Introduction
Neurodegenerative diseases, including Alzheimer's
disease, amyotrophic lateral sclerosis, Parkinson's disease and
Huntington's disease, are predicted to emerge as the second leading
cause of mortality worldwide by 2040. Therefore, understanding the
molecular mechanisms underlying neurodegeneration, and developing
effective therapeutic strategies is critical (1). Neurodegenerative diseases are also
considered to be protein misfolding disorders, and they share
similar characteristics, such as aggregation of misfolded proteins,
which may lead to neuronal dysfunction and apoptosis. Previous
research has indicated that the processing of misfolded aggregated
proteins in the endoplasmic reticulum (ER) induces ER stress and
causes mitochondrial dysfunction, and these processes are critical
in the progression of neurodegenerative diseases (2).
Diabetic retinopathy is largely considered to be a
microvascular disease, however, structural neurodegenerative
alterations, such as neuronal apoptosis, have been described in the
early stages of diabetic retinopathy (3,4).
Therefore, diabetic retinopathy is recognized as a
neurodegenerative disorder, as well as a microvascular disease. In
patients with diabetes, a non-enzymatic reaction between sugars and
the amino groups of proteins, lipids and nucleic acids leads to the
reversible formation of Schiff bases. Following rearrangement, they
form Amadori products, and these early glycation products then
undergo irreversible cross-linking, forming heterogeneous
fluorescent derivatives that are termed advanced glycation
end-products (AGEs) (5). AGEs are
a type of misfolded protein that have the ability to induce ER
stress, subsequently leading to mitochondrial dysfunction and
oxidative stress (6).
Oxidative stress has been reported to be a principal
mechanism implicated in the pathogenesis of various diseases,
including neurodegenerative disorders, diabetes and cardiovascular
disease (7). In addition,
oxidative stress has been implicated in a number of pathologies
associated with diabetes, including diabetic neuropathy and
nephropathy (8). Therefore,
antioxidative agents that prevent or delay oxidative stress-induced
neuronal apoptosis may have potential in the development of
effective therapeutic strategies to prevent the neurodegenerative
processes associated with diabetes.
Thioredoxin (Trx) was initially discovered in 1964
and belongs to the Trx family of redox proteins. It is a 12 kDa
multifunctional protein with a redox-active disulfide/dithiol
within its conserved active site sequence (−Cys-Gly-Pro-Cys-)
(9). Trx functions in several
cellular responses, including proliferation, apoptosis and
survival. In the extracellular milieu, Trx exhibits chemokine-like
activity, and functions as a scavenger of reactive oxygen species
and transcription factor activator in the cytoplasm (10). Trx has also been reported to serve
a cytoprotective role under conditions of cellular stress and
injury (11).
Proanthocyanidins are phenolic compounds that are
presented in fruit, vegetables, wine, tea, nuts and seeds (12). Grape seed proanthocyanidin extract
(GSPE) consists of a mixture of pycnogenols and flavonoids,
including oligomeric proanthocyanidins, which exhibit potent
antioxidative properties and are extracted from grape seeds and
skins (13). A previous study
reported that GSPE suppressed the apoptosis of bladder cells in
diabetic rats by activating the nuclear factor erythroid 2-like 2
(Nrf2) pathway (13). Furthermore,
a previous study by our group demonstrated that activation of the
Nrf2 pathway led to upregulation of Trx in mice with photoreceptor
degeneration (14). Therefore, in
the present study, GSPE was used to upregulate the expression of
Trx, and to investigate the antiapoptotic effects of Trx in
hyperglycemia-induced neurodegeneration. The present study aimed to
provide evidence to support the potential use of GSPE for the
prevention and treatment of diabetic retinopathy in clinical
practice.
Materials and methods
Cell culture and reagents
Mouse Neuro2a neuroblastoma cells obtained from the
Institute of Biochemistry and Cell Biology, Chinese Academy of
Sciences (Shanghai, China) were cultured in Dulbecco's modified
Eagle's medium (DMEM) supplemented with 10% fetal bovine serum
(both Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) at
37°C in a 5% CO2 atmosphere. Then, 3×105
cells/ml were seeded into a 6-well plate and medium was replaced
every 1–2 days. The cells were washed with PBS prior to the
experiments. The Trx inhibitor PX12 (Tocris Bioscience; Bio-Techne,
Minneapolis, MN, USA) was dissolved in 1 mM dimethyl sulfoxide and
stored at −20°C. The stock high-glucose (HG; 100 mM) medium (Gibco;
Thermo Fisher Scientific, Inc.) was diluted with DMEM to give a
final working concentration of 30 mM glucose and was stored at 4°C.
GSPE was purchased from Tianjin Jianfeng Natural Product R&D,
Co., Ltd. (Tianjin, China). Neuro2a cells were pretreated for 6 h
with or without 10 µM PX12, and subsequently received treatment for
24 h with or without 10 µg/ml GSPE and 30 mM glucose at 37°C in a
5% CO2 atmosphere.
Flow cytometric analysis
Neuro2a cells that were treated with HG, HG + GSPE
or HG + GSPE + PX12, and control cells, were washed with PBS three
times prior to analysis. Then, 3×105 cells/ml were
collected by centrifugation at 112 × g for 5 min at room
temperature, and then stained with annexin V (1:100)/propidium
iodide (PI) (1:500) in binding buffer (BioTools, Inc., Jupiter, FL,
USA) at room temperature for 15 min in the dark. Subsequently, the
samples were analyzed by BD Accuri™ C6 software version 1.0.264.21
(BD Biosciences, Franklin Lakes, NJ, USA).
