Introduction
Atherosclerosis (AS) is the pathological basis of
cardiovascular and cerebrovascular disease, and is a common disease
endangering health, which can affect the elastic artery and elastic
muscle arterial intima, leading to luminal sclerosis and stenosis,
and subsequent coronary heart disease, cerebral infarction and
thromboembolic disease amongst other serious complications. Due to
changes in lifestyle and dietary habits, the incidence of coronary
heart disease and stroke have increased each year, and have become
major causes of mortality (1).
Mitogen-activated protein kinases (MAPKs) are
responsible for extracellular signal transduction into the cell to
induce cellular biological responses. MAPKs are important in the
regulation of the cardiovascular system and have become a major
target of pharmacological investigations (2).
Saikosaponin has several effects, as an
anti-endotoxin and corticosterone hormone, inhibiting the
activities of Na+ and K+-ATP enzymes, immune
regulation, anti-platelet activity, anti-allergy effects, and
anti-cell adhesion. However, few studies have examined its effects
on the mechanism of AS, Therefore, examining the role of
saikosaponin in treating AS is of important clinical
significance.
Materials and methods
Cell resuscitation, culture and MTT
assay detection of the inhibitory effect of saikosaponin on
oxidized low density lipoprotein (ox-LDL)-induced HUVEC injury
The HUVECs (Type Culture Collection of the Chinese
Academy of Sciences, Shanghai, China) were resuscitated and
cultured in low-glucose Dulbecco's modified Eagle's medium (DMEM;
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) containing
10% fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.), 100
U/ml penicillin, and 100 g/ml streptomycin, in an incubator at 37°C
and 5% CO2. The cells in logarithmic growth phase were
harvested and seeded into a 96-well plate at a density of
1×104 cells, with 100 µl/well. The cells were incubated
in a 5% CO2 incubator. Following cell adherence, the
medium was replaced with serum-free medium, and the cells were
cultured for another 24 h to the stationary phase of
synchronization, or the G0 phase. The cells were divided into five
groups: Model group; control group; low dose group (10 µM
saikosaponin); middle dose group (20 µM saikosaponin); and high
dose group (40 µM saikosaponin). The cells in the model group and
the control group were treated with an equal volume of serum-free
DMEM low-glucose culture medium. After 30 min, the cells in the
model group and the saikosaponin treatment groups were exposed to
ox-LDL at a density of 100 µg/ml. An equal volume of serum-free
DMEM low-glucose medium was added to the cells in control group,
and all cells were cultured for another 24 or 48 h. To each well,
20 µl MTT solution (5 mg/ml) was added for 4 h. Following this, the
supernatant was discarded and replaced with 150 µl DMSO. The medium
was agitated for 10 min, until the dark blue crystals were
sufficiently dissolved. The absorbance at a wavelength of 570 nm
was measured with a microplate reader and the cell viability was
calculated.
Flow cytometric analysis of the
inhibitory effect of saikosaponin on the apoptosis of
ox-LDL-induced HUVECs
Cells were initially treated in the same way as in
the MTT assay being divided into 5 groups. The cells were collected
and placed into a centrifuge tube at a density of 1×106
cells. Centrifugation was performed at 111.8 × g under 4°C for 5
min and the supernatant was discarded. A total of 200 µl Binding
Buffer was added to re-suspend the cells, and a mixture of 10 µl
Annexin V-FITC and 10 µl PI staining solution was added and gently
agitated. The cells were placed at room temperature in the dark for
15 min, followed by the addition of 300 µl Binding Buffer. The
apoptosis of cells was detected within 1 h using flow
cytometry.
ELISA to detect the effects of
saikosaponin on the levels of tumor necrosis factor (TNF)-α,
interleukin (IL)-6, superoxide dismutase (SOD) and malondialdehyde
(MDA) in HUVECs induced by ox-LDL
The cells were treated as described for the MTT
assay. The supernatant was collected and cultured. The expression
levels of TNF-α, IL-6, SOD and MDA in the HUVECs were detected
using a double antibody sandwich ELISA kit, in accordance with the
manufacturer's protocol of the ELISA kit.
Immunofluorescence staining to detect
the effects of saikosaponin on the nuclear transfer of nuclear
factor (NF)-κB in HUVECs induced by ox-LDL
Sterilized coverslips were placed in a 6-well
culture plate, onto which the well-grown HUVECs in the logarithmic
phase were inoculated at a cell density of 1×104 per
well, and the cells were treated according to the method used for
the MTT assay. The climbing cell tablets were fixed in 4%
paraformaldehyde solution for 30 min, followed by washing with PBS
three times, for 5 min each time. A total of 5 ml non-immune goat
serum (Gibco; Thermo Fisher Scientific, Inc.) was added in a
drop-wise manner. The cells were incubated for 20 min at room
temperature, following which excess solution was discarded and
NF-κB monoclonal antibody solution (1:200; cat. no. 554184; BD
Biosciences, Franklin Lakes, NJ, USA) was added for overnight
incubation at 4°C. The coverslips were removed and washed three
times with PBS, for 5 min each. Subsequently, goat anti-mouse
immunoglobulin (Ig) G labeled with rhodamine (1:100; cat. no.
