Introduction
Osteosarcoma (OS) is the most common
non-haematological primary bone tumour that affects children and
young adolescents, and pulmonary metastases are the major cause of
mortality (1–3). In total, ~20% of patients present
with pulmonary metastasis at initial diagnosis, and in ~40% of
patients, metastases occur in the advanced stage (4). Furthermore, OS has a low survival
rate in patients due to chemoresistance and the high rates of
pulmonary metastasis (5). For
patients with non-metastatic OS, 5-year OS has increased to 60–70%,
but this rate is reduced to 20% when metastases occur (6). Therefore, strategies to prevent
tumour metastasis are urgently needed. However, the molecular
mechanisms of pulmonary metastasis in patients with OS remain
poorly understood. Thus, identifying key molecules associated with
pulmonary metastasis of OS is important for the development of more
effective metastasis-suppressive therapies.
During the last decade, gene expression profiling
has identified critical genes and cellular signaling pathways
related to the occurrence, development and change of human OS
(7). Microarray techniques
combined with bioinformatics analysis can determine the
differential expression levels of genes accurately and provide an
effective tool for large-scale gene expression studies (8). In the patients with OS, the lung is
the most common metastatic site, and metastases can lead to a high
rate of mortality (9). Therefore,
examining differentially expressed genes (DEGs) between
non-metastatic and metastatic OS samples is helpful for identifying
the key genes and pathways leading to pulmonary metastasis.
In the current study, by comparing the gene
expression of non-metastatic and metastatic OS samples in the GEO
database, the authors identified DEGs that may be related to
pulmonary metastasis. Subsequently, Gene Ontology (GO) and Kyoto
Encyclopedia of Genes and Genomes (KEGG) enrichment pathway
analyses were performed. In combination with protein-protein
interaction (PPI) information, not only did the authors identify
relevant genes and pathways, but also revealed existing molecular
mechanisms. In conclusion, the authors' finding can improve the
understanding of OS and identify the genes associated with
pulmonary metastasis, thereby providing potential therapeutic
targets for further studies.
Materials and methods
Gene expression microarray data
In the present study, the gene expression profiles
of GSE85537 were downloaded from Gene Expression Omnibus (GEO,
http://www.ncbi.nlm.nih.gov/geo/).
GSE85537 was based on the Affymetrix GPL570 platform (Affymetrix
Human Genome U133 Plus 2.0 Array; Affymetrix; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). The GSE85537 dataset contained
six samples, including three non-metastatic OS samples and three
metastatic OS samples.
Identification of DEGs
The raw data files used for the analysis included
TXT files. The analysis was carried out using GEO2R (https://www.ncbi.nlm.nih.gov/geo/geo2r/), which can
perform comparisons on original submitter-supplied processed data
tables using the GEOquery and limma R packages from the
Bioconductor project. The DEGs between the non-metastatic and
metastatic OS samples were selected (P<0.05), and overlapped
genes were identified.
GO enrichment and KEGG pathway
analysis of the DEGs
After obtaining the DEGs, the authors submitted the
DEG list to the online software Database for Annotation,
Visualization and Integrated Discovery (DAVID, https://david.ncifcrf.gov/) to identify
overrepresented GO categories and pathway categories. GO analysis
can determine the biological meaning in a large list of genes and
categorize gene product functions, including biological process
(BP), molecular function (MF) and cellular component (CC) (10,11).
KEGG (http://www.genome.jp/) is a knowledge
base for systematic analysis of gene functions, linking genomic
information with higher-level systemic functions (12,13).
Finally, the enriched functions of DEGs were selected via GO and
KEGG pathway analysis, and P<0.05 was considered to indicate a
statistically significant difference.
Construction of the PPI Network of
DEGs
To further investigate the molecular mechanism of
pulmonary metastasis in patients with OS, the Search Tool for the
Retrieval of Interacting Genes (STRING) database (http://www.string-db.org/) was used to evaluate the
interactive relationships among DEGs. Initially, the DEG list was
first submitted to STRING, and then, the experimentally validated
interactions were selected with a combined score >0.4.
Subsequently, the PPI networks were analyzed using Cytoscape
software (version 3.5.1; www.cytoscape.org). Then, the plug-in Molecular
Complex Detection (MCODE) was used to screen the modules of the PPI
network in Cytoscape. Furthermore, the enrichment analyses were
performed for DEGs in the corresponding modules. P<0.05 was
considered to indicate a statistically significant difference.
