Introduction
Tumors are associated with high incidence rates and
cannot be easily treated. Currently available treatment strategies
that are used in clinical practice include surgery, chemotherapy,
radiotherapy, targeted drug therapy and gene therapy. Although
numerous novel antitumor drugs have been developed, these drugs
inevitably injure healthy tissues and cells, thus inducing severe
adverse reactions that greatly affect clinical use (1). Therefore, it is of great significance
to identify novel antitumor agents characterized by high
pharmacological efficiency, few adverse events and low toxicity,
and high specificity.
As a semi-synthetic camptothecin derivative,
irinotecan overcomes low aqueous solubility, promotes
gastrointestinal absorption and exerts evident therapeutic effects
on various tumors, including colorectal cancer, gastric cancer,
cervical cancer, ovarian cancer, lung cancer and esophageal cancer
(2–5). In addition, irinotecan induces
cytotoxicity predominantly by affecting DNA topoisomerase I (DNA
Topo I), which participates in intracellular DNA replication,
transcription, recombination and repair. This enzyme resolves
topological restrictions inside DNA molecules and relaxes
excessively coiled DNA double strands, in order to maintain normal
DNA replication and transcription (6). Irinotecan only works after being
catalyzed by carboxylesterase, when it is converted into the active
SN-38 form; however, the conversion process is characterized by low
efficiency. Furthermore, it strongly inhibits acetylcholinesterase
(AchE) activity. Long-term or high-dose use of irinotecan may
trigger gastrointestinal reactions, liver and kidney toxicity, and
bone marrow suppression (7,8). To
reduce such adverse reactions, it is of great significance to
structurally modify irinotecan. We previously synthesized a series
of irinotecan derivatives by analyzing the structure-property
relationships of camptothecin derivatives and selecting proper
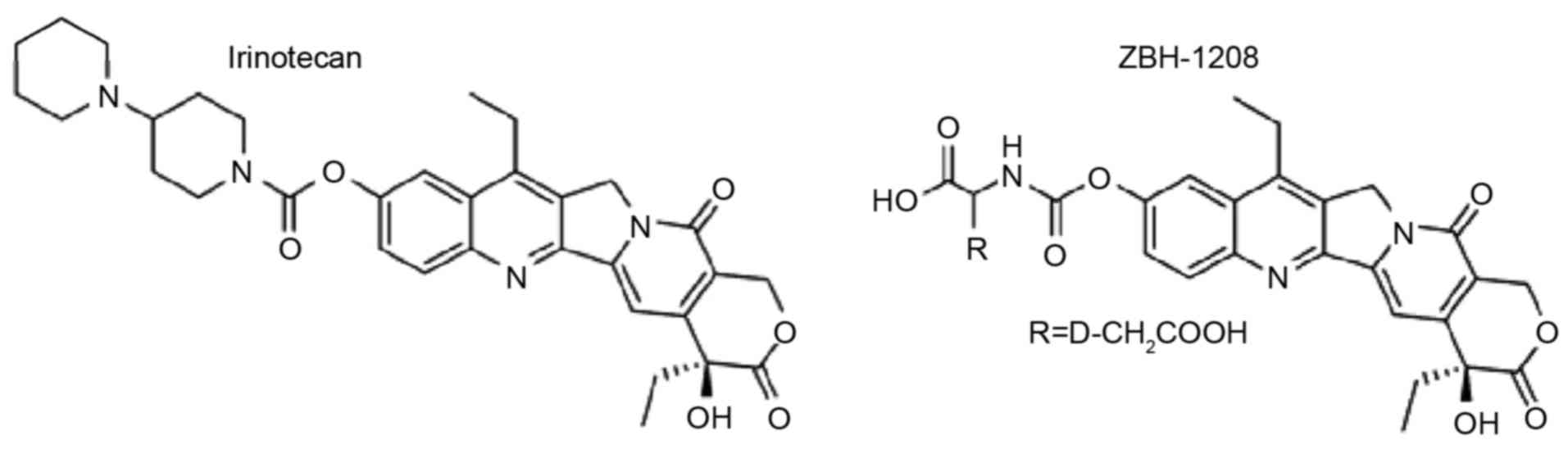
carriers (unpublished data). The screened compound, ZBH-1208
(Fig. 1), exhibits higher in
vitro antitumor activity and reduced AchE-inhibiting activity
compared with irinotecan (9).
Therefore, it is necessary to study the antitumor activity and the
underlying mechanisms of ZBH-1208, in order to develop more
eligible drugs. The present study aimed to evaluate the inhibitory
effects of ZBH-1208 on B22 mouse brain tumor xenografts, and to
clarify the underlying molecular mechanism.