SY0686; Biomics Laibo Technology Co., Ltd., Beijing, China) was
added and incubated for 1 h at room temperature. The cells were
then washed with PBS three times, for 5 min each, following which
nuclei were stained with DAPI. Finally, the cells were sealed with
50% glycerol, and positive photographic microscopy was performed
under a Nikon ECLIPSE 80i microscope (Nikon Corporation, Tokyo,
Japan).
Reverse transcription-polymerase chain
reaction (RT-PCR) assay to detect the effects of saikosaponin on
adhesion molecule gene expression in ox-LDL-induced HUVECs
A total of 1 ml TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) was added to the cells
for sufficient lysis, followed by the addition of 0.2 ml chloroform
and agitation, and standing in an ice-bath for 5 min. The cells
were then centrifuged for 20 min at 4°C and 1,609.92 × g. The
supernatant was replaced with an equal volume of isopropanol, and
placed in an ice-bath for 5 min. The cells were centrifuged again
for 20 min at 4°C and 1,609.92 × g, following which the supernatant
was discarded and 1 ml 75% ethanol was added. The cells were
centrifuged again for 20 min at 4°C and 1,609.92 × g. The
supernatant was carefully removed and the EP tube was inverted and
dried for 15 min at room temperature. RNase-Free ddH2O
(20 µl) was added to dissolve the precipitate following drying of
the sediment. Subsequently, 1 µl solution was suctioned with an
Eppendorf, and the remainder was stored in a deep freeze
refrigerator at −70°C. The 1 µl of solution was diluted into 80 µl
with diethyl pyrocarbonate ddH2O. The ratio of OD260 to
OD280 was measured using a spectrophotometer and the quantity of
RNA was calculated. The synthesis of cDNA and RT procedures were
performed according to manufacturer's protocol of the PrimeScript
RT reagent kit with the following: 12.5 µl SYBR Premix Ex Taq, 1 µl
PCR forward primer, 1 µl PCR reverse primer, 2 µl DNA template, 8.5
µl dH2O to a total of 25 µl. The fluorescence RT-PCR was
performed as follows: Amplification curve: Pre-denaturation for 5
min at 95°C, denaturation for 20 sec at 95°C, annealing at 60°C for
30 sec, extension at 72°C for 20 sec. This stage was for
fluorescence signal acquisition, comprising a total of 40 cycles;
dissolution curve: 60–95°C in increments of 0.5°C per cycle, for 20
sec, this stage was for fluorescence signal acquisition, comprising
a total of 71 cycles. The PCR product size was verified by 10 µl
electrophoresis on a 1% agarose gel. The primer sequences were as
follows: Intercellular adhesion molecule 1 (ICAM-1) forward,
5′-CAATACGTCGCACCGCCTT-3′ and reverse, 5′-ACTTGATACGGTGCTACTT-3′;
vascular cell adhesion molecule 1 (VCAM-1) forward,
5′-CGCAGAAGTACATTCGG-3′ and reverse, 5′-CCCACTGAGGCAAGTACACTG-3′;
and GAPDH forward, 5′-ACCTCCTCGTTACGACAT-3′ and reverse,
5′-GCACTTACTCACTGCCGTTG-3′.
Western blot analysis to detect the expression
levels of caspase-3, B-cell lymphoma 2 (Bcl-2), Bcl-2-associated X
protein (Bax), extracellular signal-regulated kinase (ERK)1/2,
phosphorylated (p-)ERK1/2, P-38, p-p38, c-Jun N-terminal
kinase (JNK) and p-JNK. The cells were collected and washed
twice in PBS, and 400 µl of lysate was added to each flask with 40
µl PMSF (10 mmol/l). The flasks were gently shaken and then placed
on ice 10 min for sufficient lysis. The cells were aspirated
repeatedly with a sterile syringe. The lysate was transferred to an
EP tube, and placed in an ice-bath for 30 min, followed by
centrifugation for 15 min at 12,000 × g at 4°C. The supernatant was
transferred to a new EP tube, and the protein density was
determined using the ELISA method. To each tube, 20 µl 6X buffer
was added for every 100 µl, and was boiled for 5 min. Following
mixing, the samples were stored at −80°C. The proteins (50 µg/lane)
from the above samples were separated by electrophoresis on a 12%
SDS-PAGE gel. The separated proteins were transferred onto a PVDF
membrane through a wet method, and maintained at room temperature
for 1 h. Subsequently, primary antibodies against caspase-3 (cat.