Results
Identification of DEGs
The gene expression profile GSE85537 was downloaded
from the GEO database, and the GEO2R method was used to identify
DEGs in metastatic OS samples compared with non-metastatic OS
samples. P<0.05, logFC (fold control) >2.0 or logFC <-2.0
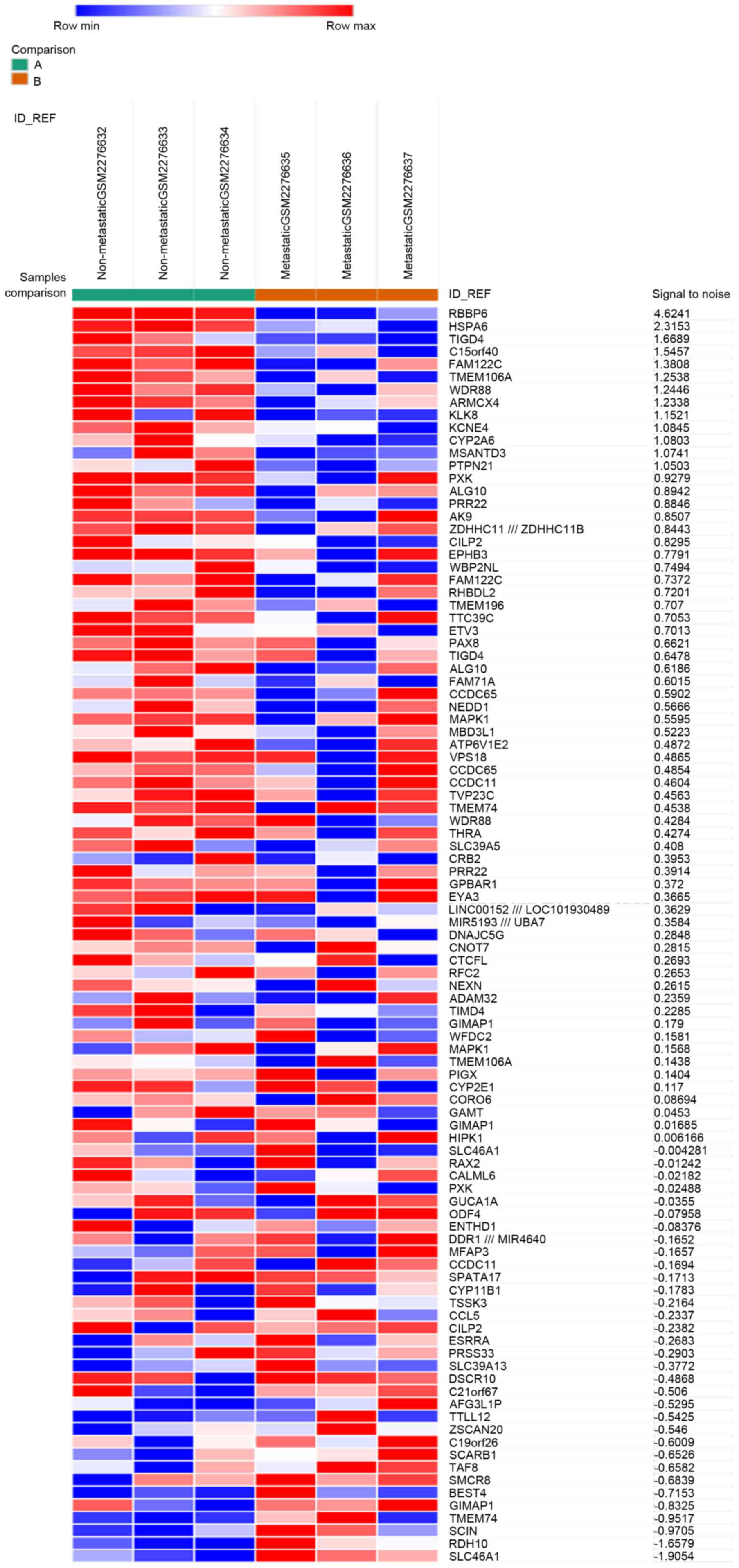
was used as the criteria, and 2,493 genes were identified as DEGs.
Among these, 485 genes (19.45%) were upregulated, and the remaining
2,008 genes (80.55%) were downregulated. Subsequently, the top 50
upregulated and downregulated DEGs were selected to generate the
heatmap (Fig. 1).
GO term enrichment analysis
To functionally categorize these 2,493 significant
genes, the DEGs were analyzed using the online software DAVID. GO
analysis revealed that the upregulated DEGs were significantly
enriched in BP, including transmembrane transport, ion transport,
and release of sequestered calcium ion into cytosol (Table I); the downregulated DEGs were
significantly enriched in BP, including cell surface receptor
signaling pathway, actin filament-based process and movement of
cell or subcellular component (Table
I). For MF, the upregulated DEGs were enriched in
ATPase-coupled ion transmembrane transporter activity and
cation-transporting ATPase activity, and the downregulated DEGs
were enriched in Ras guanyl-nucleotide exchange factor activity,
guanyl-nucleotide exchange factor activity, and growth factor
binding (Table I). In addition, GO
CC analysis also indicated that the upregulated DEGs were
significantly enriched in the synapse, integral component of plasma
membrane and intrinsic component of plasma membrane, and
downregulated DEGs were enriched in the proteinaceous extracellular
matrix, extracellular matrix and plasma membrane region (Table I).
 | Table I.Gene ontology analysis of DEGs. |
Table I.
Gene ontology analysis of DEGs.