Materials and methods
Drugs and reagents
ZBH-1208 was provided by the Institute of
Pharmacology and Toxicology of Academy of Military Medical Sciences
(Beijing, China). Irinotecan was purchased from Shandong Qilu
Pharmaceutical Co., Ltd. (Jinan, China; National Medicine Permit
no. H20068128). Sodium chloride was obtained from Sichuan Kelun
Pharmaceutical Co., Ltd. (National Medicine Permit no. H20056626;
Sichuan, China). TRIzol reagent, bicinchoninic acid (BCA) protein
quantification kit and enhanced chemiluminescent (ECL) reagent were
purchased from Thermo Fisher Scientific, Inc. (Waltham, MA, USA).
Primers were designed by and purchased from Shanghai Generay
Biotech Co., Ltd. (Shanghai, China). Rabbit anti-mouse p53 (cat.
no. sc-126), rabbit anti-mouse p21 (cat. no. sc-6246), rabbit
anti-mouse cyclin B1 (cat. no. sc-245), mouse anti-human cyclin E
(cat. no. sc-481), rabbit anti-mouse cell division cycle 2 (CDC2)
(cat. no. sc-54), rabbit anti-mouse phosphorylated (p)-CDC2 (Tyr15)
(cat. no. sc-12340), rabbit anti-mouse p-CDC2 (Thr161) (cat. no.
sc-12341), rabbit anti-mouse cyclin-dependent kinase 2 (CDK2) (cat.
no. sc-6248), rabbit anti-mouse CDK7 (cat. no. sc-56284), rabbit
anti-mouse Wee1 (cat. no. sc-5285) and rabbit anti-mouse CDC25C
(cat. no. sc-327) antibodies, and horseradish peroxidase-labeled
goat anti-rabbit (cat. no. sc-2030) and goat anti-mouse
immunoglobulin G (cat. no. sc-2005) secondary antibodies were
purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX,
USA).
Apparatus
The 7700 fluorescence quantitative polymerase chain
reaction (qPCR) system was purchased from Applied Biosystems
(Thermo Fisher Scientific, Inc.). Medonic CA620 hematology analyzer
was obtained from Boule Medical AB (Spånga, Sweden). High-speed
refrigerated centrifuge was purchased from Heraeus (Hanau,
Germany), and centrifuge was obtained from Eppendorf AG (Hamburg,
Germany).
Animals
Specific pathogen-free-grade male ICR mice (age,
56–70 days; weight, 18–22 g) were purchased from the Experimental
Animal Center of Peking University Health Science Center (Beijing,
China; certificate of conformity: SCXK (Beijing) 2006–0008). Mice
were maintained at a temperature of 20–22°C, humidity of 40–60%
under a 12-h light/dark cycle with free access to food and water.
The present study has been approved by the Ethics Committee of
Liaocheng People's Hospital (Liaocheng, China), and efforts were
made to minimize the suffering of animals.
Cell line
The B22 mouse brain tumor cell line was provided by
the China Center for Type Culture Collection (Wuhan, China). Cells
were cultured in Dulbecco's modified Eagle's medium supplemented
with 10% fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.)
in a humidified 5% CO2 atmosphere at 37°C. Cells were
inoculated into 30 ICR mice with a 2 ml intraperitoneal injection
of 1×106 cells/ml.
Animal grouping and treatment
Under sterile conditions, B22 tumor tissues, which
had been subcutaneously inoculated into the right axillary region
for 2 weeks, were collected, cut into small pieces, ground,
screened and centrifuged at 1,200 × g for 5 min at 4°C.
Subsequently, tumor cells were resuspended by adding 5X volume of
normal saline and the cells (1×106 cells/ml) were
subcutaneously inoculated into the right axillary region of male
ICR mice (0.2 ml/mouse). On the subsequent day, mice were randomly
divided into the negative control (normal saline) group (i.p., once
daily), the low-dose and high-dose ZBH-1208 administration groups
(20 and 40 mg/kg, respectively; i.p., once daily) and the positive
control (irinotecan) group (25 mg/kg; i.p., once daily) (n=8
mice/group). The irinotecan and low-dose ZBH-1208 groups were
administered freshly prepared solutions at the same molar
concentration. The mice were weighed daily and sacrificed by
cervical dislocation following 12 days of treatment, after
anesthesia with 1.5% pentobarbital sodium (35 mg/kg) intravenously.
The solid tumors were weighed; tumor inhibition rate was calculated
as follows: Tumor inhibition rate (%) = (Average tumor weight of
negative control group - average tumor weight of administration
group)/average tumor weight of negative control group × 100%.
Weight difference was calculated, as follows: Weight difference =
Weight on the last day - weight on the first day.
Measurement of visceral indices and
counting of white blood cells
Prior to sacrifice of tumor-bearing mice by cervical
dislocation, 2–5 ml peripheral blood was collected from the orbital
sinus to evaluate the effects of ZBH-1208 on mouse white blood cell
counts using the Medonic CA620 hemocytometer. After the mice were
sacrificed, spleen and thymus gland tissues were collected, in
order to calculate the visceral index, as follows: Visceral index
(mg/g) = Visceral weight/body weight.
Reverse transcription-qPCR
(RT-qPCR)
Total RNA was extracted from peripheral blood cells
using TRIzol, and its integrity was identified by electrophoresis.
