Introduction
Hand, foot, and mouth disease (HFMD) is
characterized by the development of mild febrile illness with
papulovesicular lesions on the hands, feet, and mouth. HFMD
outbreaks have been reported throughout the Asia-Pacific region and
pose a serious health threat to young children (1). Epidemics of HFMD in China recently
became an important public health concern and 1,898,760 cases of
HFMD have been reported in 2015 alone (2–5).
Enterovirus 71 (EV71) is the main causative agent of HFMD and is
responsible for the majority of deaths in children under 3 years of
age (6,7). The primary manifestations of deadly
EV17 infection is brain stem encephalitis, pulmonary edema, and
heart failure (5,8,9).
EV71 is a small non-enveloped virus composed of a
single positive-strand RNA enclosed in an icosahedral capsid
assembled with four structural proteins (VP1, VP2, VP3, and VP4)
(10). VP1, VP2, and VP3 are
exposed on the surface of the virion, while VP4 is hidden inside
(11). VP1 is a promising target
for the development of vaccines as it is reported to elicit
neutralizing antibody responses. Foo et al recently reported
that the highly conserved SP70 peptide derived from amino acids
(aa) 208–222 of VP1 (YPTFGEHKQEKDLEYC) can elicit neutralizing
antibodies in mice (12,13). This response was comparable to the
neutralizing antibody response elicited by whole virion, and
primarily involved IgG1 (14,15).
Nevertheless, the mechanisms by which SP70-directed
antibodies neutralize EV71 are not yet understood. In this study,
we synthesized a set of SP70 mutated peptides using alanine
scanning mutagenesis. These peptides were used to characterize
species-specific antibodies induced by EV71-based antigens.
Materials and methods
Cells and virus
Vero cells and myeloma SP2/0 and YB2/0 cells
[American Type Culture Collection (ATCC), Manassas, VA, USA] were
maintained in RPMI 1640 medium supplemented with 10% fetal bovine
serum (FBS) at 37°C in a humidified 5% CO2
atmosphere.
The EV71 strain H3-TY (Genbank no. GU129025.1) was
provided by Zhejiang Pukang Biotechnology Co., Ltd. (Hangzhou,
China). EV71 H3-TY is a subtype C4 virus and was isolated from a
child with HFMD in Hangzhou (Zhejiang, China). Viral stocks were
prepared in Vero cells. Virus titers were determined using the
REED-Muench method.
Animals
BALB/c mice (8-week-old, female, n=9), Lou/C rats
(3-week-old, female, n=3), and New Zealand white rabbits (2.5–3 kg,
female, n=6) were obtained from the Laboratory Animal Center of
Zhejiang Academy of Medical Science (Hangzhou, China). All
procedures and animal experiments were approved by the Animal Care
and Use Committee of Zhejiang Academy of Medical Science.
Peptide synthesis
The SP70 epitope of EV71 VP1 (aa 208–222) was
synthesized, along with 15 similar peptides in which one alanine
was substituted for another aa at each position (Table I). The coxsackievirus A16 VP1
epitope (aa 208–222) was synthesized as a negative control
(Table I). A keyhole limpet
hemocyanin (KLH)-conjugated EV71 SP70 peptide was synthesized. All
peptides were synthesized with 95% HPLC purity grade by GenScript
(Piscataway, NJ, USA) and confirmed by mass spectrometry. The
peptides were dissolved in 100% dimethyl sulfoxide (DMSO) at 2
mg/ml and stored at −80°C.
 | Table I.Sequences of synthesized
peptides. |
Table I.
Sequences of synthesized
peptides.
| No. | Peptide
sequence |
|---|
| WT |
YPTFGEHKQEKDLEY |
| 1 |
APTFGEHKQEKDLEY |
| 2 |
YATFGEHKQEKDLEY |
| 3 |
YPAFGEHKQEKDLEY |
| 4 |
YPTAGEHKQEKDLEY |
| 5 |
YPTFAEHKQEKDLEY |
| 6 |
YPTFGAHKQEKDLEY |
| 7 |
YPTFGEAKQEKDLEY |
| 8 |
YPTFGEHAQEKDLEY |
| 9 |
YPTFGEHKAEKDLEY |
| 10 |
YPTFGEHKQAKDLEY |
| 11 |
YPTFGEHKQEADLEY |
| 12 |
YPTFGEHKQEKALEY |
| 13 |
YPTFGEHKQEKDAEY |
| 14 |
YPTFGEHKQEKDLAY |
| 15 |
YPTFGEHKQEKDLEA |
| NC |
YPTFGEHLQANDLDY |
Production of purified inactivated
EV71 from Vero cells
Vero cells were transduced with the EV71 H3-TY
virus, centrifuged at 10,000 × g for 15 min, and inactivated by
incubation with 0.025% formalin at 37°C for 48 h. After
inactivation, the virus was concentrated 100 folds using Pellicon
ultrafiltration (EMD Millipore, Billerica, MA, USA) and purified on
a sepharose 6-FF column (GE Healthcare, Waukesha, WI, USA). Virus
concentration was determined by sandwich ELISA (AbMax Biotechnology
Co., Ltd., Beijing, China).
