Introduction
Type 2 diabetes mellitus (T2DM) is a major public
health problem worldwide. According to the International Diabetes
Federation Diabetes Atlas 2015, there are 415 million people living
with diabetes worldwide, and this number is expected to rise to 642
million by 2040 (1). T2DM
constitutes >90% of the cases of diabetes, and its prevalence
has been dramatically increasing in developing countries,
especially in China. T2DM is characterized by hyperglycemia,
hyperinsulinemia, and subclinical chronic inflammation (2,3).
Diabetic patients have increased risk of several types of
malignancies, including breast, pancreas, liver, urinary tract,
female reproductive organ, and colorectal cancer (4). Breast cancer is one of the most
common cancers worldwide and has a high mortality rate in women
(5). A recently reported
meta-analysis revealed that women with diabetes had a significantly
higher risk (~20%) of breast cancer than those without diabetes
(6). Hyperinsulinemia and T2DM
were demonstrated to be independent risk factors for postmenopausal
breast cancer (7). In addition,
accumulating data suggest that diabetes and its complications can
adversely affect cancer therapy (8) and increase mortality (9), thus affecting the outcome of breast
cancer patients (10,11).
The insulin-like growth factor (IGF) and insulin
receptors (IR) are important in breast cancer development and
progression in T2DM patients (12–14).
Insulin, not only elicits direct mitogenic effects through its
actions on tumor cell growth, invasion and tumor-related
angiogenesis (15), but also
indirectly promotes estrogen and IGF response in both normal and
malignant breast tissues (16).
Insulin receptor substrate 1 (IRS1) is a regulator of insulin, IGF,
and cytokine signaling, and therefore serves an important role in
the proliferation, survival, and transformation of cells, by
conveying signals to the phosphatidylinositol 3-kinase (PI3K)/AKT
serine/threonine kinase 1 (Akt) and extracellular signal-regulated
kinase (ERK) 1/2 pathways. In general, the metabolic effects of
insulin, such as glucose transport, are mediated by the PI3K
pathway, whereas the mitogenic effects of insulin involve the
mitogen-activated protein kinase (MAPK) pathway (17).
To date, the molecular mechanisms responsible for
the association between diabetes and breast cancer are not well
understood. The purpose of the present study was therefore to
investigate the effect of high glucose and high insulin conditions
on the proliferation and invasion of MCF-7 cells and to understand
the molecular mechanisms underlying this effect.
Materials and methods
Cell culture and treatment
MCF-7 breast cancer cells, purchased from the
Institute of Cell Research of the Chinese Academy of Sciences
(Shanghai, China), were cultured in Dulbecco's modified Eagle's
medium (DMEM; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo
Fisher Scientific, Inc.) and antibiotics (100 IU/ml penicillin and
100 mg/ml streptomycin) in 5% CO2 at 37°C. The cells
were cultured in normal glucose + low insulin (5.6 mM glucose + 5
nM insulin) or high glucose + high insulin (25 mM glucose + 25 nM
insulin) conditions.
Cell viability and cell proliferation
assays
The MTT cell viability assay is based on the
conversion of MTT to violet-colored formazan crystals by
mitochondrial dehydrogenases (18). For the MTT assay, the cells were
seeded in 96-well plates at a density of 1×104
cells/well. The cells were allowed to attach and proliferate for 24
h, and then subjected to the normal glucose + low insulin (5.6 mM
glucose + 5 nM insulin) or high glucose + high insulin (25 mM
glucose + 25 nM insulin) conditions for 24 h. Then, the cells were
incubated with 0.1 mg/ml MTT at 37°C for 4 h and lysed in dimethyl
sulfoxide at room temperature for 10 min to dissolve the formazan
crystals. The absorbance in each well was measured at 570 nm using
a SpectraMax i3 spectrophotometer (Molecular Devices, LLC,
Sunnyvale, CA, USA) and the results were expressed as % of cell
viability relative to the control cells.
For the proliferation assay,
5-Ethynyl-2′-deoxyuridine (EdU) was used. EdU is a nucleoside
analog of thymidine that is readily incorporated into cellular DNA
during DNA replication. Cell proliferation was evaluated using a
Cell-Light EdU Apollo 567 In Vitro Imaging kit (Ribobio Co.,
Ltd., Guangzhou, China), according to the manufacturer's protocol.
Briefly, the cells were incubated with 50 µM EdU for 2 h at 37°C,
fixed with 4% formaldehyde, stained with the Apollo reaction
cocktail and Hoechst 33342 as a nuclear counterstain, and protected
from light. Images were acquired under a fluorescent microscope and
the EdU positive cells (red cells) were counted in five random
fields per sample. The EdU incorporation rate was expressed as the
ratio of EdU positive cells to total Hoechst 33342 positive cells
(blue cells). All experiments were repeated independently at least
three times. ImageJ software v1.48 (National Institutes of Health,
Bethesda, MD, USA) was used to generate overlapping images
(19).
Total RNA extraction and reverse
transcription (RT)
Total RNA was extracted from cells using the TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocol. The total RNA was then converted to
cDNA using the PrimeScript 1st strand cDNA Synthesis kit (Takara
Bio, Inc., Otsu, Japan), according to the manufacturer's
protocol.