Subsequently, 1 µg total RNA was reverse-transcribed into cDNA
using the QuantiTect Reverse Transcription kit (Qiagen, Inc.,
Valencia, CA, USA), as previously described (10). The total reaction volume was 10 µl.
qPCR was performed using the 7700 fluorescent qPCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). Primer sequences were
as follows: p53 sense, 5′-TACTCCCCTGCCCTCAACAAGA-3′ and antisense,
5′-ACAACCTCCGTCATGTGCTGTG-3′; p21 sense,
5′-TACTCCCCTGCCCTCAACAAGA-3′ and antisense,
5′-CGCTATCTGAGCAGCGCTCAT-3′; cyclin B1 sense,
5′-GCAACCTCCAAGCCCGGACTG-3′ and antisense,
5′-AAATAGGCTCAGGCGAAAGTT-3′; Wee1 sense,
5′-ATTTCTCTGCGTGGGCAGAAG-3′ and antisense,
5′-CAAAAGGAGATCCTTCAACTCTGC-3′; CDC2 sense,
5′-TCTATCCCTCCTGGTCAGTTC-3′ and antisense,
5′-TGTCCACTGGAGTTGAGTAGC-3′; CDC25c sense,
5′-ACCTCTTTCATACCGTTGCTGG-3′ and antisense,
5′-AACTCCTTGTATCCGCCCTTCA-3′; CDK7 sense, 5′-GTGGGCTGTTTGCTGTAT-3′
and antisense, 5′-TTCTTGGGCAATCCTCCT-3′; cyclin E sense,
5′-CAGGGTATCAGTGGTGCGACA-3′ and antisense,
5′-TCTTTGCTCGGGCTTTGTCC-3′; CDK2 sense, 5′-CCACCGAGACCTTAAACC-3′
and antisense, 5′-GTGTAAGTACGAACAGGG-3′; and GAPDH sense,
5′-AAGGTGGTGAAGCAGGCGGC-3′ and antisense,
5′-GAGCAATGCCAGCCCCAGCA-3′. The PCR reaction system (25 µl)
contained 12.5 µl 2X Premix Ex Taq (Takara Bio, Inc., Otsu, Japan),
1 µl each sense and antisense primer, 1 µl cDNA and
ddH2O. The thermocycling conditions for PCR were as
follows: 95°C for 10 min, followed by 40 cycles at 95°C for 10 sec,
60°C for 30 sec and 72°C for 20 sec. The expression ratio of target
gene to internal reference gene (GAPDH) was calculated based on the
fluorescent curve and Cq values were used to determine the
difference in gene expression between the samples (11).
Western blot analysis
Total protein was extracted from tumor tissues by
chemical lysis using lysis buffer (Abcam, Cambridge, MA, USA) and
protein concentration was determined using the BCA method. Total
proteins (25–50 µg) were separated by 8% SDS-PAGE and were
transferred to a nitrocellulose membrane. The membrane was then
stained with Ponceau S (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany), blocked in 1% skimmed milk at room temperature for 1 h
and incubated overnight with primary antibodies diluted at 1:800 in
1.5% bovine serum albumin (BSA; Sigma-Aldrich; Merck KGaA) at 4°C.
Subsequently, the membrane was washed three times with TBS
containing 10% Tween (TBST; 10 min/wash) and incubated with 1.5%
BSA-diluted secondary antibodies (1:1,500) at room temperature for
1 h. The membrane then underwent four further washes with TBST (15
min/wash), after which the blot was visualized with an ECL reagent
for ~1 min, exposed to X-ray film for 5–30 min and scanned. The
grayscale values were analyzed by ImageJ software version 1.41
(National Institutes of Health, Bethesda, MD, USA). GAPDH was used
as the housekeeping protein.
Statistical analysis
All data were analyzed using Microsoft Excel version
2012 (Microsoft Corporation, Redmond, WA, USA) and expressed as
mean ± standard deviation of 3 independent experiments. Data were
analyzed using a two-way analysis of variance, followed by
Dunnett's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Inhibitory effects of ZBH-1208 on B22
solid tumor
As presented in Table
I, compared with the negative control group, the average tumor
weight of the irinotecan group was significantly decreased
(P<0.01), with a high tumor inhibition rate. In addition, the
average tumor weight of the 40 mg/kg ZBH-1208 group was
significantly reduced compared with the negative control group
(P<0.05), with an inhibition rate of >50%.
 | Table I.Inhibitory effects of ZBH-1208 on B22
solid tumors (n=8; mean ± standard deviation). |
Table I.