no. 9661; Cell Signaling Technology Inc., Danvers, MA, USA),
anti-Bcl-2 (cat. no. sc-7382; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) anti-Bax (cat. no. sc-6236; Santa Cruz
Biotechnology, Inc.), anti-ERK1/2 (cat. no. LT9996; LifeTein, LLC.,
Somerset, NJ, USA), anti-p-ERK1/2 (cat. no. LT5646; LifeTein,
LLC.), anti-P-38 (cat. no. sc-166182; Santa Cruz Biotechnology,
Inc.), anti-p-p38, anti-JNK (cat. no. RB020016; RayBiotech, Inc,
Norcross, GA, USA) and anti-p-JNK (cat. no. sc-6254; Santa Cruz
Biotechnology, Inc.) were added to the membranes (all at 1:1,000)
and the cells were incubated at 4°C overnight. The cells were then
washed with PBST three times and the secondary horse radish
peroxidase conjugated antibody goat anti-mouse IgG (cat. no.
A00160; 1:1,000; Kings Global Biotech, Ltd., Maharashtra, India)
was added. The cells were incubated for 1 h at 4°C, and then washed
with PBST three times. Subsequent color development and fixation
were performed following visualization with the enhanced
chemiluminescence luminous liquid kit (cat. no. WBKLS0050; EMD
Millipore, Billerica, MA, USA). The expression levels of the above
proteins were measured and quantified using Image J analysis
software (version 1.48U; National Institutes of Health, Bethesda,
MD, USA).
Statistical analysis
All data are expressed as the mean ± standard
deviation. Comparisons between the two groups were performed using
a unpaired Student's t-test, and multiple sets of data (>2) were
compared using one-way analysis of variance. P<0.05 was
considered to indicate a statistically significant difference. All
data were analyzed using GraphPad Prism 5.0 (GraphPad Software,
Inc., La Jolla, CA, USA).
Results
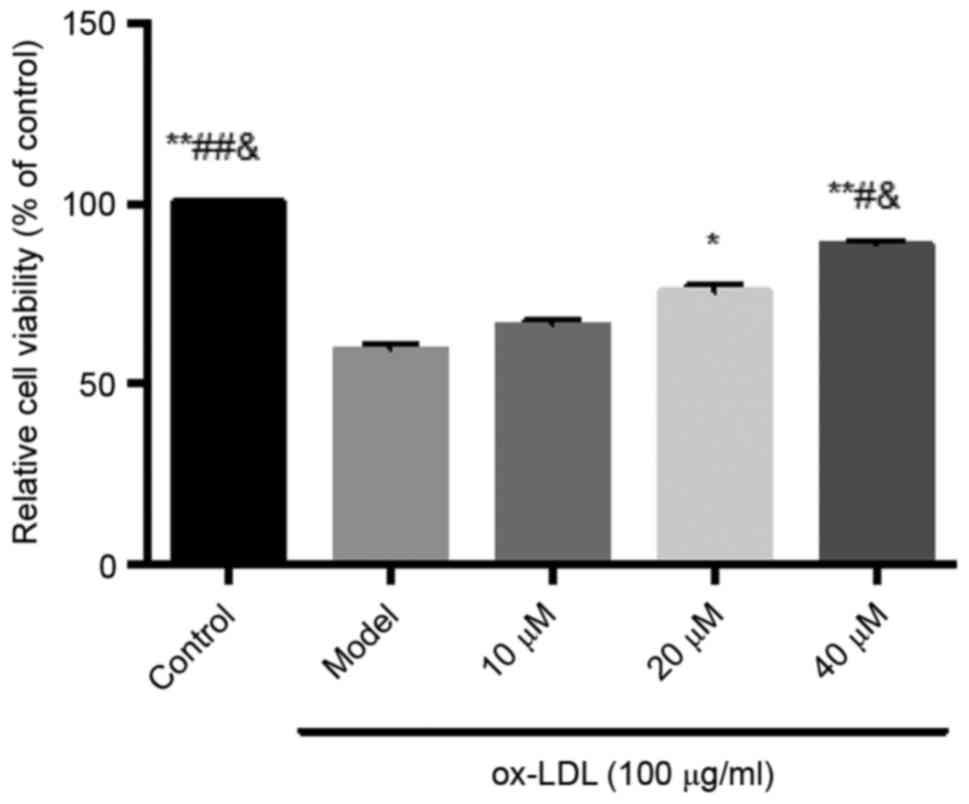
Inhibitory effects of saikosaponin on
ox-LDL-induced HUVEC injury
The results showed that ox-LDL at a concentration of
100 µg/ml caused significant cell damage, and decreased cell
viability. Saikosaponin at all concentrations (10, 20 and 40 µM)
inhibited the ox-LDL-induced HUVEC injury and improved cell
viability, particularly at a concentration of 40 µM, as shown in
Fig. 1.