| Expression | Category | GO-ID | Term | Gene count | % | P-value |
|---|
| Upregulated | BP | GO:0055085 | Transmembrane
transport | 43 | 12.6 |
5.10×10−6 |
|
| BP | GO:0006811 | Ion transport | 45 | 13.2 |
1.40×10−5 |
|
| BP | GO:0051209 | Release of
sequestered calcium ion into cytosol | 10 | 2.9 |
3.20×10−5 |
|
| BP | GO:0051283 | Negative regulation
of sequestering of calcium ion | 10 | 2.9 |
3.20×10−5 |
|
| BP | GO:0097553 | Calcium ion
transmembrane import into cytosol | 10 | 2.9 |
3.50×10−5 |
|
| MF | GO:0042625 | ATPase coupled ion
transmembrane transporter activity | 7 | 2.0 |
1.10×10−3 |
|
| MF | GO:0019829 | Cation-transporting
ATPase activity | 6 | 1.8 |
2.90×10−3 |
|
| MF | GO:0022853 | Active ion
transmembrane transporter activity | 7 | 2.0 |
6.10×10−3 |
|
| MF | GO:0042626 | ATPase activity,
coupled to transmembrane movement of substances | 7 | 2.0 |
7.20×10−3 |
|
| MF | GO:0070330 | Aromatase
activity | 4 | 1.2 |
7.50×10−3 |
|
| CC | GO:0045202 | Synapse | 30 | 8.8 |
5.30×10−6 |
|
| CC | GO:0005887 | Integral component
of plasma membrane | 46 | 13.5 |
5.10X10−5 |
|
| CC | GO:0031226 | Intrinsic component
of plasma membrane | 46 | 13.5 |
1.30×10−4 |
|
| CC | GO:0098794 | Postsynapse | 17 | 5.0 |
1.50×10−4 |
|
| CC | GO:0044456 | Synapse part | 23 | 6.7 |
1.50×10−4 |
| Downregulated | BP | GO:0007166 | Cell surface
receptor signaling pathway | 226 | 17.2 |
2.00×10−9 |
|
| BP | GO:0030029 | Actin
filament-based process | 79 | 6.0 |
4.30×10−9 |
|
| BP | GO:0006928 | Movement of cell or
subcellular component | 165 | 12.5 |
4.30×10−9 |
|
| BP | GO:2000145 | Regulation of cell
motility | 85 | 6.5 |
1.10×10−8 |
|
| BP | GO:0040012 | Regulation of
locomotion | 87 | 6.6 |
1.70×10−8 |
|
| MF | GO:0005088 | Ras
guanyl-nucleotide exchange factor activity | 34 | 2.6 |
1.10×10−6 |
|
| MF | GO:0005085 | Guanyl-nucleotide
exchange factor activity | 41 | 3.1 |
1.30×10−6 |
|
| MF | GO:0019838 | Growth factor
binding | 23 | 1.7 |
4.00×10−6 |
|
| MF | GO:0005102 | Receptor
binding | 126 | 9.6 |
4.90×10−6 |
|
| MF | GO:0004713 | Protein tyrosine
kinase activity | 26 | 2.0 |
4.20×10−5 |
|
| CC | GO:0005578 | Proteinaceous
extracellular matrix | 55 | 4.2 |
3.90×10−10 |
|
| CC | GO:0031012 | Extracellular
matrix | 67 | 5.1 |
1.10×10−8 |
|
| CC | GO:0098590 | Plasma membrane
region | 101 | 7.7 |
1.20×10−8 |
|
| CC | GO:0044421 | Extracellular
region part | 295 | 22.4 |
2.00×10−7 |
|
| CC | GO:0005576 | Extracellular
region | 333 | 25.3 |
4.00×10−6 |
KEGG pathway analysis
KEGG analysis revealed that the upregulated DEGs
were enriched in focal adhesion, ovarian steroidogenesis, Rap1
signaling pathway, chemokine signaling pathway and PI3K-Akt
signaling pathway, while the downregulated DEGs were enriched in
the Rap1 signaling pathway, pathways in cancer, platelet
activation, rheumatoid arthritis and cAMP signaling pathway
(Table II).
| Table II.KEGG pathway analysis of DEGs. |
Table II.
KEGG pathway analysis of DEGs.
| Expression | Pathway-ID | Name | Gene count | % | P-value | Genes |
|---|
| Upregulated | 4510 | Focal adhesion | 12 | 3.5 |
9.20×10−4 | RASGRF1, ROCK1,
SHC4, COL4A3, COL4A4, COL6A5, EGFR, FLT1, HGF, IGF1R, SPP1,
VAV2 |
|
| 4913 | Ovarian
steroidogenesis | 6 | 1.8 |
1.60×10−3 | CYP1A1, CYP19A1,
CGA, HSD3B1, IGF1R, INS |
|
| 4015 | Rap1 signaling
pathway | 11 | 3.2 |
3.60×10−3 | GNAO1, RASGRP2,
EGFR, FGF22, FGFR4, FLT1, HGF, IGF1R, INS, LPAR4, PLCB2 |
|
| 4062 | Chemokine signaling
pathway | 9 | 2.6 |
1.60×10−2 | CCL16, CCL25, GRK4,
RASGRP2, ROCK1, SHC4, PLCB2, PPBP, VAV2 |
|
| 4151 | PI3K-Akt signaling
pathway | 13 | 3.8 |
1.70×10−2 | COL4A3, COL4A4,
COL6A5, CSF3R, EGFR, FGF22, FGFR4, FLT1, HGF, IGF1R, INS, LPAR4,
SPP1 |
| Downregulated | 4015 | Rap1 signaling
pathway | 31 | 2.4 |
2.50×10−6 | GNAO1, KITLG,
RAPGEF5, ADCY1, APBB1IP, CDH1, CNR1, F2R, CSF1, DRD2, EGFR, FGF14,
FGF20, FGF7, FGFR2, IGF1R, IGF1, ITGB1, ITGB3, KDR, LCP2, MAGI1,
MAP2K6, PIK3CD, PLCB1, PDGFRB, PFN4, PRKD2, RAC2, SIPA1L2,
TLN1 |
|
| 5200 | Pathways in
cancer | 46 | 3.5 |
4.50×10−6 | BAD, BCR, CXCL12,
CXCL8, CXCR4, FAS, GNG11, KITLG, RUNX1T1, ARHGEF12, SPI1, WNT5B,
ADCY1, AGTR1, CDH1, F2R, COL4A1, COL4A2, CDK6, EPAS1, EGFR, ERBB2,
FGF14, FGF20, FGF7, FGFR2, FZD4, FZD5, IGF1R, IGF1, ITGB1, IL6,