Inhibitory effects of ZBH-1208 on B22
solid tumors (n=8; mean ± standard deviation).
| Group | Dose (mg/kg) | Average tumor weight
(g) | Inhibition rate
(%) |
|---|
| Negative control | – |
3.094±1.022 | – |
| Irinotecan | 25 |
0.391±0.112a | 87.7 |
| ZBH-1208 | 20 |
2.739±1.131 | 11.7 |
|
| 40 |
1.422±0.224b | 54.1 |
Effects of ZBH-1208 on immune organs
and white blood cell count
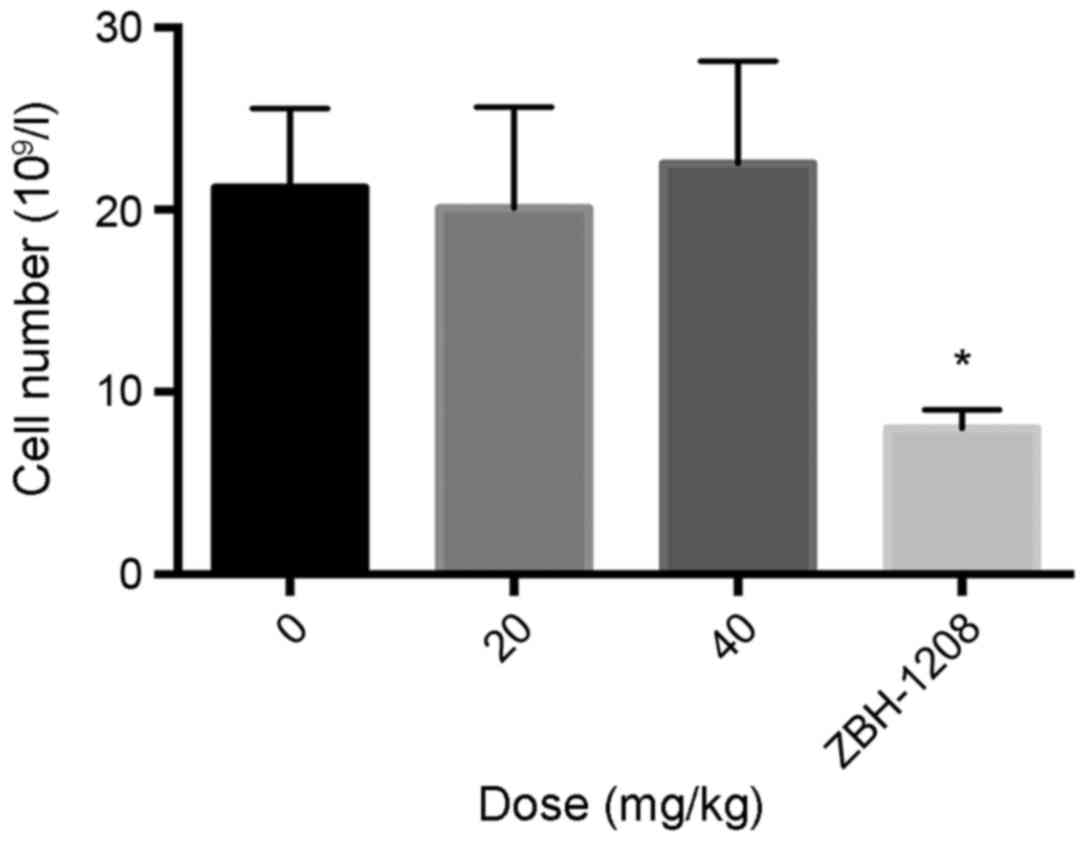
The average white blood cell count of the irinotecan
group (4.5×109 l−1) was significantly lower
than that of the negative control group (21.3×109
l−1); however, the average white blood cell count of the
20 and 40 mg/kg ZBH-1208 groups was similar to the negative control
group (20.7×109 and 21.5×109 l−1
respectively) (Fig. 2).
As presented in Table
II, the irinotecan group exhibited a significantly reduced
weight difference, and reduced thymic and splenic indices, compared
with the negative control group (P<0.05). However, the values of
the ZBH-1208 groups were similar to those of the negative control
group.
| Table II.Effects of ZBH-1208 on visceral
indices (n=8, mean ± standard deviation). |
Table II.
Effects of ZBH-1208 on visceral
indices (n=8, mean ± standard deviation).