Quantitative polymerase chain reaction
(qPCR)
qPCR was performed in a LightCycler 480 system
(Roche Applied Science, Penzberg, Germany) using the SYBR Green
Master Mix (Takara Bio, Inc.) (20,21).
The following primers were used: IRS1, forward
5′-TTTGTGGTCCTTCCGTAGTT-3′ and reverse 5′-CCTGCCCCTAATGTGATGCT-3′;
β-actin, forward 5′-AAGGTGACAGCAGTCGGTT-3′ and reverse
5′-GTGTGGACTTGGGAGAGG-3′. The PCR conditions were: 95°C for 30 sec,
followed by 40 cycles of 95°C for 5 sec and 60°C for 30 sec. IRS1
gene expression was normalized to β-actin expression for each
sample. The relative mRNA expression was calculated using the -ΔΔCq
method (22).
Western blot analysis
Western blot analysis was conducted as described
previously (23,24). Briefly, the cells were cultured
under normal (5.6 nM) or high-glucose conditions (25 nM) for 2
weeks. Following overnight serum starvation, the cells were then
incubated with low insulin (5 nM) or high insulin (25 nM) for 2 h.
Cells were washed with ice-cold PBS and lysed in
Radioimmunoprecipitation Lysis Buffer (Beyotime Institute of
Biotechnology, Shanghai, China) containing a 1% protease inhibitor
cocktail (Beyotime Institute of Biotechnology). The cell lysates
were spun at 10,000 × g for 10 min at 4°C, and the resulting
supernatant was stored at −80°C. Protein concentration was
determined using the bicinchoninic acid protein assay (Biyuntian
Biotechnology Co.). The proteins were separated by 8–12% SDS-PAGE
and transferred to polyvinylidene difluoride membranes (EMD
Millipore, Billerica, MA, USA). The membranes were initially
blocked with 5% nonfat dry milk in TBS/0.1% Tween 20 for 1 h and
then incubated with primary antibodies specific to IRS1 (cat. no.
ab52167; 1:1,000; Abcam, Cambridge, MA, USA), tubulin (cat. no.
MB0009; 1:5,000; Bioworld Technology, Inc., St Louis Park, MN,
USA), phosphorylated (p)-ERK1/2 (cat. no. #4377; 1:1,000; Cell
Signaling Technology, Inc., Danvers, MA, USA), total ERK1/2 (cat.
no. #4695; 1:1,000; Cell Signaling Technology, Inc.), RAS
proto-oncogene (Ras; cat. no. ab108602; 1:1,000; Abcam) and RAF1
proto-oncogene (Raf-1; cat. no. ab137435; 1:1,000; Abcam) at 4°C
overnight. The membranes were then incubated with the appropriate
horseradish peroxidase-conjugated secondary antibody (anti-rabbit
IgG; cat. no. #7074; 1:1,000; Cell Signaling Technology, Inc.) at
room temperature for 2 h. The immunoreactions were visualized using
the Amersham Enhanced Chemiluminescence Plus western blotting
detection reagents (GE Healthcare, Chicago, IL, USA), and the
resulting band intensity was quantified using ImageJ software v1.48
(National Institutes of Health) (19).
Wound healing assay
To determine cell migration, MCF-7 cells were seeded
in 6-well plates, incubated in normal glucose + low insulin (5.6 mM
glucose + 5 nM insulin) or high glucose + high insulin (25 mM
glucose + 25 nM insulin) conditions and grown to confluence
overnight. Wounds were made by scraping with a sterilized 10 µl
pipette tip, and cells were photographed under a phase contrast
microscope (Carl Zeiss AG, Oberkochen, Germany) at 0, 24 and 72 h.
The wound width was evaluated by measuring the distance between the
two edges of the scratch in 5 random fields per plate; 3
plates/group were analyzed in total. Relative wound recovery was
determined using the following formula: [(Wound width End time
point-Wound width Starting time point)/Wound
widthStarting time point] × 100%. All experiments were
repeated independently at least three times.
Invasion assay
The cell invasion assay was performed using 24-well
transwell chambers (8.0 µm; Corning Incorporated, Corning, NY,
USA). Cell suspensions were prepared in serum-free media containing
2×104 cells/ml, and 200 µl of cell suspension was seeded
into the upper chamber of transwells that were pre-coated with
Matrigel (cat. no. 354234; Corning Incorporated). Then, 600 µl of
medium containing 10% serum was added to the lower chamber as the
chemoattractant. After 24 h of incubation, the cells on the upper
side of the transwell were removed using a cotton swab, and the
filters were fixed with 100% methanol for 30 min, followed by
staining with Crystal Violet Staining Solution (Beyotime Institute
of Biotechnology). For each chamber, four random fields were
observed by phase contrast microscopy, and the average number of
invading cells in each group was counted.
Statistical analysis
All experiments were repeated independently at least
three times. Results were expressed as mean ± standard deviation.