LAMA2, LAMB1, MMP2, MAPK10, NFKB2, PTCH1, PIK3CD, PLCB1, PDGFRB,
PML, PTGER3, PIAS2, RAC2, TPM3 |
|
| 4611 | Platelet
activation | 19 | 1.4 |
3.90×10−4 | FYN, ARHGEF12,
ADCY1, APBB1IP, F2R, COL1A2, COL3A1, COL5A3, GUCY1A3, GUCY1B3,
ITGB1, ITGB3, LCP2, PIK3CD, PLA2G4C, PLCB1, PTGS1, TLN1,
TBXAS1 |
|
| 5323 | Rheumatoid
arthritis | 15 | 1.1 |
4.20×10−4 | ATP6V0D2, ATP6V0E1,
ATP6V1B1, CCL2, CXCL12, CXCL8, CD86, CTSK, CSF1, IL1B, IL11, IL6,
HLA-DOA, HLA-DRB4, TNFSF13 |
|
| 4024 | cAMP signaling
pathway | 24 | 1.8 |
8.60×10−4 | HTR1D, HTR1F,
ABCC4, ATP1B4, ATP1B2, BAD, ADCY1, CAMK2A, CHRM2, F2R, DRD2, GIPR,
GRIN3A, MC2R, MAPK10, PTCH1, PPARA, PIK3CD, PDE3A, PDE4D, PLN,
PLD1, PTGER3, RAC2 |
PPI network of the DEGs and core genes
in the PPI network
Based on the information in the STRING database, the
PPI network contained 1,123 nodes and 4,986 edges. The nodes
indicate the DEGs, and the edges indicate the interactions between
the DEGs. The top 10 high-degree hub nodes included albumin (ALB),
epidermal growth factor receptor (EGFR), insulin (INS), interleukin
6 (IL-6), cadherin 1 (CDH1), FYN proto-oncogene (FYN), erb-b2
receptor tyrosine kinase 2 (ERBB2), interleukin 8 (IL8), C-X-C
motif chemokine ligand 12 (CXCL12) and Ras-related C3 botulinum
toxin substrate 2 (RAC2). Among these genes, ALB presented the
highest node degree, which was 132. The core genes and their
corresponding degree are shown in Table III. Then, the authors used MCODE
to screen the modules of the PPI network (Fig. 2), and performed an enrichment
analysis of the genes involved in the top three significant
modules. The results demonstrated that the DEGs in modules 1–3 were
principally related to neuroactive ligand-receptor interaction, the
Rap1 signaling pathway, and protein digestion and absorption
(Table IV).
| Table III.The core genes and their
corresponding degree. |
Table III.
The core genes and their
corresponding degree.
| Gene | Degree |
|---|
| ALB | 132 |
| EGFR | 113 |
| INS | 107 |
| IL6 | 77 |
| CDH1 | 76 |
| FYN | 72 |
| ERBB2 | 71 |
| IL8 | 67 |
| CXCL12 | 62 |
| RAC2 | 57 |
| GNAO1 | 55 |
| MMP2 | 54 |
| TOP2A | 54 |
| EDN1 | 53 |
| MYH14 | 52 |
| ITGB1 | 50 |
| ACTA2 | 50 |
| KDR | 49 |
| IGF1 | 48 |
| PPARA | 48 |
| Table IV.The enriched pathways of modules. |
Table IV.
The enriched pathways of modules.
| Modules | Enriched
pathways | P-value | False discovery
rate | Nodes |
|---|
| 1 | Neuroactive
ligand-receptor interaction |
9.60×10−10 |
2.68×10−14 | ADORA3, CHRM2,
CHRM4, CNR1, DRD2, GRM3, GRM6, HTR1D, HTR1F, PTGER3, S1PR1,
S1PR5 |
|
| Chemokine
signaling |
6.10×10−6 |
2.03×10−7 | CCL16, CCL25,
CXCL12, CXCL2, CXCR4, IL8, PPBP |
|
|
Cytokine-cytokine |
5.00×10−4 |
1.87×10−6 | CCL16, CCL25,
CXCL12, CXCL2, CXCR4, IL8, PPBP |
| 2 | Rap1 signaling
pathway |
2.40×10-16 |
5.46×10−21 | CDH1, EGFR, F2R,
FGF7, FLT1, GNAO1, HGF, IGF1, IGF1R, INS, KDR, LPAR4, PDGFRB,
PLCB1, PLCB2 |
|
| PI3K-Akt signaling
pathway |
3.20×10−9 |
1.09×10−12 | EGFR, F2R, FGF7,
FLT1, HGF, IGF1, IGF1R, IL6, INS, KDR, LPAR4, PDGFRB |
|
| Calcium signaling
pathway |
6.10×10−8 |
1.09×10−10 | AGTR1, EGFR, ERBB2,
F2R, GRPR, PDGFRB, PLCB1, PLCB2, PTGFR |
| 3 | Protein digestion
and absorption |
4.30×10−14 |
1.48×10−16 | COL1A2, COL21A1,
COL3A1, COL4A1, COL4A2, COL4A3, COL4A4, COL5A3 |
|
| ECM-receptor
interaction |
2.30×10−11 |
8.52×10−14 | COL1A2, COL3A1,
COL4A1, COL4A2, COL4A3, COL4A4, COL5A3 |
|
| Amoebiasis |
7.80×10−11 |
2.44×10−13 | COL1A2, COL3A1,
COL4A1, COL4A2, COL4A3, COL4A4, COL5A3 |
Discussion
Osteosarcoma is the most common primary malignant
bone tumour, is commonly observed in children and adolescents and
shows a strong tendency for pulmonary metastasis (14). Pulmonary metastasis can cause
medical therapy failure and high mortality rate in osteosarcoma
patients (15,16). However, the present knowledge of
the molecular mechanism of pulmonary metastasis in patients with OS
remains insufficient. Therefore, the elucidation of molecular
mechanisms to inhibit the metastasis of OS is imperative.
High-throughput profiling technologies, such as microarrays, have
been widely used and regarded as invaluable tools to identify
potential therapeutic targets (17,18).