| Group | Dose (mg/kg) | Weight difference
(g) | Splenic index
(mg/g) | Thymic index
(mg/g) |
|---|
| Negative control | – |
18.7±2.1 |
11.405±4.234 |
2.425±0.421 |
| Irinotecan | 25 |
15.1±2.1a |
6.237±1.002a |
1.235±0.112b |
| ZBH-1208 | 20 |
17.3±1.1b |
12.325±5.210 |
4.157±0.112 |
|
| 40 |
18.4±2.2 |
10.535±2.117 |
2.279±0.471 |
mRNA expression levels of cell
cycle-related genes, as detected by RT-qPCR
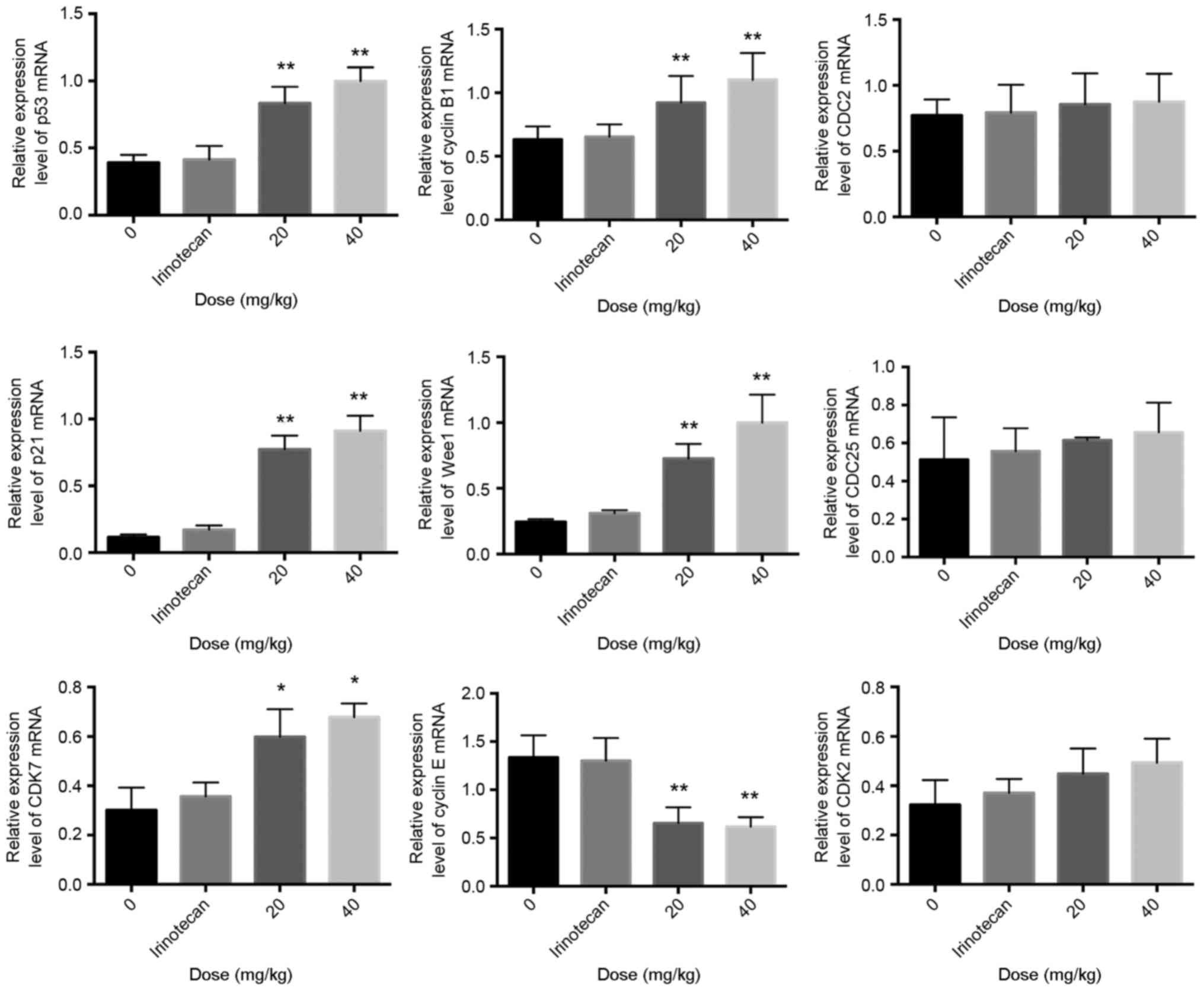
Compared with the negative control group, the mRNA
expression levels of p53, p21, cyclin B1, Wee1 and CDK7 in the
ZBH-1208 groups were upregulated to different extents in a
dose-dependent manner. Conversely, the mRNA expression levels of
cyclin E were significantly downregulated in response to ZBH-1208,
in a dose-independent manner. In addition, ZBH-1208 treatment did
not affect the mRNA expression levels of CDC2, CDK2 or CDC25C
(Fig. 3).