Differences between two groups were assessed using the Student's
t-test (two-tailed). Statistical analysis was performed using
GraphPad PRISM 6 software (GraphPad Software, Inc., La Jolla, CA,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Effect of high glucose and high
insulin on the proliferation of MCF-7 cells
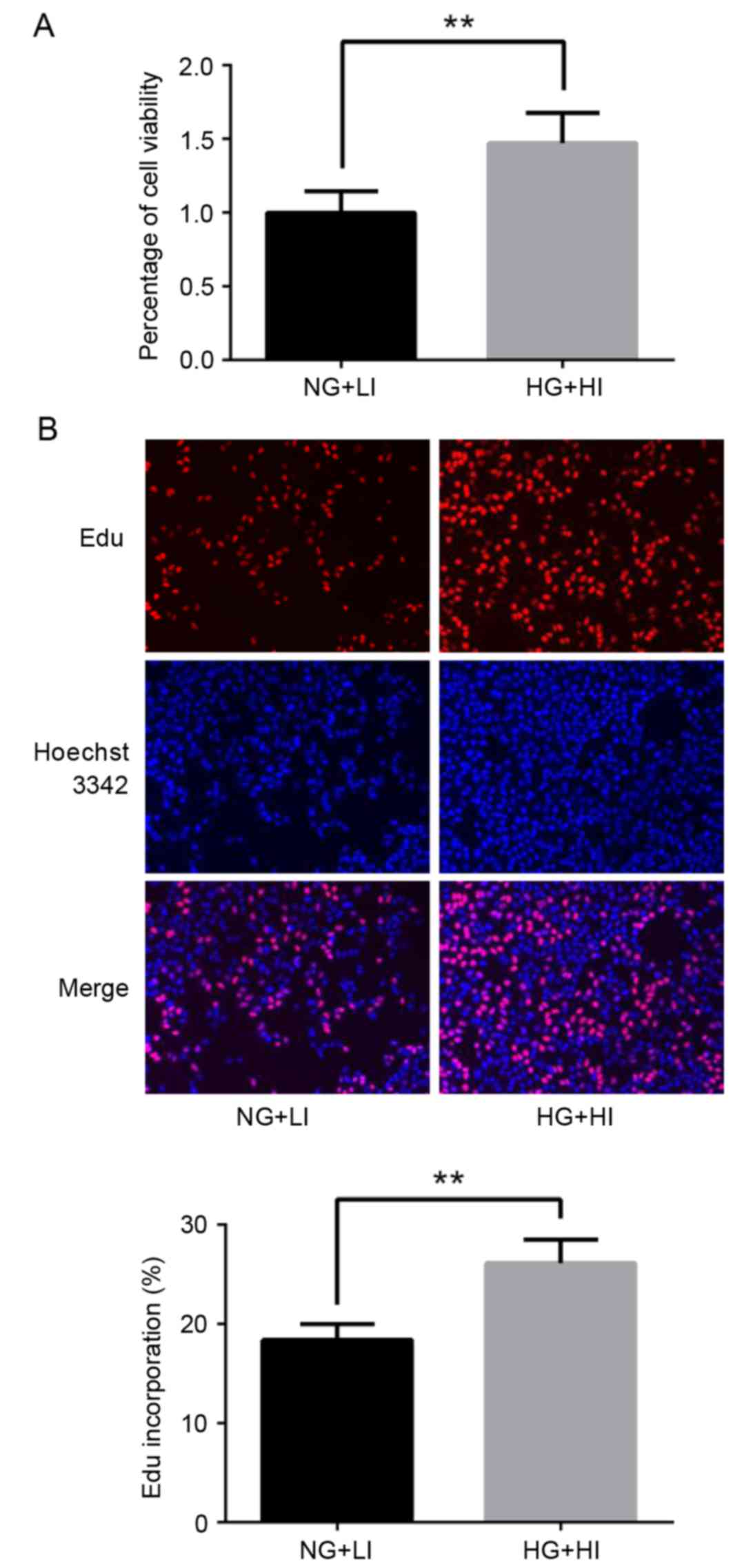
Results from the MTT assay revealed that high
glucose and high insulin culturing conditions resulted in increased
viability in the MCF-7 cells compared with the control normal
glucose and low insulin conditions (Fig. 1A). Additionally, results from the
EDU cell proliferation assay demonstrated that MCF-7 cells
exhibited increased proliferation under high glucose and high
insulin culturing conditions, comparing with control conditions
(Fig. 1B).