The present study performed a comprehensive analysis and built a
gene interaction network based on the gene expression profiles
(GSE85537), comprising three non-metastatic OS samples and three
metastatic OS samples. The results of the analysis demonstrated
that there were 2,493 DEGs in the metastatic OS tumour samples
compared with non-metastatic OS tumour samples, and among these
DEGs, 485 were upregulated and 2,008 were downregulated. Moreover,
GO and KEGG pathway analyses were performed to obtain a better
understanding of the interactions of DEGs. Combining with the PPI
network, key potential genes and pathways that may be associated
with pulmonary metastasis of OS were identified.
The results of GO analyses indicated that the
significant ontology categories included transmembrane transport,
ion transport and aromatase activity. The system of transport is
essential to every living cell and it had multiple functions,
including allowing the entry to all essential molecules, exporting
macromolecules such as proteins and DNA, providing cellular
concentrations of ions (19,20).
In a recent study on papillary thyroid carcinoma progression,
downregulated DEGs were also predominantly enriched in
transmembrane transport process (21). Additionally, ion transport system
also stimulates the progression of pithelial-mesenchymal transition
(EMT) in cervical carcinoma cells (22). Therefore, transmembrane transport
and ion transport are fundamental processes and may serve roles in
pulmonary metastasis of OS. Moreover, aromatase is an enzyme that
converts testosterone into estradiol, and the control of aromatase
activity mainly depends on Ca2+ transients (23). It has been reported that aromatase
activity is associated with tumour-node-metastasis staging in human
breast cancer (24). Cell motility
is essential in the regulation of cancer progression, and aberrant
cell motility occurs in malignant cancers and results in tumour
metastasis (25). The activation
of ERBB3-dependent signaling serves a key role in the regulation of
cell motility, and regulation of cell motility contributes to
intrahepatic metastasis and early recurrence of hepatocellular
carcinoma (HCC) (26). Cell
motility is also considered as an important process in the
regulation of metastasis of oral cancer (27). Cell motility may therefore be
important to pulmonary metastasis of OS. In addition, many studies
have demonstrated that receptor binding is closely correlated with
tumour metastasis, and these receptor bindings include fibroblast
growth factor and vascular endothelial growth factor receptor
binding, duffy antigen receptor for chemokines and growth factor
receptor bound protein 2 Src homology 2 domain binding (28–30).
Therefore, GO analyses can help identify the possible biological
processes, molecular functions and cellular components involved in
tumour metastasis.
Moreover, the KEGG pathways revealed that DEGs were
enriched in focal adhesion, platelet activation, the Rap1 signaling
pathway and the PI3K-Akt signaling pathway. Focal adhesion is a
dynamic multi-protein complex that connects intracellular actin
fibres and extracellular substrates (31,32).
Focal adhesion kinase (FAK) is an effective therapeutic target of
cell migration in various tumour cells (33–35).
Furthermore, Rap1 is considered as a protein that can revert Ras
transformation, and it is a key mediator in the control of integrin
activation (36). A recent study
revealed that the cAMP/Epac/Rap1 signaling cascade had a crucial
role in blood-tumour barrier hyperpermeability (37). Cancer cell-induced platelet
activation is identified as a critical event responsible for
prometastatic activity of platelets, and blocking platelet
aggregation can inhibit the progression of skeletal metastases, yet
the mechanism underlying this effect is still unknown (38). Moreover, the PI3 K/Akt signaling
pathway is involved in various cellular processes, such as
inflammation, autophagy and cancer progression (39), and many studies have demonstrated
that activation of the PI3K-Akt signaling pathway can increase the
metastatic potential (40–42). Therefore, further understanding of
these signaling pathways can help us to elucidate the crucial
mechanism of tumour metastasis.
Furthermore, the authors analyzed the PPI network
and found that ALB, EGFR, INS, IL6, CDH1, FYN, ERBB2, IL8, CXCL12
and RAC2 were the top 10 core genes, which may be potential
therapeutic targets for pulmonary metastasis of OS. Node degree
refers to the number of interacting partners per protein. Among
these genes, ALB showed the highest node degree. Albumin is one of
the major plasma proteins and is synthesized and secreted primarily
by hepatocytes, and it is often used for diagnosis of HCC (43). Albumin is a major indicator of a
favourable outcome of HCC, and it can suppress cell proliferation
via reduction of phosphorylation of Rb (retinoblastoma) proteins
and increase of p21 and p57 (44).
A recent study indicated that the expression of albumin increased
in human hepatoma BEL-7402 cells after bufalin was used, and ALB
may be associated with bufalin-mediated inhibition of the invasion
and metastasis of HCC cells (45).
Albumin mRNA was also found in lung by means of
reverse-transcription polymerase chain reaction, so albumin may
serve a role in pulmonary metastasis (46). The present study indicated that ALB
was a key gene with the highest node degree in OS, therefore ALB
may be a potential target for inhibiting OS tumour metastasis, but
further experimental verification is necessary.
The EGFR is a key component in the mitogen-activated
protein kinase (MAPK) pathway, and anti-EGFR therapy can lead to
inhibition of tumour growth, invasion and metastasis via inhibition
of the MAPK pathway (47).
Moreover, nasopharyngeal carcinoma (NPC), which has the highest
rate of metastasis, has been extensively studied, and inhibition of
EGFR can inhibit NPC cell migration and invasion (48). Additionally, EGFR has been shown to
promote survival of prostate tumour-initiating cells and
circulating tumour cells that metastasize to bone (49). These results corresponded with the
function analysis in the present study, and EGFR is likely to be an
important regulated target in pulmonary metastasis of OS.
Therefore, EGFR may be a potential therapeutic target and
prognostic indicator in OS patients, and a thorough study of EGFR
in OS is needed. Furthermore, a recent study demonstrated that
inflammatory cytokines, such as IL-6, promoted proliferation,
migration, invasion and EMT of gallbladder cancer both in
vitro and in vivo (50). Overexpression of human epidermal
growth factor receptor 2 (ErbB2 or HER2) occurs in ~30% of breast
cancer patients, and overexpression of activated ErbB2 in the
mammary epithelium can lead to the rapid induction of metastatic
multifocal mammary tumours (51,52).