Protein expression levels of cell
cycle-related proteins, as detected by western blotting
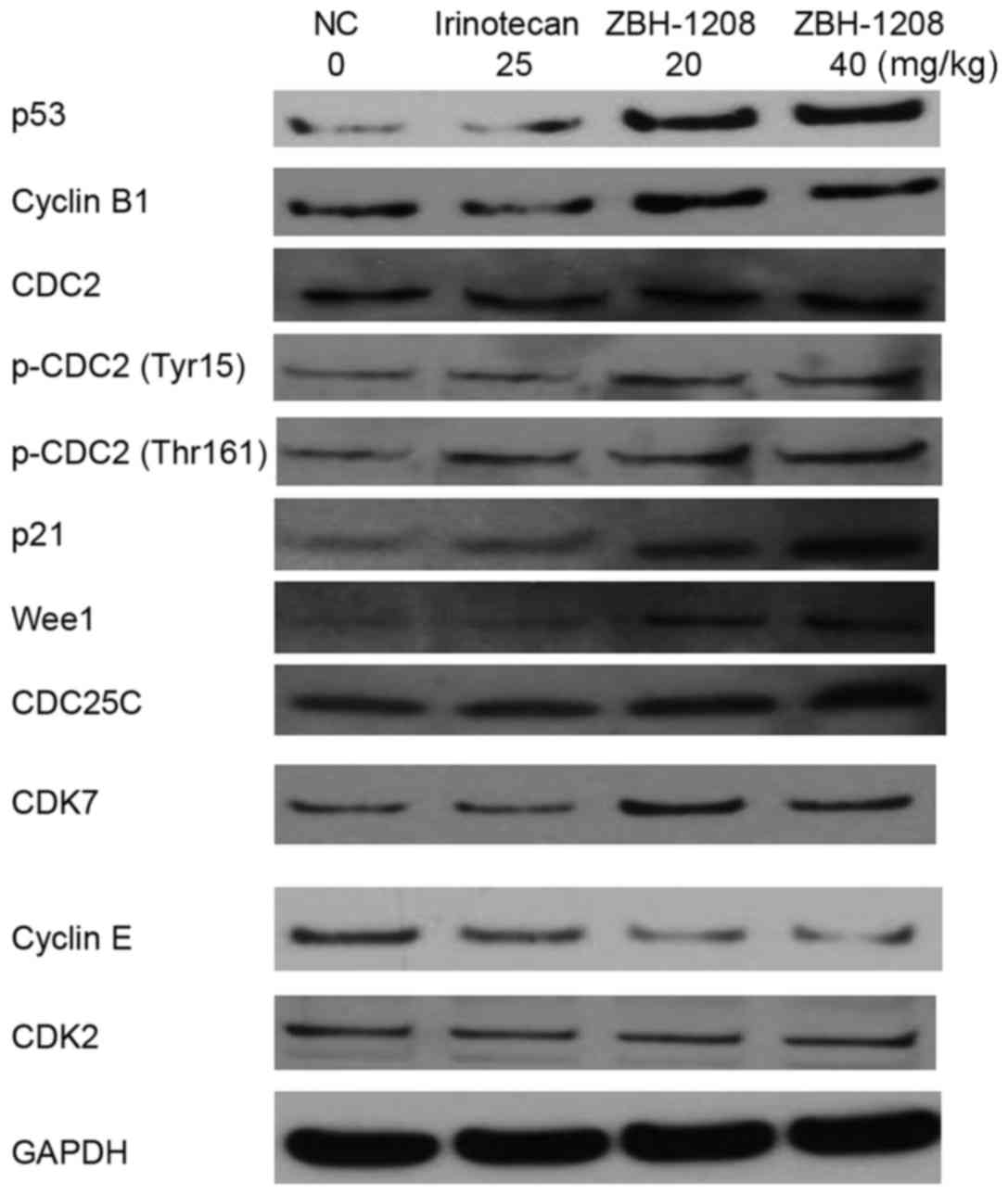
Compared with the negative control group, the
expression levels of p53, p21, CDK7, Wee1, p-CDC2 (Tyr15), p-CDC2
(Thr161) and cyclin B1 were upregulated, whereas the expression of
cyclin E was downregulated in the ZBH-1208 groups. However, the
expression levels of CDC2, CDK2 and CDC25C were barely altered in
response to ZBH-1208 treatment (Fig.
4).
Discussion
Irinotecan and its derivatives commonly exist as
non-bioactive hydroxylated forms or bioactive lactone forms in
vivo. At low pH, the non-bioactive hydroxylated forms can
easily transform into bioactive lactones, following an
interconvertible, dynamically balanced reaction. SN-38 is the
active metabolite of irinotecan in vivo; previous
unpublished data from our group suggested that SN-38 may
predominantly affect the S phase of tumor cells and target DNA Topo
I. In the liver, irinotecan is metabolized by uridine diphosphate
glucuronosyltransferase 1A1 (UGT1A1) into the nonactive glucuronide
product SN-38G (7), which is
secreted into the intestinal tract through bile and converted into
active SN-38 by β-glucuronidase. As a result, the intestinal mucosa
is injured, thus causing toxic side effects, such as delayed
diarrhea. Furthermore, UGT1A1 catalyzes SN-38 into SN-38G for
detoxification (12,13). Notably, irinotecan and its
derivatives only serve biological roles following in vivo
metabolism; therefore, the present study performed in vivo
experiments. Since drugs for the treatment of brain tumors remain
limited in clinical practice, the present study focused on the
anticancer effects of ZBH-1208 on B22 mouse xenografts. As a novel
derivative of irinotecan, ZBH-1208 inhibited subcutaneously
implanted B22 tumors in mice; however, its role in treating
orthotopically implanted tumors requires further verification.