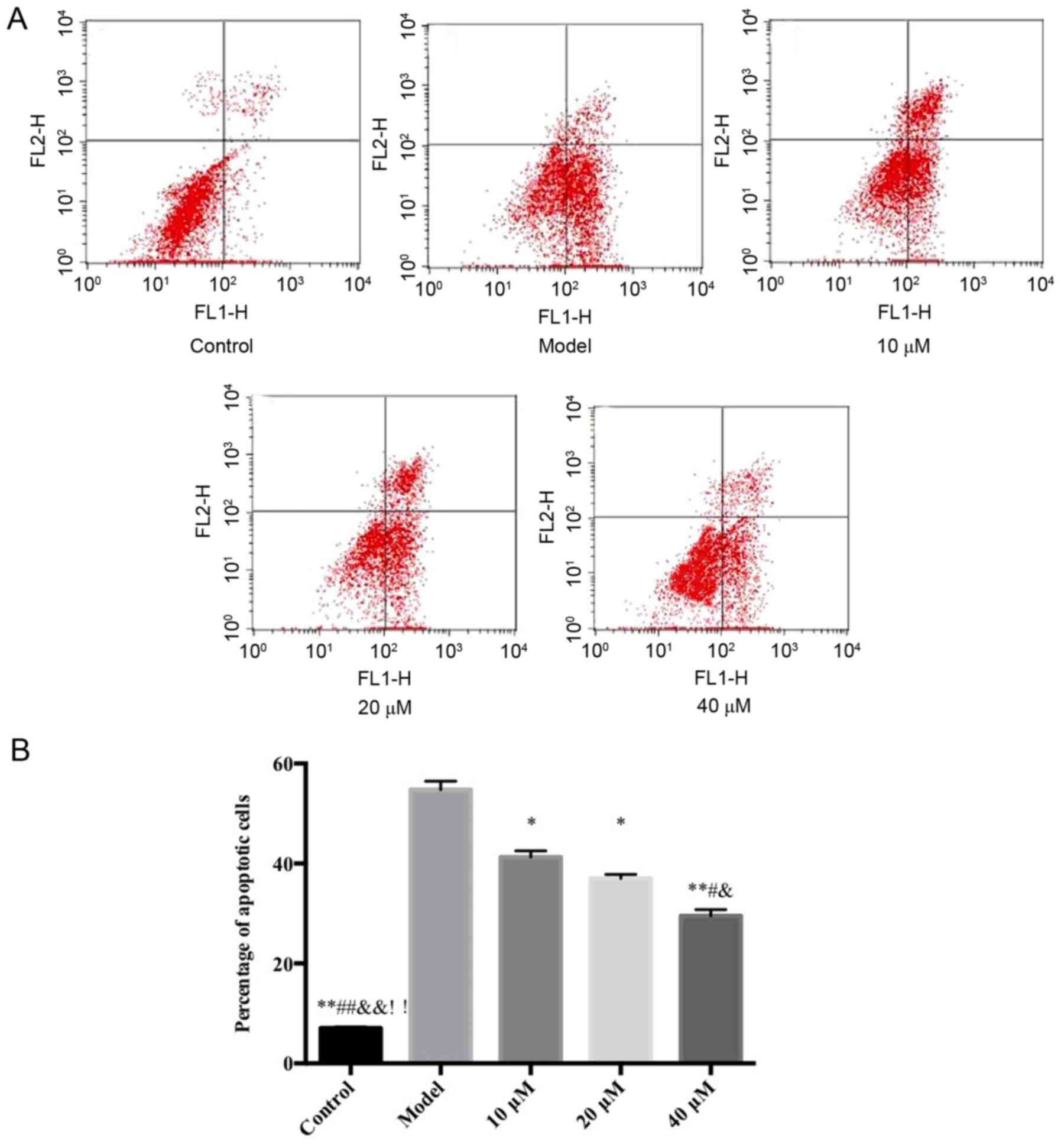
Flow cytometry detection of the
inhibitory effects of saikosaponin on ox-LDL-induced HUVEC
apoptosis
The results demonstrated that, compared with the
control group, the proportion of HUVECs in early stage apoptosis in
the model group increased significantly. However, compared with the
model group, the saikosaponin-treated groups showed significant
decreases in early stage apoptosis of HUVECs, particularly at in
the 40 µM group, as shown in Fig.
2.
Effects of saikosaponin on expression
levels of TNF-α, IL-6, SOD and MDA in ox-LDL-induced HUVECs
The results showed that, compared with the control
group, the levels of TNF-α, IL-6, SOD and MDA in the model group
were significantly increased, and the level of SOD was
significantly decreased. Compared with the model group, the levels
of TNF-α, IL-6 and MDA in the saikosaponin-treated groups were
decreased, and the level of SOD was increased, particularly in the
40 µM group with statistical significance (Table I).
 | Table I.Effects of saikosaponin on the levels
of TNF-α, IL-6, MDA and SOD in oxidized low-density
lipoprotein-induced human umbilical vein endothelial cells. |
Table I.