High glucose and high insulin
conditions promote the migration and invasion of MCF-7 cells
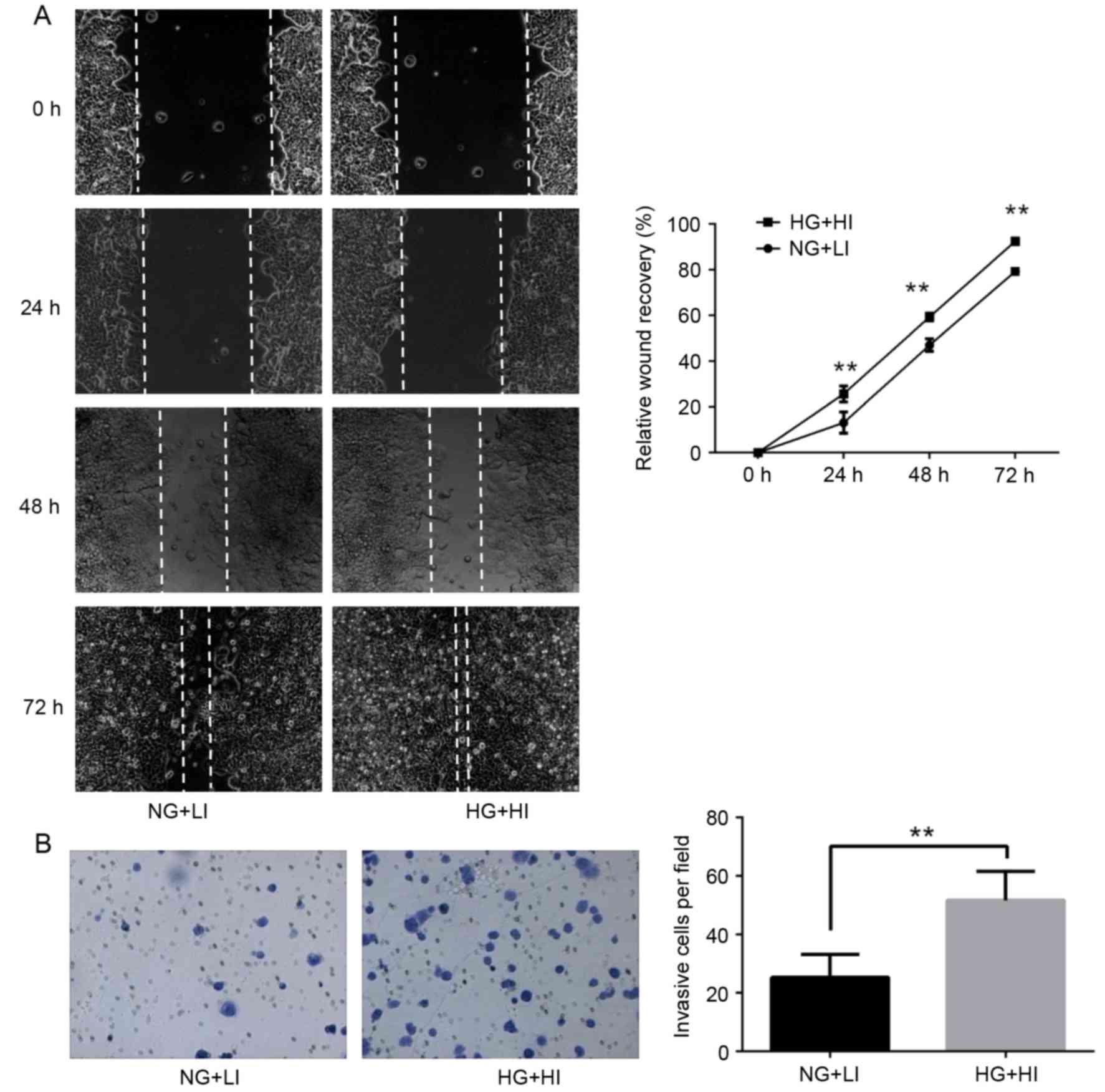
To determine whether high glucose and high insulin
conditions are associated with the progression of breast cancer,
the effect of these conditions on the invasive behavior of MCF-7
cells was examined. Using a wound healing assay, the results
demonstrated that high glucose and high insulin culturing
conditions promoted MCF-7 cell migration, compared with control
culturing conditions (Fig. 2A). In
addition, the invasive ability of MCF-7 cells was significantly
increased following exposure to high glucose and high insulin
conditions, as determined using a transwell invasion assay
(Fig. 2B). Taken together, these
results indicated that exposure of MCF-7 cells to high glucose and
high insulin conditions increased their migration and invasion
ability.
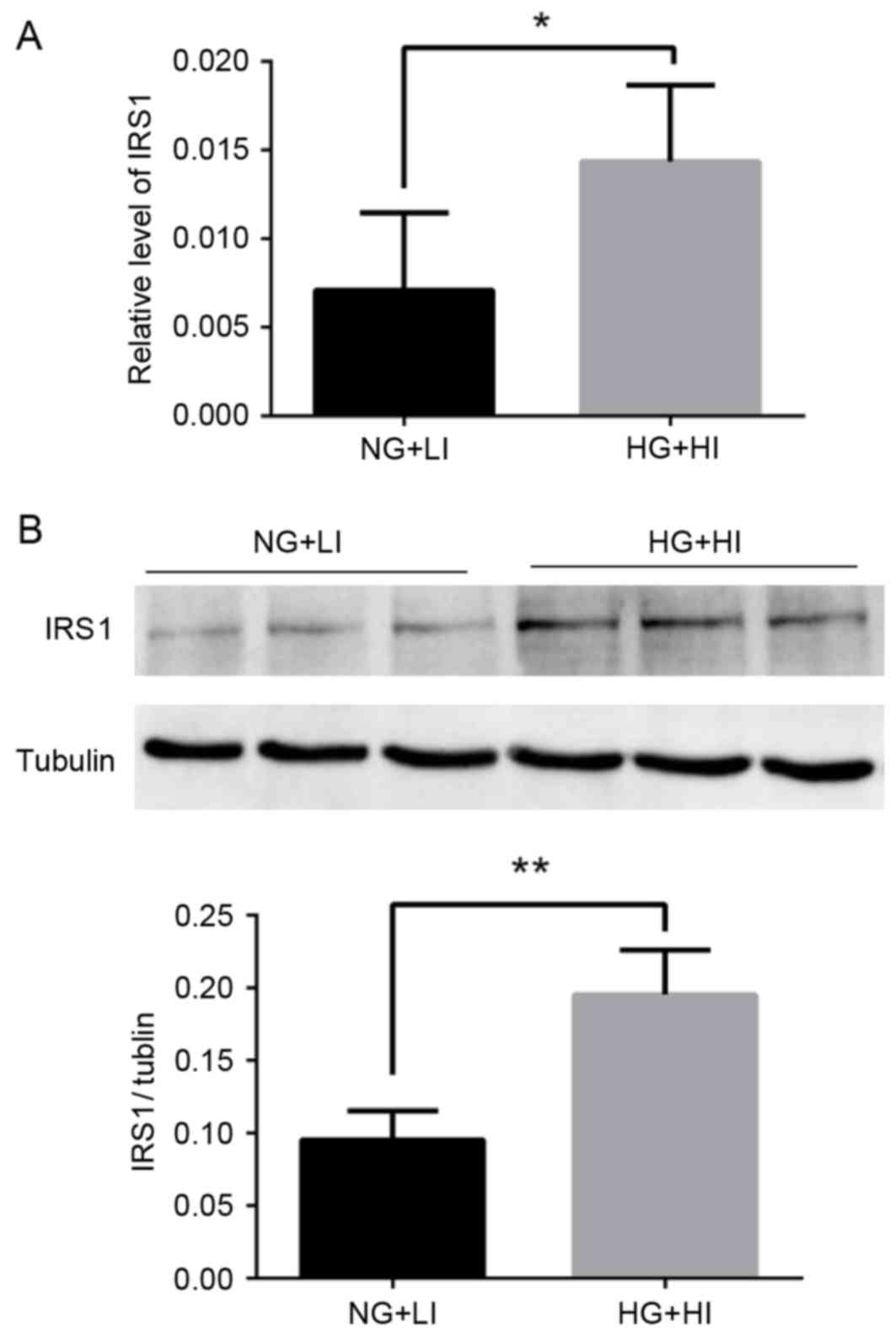
Exposure to high glucose and high
insulin conditions upregulates IRS1 in MCF-7 cells
In order to explore the potential mechanisms
underlying the behavior of the MCF-7 cells exposed to high glucose
and high insulin, the expression of IRS1 was examined by RT-qPCR
and western blotting. IRS1 mRNA and protein expression was
significantly upregulated following exposure to high glucose and
high insulin culturing conditions compared with normal glucose and
low insulin conditions (Fig.
3).
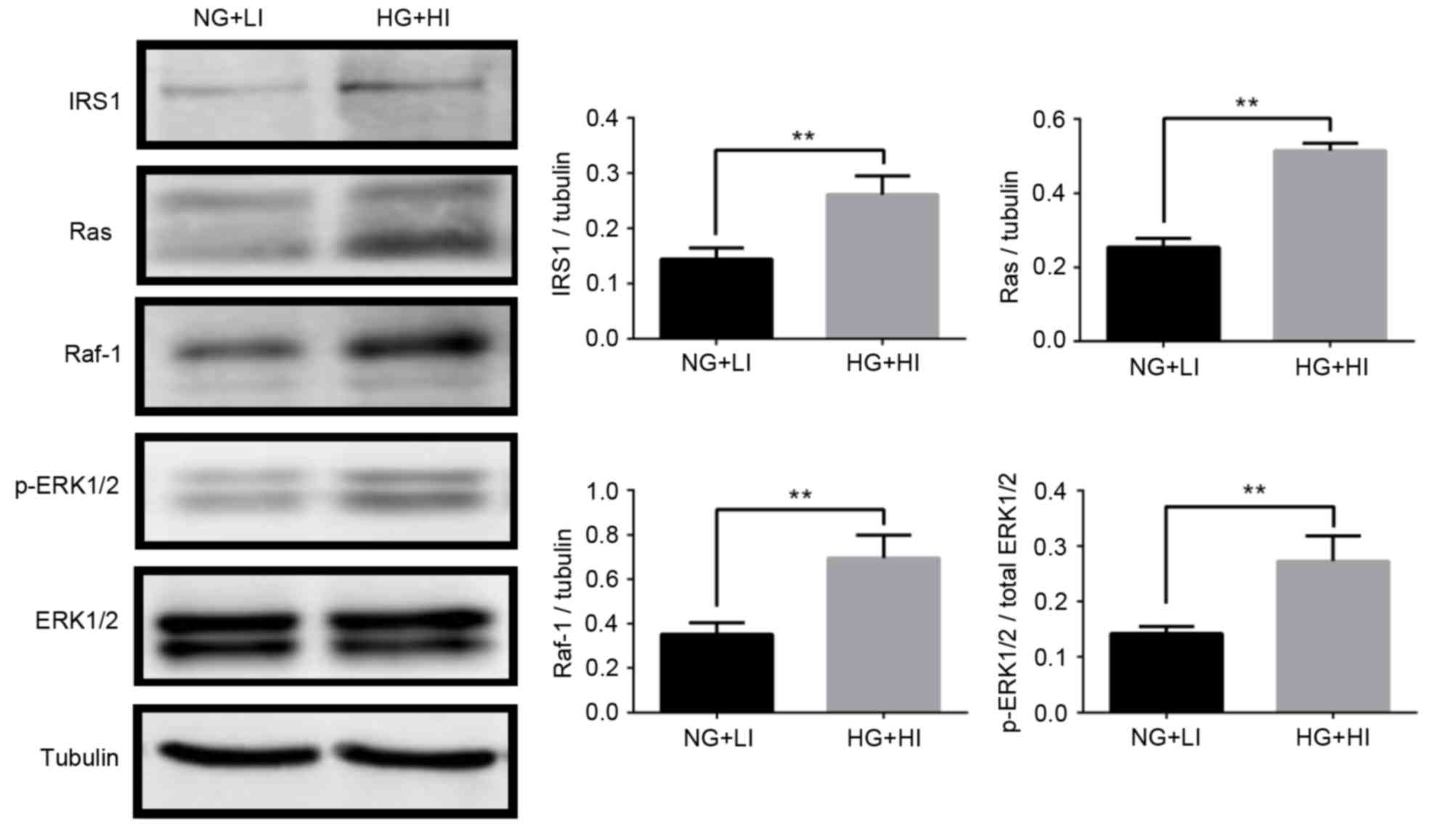
IRS1 promotes Ras-ERK pathway
activation in MCF-7 cells
IRS1 promotes activation of the downstream
Ras/Raf/ERK signaling pathway, which is important for the
regulation of cell proliferation, apoptosis, and differentiation
(25). This pathway was thus
examined in the MCF-7 cells by western blotting. The results
demonstrated that ERK1/2 phosphorylation increased significantly
under the high glucose and high insulin culturing conditions
compared with the normal glucose and low insulin conditions
(Fig. 4). In addition, under the
high glucose and high insulin culturing conditions, protein
expression levels of Ras and Raf-1 were significantly upregulated
(Fig. 4). The results suggested
that, in MCF-7 breast cancer cells, ERK may be phosphorylated by