Western blot analysis
Total proteins were extracted from Neuro2a cells
that were treated with HG, HG + GSPE or HG + GSPE + PX12, and
control cells. The cells were lysed in ice-cold lysis buffer (Geno
Technology, Inc., St., Louis, MO, USA) for 30 min. The lysates were
centrifuged at 12,000 × g for 30 min at 4°C, and the supernatants
were collected as protein samples. The concentration of protein was
measured by BCA assay and equal amounts of extracted protein
samples (50 µg protein/lane) were separated by 10% SDS-PAGE and
transferred onto polyvinylidene difluoride membranes (EMD
Millipore, Billerica, MA, USA). The membranes were blocked in 5%
non-fat milk for 1 h at room temperature and then incubated with
the following primary antibodies overnight at 4°C: Anti-Trx
(rabbit; 2429; 1:1,000 Cell Signaling Technology, Inc., Danvers,
MA, USA), anti-78 kDa glucose-regulated protein (rabbit; GRP78;
11587-1-AP; 1:1,000; ProteinTech Group, Inc., Chicago, IL, USA),
anti-Trx-interacting protein (Txnip; rabbit; 18243-1-AP; 1:1,000;
ProteinTech Group, Inc.), anti-apoptosis signal-regulating kinase
(ASK) 1 (rabbit; ab45178; 1:2,000; Abcam, Cambridge, UK), anti-Nrf2
(rabbit; sc-722; 1:500; Santa Cruz Biotechnology, Inc., Dallas, TX,
USA) and anti-β-actin (mouse; sc-4778; 1:1,000; Santa Cruz
Biotechnology, Inc.). All antibodies were diluted in 5% non-fat
milk. The membranes were washed three times with 1X TBS containing
0.1% Tween-20 (TBST) for 15 min and then incubated with secondary
antibodies horseradish peroxidase (HRP) conjugated goat anti-rabbit
immunoglobulin (Ig)G (ab6721; 1:2,000; Abcam) and HRP goat
anti-mouse IgG (ab205719; 1:2,000; Abcam) for 1 h at room
temperature. Following three washes with 1X TBST for 20 min, the
protein bands were visualized using enhanced chemiluminescence (GE
RPN2232; GE Healthcare Life Sciences) and exposed to X-ray film.
Blots were semi-quantified by densitometry using Labworks software
version 4.5 (Perkin Elmer, Inc., Waltham, MA, USA).
Terminal deoxynucleotidyl transferase
2′-deoxyuridine, 5′-triphosphate nick-end labeling (TUNEL)
assay
To detect cell apoptosis, 3×104 cells/ml
was seeding into 8-well chamber slides (Thermo Fisher Scientific,
Inc.) and a TUNEL assay was performed using a FITC labeled TUNEL
Cell Death Detection kit (Nanjing KeyGen Biotech Co., Ltd.,
Nanjing, China), according to the manufacturer's protocol. Briefly,
following control, HG, HG + GSPE or HG + GSPE + PX12 treatment,
cells were fixed in 4% paraformaldehyde for 15 min at room
temperature, washed in PBS three times for 10 min and incubated
with 50 µl reaction mixture for 60 min at 37°C in a dark humidified
chamber. The cells were then counterstained with PI (Vector
Laboratories, Inc., Burlingame, CA, USA) diluted 1:500 for 10 min
at room temperature in the dark. Apoptotic cells were observed in
at least 5 fields under a fluorescence microscope (BZ 7000; Keyence
Corporation, Osaka, Japan) and Image Pro Plus version 5.0 (Media
Cybernetics, Inc., Rockville, MD, USA) was used to analyze
data.
Animal experiments
A total 24 male inbred BALB/c mice (age, 6 weeks;
weight, 20–25 g) were supplied by Dalian Medical University
(Liaoning, China) and were housed in an animal control facility for
2 weeks in a 12-h light/dark cycle with a mean illumination of 80
lx and 30–70% humidity. The animals were maintained at 22±2°C. Tap
water and food pellets (Beijing Solarbio Science & Technology
Co., Ltd., Beijing, China) were available ad libitum. All
experimental procedures were conducted in accordance with the
institutional guidelines for the care and use of laboratory
animals, and experimental protocols were approved by the
Institutional Animal Care and Use Committee at Dalian Medical
University. A total of 6 mice were assigned to 4 groups; control,
diabetic, diabetic + GSPEI and diabetic + GSPEII. The diabetic mice
were fed a high fat diet (10% lard, 20% yolk, 1% cholesterol, 0.5%
cholate, 20% sucrose and 48.5% standard diet) for 8 weeks and
intraperitoneally injected with streptozotocin (STZ; 60 mg/kg;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) twice every 3 days
to induce diabetes. When blood glucose levels reached 250 mg/dl,
the model was considered to be successfully established. Subsequent
experiments were conducted between 10.00 am and 2.00 pm. STZ was
dissolved in cold 50 mM citric acid buffer (pH 4.5). The diabetic
mice were also administered with various concentrations of GSPE (50
and 100 mg/kg) via oral gavage.