Effects of saikosaponin on the levels
of TNF-α, IL-6, MDA and SOD in oxidized low-density
lipoprotein-induced human umbilical vein endothelial cells.
| Group | TNF-α (pg/ml) | IL-6 (pg/ml) | MDA
(nmol/mgprot) | SOD
(nmol/mgprot) |
|---|
| Control |
82.11±10.21a,c,f |
63.89±7.53a,c,e |
15.02±1.45a,c,f |
170.22±8.73a,c,f |
| Model |
125.28±12.90 |
108.11±6.51 |
32.99±2.73 |
112.09±4.89 |
| 10 µM |
110.35±10.28b |
88.49±1.72 |
29.02±2.09 |
120.67±9.62 |
| 20 µM |
96.12±2.84b |
80.65±5.83b |
27.84±1.75 |
133.89±4.91 |
| 40 µM |
90.23±2.21a,d |
71.89±6.66a,c,f |
19.04±1.10a,c,f |
156.69±1.23a,c |
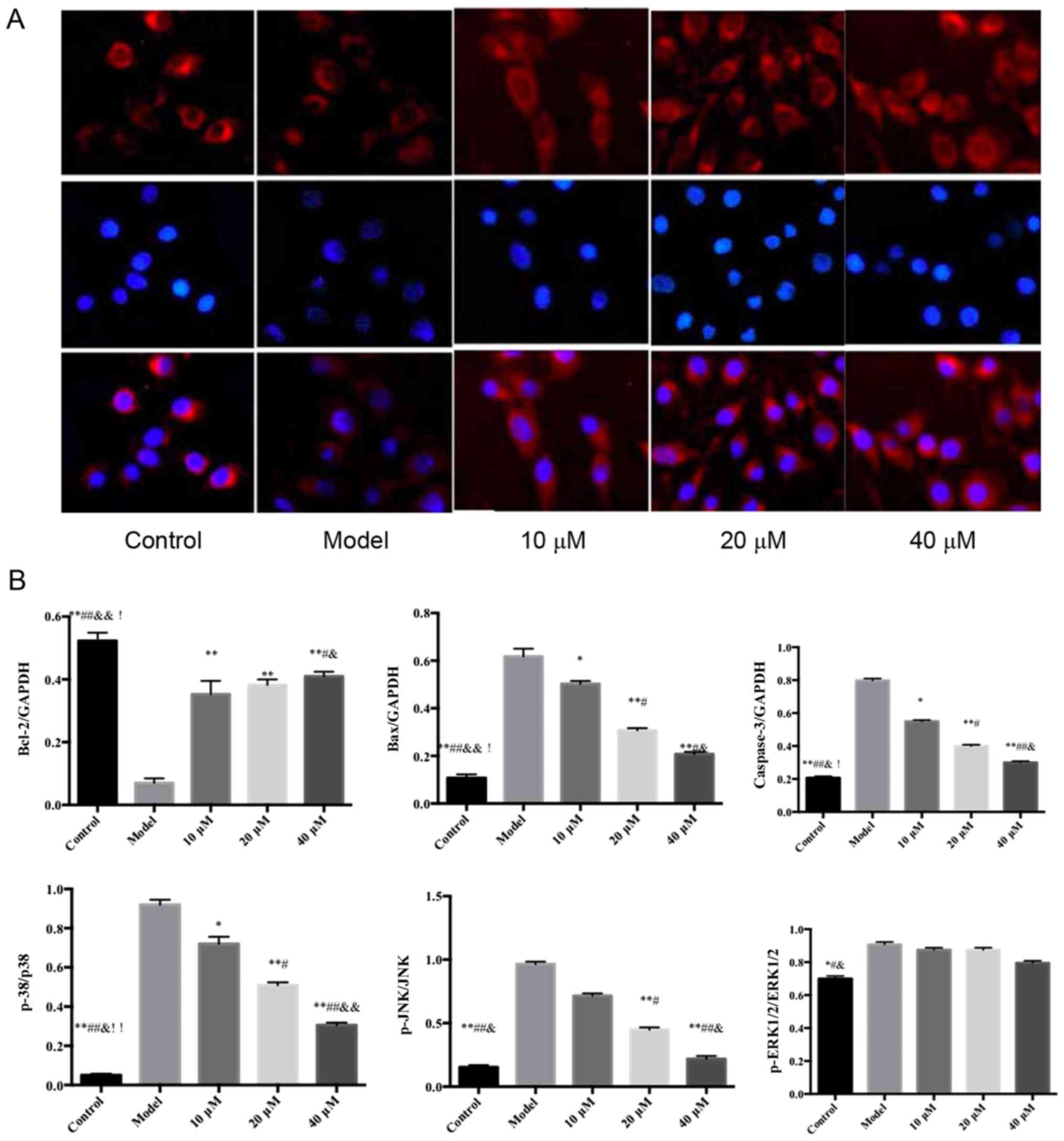
Effects of saikosaponin on the nuclear
transfer of NF-κB p65 in ox-LDL-induced HUVECs
The results of the immunofluorescence analysis
showed that NF-κB p65, visible in red, was expressed in the
cytoplasm and nucleus. Following DAPI staining, the
positively-stained nuclei were blue, which indicated the location
of the nucleus. Compared with the control group, the level of NF-κB
p65 was significantly increased in the nucleus of thr model group.