the sequential activation of Ras and Raf-1, thereby inducing cell
survival, proliferation, and invasion.
Discussion
With the changing global lifestyle, the number of
people suffering from T2DM is constantly increasing. Female
diabetic patients have an increased risk of breast cancer and its
related mortality. However, the relationship between diabetes
mellitus and breast cancer remains unclear. The results of the
present study demonstrated that high glucose and high insulin
conditions promoted the proliferation and invasion of MCF-7 breast
cancer cells by upregulating IRS1 and activating the Ras/Raf/ERK
pathway.
Insulin resistance, which is characterized by
hyperglycemia and hyperinsulinemia, is a major cause in the
pathogenesis of T2DM. Insulin is a peptide hormone produced by the
pancreatic b-cells, and it is well known for its involvement in
cell survival and proliferation, as well as for its effect on
mitogenic signals (26).
Additionally, insulin receptors are frequently overexpressed in
breast cancer cells (27,28). Among the metabolic changes
exhibited by cancer cells, an increase in glucose metabolism and
glucose dependence is common (29). Elevated glucose levels directly
promote the proliferation of tumor cells by functioning as a source
of energy. As an adaptor of insulin, IRS1 has been demonstrated to
act as an oncogene, and serve major roles in the growth,
proliferation, migration, invasion, and differentiation of cells
(30). Constitutive IRS1
activation is implicated in a variety of solid tumors, including
breast cancer (31). The
expression of IRS1 is increased in breast cancer tissues, with
higher levels in the well-differentiated tumors compared with the
poorly differentiated tumors (32). Because of the cross-talk between
IGF signaling and estrogen receptor (ER) signaling pathways, IRS1
expression is regulated by estrogen in MCF-7 cells (33). MCF-7 is a hormone-dependent breast
cancer cell line due to its expression of ER. Expression and
activation of the ER enhances insulin mitogenicity by upregulating
IRS1 and increasing PI3 K/Akt and MAPK signaling (34). High IRS1 expression is an indicator