Therefore, these core genes in the current study have been
suggested to be associated with tumour metastasis in the previous
studies, so the analysis of these core genes is useful for
understanding the molecular mechanisms and identifying therapeutic
targets of OS with pulmonary metastasis. However, subsequent
prospective studies will be required to further confirm the
function of these core genes in pulmonary metastasis of OS.
Module analysis of the PPI network demonstrated that
the pulmonary metastasis of OS was associated with neuroactive
ligand-receptor interaction, the Rap1 signaling pathway, protein
digestion and absorption and other processes. It has been
demonstrated that neuroactive ligand-receptor interaction is
important in HCC progression and it is present in the early-,
middle- and late-stages of HCC (53,54).
Moreover, the neuroactive ligand-receptor interaction pathway was
enriched in prostate tumours from African-American patients
(55). Therefore, neuroactive
ligand-receptor interaction appears to be important in pulmonary
metastasis of OS. Moreover, the protein digestion and absorption
pathway has been reported to be associated with pancreatic
neuroendocrine tumours and breast cancer (56,57).
Thus, the roles of these pathways in pulmonary metastasis of OS
need to be confirmed by the future studies, and these results may
lead to additional therapeutic alternatives in the patients with
OS.
In conclusion, the present study identified 2,493
DEGs, which may be involved in the progress of pulmonary metastasis
in OS patients, via a comprehensive bioinformatics analysis. GO
term, KEGG pathway and PPI network analyses provided a set of
related genes and pathways to help elucidate the molecular
mechanisms of pulmonary metastasis. Further experimental studies
are needed to confirm these results and should help determine
potential targets for inhibiting pulmonary metastasis in OS
patients.
Acknowledgements
The present study was supported by the State Key
Program of National Natural Science Foundation of China (grant no.
81330042), International Cooperation Program of National Natural
Science Foundation of China (grant no. 81620108018), Special
Program for Sino-Russian Joint Research Sponsored by the Ministry
of Science and Technology, China (grant no. 2014DFR31210) and Key
Program Sponsored by the Tianjin Science and Technology Committee,
China (grant nos. 13RCGFSY19000 and 14ZCZDSY00044).
References
|
1
|
Klein MJ and Siegal GP: Osteosarcoma:
Anatomic and histologic variants. Am J Clin Pathol. 125:555–581.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Neklyudova O, Arlt MJ, Brennecke P, Thelen
M, Gvozdenovic A, Kuzmanov A, Robl B, Botter SM, Born W and Fuchs
B: Altered CXCL12 expression reveals a dual role of CXCR4 in
osteosarcoma primary tumor growth and metastasis. J Cancer Res Clin
Oncol. 142:1739–1750. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zandueta C, Ormazábal C, Perurena N,
Martínez-Canarias S, Zalacaín M, Julián MS, Grigoriadis AE,
Valencia K, Campos-Laborie FJ, Jde L Rivas, et al: Matrix-Gla
protein promotes osteosarcoma lung metastasis and associates with
poor prognosis. J Pathol. 239:438–449. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Qu L and Liu B: Cyclooxygeanse-2 promotes
metastasis in osteosarcoma. Cancer Cell Int. 15:692015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Weekes D, Kashima TG, Zandueta C, Perurena
N, Thomas DP, Sunters A, Vuillier C, Bozec A, El-Emir E, Miletich
I, et al: Regulation of osteosarcoma cell lung metastasis by the
c-Fos/AP-1 target FGFR1. Oncogene. 35:2852–2861. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gorlick R, Anderson P, Andrulis I, Arndt
C, Beardsley GP, Bernstein M, Bridge J, Cheung NK, Dome JS, Ebb D,
et al: Biology of childhood osteogenic sarcoma and potential
targets for therapeutic development: Meeting summary. Clin Cancer
Res. 9:5442–5453. 2003.PubMed/NCBI
|
|
7
|
Selvarajah GT, Kirpensteijn J, van
Wolferen ME, Rao NA, Fieten H and Mol JA: Gene expression profiling
of canine osteosarcoma reveals genes associated with short and long
survival times. Mol Cancer. 8:722009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wolf M, El-Rifai W, Tarkkanen M, Kononen
J, Serra M, Eriksen EF, Elomaa I, Kallioniemi A, Kallioniemi OP and
Knuutila S: Novel findings in gene expression detected in human
osteosarcoma by cDNA microarray. Cancer Genet Cytogenet.