Morphological analysis by quantitative
histology
The animals were sacrificed with an overdose of
carbon dioxide 2 weeks after commencement of the different
treatments. The enucleated eyes were immersed in Bouin's solution
for 24 h and then fixed in 70% ethanol for 24 h at room
temperature. Following alcohol dehydration, the eyes were embedded
in paraffin and 5-µm sagittal sections containing the entire
retina, including the optic disc, were obtained. The retinal
sections were stained with 0.5% hematoxylin and 1% eosin (H&E)
at room temperature to observe under a light microscope. In each of
the superior and inferior hemispheres, the thickness of the outer
nuclear layer (ONL) was measured at 9 defined points. Each point
was centered on adjacent 220-µm lengths of the retina. The first
point of measurement was at a ~220 µm distance from the optic nerve
head, and subsequent measurement points were located in the
periphery. The average ONL thickness was determined based on
measurements from 18 points in each section.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was obtained from each retina sample using
TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.). RT was
performed with the PrimeScript RT Reagent kit (Perfect Real Time;
Takara Bio, Inc., Otsu, Japan). RT-qPCR was performed to measure
Trx mRNA expression using SYBR Premix DimerEraser (Takara Bio,
Inc.), with reverse-transcribed cDNA as the template. All PCR
reactions were conducted in a final volume of 20 µl. The
amplification was performed using an ABI Prism 7000 Sequence
Detection System (Applied Biosystems; Thermo Fisher Scientific,
Inc.) under the following conditions: 95°C for 30 sec followed by
40 cycles of 95°C for 3 sec, 72°C for 30 sec and 55°C for 30 sec.
GAPDH was used as an internal reference gene. The primers used were
as follows: Forward, 5′-GGAATGGTGAAGCAGATCGAG-3′ and reverse,
5′-ACGCTTAGACTAATTCATTAAT-3′ for Trx; forward,
5′-TGTGATGGGTGTGAACCACGAGAA-3′ and reverse,
5′-GAGCCCTTCCACAATGCCAAAGTT-3′ for GAPDH. The 2−ΔΔCq
method was used to quantify the results (15).
Statistical analysis
Data are presented as the mean + standard deviation.
Each experiment was performed three times. The statistical
significance of the differences between groups was assessed using
one-way analysis of variance. Statistical analysis was performed
using SPSS software version 17.0 (SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Antiapoptotic effects of GSPE during
in vivo hyperglycemia
In the present study, diabetes was induced in mice
in vivo using an STZ injection and a high-fat diet. The
morphology of the retina was observed following H&E staining
and the present results demonstrated that ONL (photoreceptor cell
layer) thickness was diminished in diabetic mice compared with in
healthy mice. However, following treatment with various
concentrations of GSPE, ONL degeneration was revealed to be
prevented (Fig. 1A and B). Reverse
transcription-quantitative polymerase chain reaction was used to
detect the expression of Trx in tissue samples isolated from mice
in the various groups. As presented in Fig. 1C, the mRNA expression of Trx was
significantly downregulated in diabetic mice compared with in
healthy mice. Conversely, following GSPE administration, Trx mRNA
expression levels were significantly increased compared with in
diabetic mice.
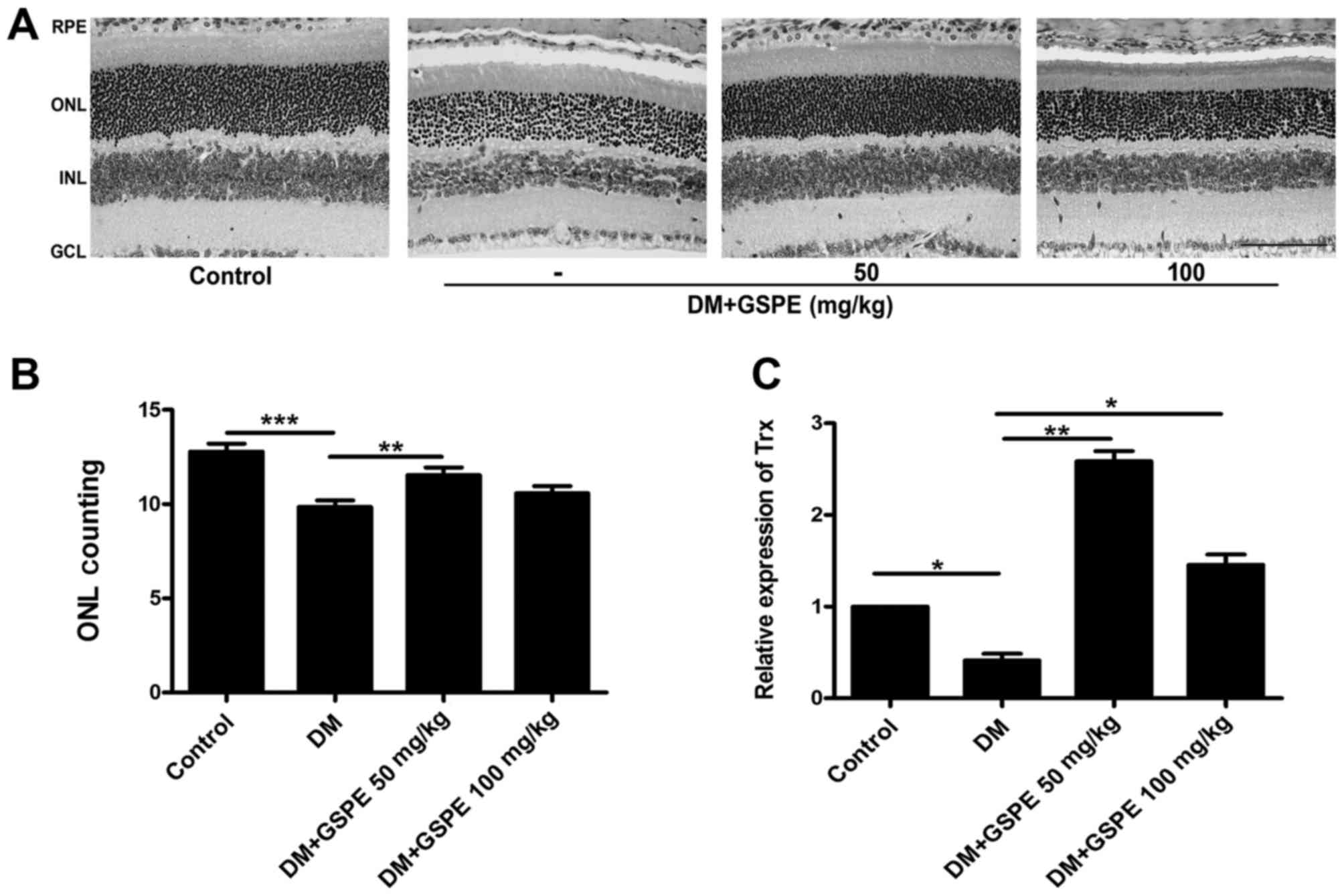 | Figure 1.GSPE prevents photoreceptor cell
damage induced by hyperglycemia. (A) The morphology of
photoreceptor cells was observed following staining with
hematoxylin and eosin. Scale bar, 50 µm. (B) The average nuclear
number of photoreceptor cells in 220-µm sections from the optic
nerve of the superior and inferior mouse retina. (C) Trx expression
following various treatments was assessed using reverse
transcription-quantitative polymerase chain reaction. Data are
expressed as the mean + standard deviation (n=3 mice/group).