However, compared with the model group, the level of NF-κB p65 was
significantly decreased in the nucleus of the saikosaponin-treated
groups, particularly in the 40 µM group, as shown in Fig. 3.
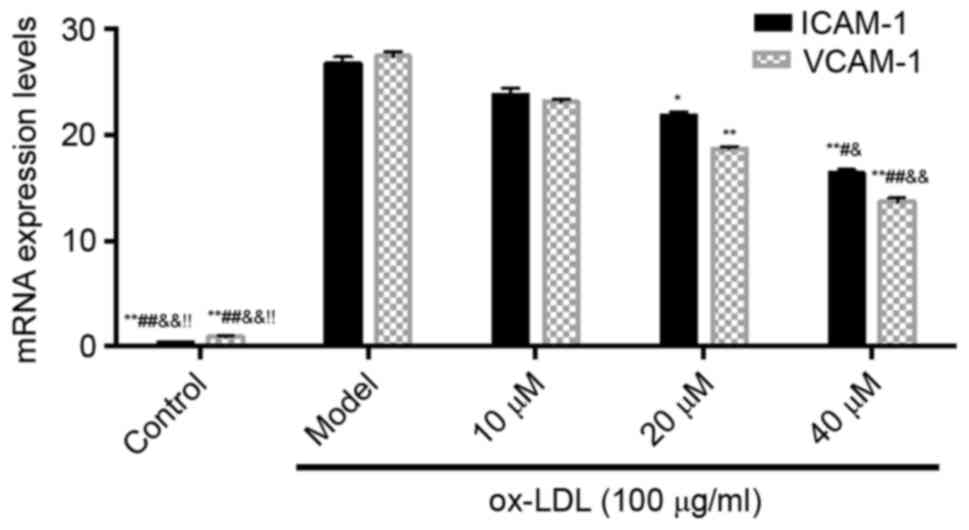
Effect of saikosaponin on the gene
expression of adhesion molecules in ox-LDL-induced HUVECs
The results showed that, compared with the control
group, the mRNA expression levels of ICAM-1 and VCAM-1 in the model
group were increased. However, compared with the model group, the
mRNA levels of ICAM-1 and VCAM-1 in the saikosaponin-treated groups
were significantly decreased, particularly in the 40 µM group, as
shown in Fig. 4.
Effects of saikosaponin on the protein
expression levels of caspase-3, Bcl-2, Bax, ERK1/2, p-ERK1/2, p38,
p-p38, JNK and p-JNK
The protein expression levels of ERK1/2, p38 and JNK
in the ox-LDL-induced HUVECs were similar among all groups;
however, the expression levels of p-p38 and p-JNK were
significantly different among groups (Fig. 5).
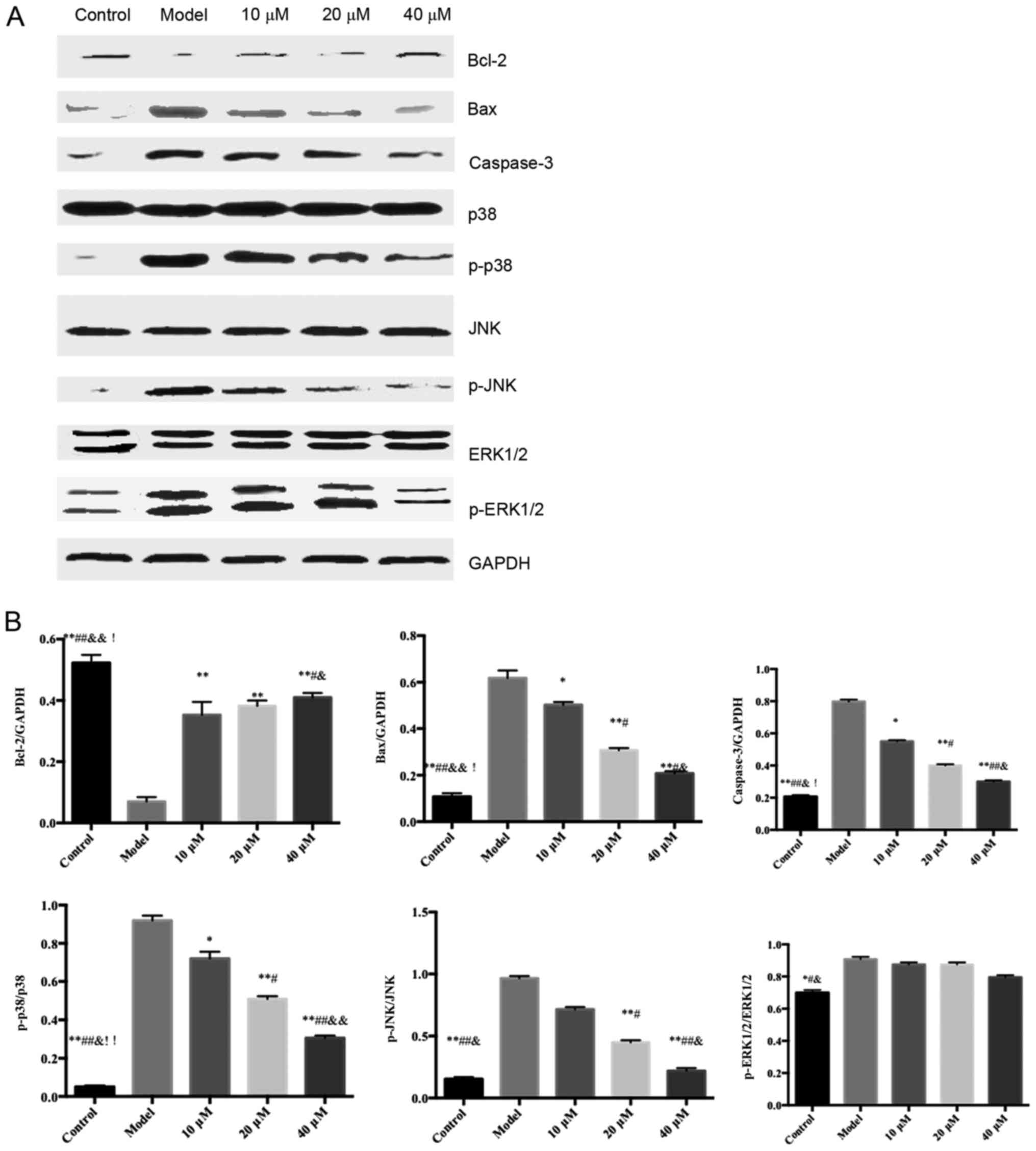 | Figure 5.Effects of saikosaponin on the protein
expression levels of caspase-3, Bcl-2, Bax, ERK1/2, p-ERK1/2, p38,
p-p38, JNK and p-JNK. (A) Blots demonstrate the expression levels
of proteins. (B) Quantitative results of the western blotting.
*P<0.05 vs. model, **P<0.01 vs. model; #P<0.05
vs. 10 µM; ##P<0.01 vs. 10 µM;
&P<0.05 vs. 20 µM; &&P<0.01
vs. 20 µM; !P<0.05 vs. 40 µM, !!P<0.01
vs. 40 µM. Bcl-2, B-cell lymphoma 2; Bax, Bcl-2-associated X
protein; ERK, extracellular signal-regulated kinase; p-,
phosphorylated. |
Discussion
The MAPK cascade is one of the most important signal
transduction pathways in cells, and is involved in multiple