Furthermore, the present study observed the immunotoxic effects of
ZBH-1208. Briefly, irinotecan (25 mg/kg) decreased weight, visceral
index and white blood cell count, whereas ZBH-1208 at the same
concentration (20 mg/kg) or higher (40 mg/kg) did not induce
discernible toxicity.
Suppression of cell cycle progression is of great
significance to the inhibition of tumor growth. In eukaryotes,
there are two crucial regulatory points in the cell cycle:
G1/S and G2/M phases. Cell cycle progression
is regulated by cyclins and CDKs, which are negatively regulated by
CDK inhibitors (CKIs) (14,15).
As a CKI, p21 is markedly expressed in the early G2/M
phase, which induces temporary arrest in the G2 phase,
blocks cell division and proliferation, and finally suppresses
tumor cell growth. In addition, it inhibits the activities of
various cyclin-CDK complexes and negatively regulates their
functions to affect cell cycle progression. For example, p21 is
able to independently bind cyclin E and CDK2 and deactivate them,
thereby arresting cells in the G1/S phase (15–18).
The product of the antioncogene p53, i.e., the p53 protein, serves
a critical role in the G1/S phase transition. As an
upstream gene of p21, it initiates the transcription and expression
of p21 following transcription (16–19).
It has previously been reported that irinotecan may affect the
expression levels of cell cycle-related proteins, including cyclin
D and cyclin B, to nonspecifically block the cell cycle (20). To clarify the mechanism by which
ZBH-1208 inhibits the growth of B22 brain tumor xenografts, the
present study assessed its effects on the expression levels of cell
cycle-related proteins. In a dose-dependent manner, ZBH-1208
upregulated the expression levels of p53 and p21, which may lead to
p53/p21 pathway-mediated cell cycle arrest. However, whether
ZBH-1208 affects other CKIs, such as p16 and p27, requires further
study.
Cyclin E-CDK2 has an essential role in
G1/S transition. Synthesized in the middle of the
G1 phase, cyclin E has a high content in S phase and
gradually degrades thereafter. The expression levels of CDK2 remain
unchanged throughout the cell cycle. After binding CDK2, cyclin E
can activate it and form a complex to promote phosphorylation of
retinoblastoma protein, thus releasing transcription factor E2F and
accelerating the progression of G1/S phase (21,22).
In the present study, ZBH-1208 downregulated cyclin E expression;
however, it did not affect CDK2 expression. Regulation of the
G2/M phase depends on formation of the cyclin B1/CDC2
complex, i.e., mitosis promoting factor (MPF). Cyclin B1 is
specifically synthesized from the S phase, which gradually
increases and peaks in the G2/M phase. Activation of
CDC2 requires dephosphorylation of Thr14 and Tyr15 residues, as
well as phosphorylation of Thr161. In addition, a decrease in the
activity of Wee1 kinase and CDC25 can dephosphorylate Thr14 and
Tyr15 of CDC2, and Thr161 is phosphorylated in the presence of CDK7
(23,24). In a dose-dependent manner, ZBH-1208
upregulated cyclin B1 expression in B22 xenografts; however, CDC2
expression remained unchanged. Although the resulting upregulated
expression of CDK7 elevated that of p-CDC2 (Thr161), which may
benefit MPF activation, the expression levels of Wee1 were
upregulated and those of CDC25C were unaltered. Accordingly,
phosphorylation of CDC2 Tyr15 was markedly enhanced to inhibit the
activation of CDC2 and the activity of MPF.
In conclusion, ZBH-1208 moderately suppressed B22
mouse brain tumor growth, without significantly reducing visceral
indices of immune organs or white blood cell counts. In addition,
it exerted its effects by upregulating the protein expression
levels of p53, p21, Wee1 and p-CDC2 (Tyr15), and by downregulating
cyclin E expression. Nevertheless, the detailed mechanism remains
unclear. For example, whether ZBH-1208 induces DNA damage via in
vivo metabolism, similar to irinotecan; whether it activates
the p53 protein via the ataxia telangiectasia mutated
serine/threonine kinase/checkpoint kinase 2 pathway; and whether it
may induce immune system toxicity requires further
investigation.
References
|
1
|
DeSantis CE, Lin CC, Mariotto AB, Siegel
RL, Stein KD, Kramer JL, Alteri R, Robbins AS and Jemal A: Cancer
treatment and survivorship statistics, 2014. CA Cancer J Clin.
64:252–271. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Eba J, Shimokawa T, Nakamura K, Shibata T,
Misumi Y, Okamoto H, Yamamoto N and Ohe Y; Lung Cancer Study Group
of the Japan Clinical Oncology Group, : A Phase II/III study
comparing carboplatin and irinotecan with carboplatin and etoposide
for the treatment of elderly patients with extensive-disease
small-cell lung cancer (JCOG1201). Jpn J Clin Oncol. 45:115–118.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Misumi Y, Nishio M, Takahashi T, Ohyanagi
F, Horiike A, Murakami H, Kenmotsu H, Yamamoto N, Ishii M,
Shimokawa T, et al: A feasibility study of carboplatin plus
irinotecan treatment for elderly patients with extensive disease
small-cell lung cancer. Jpn J Clin Oncol. 44:116–121. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Peleg R, Romzova M, Kogan-Zviagin I, Apte
RN and Priel E: Modification of topoisomerases in mammospheres
derived from breast cancer cell line: Clinical implications for
combined treatments with tyrosine kinase inhibitors. BMC Cancer.