*P<0.05, **P<0.01 and ***P<0.001, as indicated. RPE,
retinal pigment epithelium; INL, inner nuclear layer; GCL, ganglion
cell layer; GSPE, grape seed proanthocyanidin extract; Trx,
thioredoxin; ONL, outer nuclear layer; DM, diabetic mice. |
Antiapoptotic effects of GSPE during
in vitro hyperglycemia
During the in vitro experiments used in the
present study, mouse Neuro2a cells were maintained in hyperglycemic
conditions (30 mM glucose) to induce an HG damage model with or
without GSPE treatment. Hyperglycemia-induced apoptosis in Neuro2a
cells was quantitatively analyzed using flow cytometry. As
presented in Fig. 2A and B, HG
treatment increased the percentage of apoptotic cells compared with
the control group, whereas treatment with GSPE counteracted the
proapoptotic effects of hyperglycemia.
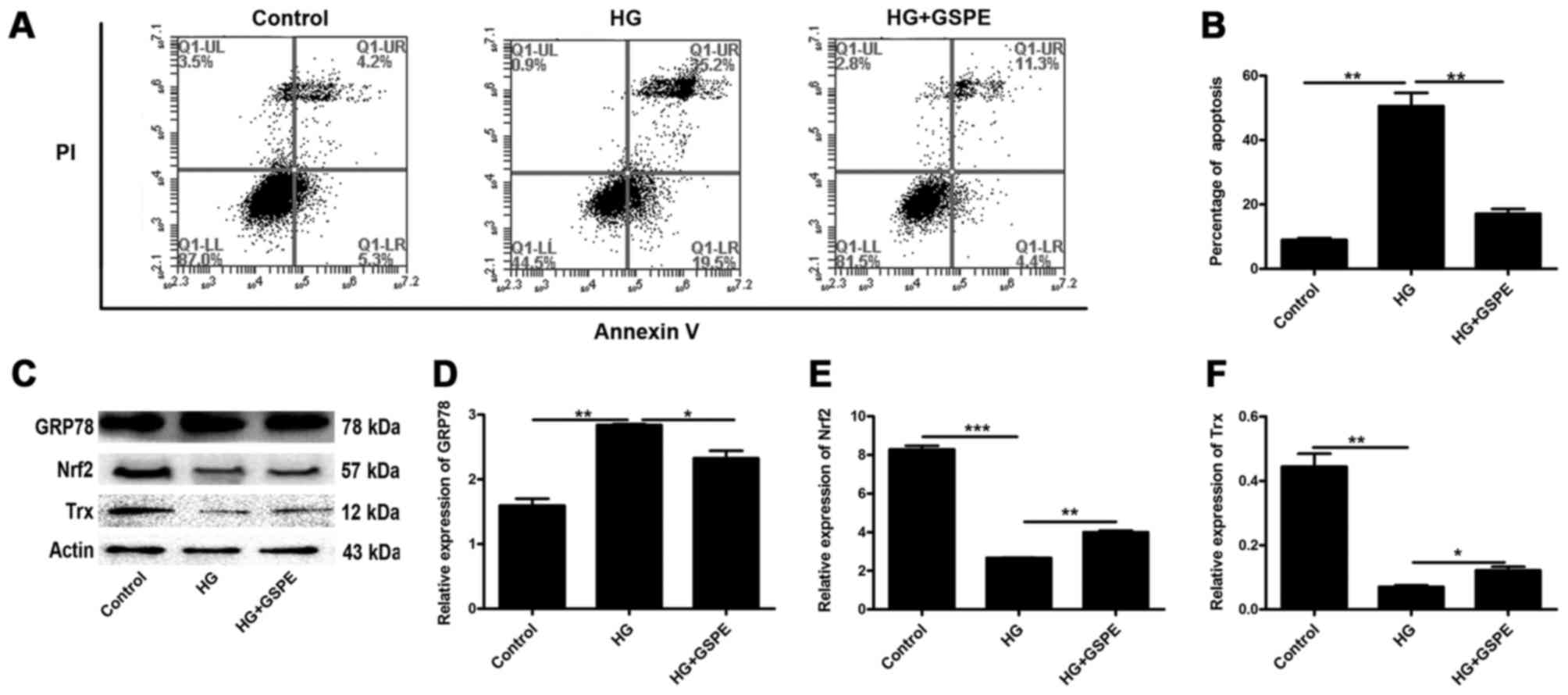 | Figure 2.Apoptosis of mouse Neuro2a cells
induced by hyperglycemia in vitro. (A) Apoptosis induced by
HG treatment was analyzed using flow cytometry. (B) Apoptosis
percentages obtained by flow cytometry were statistically analyzed.