biological responses. There are three MAPK signaling pathways in
mammalian cells, namely the ERK, JNK and p38 MAPK pathways
(2). The MAPK signaling pathway is
dormant when unstimulated, but is activated by the cascade
phosphorylation of upstream kinases. The activated MAPKs enter the
nucleus and facilitate the phosphorylation of transcription
factors, which alter the gene expression of the cell, and are
involved in the processes of cell proliferation, differentiation,
transformation and apoptosis (3,4).
In the p38 MAPK pathway, MAPK kinase (MKK)4 can
inhibit the MAPK signaling pathway, whereas MKK3/6 can
phosphorylate and activate MKK, thus activating MAPK and further
phosphorylating downstream molecules, including protein kinase,
phospholipase and transcription factors. MAPK specifically
regulates gene expression and is involved in the transduction
processes of cell proliferation, differentiation and apoptosis
(5–7).
In mammalian cells, the ERK pathway is considered to
be the classic MAPK signaling pathway involved in cell
proliferation and differentiation, mediated by growth cytokines,
whose signal transduction requires the activation of ERK1/2
(8,9).
JNK is a member of the MAPK superfamily. Being
expressed in the cytoplasm, JNK can activate downstream nuclear
transcription factors through transcriptional- and
non-transcriptional-dependent forms. The activation of JNK occurs
through the phosphorylation of amino terminal residues. Following
activation, JNK shifts to the nucleus (10–12).
JNK is important in cell stress response and is involved in
regulating apoptosis (13).
Normally, the phosphorylation levels of MAPK determine the activity
of JNK (14,15).
