. Cell Tissue Res.
345:1–19. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jessberger S, Römer B, Babu H and
Kempermann G: Seizures induce proliferation and dispersion of
doublecortin-positive hippocampal progenitor cells. Exp Neurol.
196:342–351. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Corbo JC, Deuel TA, Long JM, LaPorte P,
Tsai E, Wynshaw-Boris A and Walsh CA: Doublecortin is required in
mice for lamination of the hippocampus but not the neocortex. J
Neurosci. 22:7548–7557. 2002.PubMed/NCBI
|
|
10
|
Kim JH, Jang BG, Choi BY, Kwon LM, Sohn M,
Song HK and Suh SW: Zinc chelation reduces hippocampal neurogenesis
after pilocarpine-induced seizure. PLoS One. 7:e485432012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Petersen AV, Cotel F and Perrier JF:
Plasticity of the axon initial segment: Fast and slow processes
with multiple functional roles. Neuroscientist. May 3–2016.(Epub
ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hamada MS and Kole MH: Myelin loss and
axonal ion channel adaptations associated with gray matter neuronal
hyperexcitability. J Neurosci. 35:7272–7286. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Harty RC, Kim TH, Thomas EA, Cardamone L,
Jones NC, Petrou S and Wimmer VC: Axon initial segment structural
plasticity in animal models of genetic and acquired epilepsy.
Epilepsy Res. 105:272–279. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hedstrom KL, Ogawa Y and Rasband MN:
AnkyrinG is required for maintenance of the axon initial segment
and neuronal polarity. J Cell Biol. 183:635–640. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Evans MD, Dumitrescu AS, Kruijssen DL,
Taylor SE and Grubb MS: Rapid modulation of axon initial segment
length influences repetitive spike firing. Cell Rep. 13:1233–1245.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kole MH and Stuart GJ: Signal processing
in the axon initial segment. Neuron. 73:235–247. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Grubb MS and Burrone J: Activity-dependent
relocation of the axon initial segment fine-tunes neuronal
excitability. Nature. 465:1070–1074. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Racine RJ: Modification of seizure
activity by electrical stimulation: Cortical areas.
Electroencephalogr Clin Neurophysiol. 38:1–12. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen YL, Tsaur ML, Wang SW, Wang TY, Hung
YC, Lin CS, Chang YF, Wang YC, Shiue SJ and Cheng JK: Chronic
intrathecal infusion of mibefradil, ethosuximide and nickel
attenuates nerve ligation-induced pain in rats. Br J Anaesth.
115:105–111. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee TS, Kaku T, Takebayashi S, Uchino T,
Miyamoto S, Hadama T, Perez-Reyes E and Ono K: Actions of
mibefradil, efonidipine and nifedipine block of recombinant T- and
L-type Ca channels with distinct inhibitory mechanisms.
Pharmacology. 78:11–20. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Parent JM, Tada E, Fike JR and Lowenstein
DH: Inhibition of dentate granule cell neurogenesis with brain
irradiation does not prevent seizure-induced mossy fiber synaptic
reorganization in the rat. J Neurosci. 19:4508–4519.
1999.PubMed/NCBI
|
|
22
|
Alam AM and Starr MS: Regional changes in
brain dopamine utilization during status epilepticus in the rat
induced by systemic pilocarpine and intrahippocampal carbachol.
Neuropharmacology. 35:159–167. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shu Y, Xiao B, Wu Q, Liu T, Du Y, Tang H,
Chen S, Feng L, Long L and Li Y: The Ephrin-A5/EphA4 interaction
modulates neurogenesis and angiogenesis by the p-Akt and p-ERK
pathways in a mouse model of TLE. Mol Neurobiol. 53:561–576. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bender RA, Dubé C, Gonzalez-Vega R, Mina
EW and Baram TZ: Mossy fiber plasticity and enhanced hippocampal
excitability, without hippocampal cell loss or altered
neurogenesis, in an animal model of prolonged febrile seizures.
Hippocampus. 13:399–412. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Parent JM, Yu TW, Leibowitz RT, Geschwind
DH, Sloviter RS and Lowenstein DH: Dentate granule cell
neurogenesis is increased by seizures and contributes to aberrant
network reorganization in the adult rat hippocampus. J Neurosci.
17:3727–3738. 1997.PubMed/NCBI
|
|
26
|
Kron MM, Zhang H and Parent JM: The
developmental stage of dentate granule cells dictates their
contribution to seizure-induced plasticity. J Neurosci.
30:2051–2059. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ribak CE, Tran PH, Spigelman I, Okazaki MM
and Nadler JV: Status epilepticus-induced hilar basal dendrites on
rodent granule cells contribute to recurrent excitatory circuitry.
J Comp Neurol. 428:240–253. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Song C, Xu W, Zhang X, Wang S, Zhu G, Xiao
T, Zhao M and Zhao C: CXCR4 Antagonist AMD3100 Suppresses the
long-term abnormal structural changes of newborn neurons in the
Intraventricular kainic acid model of epilepsy. Mol Neurobiol.
53:1518–1532. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yamada R and Kuba H: Structural and
functional plasticity at the axon initial segment. Front Cell
Neurosci. 10:2502016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Grubb MS, Shu Y, Kuba H, Rasband MN,
Wimmer VC and Bender KJ: Short- and long-term plasticity at the
axon initial segment. J Neurosci. 31:16049–16055. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kole MH, Letzkus JJ and Stuart GJ: Axon
initial segment Kv1 channels control axonal action potential
waveform and synaptic efficacy. Neuron. 55:633–647. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Grubb MS and Burrone J: Building and
maintaining the axon initial segment. Curr Opin Neurobiol.
20:481–488. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kuba H, Oichi Y and Ohmori H: Presynaptic
activity regulates Na(+) channel distribution at the axon initial
segment. Nature. 465:1075–1078. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang Y, Sun D, Yue Z, Tang W, Xiao B and
Feng L: Rats with malformations of cortical development exhibit
decreased length of AIS and hypersensitivity to Pilocarpine-induced
status epilepticus. Neurochem Res. 41:2215–2222. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Evans MD, Sammons RP, Lebron S, Dumitrescu
AS, Watkins TB, Uebele VN, Renger JJ and Grubb MS: Calcineurin
signaling mediates activity-dependent relocation of the axon
initial segment. J Neurosci. 33:6950–6963. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Schafer DP, Jha S, Liu F, Akella T,
McCullough LD and Rasband MN: Disruption of the axon initial
segment cytoskeleton is a new mechanism for neuronal injury. J
Neurosci. 29:13242–13254. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Baranauskas G, David Y and Fleidervish IA:
Spatial mismatch between the Na+ flux and spike initiation in axon
initial segment. Proc Natl Acad Sci USA. 110:4051–4056. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gulledge AT and Bravo JJ: Neuron
morphology influences axon initial segment plasticity. eNeuro.
3:pii: ENEURO.0085–15.2016. 2016. View Article : Google Scholar
|
|
39
|
Gutzmann A, Ergül N, Grossmann R, Schultz
C, Wahle P and Engelhardt M: A period of structural plasticity at
the axon initial segment in developing visual cortex. Front
Neuroanat. 8:112014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Jones SL and Svitkina TM: Axon initial
segment cytoskeleton: Architecture, development, and role in neuron
polarity. Neural Plast. 2016:68082932016. View Article : Google Scholar : PubMed/NCBI
|