of early disease recurrence in ER-positive human primary breast
tumors (35), and it indicates
increased sensitivity to IGF-1 stimulated cell migration (36). Downregulation of IRS1 can suppress
the growth of MCF-7 cells (36)
and enhance the cytotoxic effects of tamoxifen (37). In vivo, transgenic mice
overexpressing IRS1 have been reported to develop breast cancer and
subsequent metastasis (38).
Consistent with these previous studies, the present results
demonstrated that IRS1 expression in MCF-7 breast cancer cells was
increased following high glucose and high insulin culturing
conditions compared with normal glucose and low insulin
conditions.
MAPKs are known to be involved in transmitting
extracellular signals that regulate cell growth, differentiation,
and apoptosis (39), and to serve
an essential role in MCF-7 cell cycle progression (40). Four dominating MAPK signaling
cascades, the ERK1/2, ERK5, c-Jun N-terminal kinase (JNK) and p38
pathways, are implicated in normal function of mammary epithelial
cells as well as the pathogenesis of breast cancer (41). Of these, the ERK-1/2 pathway is the
most relevant to breast cancer (42). The Ras family of proto-oncogenes
(comprising of H-Ras, N-Ras and K-Ras) encodes small GTP-binding
proteins that transmit growth-promoting signals from the plasma
membrane to the nucleus. This cascade involves predominantly three
kinases. Upon the activation of Ras (a GTPase), Raf-1 acts as an
MAPK kinase kinase and phosphorylates and activates MEK1/2, which
then activates ERK1/2. Raf-1, a highly-conserved serine/threonine
kinase of the MAPK pathway, serves a central role in the MAPK
signaling pathway and is, thus, involved in proliferation,
transformation, survival, and metastasis of cells. Active MAPK
expression is significantly higher in breast tumors than in the
adjacent normal breast tissue and may be a marker of breast cancer
metastasis (43). The present
results revealed that ERK phosphorylation and Ras/Raf-1 protein
expression were significantly increased under high glucose and high
insulin conditions, thereby indicating that the enhanced
proliferation and invasion of MCF-7 cells may be due to activation
of the Ras/Raf/ERK pathway.