14:9102014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu Y, Wang KY, Li Z, Liu YP, Izumi H,
Uramoto H, Nakayama Y, Ito K and Kohno K: Y-box binding protein 1
enhances DNA topoisomerase 1 activity and sensitivity to
camptothecin via direct interaction. J Exp Clin Cancer Res.
33:1122014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pitot HC: US pivotal studies of irinotecan
in colorectal carcinoma. Oncology (Williston Park). 12 8 Suppl
6:S48–S53. 1998.
|
|
7
|
Hind D, Tappenden P, Tumur I, Eggington S,
Sutcliffe P and Ryan A: The use of irinotecan, oxaliplatin and
raltitrexed for the treatment of advanced colorectal cancer:
Systematic review and economic evaluation. Health Technol Assess.
12:iii–ix, xi–162. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sadahiro S, Suzuki T, Tanaka A, Okada K,
Saito G and Kamijo A: A phase II trial of combined chemotherapy
with oral S-1 and 24-hour infusions of irinotecan plus bevacizumab
in patients with metastatic colorectal cancer. Oncology.
88:353–359. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao J: Study on anti-tumor activity and
mechanism of novel Irinotecan derivative ZBH-1208 in vitro
(unpublished PhD thesis). Jilin University. 2015.
|
|
10
|
Yin Q, Fischer L, Noethling C and Schaefer
WR: In vitro-assessment of putative antiprogestin activities of
phytochemicals and synthetic UV absorbers in human endometrial
Ishikawa cells. Gynecol Endocrinol. 31:578–581. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Graeven U, Arnold D, Reinacher-Schick A,
Heuer T, Nusch A, Porschen R and Schmiegel W: A randomised phase II
study of irinotecan in combination with 5-FU/FA compared with
irinotecan alone as second-line treatment of patients with
metastatic colorectal carcinoma. Onkologie. 30:169–174.
2007.PubMed/NCBI
|
|
13
|
Mackenzie PI, Bock KW, Burchell B,
Guillemette C, Ikushiro S, Iyanagi T, Miners JO, Owens IS and
Nebert DW: Nomenclature update for the mammalian UDP
glycosyltransferase (UGT) gene superfamily. Pharmacogenet Genomics.
15:677–685. 2015. View Article : Google Scholar
|
|
14
|
Stewart ZA, Westfall MD and Pietenpol JA:
Cell-cycle dysregulation and anticancer therapy. Trends Pharmacol
Sci. 24:139–145. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Arellano M and Moreno S: Regulation of
CDK/cyclin complexes during the cell cycle. Int J Biochem Cell
Biol. 29:559–573. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xiong Y, Hannon GJ, Zhang H, Casso D,
Kobayashi R and Beach D: p21 is a universal inhibitor of cyclin
kinases. Nature. 366:701–704. 1993. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Goldstein M, Roos WP and Kaina B:
Apoptotic death induced by the cyclophosphamide analogue
mafosfamide in human lymphoblastoid cells: Contribution of DNA
replication, transcription inhibition and Chk/p53 signaling.
Toxicol Appl Pharmacol. 229:20–32. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen P: Research progress on tumor-related
genes. J Basic Clin Oncol. 21:90–92. 2008.(In Chinese).
|
|
19
|
Nigam N, Prasad S, George J and Shukla Y:
Lupeol induces p53 and cyclin-B-mediated G2/M arrest and targets
apoptosis through activation of caspase in mouse skin. Biochem
Biophys Res Commun. 381:253–258. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Huang MY, Pan H, Xu LH, Hou XF, Zha QB,
Shi ZJ, He XH and Ouyang DY: Effect of irinotecan on proliferation
and apoptosis of RAW 264.7 macrophages. Immunol J. 31:776–780.
2015.(In Chinese).
|
|
21
|
Chen J and Song WZ: Mechanism of molecular
regulation of CyclinE-CDK2. J Med Mol Biol. 3:220–222. 2006.
|
|
22
|
Shu W, Ma QJ and Ye X: CyclinE-CDK2 and
its related proteins in the cell cycle. Lett Biotechnol. 19:97–100.
2008.
|
|
23
|
O'Connell MJ, Walworth NC and Carr AM: The
G2-phase DNA-damage checkpoint. Trends Cell Biol. 10:296–303. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yan XL, Xu B, Li M, Zhou Y and Cui JR:
Inhibition of diacetyldianhydrogalactitol on ornithine
decarboxylase activity and its possible mechanisms. Chin Pharm J.
44:904–908. 2009.(In Chinese).
|