The present study found that saikosaponin inhibited
ox-LDL-induced HUVEC injury. The activity of SOD was increased,
whereas MDA content was decreased. The levels of JNK and p38
phosphorylation were decreased; therefore, the oxidative stress
injury in the HUVECs induced by ox-LDL was inhibited. In addition,
saikosaponin prevented NF-κB from being induced into the nucleus,
inhibited the secretion of inflammatory cytokines (TNF-α and IL-6)
and adhesion molecules (ICAM-1 and VCAM-1), increased the
expression of Bcl-2, and decreased the expression of caspase-3 and
Bax, thereby inhibiting the apoptosis of ox-LDL-induced HUVECs. The
different effects of saikosaponin on the phosphorylation of ERK1/2,
JNK and p38 in the HUVECs suggested that HUVECs may be involved in
the occurrence and development of AS through their unique
respective signaling pathways.
In conclusion, saikosaponin significantly inhibited
AS, which may have been through inhibiting the activation of p38
and ERK1/2 signaling pathways, thus having an anti-proliferative
effect on smooth muscle cells. Through inhibiting the JNK and p38
signaling pathways, inhibiting the p38 and NF-κB protein cascade,
saikosaponin inhibited injury and apoptosis of the endothelial
cells.
Acknowledgements
This study was supported by the Foundation of
Ministry of Science of Shaanxi Province, China (grant no.
2012K15-02-05).
References
|
1
|
Lee SM, Lee YJ, Kim YC, Kim JS, Kang DG
and Lee HS: Vascular protective role of vitexicarpin isolated from
Vitex rotundifolia in human umbilical vein endothelial cells.
Inflammation. 35:584–593. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Madonna R, Massaro M, Pandolfi A, Consoli
A and De Caterina R: The prominent role of p38 mitogen-activated
protein kinase in insulin-mediated enhancement of VCAM-1 expression
in endothelial cells. Int J Immunopathol Pharmacol. 20:539–555.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang Z, Niu Q, Peng X, Li M, Liu Y, Liu J,
Wen S and Wei Y: Mitofusin 2 ameliorates aortic remodeling by
suppressing ras/raf/ERK pathway and regulating mitochondrial
function in vascular smooth muscle cells. Int J Cardiol.
178:165–167. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Park B, Yim JH, Lee HK, Kim BO and Pyo S:
Ramalin inhibits VCAM-1 expression and adhesion of monocyte to
vascular smooth muscle cells through MAPK and PADI4-dependent NF-κB
and AP-1 pathways. Biosci Biotechnol Biochem. 79:539–552. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Martin-Blanco E: p38 MAPK signalling
cascades: Ancient roles and new functions. Bioessays. 22:637–645.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chahine MN, Blackwood DP, Dibrov E,
Richard MN and Pierce GN: Oxidized LDL affects smooth muscle cell
growth through MAPK-mediated actions on nuclear protein import. J
Mol Cell Cardiol. 46:431–441. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang GF, Shi CG, Sun MZ, Wang L, Wu SX,
Wang HF, Xu ZQ and Chen DM: Tetramethylpyrazine attenuates
atherosclerosis development and protects endothelial cells from
ox-LDL. Cardiovasc Drugs Ther. 27:199–210. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fisk M, Gajendragadkar PR, Mäki-Petäjä KM,
Wilkinson IB and Cheriyan J: Therapeutic potential of p38 MAP
kinase inhibition in the management of cardiovascular disease. Am J
Cardiovasc Drugs. 14:155–165. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sun Y, Liu WZ, Liu T, Feng X, Yang N and
Zhou HF: Signaling pathway of MAPK/ERK in cell proliferation,
differentiation, migration, senescence and apoptosis. J Recept
Signal Transduct Res. 35:600–604. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Glaros EN, Kim WS and Garner B:
Myriocin-mediated up-regulation of hepatocyte apoA-I synthesis is
associated with ERK inhibition. Clin Sci (Lond). 118:727–736. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dhanasekaran DN and Reddy EP: JNK
signaling in apoptosis. Oncogene. 27:6245–6251. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yarza R, Vela S, Solas M and Ramirez MJ:
c-Jun N-terminal Kinase (JNK) signaling as a therapeutic target for
Alzheimer's disease. Front Pharmacol. 6:3212016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wei L, Deng W, Cheng Z, Guo H, Wang S,
Zhang X, He Y and Tang Q: Effects of hesperetin on platelet-derived
growth factor-BB-induced pulmonary artery smooth muscle cell
proliferation. Mol Med Rep. 13:955–960. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cui Y, Sun YW, Lin HS, Su WM, Fang Y, Zhao
Y, Wei XQ, Qin YH, Kohama K and Gao Y: Platelet-derived growth
factor-BB induces matrix metalloproteinase-2 expression and rat
vascular smooth muscle cell migration via ROCK and ERK/p38 MAPK
pathways. Mol Cell Biochem. 393:255–263. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim MH, Kang HM, Kim CE, Han S and Kim SW:
Ramipril inhibits high glucose-stimulated up-regulation of adhesion
molecules via the ERK1/2 MAPK signaling pathway in human umbilical
vein endothelial cells. Cell Mol Biol Lett. 20:937–947. 2015.
View Article : Google Scholar : PubMed/NCBI
|