123:128–132. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang N, Xie T, Xian M, Wang YJ, Li HY,
Ying MD and Ye ZM: SIRT1 promotes metastasis of human osteosarcoma
cells. Oncotarget. 7:79654–79669. 2016.PubMed/NCBI
|
|
10
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The Gene
Ontology Consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lammers G, Gilissen C, Nillesen ST,
Uijtdewilligen PJ, Wismans RG, Veltman JA, Daamen WF and van
Kuppevelt TH: High density gene expression microarrays and gene
ontology analysis for identifying processes in implanted tissue
engineering constructs. Biomaterials. 31:8299–8312. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wixon J and Kell D: The Kyoto encyclopedia
of genes and genomes-KEGG. Yeast. 17:48–55. 2000.PubMed/NCBI
|
|
14
|
Lu J, Song G, Tang Q, Zou C, Han F, Zhao
Z, Yong B, Yin J, Xu H, Xie X, et al: IRX1 hypomethylation promotes
osteosarcoma metastasis via induction of CXCL14/NF-κB signaling. J
Clin Invest. 125:1839–1856. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Isakoff MS, Bielack SS, Meltzer P and
Gorlick R: Osteosarcoma: Current treatment and a collaborative
pathway to success. J Clin Oncol. 33:3029–3035. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Siegel HJ and Pressey JG: Current concepts
on the surgical and medical management of osteosarcoma. Expert Rev
Anticancer Ther. 8:1257–1269. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liang B, Li C and Zhao J: Identification
of key pathways and genes in colorectal cancer using bioinformatics
analysis. Med Oncol. 33:1112016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yin F, Shu L, Liu X, Li T, Peng T, Nan Y,
Li S, Zeng X and Qiu X: Microarray-based identification of genes
associated with cancer progression and prognosis in hepatocellular
carcinoma. J Exp Clin Cancer Res. 35:1272016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hollenstein K, Dawson RJ and Locher KP:
Structure and mechanism of ABC transporter proteins. Curr Opin
Struct Biol. 17:412–418. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yen MR, Choi J and Saier MH Jr:
Bioinformatic analyses of transmembrane transport: Novel software
for deducing protein phylogeny, topology and evolution. J Mol
Microbiol Biotechnol. 17:163–176. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Qiu J, Zhang W, Xia Q, Liu F, Li L, Zhao
S, Gao X, Zang C, Ge R and Sun Y: RNA sequencing identifies crucial
genes in papillary thyroid carcinoma (PTC) progression. Exp Mol
Pathol. 100:151–159. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee MY and Shen MR: Epithelial-mesenchymal
transition in cervical carcinoma. Am J Transl Res. 4:1–13.
2012.PubMed/NCBI
|
|
23
|
Fester L, Zhou L, Ossig C, Labitzke J,
Bläute C, Bader M, Vollmer G, Jarry H and Rune GM: Synaptopodin is
regulated by aromatase activity. J Neurochem. 140:126–139. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bolufer P, Ricart E, Lluch A, Vazquez C,
Rodriguez A, Ruiz A, Llopis F, Garcia-Conde J and Romero R:
Aromatase activity and estradiol in human breast cancer: Its
relationship to estradiol and epidermal growth factor receptors and
to tumor-node-metastasis staging. J Clin Oncol. 10:438–446. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hammer A and Diakonova M:
Prolactin-induced PAK1 tyrosyl phosphorylation promotes FAK
dephosphorylation, breast cancer cell motility, invasion and
metastasis. BMC Cell Biol. 17:312016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hsieh SY, He JR, Hsu CY, Chen WJ, Bera R,
Lin KY, Shih TC, Yu MC, Lin YJ, Chang CJ, et al:
Neuregulin/erythroblastic leukemia viral oncogene homolog 3
autocrine loop contributes to invasion and early recurrence of
human hepatoma. Hepatology. 53:504–516. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Noguti J, De Moura CF, De Jesus GP, Da
Silva VH, Hossaka TA, Oshima CT and Ribeiro DA: Metastasis from
oral cancer: An overview. Cancer Genomics Proteomics. 9:329–335.
2012.PubMed/NCBI
|
|
28
|
Aviezer D, Cotton S, David M, Segev A,
Khaselev N, Galili N, Gross Z and Yayon A: Porphyrin analogues as
novel antagonists of fibroblast growth factor and vascular
endothelial growth factor receptor binding that inhibit endothelial
cell proliferation, tumor progression, and metastasis. Cancer Res.
60:2973–2980. 2000.PubMed/NCBI
|
|
29
|
Giubellino A, Gao Y, Lee S, Lee MJ,
Vasselli JR, Medepalli S, Trepel JB, Burke TR Jr and Bottaro DP:
Inhibition of tumor metastasis by a growth factor receptor bound
protein 2 Src homology 2 domain-binding antagonist. Cancer Res.
67:6012–6016. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang J, Ou ZL, Hou YF, Luo JM, Shen ZZ,
Ding J and Shao ZM: Enhanced expression of Duffy antigen receptor
for chemokines by breast cancer cells attenuates growth and
metastasis potential. Oncogene. 25:7201–7211. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Case LB and Waterman CM: Integration of
actin dynamics and cell adhesion by a three-dimensional,
mechanosensitive molecular clutch. Nat Cell Biol. 17:955–963.
2015.PubMed/NCBI
|
|
32
|
Sulzmaier FJ, Jean C and Schlaepfer DD:
FAK in cancer: Mechanistic findings and clinical applications. Nat
Rev Cancer. 14:598–610. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kishi T, Mayanagi T, Iwabuchi S, Akasaka T
and Sobue K: Myocardin-related transcription factor A (MRTF-A)
activity-dependent cell adhesion is correlated to focal adhesion
kinase (FAK) activity. Oncotarget. 7:72113–72130. 2016.PubMed/NCBI
|
|
34
|
Mendoza P, Ortiz R, Díaz J, Quest AF,
Leyton L, Stupack D and Torres VA: Rab5 activation promotes focal
adhesion disassembly, migration and invasiveness in tumor cells. J
Cell Sci. 126:3835–3847. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Salem I, Alsalahi M, Chervoneva I, Aburto
LD, Addya S, Ott GR, Ruggeri BA, Cristofanilli M and Fernandez SV:
The effects of CEP-37440, an inhibitor of focal adhesion kinase, in
vitro and in vivo on inflammatory breast cancer cells. Breast
Cancer Res. 18:372016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Riedl JA, Brandt DT, Batlle E, Price LS,
Clevers H and Bos JL: Down-regulation of Rap1 activity is involved
in ephrinB1-induced cell contraction. Biochem J. 389:465–469. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li Z, Liu XB, Liu YH, Xue YX, Wang P, Liu
LB, Yao YL and Ma J: Functions for the cAMP/Epac/Rap1 signaling
pathway in low-dose endothelial monocyte-activating
polypeptide-II-induced opening of blood-tumor barrier. J Mol
Neurosci. 57:1–10. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Leblanc R and Peyruchaud O: The role of
platelets and megakaryocytes in bone metastasis. J Bone Oncol.