Taken together, the present findings indicate that,
in MCF-7 breast cancer cells, high glucose and high insulin
conditions promoted cell proliferation and invasion by upregulating
IRS1 and activating the Ras/Raf/ERK pathway. A better understanding
of the mechanisms underlying the effects of high glucose and high
insulin on breast cancer cells may aid in developing prevention
strategies against disease progression.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81260133).
References
|
1
|
http://www.diabetesatlas.org/
|
|
2
|
American Diabetes Association, . Diagnosis
and classification of diabetes mellitus. Diabetes Care. 33 Suppl
1:S62–S69. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
DeFronzo RA: Pathogenesis of type 2
diabetes mellitus. Med Clin North Am. 88:787–835, ix. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vigneri P, Frasca F, Sciacca L, Pandini G
and Vigneri R: Diabetes and cancer. Endocr Relat Cancer.
16:1103–1123. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Larsson SC, Mantzoros CS and Wolk A:
Diabetes mellitus and risk of breast cancer: A meta-analysis. Int J
Cancer. 121:856–862. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vona-Davis L and Rose DP: Type 2 diabetes
and obesity metabolic interactions: Common factors for breast
cancer risk and novel approaches to prevention and therapy. Curr
Diabetes Rev. 8:116–130. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zeng L, Biernacka KM, Holly JM, Jarrett C,
Morrison AA, Morgan A, Winters ZE, Foulstone EJ, Shield JP and
Perks CM: Hyperglycaemia confers resistance to chemotherapy on
breast cancer cells: The role of fatty acid synthase. Endocr Relat
Cancer. 17:539–551. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lipscombe LL, Goodwin PJ, Zinman B,
McLaughlin JR and Hux JE: The impact of diabetes on survival
following breast cancer. Breast Cancer Res Treat. 109:389–395.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jiralerspong S, Kim ES, Dong W, Feng L,
Hortobagyi GN and Giordano SH: Obesity, diabetes and survival
outcomes in a large cohort of early-stage breast cancer patients.
Ann Oncol. 24:2506–2514. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hou G, Zhang S, Zhang X, Wang P, Hao X and
Zhang J: Clinical pathological characteristics and prognostic
analysis of 1,013 breast cancer patients with diabetes. Breast
Cancer Res Treat. 137:807–816. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Novosyadlyy R, Lann DE, Vijayakumar A,
Rowzee A, Lazzarino DA, Fierz Y, Carboni JM, Gottardis MM, Pennisi
PA, Molinolo AA, et al: Insulin-mediated acceleration of breast
cancer development and progression in a nonobese model of type 2
diabetes. Cancer Res. 70:741–751. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gallagher EJ and LeRoith D: The
proliferating role of insulin and insulin-like growth factors in
cancer. Trends Endocrinol Metab. 21:610–618. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rostoker R, Abelson S, Bitton-Worms K,
Genkin I, Ben-Shmuel S, Dakwar M, Orr ZS, Caspi A, Tzukerman M and
LeRoith D: Highly specific role of the insulin receptor in breast
cancer progression. Endoc-Relat Cancer. 22:145–157. 2015.
View Article : Google Scholar
|
|
15
|
Michels KB, Solomon CG, Hu FB, Rosner BA,
Hankinson SE, Colditz GA and Manson JE: Nurses' Health Study: Type
2 diabetes and subsequent incidence of breast cancer in the Nurses'
Health Study. Diabetes care. 26:1752–1758. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mawson A, Lai A, Carroll JS, Sergio CM,
Mitchell CJ and Sarcevic B: Estrogen and insulin/IGF-1
cooperatively stimulate cell cycle progression in MCF-7 breast
cancer cells through differential regulation of c-Myc and cyclin
D1. Mol Cell Endocrinol. 229:161–173. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rose DP and Vona-Davis L: The cellular and
molecular mechanisms by which insulin influences breast cancer risk
and progression. Endocr-Relat Cancer. 19:R225–R241. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li Y, Wang K, Zou QY, Magness RR and Zheng
J: 2,3,7,8-Tetrachlorodibenzo-p-dioxin differentially suppresses
angiogenic responses in human placental vein and artery endothelial
cells. Toxicology. 336:70–78. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Schneider CA, Rasband WS and Eliceiri KW:
NIH Image to ImageJ: 25 years of image analysis. Nat Methods.