(C) Western blot analysis was used to detect GRP78, Nrf2 and Trx
protein expression. Densitometric analysis was used to
semi-quantify the protein expression levels of (D) GRP78, (E) Nrf2
and (F) Trx, normalized to β-actin. Data are expressed as the mean
+ standard deviation (n=3 for each group). *P<0.05, **P<0.01
and ***P<0.001, as indicated. HG, high glucose; GRP78, 78 kDa
glucose-regulated protein; Nrf2, nuclear factor erythroid 2-like 2;
Trx, thioredoxin; PI, propidium iodide; GSPE, grape seed
proanthocyanidin extract. |
Western blot analysis was used to detect the
expression of GRP78, which is one of the markers of ER stress, Nrf2
and Trx in Neuro2a cells. The present results demonstrated that the
protein expression of GRP78 was significantly upregulated in
HG-exposed cells, whereas the protein expression of Nrf2 and Trx
was significantly downregulated (Fig.
2C-F). However, following GSPE treatment, GRP78 expression was
significantly decreased, whereas Nrf2 and Trx expression was
significantly increased (Fig.
2C-F). These results indicate that HG treatment may induce ER
stress in neuronal cells and downregulate the expression of Trx,
thus leading to cell apoptosis; Trx may therefore be implicated in
the antiapoptotic effects observed following GSPE treatment.
Roles of Trx during
hyperglycemia-induced Neuro2a cell apoptosis
Neuro2a cells were pretreated for 6 h with or
without the Trx system inhibitor PX12 (10 µM), following which they
were treated for 24 h with or without 10 µg/ml GSPE and 30 mM
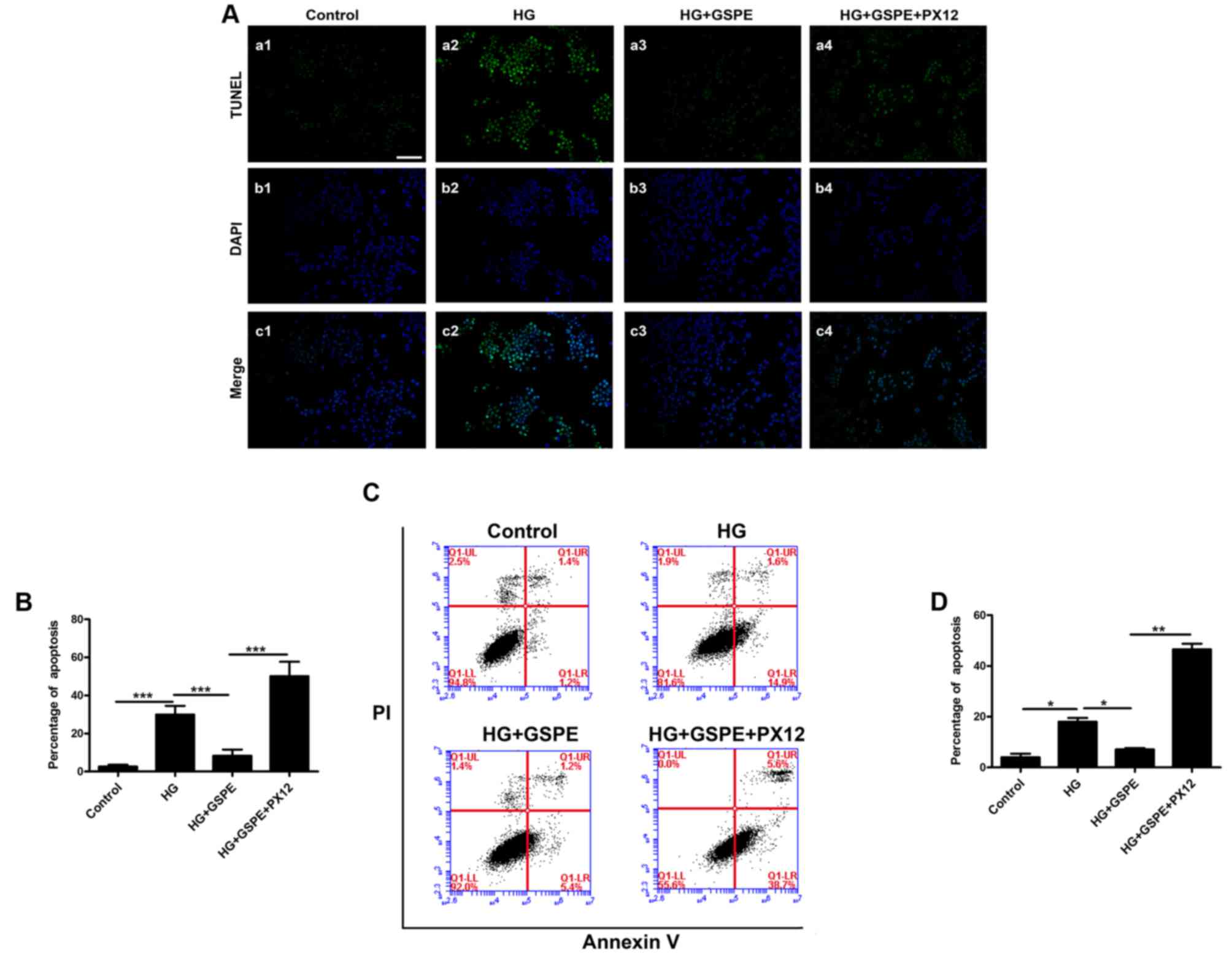
glucose. Neuro2a cell apoptosis was analyzed using TUNEL staining
(Fig. 3A and B) and flow cytometry
(Fig. 3C and D). The present
results demonstrated that GSPE inhibited Neuro2a apoptosis induced
by hyperglycemia. Conversely, PX12 treatment was revealed to
significantly enhance the HG-induced Neuro2a cell apoptosis
compared with GSPE-treated cells (Fig.