5:109–111. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chen XS, Li LY, Guan YD, Yang JM and Cheng
Y: Anticancer strategies based on the metabolic profile of tumor
cells: Therapeutic targeting of the Warburg effect. Acta Pharmacol
Sin. 37:1013–1019. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cohen-Solal KA, Boregowda RK and Lasfar A:
RUNX2 and the PI3K/AKT axis reciprocal activation as a driving
force for tumor progression. Mol Cancer. 14:1372015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu JF, Tsao YT and Hou CH: Amphiregulin
enhances intercellular adhesion molecule-1 expression and promotes
tumor metastasis in human osteosarcoma. Oncotarget. 6:40880–40895.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yang Y, Gao Z, Ma Y, Teng H, Liu Z, Wei H,
Lu Y, Cheng X, Hou L and Zou X: Fucoidan inhibits lymphangiogenesis
by downregulating the expression of VEGFR3 and PROX1 in human
lymphatic endothelial cells. Oncotarget. 7:38025–38035. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ohguchi S, Nakatsukasa H, Higashi T,
Ashida K, Nouso K, Ishizaki M, Hino N, Kobayashi Y, Uematsu S and
Tsuji T: Expression of alpha-fetoprotein and albumin genes in human
hepatocellular carcinomas: Limitations in the application of the
genes for targeting human hepatocellular carcinoma in gene therapy.
Hepatology. 27:599–607. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Nojiri S and Joh T: Albumin suppresses
human hepatocellular carcinoma proliferation and the cell cycle.
Int J Mol Sci. 15:5163–5174. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gai JQ, Sheng X, Qin JM, Sun K, Zhao W and
Ni L: The effect and mechanism of bufalin on regulating
hepatocellular carcinoma cell invasion and metastasis via
Wnt/β-catenin signaling pathway. Int J Oncol. 48:338–348. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chou HC, Sheu JC, Huang GT, Wang JT and
Chen DS: Albumin messenger RNA is not specific for circulating
hepatoma cells. Gastroenterology. 107:630–631. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yang J, Li S, Wang B, Wu Y, Chen Z, Lv M,
Lin Y and Yang J: Potential biomarkers for anti-EGFR therapy in
metastatic colorectal cancer. Tumour Biol. 37:11645–11655. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zheng LS, Yang JP, Cao Y, Peng LX, Sun R,
Xie P, Wang MY, Meng DF, Luo DH, Zou X, et al: SPINK6 promotes
metastasis of nasopharyngeal carcinoma via binding and activation
of epithelial growth factor receptor. Cancer Res. 77:579–589. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Day KC, Hiles GL, Kozminsky M, Dawsey SJ,
Paul A, Broses LJ, Shah R, Kunja LP, Hall C, Palanisamy N, et al:
HER2 and EGFR overexpression support metastatic progression of
prostate cancer to bone. Cancer Res. 77:74–85. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhang M, Gong W, Zuo B, Chu B, Tang Z,
Zhang Y, Yang Y, Zhou D, Weng M, Qin Y, et al: The microRNA miR-33a
suppresses IL-6-induced tumor progression by binding Twist in
gallbladder cancer. Oncotarget. 7:78640–78652. 2016.PubMed/NCBI
|
|
51
|
Bertucci F, Finetti P, Guille A, Adélaïde
J, Garnier S, Carbuccia N, Monneur A, Charafe-Jauffret E, Goncalves
A, Viens P, et al: Comparative genomic analysis of primary tumors
and metastases in breast cancer. Oncotarget. 7:27208–27219. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Schade B, Lesurf R, Sanguin-Gendreau V,
Bui T, Deblois G, O'Toole SA, Millar EK, Zardawi SJ, Lopez-Knowles
E, Sutherland RL, et al: β-Catenin signaling is a critical event in
ErbB2-mediated mammary tumor progression. Cancer Res. 73:4474–4487.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhang MH, Shen QH, Qin ZM, Wang QL and
Chen X: Systematic tracking of disrupted modules identifies
significant genes and pathways in hepatocellular carcinoma. Oncol
Lett. 12:3285–3295. 2016.PubMed/NCBI
|
|
54
|
Zhao Y, Xue F, Sun J, Guo S, Zhang H, Qiu
B, Geng J, Gu J, Zhou X, Wang W, et al: Genome-wide methylation
profiling of the different stages of hepatitis B virus-related
hepatocellular carcinoma development in plasma cell-free DNA
reveals potential biomarkers for early detection and high-risk
monitoring of hepatocellular carcinoma. Clin Epigenetics. 6:302014.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Myers JS, von Lersner AK and Sang QX:
Proteomic upregulation of fatty acid synthase and fatty acid
binding protein 5 and identification of cancer- and race-specific
pathway associations in human prostate cancer tissues. J Cancer.
7:1452–1464. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Akkiprik M, Peker I, Özmen T, Amuran GG,
Güllüoğlu BM, Kaya H and Özer A: Identification of differentially
expressed IGFBP5-related genes in breast cancer tumor tissues using
cDNA microarray experiments. Genes (Basel). 6:1201–1214. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wang DD, Liu ZW, Han MM, Zhu ZM, Tu YL,
Dou CQ, Jin X, Cai SW and Du N: Microarray based analysis of gene
expression patterns in pancreatic neuroendocrine tumors. Eur Rev
Med Pharmacol Sci. 19:3367–3374. 2015.PubMed/NCBI
|