9:671–675. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li Y, Wang K, Jiang YZ, Chang XW, Dai CF
and Zheng J: 2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD) inhibits
human ovarian cancer cell proliferation. Cellular oncology
(Dordrecht). 37:429–437. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang K, Li Y, Jiang YZ, Dai CF, Patankar
MS, Song JS and Zheng J: An endogenous aryl hydrocarbon receptor
ligand inhibits proliferation and migration of human ovarian cancer
cells. Cancer letters. 340:63–71. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li Y, Zhao YJ, Zou QY, Zhang K, Wu YM,
Zhou C, Wang K and Zheng J: Preeclampsia does not alter vascular
growth and expression of CD31 and vascular endothelial cadherin in
human placentas. J Histochem Cytochem. 63:22–31. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li HH, Zhao YJ, Li Y, Dai CF, Jobe SO,
Yang XS, Li XF, Patankar MS, Magness RR and Zheng J: Estradiol 17β
and its metabolites stimulate cell proliferation and antagonize
ascorbic acid-suppressed cell proliferation in human ovarian cancer
cells. Reprod Sci. 21:102–111. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shaw LM: The insulin receptor substrate
(IRS) proteins: at the intersection of metabolism and cancer. Cell
Cycle. 10:1750–1756. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pollak M: Insulin and insulin-like growth
factor signalling in neoplasia. Nat Rev Cancer. 8:915–928. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Papa V and Belfiore A: Insulin receptors
in breast cancer: Biological and clinical role. J Endocrinol
Invest. 19:324–333. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Belfiore A, Frittitta L, Costantino A,
Frasca F, Pandini G, Sciacca L, Goldfine ID and Vigneri R: Insulin
receptors in breast cancer. Ann N Y Acad Sci. 784:173–188. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hamanaka RB and Chandel NS: Targeting
glucose metabolism for cancer therapy. J Exp Med. 209:211–215.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bergmann U, Funatomi H, Kornmann M, Beger
HG and Korc M: Increased expression of insulin receptor substrate-1
in human pancreatic cancer. Biochem Biophys Res Commun.
220:886–890. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chang Q, Li Y, White MF, Fletcher JA and
Xiao S: Constitutive activation of insulin receptor substrate 1 is
a frequent event in human tumors: Therapeutic implications. Cancer
Res. 62:6035–6038. 2002.PubMed/NCBI
|
|
32
|
Sisci D, Morelli C, Garofalo C, Romeo F,
Morabito L, Casaburi F, Middea E, Cascio S, Brunelli E, Andò S and
Surmacz E: Expression of nuclear insulin receptor substrate 1 in
breast cancer. J Clin Pathol. 60:633–641. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Molloy CA, May FE and Westley BR: Insulin
receptor substrate-1 expression is regulated by estrogen in the
MCF-7 human breast cancer cell line. J Biol Chem. 275:12565–12571.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mauro L, Salerno M, Panno ML, Bellizzi D,
Sisci D, Miglietta A, Surmacz E and Andò S: Estradiol increases
IRS1 gene expression and insulin signaling in breast cancer cells.
Biochem Biophys Res Commun. 288:685–689. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee AV, Jackson JG, Gooch JL, Hilsenbeck
SG, Coronado-Heinsohn E, Osborne CK and Yee D: Enhancement of
insulin-like growth factor signaling in human breast cancer:
Estrogen regulation of insulin receptor substrate-1 expression in
vitro and in vivo. Mol Endocrinol. 13:787–796. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
de Blaquiere GE, May FE and Westley BR:
Increased expression of both insulin receptor substrates 1 and 2
confers increased sensitivity to IGF-1 stimulated cell migration.
Endocr Relat Cancer. 16:635–647. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cesarone G, Garofalo C, Abrams MT,
Igoucheva O, Alexeev V, Yoon K, Surmacz E and Wickstrom E:
RNAi-mediated silencing of insulin receptor substrate 1 (IRS1)
enhances tamoxifen-induced cell death in MCF-7 breast cancer cells.
J Cell Biochem. 98:440–450. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Dearth RK, Cui X, Kim HJ, Kuiatse I,
Lawrence NA, Zhang X, Divisova J, Britton OL, Mohsin S, Allred DC,
et al: Mammary Tumorigenesis and Metastasis Caused by
Overexpression of Insulin Receptor Substrate 1 (IRS1) or IRS-2. Mol
Cell Biol. 26:9302–9314. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Raman M, Chen W and Cobb MH: Differential
regulation and properties of MAPKs. Oncogene. 26:3100–3112. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Weng LP, Smith WM, Brown JL and Eng C:
PTEN inhibits insulin-stimulated MEK/MAPK activation and cell
growth by blocking IRS1 phosphorylation and IRS1/Grb-2/Sos complex
formation in a breast cancer model. Hum Mol Genet. 10:605–616.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Whyte J, Bergin O, Bianchi A, McNally S
and Martin F: Key signalling nodes in mammary gland development and
cancer. Mitogen-activated protein kinase signalling in experimental
models of breast cancer progression and in mammary gland
development. Breast Cancer Res. 11:2092009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Santen RJ, Song RX, McPherson R, Kumar R,
Adam L, Jeng MH and Yue W: The role of mitogen-activated protein
(MAP) kinase in breast cancer. J Steroid Biochem Mol Biol.
80:239–256. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Adeyinka A, Nui Y, Cherlet T, Snell L,
Watson PH and Murphy LC: Activated mitogen-activated protein kinase
expression during human breast tumorigenesis and breast cancer
progression. Clin Cancer Res. 8:1747–1753. 2002.PubMed/NCBI
|