3) and inhibit the expression of Trx (Fig. 4). These results indicate that,
under hyperglycemic conditions, GSPE may exhibit neuroprotective
effects in Neuro2a cells may depend on a Trx-mediated
mechanism.
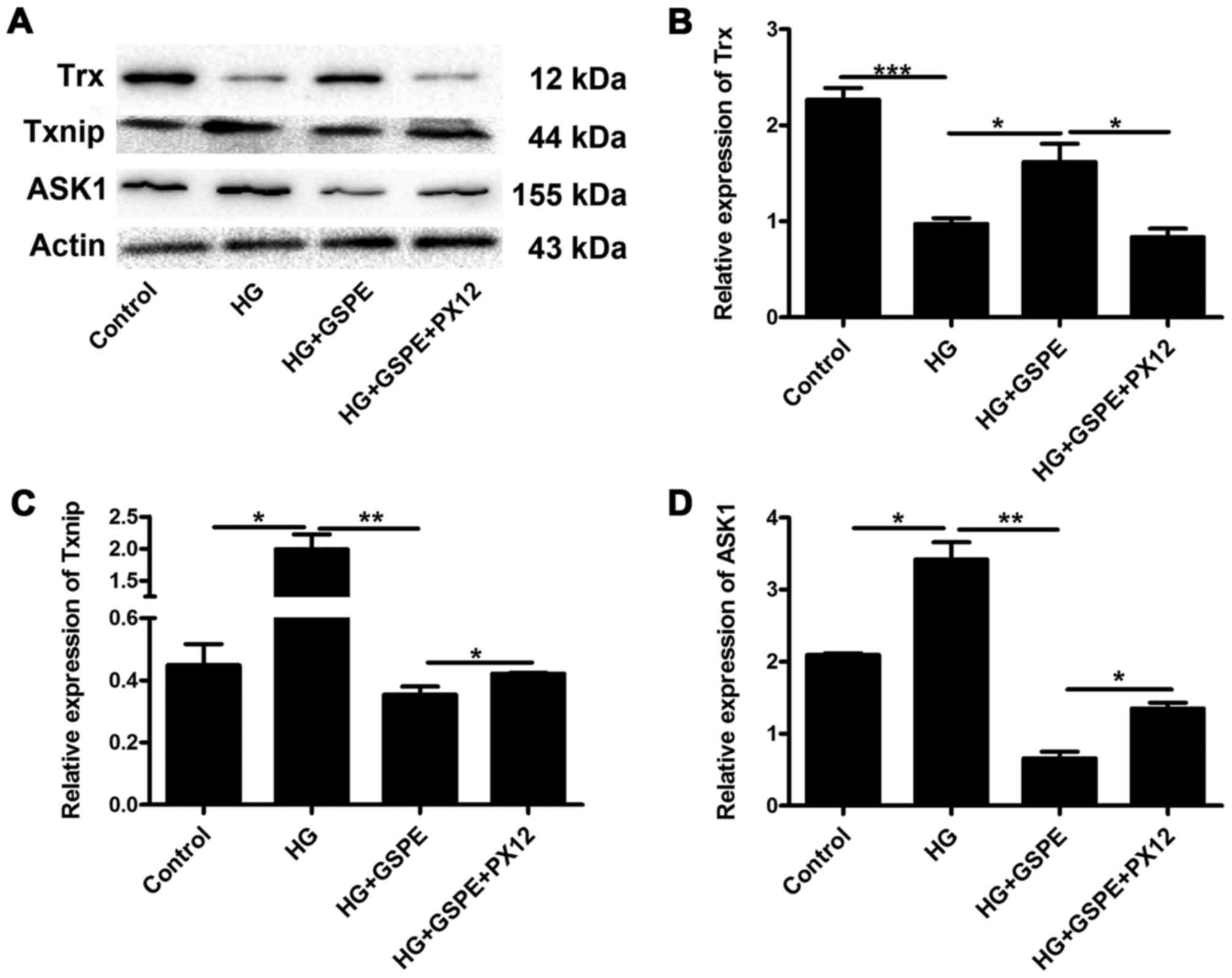 | Figure 4.Inhibition of Trx by PX12 alters the
expression of molecules implicated in the Trx cell signaling
pathway under hyperglycemic conditions. Western blot analysis was
used to detect the expression of Trx, Txnip and ASK1 in Neuro2a
cells. (A) Representative blot demonstrating expression of Trx,
Txnip and ASK1. β-actin was used as the loading control. The
protein expression levels of (B) Trx, (C) Txnip and (D) ASK1 were
assessed using densitometric analysis. Data are expressed as the
mean + standard deviation (n=3 for each group). *P<0.05,
**P<0.01 and ***P<0.001, as indicated. Trx, thioredoxin;
Txnip, Trx-interacting protein; ASK, apoptosis signal-regulating
kinase; HG, high glucose; GSPE, grape seed proanthocyanidin
extract. |
Effects of Trx expression inhibition
in HG-induced Neuro2a cell apoptosis
In order to investigate the molecular mechanisms
involved in Trx-mediated neuroprotection, Neuro2a cells were
treated with HG, GSPE and PX12, and the protein expression levels
of ASK1, Txnip and Trx were assessed using western blot analysis.
The present results demonstrated that the expression of ASK1 and
Txnip was upregulated in HG-treated cells, whereas the expression
of Trx was downregulated compared with control cells (Fig. 4). Compared with the HG group, GSPE
treatment significantly increased the protein expression levels of
Trx, and decreased the levels of ASK1 and Txnip (Fig. 4). However, in cells pretreated with
PX12, the expression of ASK1 and Txnip was increased, whereas the
expression of Trx was decreased compared with GSPE-treated cells
(Fig. 4). These results indicate
that Trx-associated pathways may be implicated in the antiapoptotic
effects of GSPE in hyperglycemic Neuro2a cells.
Discussion
Acute and chronic neurodegenerative diseases,
including stroke, traumatic brain injury, and Alzheimer's and
Parkinson's disease, are associated with high morbidity and
mortality, and few effective therapeutic strategies are available
for the treatment of these diseases (16). Previous studies have indicated that
diabetic retinopathy may represent a novel type of
neurodegenerative disease (4,17,18).
Diabetic retinopathy is the most common cause of
irreversible blindness in adults and leads to loss of central
vision. It has long been considered a microvasculopathy, however,
retinal diabetic neuropathy may also occur in patients with
diabetes mellitus (19). Several
events may lead to neuronal apoptosis in diabetic retinopathy,
including mitochondrial dysfunction and ER stress. Previous studies
have reported that ER stress is implicated in numerous diseases,
including diabetes, neurodegeneration and cancer (20–22).
In the present study, the protein expression of GRP78, which is an
ER stress marker, was revealed to be upregulated following HG
treatment in Neuro2a cells in vitro. These results indicate
that ER stress-associated mechanisms were activated during
hyperglycemia. In addition, Nrf2 protein expression was
downregulated, which led to a decrease in Trx protein expression,
subsequently leading to cell apoptosis. The present results
indicate that Trx may have potential as a novel therapeutic target
for the inhibition of HG-induced apoptosis.
GSPE is extracted from grape seeds and skins, and
has been demonstrated to exert protective effects against various
diseases, including diabetic nephropathy, drug-induced renal
toxicity, cancer metastasis and ischemic cardiomyopathy in animal
models (23). In addition, GSPE
has been reported to activate the Nrf2 pathway, and Nrf2 pathway
activation has been reported to be associated with the upregulation
of Trx expression (13,14), thus indicating that Trx may be
implicated in the antiapoptotic effects of GSPE. In the present
study, GSPE was used to treat diabetic mice in vivo, and
morphological analysis revealed that it was able to prevent ONL
degeneration compared with untreated diabetic mice. In addition,
the expression of Trx was significantly upregulated in mouse tissue
samples following GSPE treatment compared with in untreated
diabetic mice.
Based on the aforementioned findings, the Trx
inhibitor PX12 was used to pretreat Neuro2a cells. Flow cytometry
and TUNEL staining demonstrated that the percentage of apoptotic
cells was significantly reduced following GSPE treatment under
hyperglycemic conditions. However, PX12 pretreatment inhibited the
protective effects associated with GSPE treatment in
vitro.
In the present study, the molecular mechanisms
underlying the effects of GSPE were also investigated in
vitro. Txnip is an endogenous Trx inhibitor that inhibits the
intracellular actions of Trx, and Txnip has been reported to be
upregulated in a hyperglycemic environment (24). ASK1 is a member of the
mitogen-activated protein kinase kinase kinase family that serves a
major role during stress-induced apoptosis (25) and is regulated by Trx. In the
present study, the protein expression levels of ASK1 and Txnip were
increased following HG exposure, whereas it was decreased following
GSPE treatment compared with untreated cells exposed to HG.
However, ASK1 and Txnip expression was upregulated in GSPE-treated
cells that were pretreated with PX12 compared with cells treated
with GSPE alone. These results indicate that hyperglycemia may
induce the apoptosis of Neuro2a cells by activating ER stress, thus
leading to the activation of the Trx/ASK1/Txnip pathway. Based on
these results, it may be hypothesized that Trx serves a key role in
the antiapoptotic actions of GSPE.
In conclusion, the results of the present study
demonstrated that hyperglycemia induced Neuro2a cell apoptosis via
activation of ER stress-associated mechanisms and the
downregulation of Trx expression. In addition, Trx may serve an
important role in GSPE-mediated antiapoptotic mechanisms, thus
implicating the Trx/ASK1/Txnip signaling pathway in HG-induced
neurodegeneration. It can provide some evidence for potential
treatment of patients with diabetic retinopathy.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 30850001, 31371218
and 31300812).
References
|
1
|
Garcia-Huerta P, Troncoso-Escudero P,
Jerez C, Hetz C and Vidal RL: The intersection between growth
factors, autophagy and ER stress: A new target to treat
neurodegenerative diseases? Brain Res. 1649:173–180. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shah SZ, Zhao D, Khan SH and Yang L:
Unfolded protein response pathways in neurodegenerative diseases. J
Mol Neurosci. 57:529–537. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
van Dijk HW, Verbraak FD, Kok PH,
Stehouwer M, Garvin MK, Sonka M, DeVries JH, Schlingemann RO and
Abramoff MD: Early neurodegeneration in the retina of type 2
diabetic patients. Invest Ophthalmol Vis Sci. 53:2715–2719. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Simó R and Hernández C: European
Consortium for the Early Treatment of Diabetic Retinopathy
(EUROCONDOR): Neurodegeneration in the diabetic eye: New insights
and therapeutic perspectives. Trends Endocrinol Metab. 25:23–33.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yamagishi S, Fukami K and Matsui T:
Evaluation of tissue accumulation levels of advanced glycation end
products by skin autofluorescence: A novel marker of vascular
complications in high-risk patients for cardiovascular disease. Int
J Cardiol. 185:263–268. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sánchez-Chávez G, Hernández-Ramírez E,
Osorio-Paz I, Hernández-Espinosa C and Salceda R: Potential role of
endoplasmic reticulum stress in pathogenesis of diabetic
retinopathy. Neurochem Res. 41:1098–1106. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zeng XS, Jia JJ and Ma LF: Gensenoside Rb1
protects rat PC12 cells from oxidative stress-induced endoplasmic
reticulum stress: The involvement of thioredoxin-1. Mol Cell
Biochem. 410:239–246. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Weinberg E, Maymon T and Weinreb M: AGEs
induce caspase-mediated apoptosis of rat BMSCs via TNFα production
and oxidative stress. J Mol Endocrinol. 52:67–76. 2014.PubMed/NCBI
|
|
9
|
Laurent TC, Moore EC and Reichard P:
Enzymatic synthesis of deoxyribonucleotides. Iv. Isolation and
characterization of thioredoxin, the hydrogen donor from
escherichia coli B. J Biol Chem. 239:3436–3444. 1964.PubMed/NCBI
|
|
10
|
Huh KH, Cho Y, Kim BS, Do JH, Park YJ, Joo
DJ, Kim MS and Kim YS: The role of thioredoxin 1 in the
mycophenolic acid-induced apoptosis of insulin-producing cells.
Cell Death Dis. 4:e7212013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zeng XS, Jia JJ, Kwon Y, Wang SD and Bai
J: The role of thioredoxin-1 in suppression of endoplasmic
reticulum stress in Parkinson disease. Free Radic Biol Med.
67:10–18. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li SG, Ding YS, Niu Q, Xu SZ, Pang LJ, Ma
RL, Jing MX, Feng GL, Liu JM and Guo SX: Grape seed
proanthocyanidin extract alleviates arsenic-induced oxidative
reproductive toxicity in male mice. Biomed Environ Sci. 28:272–280.
2015.PubMed/NCBI
|
|
13
|
Chen S, Zhu Y, Liu Z, Gao Z, Li B, Zhang
D, Zhang Z, Jiang X, Liu Z, Meng L, et al: Grape seed
proanthocyanidin extract ameliorates diabetic bladder dysfunction
via the activation of the Nrf2 pathway. PLoS One. 10:e01264572015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kong L, Tanito M, Huang Z, Li F, Zhou X,
Zaharia A, Yodoi J, McGinnis JF and Cao W: Delay of photoreceptor
degeneration in tubby mouse by sulforaphane. J Neurochem.
101:1041–1052. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tarozzi A, Angeloni C, Malaguti M, Morroni
F, Hrelia S and Hrelia P: Sulforaphane as a potential protective
phytochemical against neurodegenerative diseases. Oxid Med Cell
Longev. 2013:4150782013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Simó R and Hernández C: Novel approaches
for treating diabetic retinopathy based on recent pathogenic
evidence. Prog Retin Eye Res. 48:160–180. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang X, Wang N, Barile GR, Bao S and
Gillies M: Diabetic retinopathy: Neuron protection as a therapeutic
target. Int J Biochem Cell Biol. 45:1525–1529. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sohn EH, van Dijk HW, Jiao C, Kok PH,
Jeong W, Demirkaya N, Garmager A, Wit F, Kucukevcilioglu M, van
Velthoven ME, et al: Retinal neurodegeneration may precede
microvascular changes characteristic of diabetic retinopathy in
diabetes mellitus. Proc Natl Acad Sci USA. 113:E2655–E2664. 2016;
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Oakes SA and Papa FR: The role of
endoplasmic reticulum stress in human pathology. Annu Rev Pathol.
10:173–194. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Placido AI, Pereira CM, Duarte AI,
Candeias E, Correia SC, Carvalho C, Cardoso S, Oliveira CR and
Moreira PI: Modulation of endoplasmic reticulum stress: An
opportunity to prevent neurodegeneration? CNS Neurol Disord Drug
Targets. 14:518–533. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu H, Cao MM, Wang Y, Li LC, Zhu LB, Xie
GY and Li YB: Endoplasmic reticulum stress is involved in the
connection between inflammation and autophagy in type 2 diabetes.
Gen Comp Endocrinol. 210:124–129. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sönmez MF and Tascioglu S: Protective
effects of grape seed extract on cadmium-induced testicular damage,
apoptosis, and endothelial nitric oxide synthases expression in
rats. Toxicol Ind Health. 32:1486–1494. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wei J, Shi Y, Hou Y, Ren Y, Du C, Zhang L,
Li Y and Duan H: Knockdown of thioredoxin-interacting protein
ameliorates high glucose-induced epithelial to mesenchymal
transition in renal tubular epithelial cells. Cell Signal.
25:2788–2796. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ishaq M, Kumar S, Varinli H, Han ZJ, Rider
AE, Evans MD, Murphy AB and Ostrikov K: Atmospheric gas
plasma-induced ROS production activates TNF-ASK1 pathway for the
induction of melanoma cancer cell apoptosis. Mol Biol Cell.
25:1523–1531. 2014. View Article : Google Scholar : PubMed/NCBI
|