Production of recombinant EV71 VP1
(rEV71-VP1) protein
A recombinant pET32a plasmid (Novagen; Merck KGaA,
Darmstadt, Germany) containing a N-terminal His-tagged EV71 VP1
gene (Genbank no. GU129025.1) was constructed using NdeI and
XhoI restriction endonucleases. The plasmid pET32a-EV71 VP1
was transformed into BL21 (DE3) chemically competent cells.
Transformed cells were incubated in a shaker at 37°C until their
optical density (OD)600 reached 0.6. They were then
induced with 0.5 mM IPTG (Amresco, Solon, OH, USA) at 37°C for 6 h.
The cells were harvested by centrifugation and the pellet was
re-suspended in 50 mM Tris-HCl buffer (pH 9.0). After ultrasound
treatment, inclusion protein was separated by centrifugation at
14,000 × g for 30 min and dissolved in 20 mM glycine and 8 M Urea
(pH 8.5). The recombinant protein was purified by HiTrap HP 5 ml
(GE Healthcare) and dialyzed by D-tube Mega with a cut-off at 6–8
kDa (Merck Serono GmbH, Darmstadt, Germany) using 20 mM glycine and
0.1% Tween-20 (pH 9.0). The concentration of the purified proteins
was determined using a BCA kit (Pierce Chemical, Dallas, TX, USA).
The proteins were identified by 10% SDS-PAGE.
Generation of monoclonal antibodies
(Mabs) elicited by SP70
Three BALB/c mice and three Lou/C rats were
immunized by injection with a suspension containing KLH-conjugated
EV71 SP70 peptide (20 µg for mice and 50 µg for rats) emulsified
with Freund's complete adjuvant (100 µl/animal), as described by
Lim et al (16). After four
weeks, the animals were sacrificed. The cells harvested from the
mouse spleens were fused with myeloma SP2/0 cells (ATCC), as
described by Yokoyama (17) and
Yokoyama et al (18). The
cells harvested from the rat spleens were fused with myeloma YB2/0
cells (ATCC) according to the same methods (16–18).
The fused hybridoma cells were re-suspended and incubated in DMEM
with 20% FBS for 10 days. Supernatants were screened by ELISA for
SP70 peptide binding. The hybridoma clone with the strongest
reactivity to the EV71 SP70 peptide was re-cloned twice by limiting
dilution and its reactivity was re-confirmed using a home-made
ELISA. Supernatants generated by sub-cloned hybridoma cells were
typed for Ig type and affinity using a home-made indirect ELISA.
Antibody neutralization was assessed by a micro-neutralization
assay.
Generation of serum polyclonal
antibodies (Pabs) elicited by inactivated EV71 or rEV71-VP1
protein
Three BALB/c mice received leg muscle injection of
Freund's complete adjuvant containing 480 EU of inactivated EV71 or
50 µg of rEV71-VP1 protein (100 µl/mouse), and boosted with the
same immunogen after 7 days. Two weeks after the final boost, serum
was collected and purified using a HiTrap rProtein A FF column (GE
Healthcare). The animals vaccinated with Freund's complete adjuvant
were used as negative controls. Antibody neutralization activity
was assessed by a micro-neutralization assay.
Three rabbits received leg muscle injection of
Freund's complete adjuvant containing 960 EU of inactivated EV71 or
500 µg of rEV71-VP1 protein (500 µl/rabbit), and were boosted
thrice with the same amount of immunogen at 2-week intervals. Ten
days after the final boost, serum was collected and purified by
HiTrap rProtein A FF column (GE Healthcare). The animals vaccinated
with Freund's complete adjuvant were used as negative controls.
Micro-neutralization assay
Antibody samples were serially diluted in minimal
essential medium (MEM) in a 96-well plate. An equal volume of virus
at 1×102 TCID50 (50% tissue culture infective
dose)/ml was added. The mixture was cultured for 2 h at 37°C. Then,
Vero cells (1×104/well) in a 96-well plate were
inoculated with 100 µl of this mixture and incubated at 37°C in a
5% CO2 incubator. After 1 h, 100 µl of MEM supplemented
with 10% bovine serum was added to each well. The plate was
incubated at 35°C in 5% CO2 for 5–7 days. Cytopathic
effects (CPEs) were observed every day using an IX51 inverted
microscope (Olympus, Tokyo, Japan). Antibody neutralization titers
were calculated using the Reed-Muench method (18).
Immunofluorescence assay
Vero cells infected with EV71 H3-TY were incubated
in a 96-well plate at 35°C in 5% CO2. When CPEs were
observed (at least 25%), cells were fixed with precooled acetone.
Antibodies diluted to 1:100 in PBS were added to the plates blocked
with 1% BSA in PBS, and incubated overnight at 4°C. After washing
five times with PBS containing 0.01% Tween-20 (PBS-T),
FITC-conjugated goat anti-mouse IgG, goat anti-rat IgG, or goat
anti-rabbit IgG (diluted 1:50 in PBS-T; Santa Cruz Biotechnology,
Santa Cruz, CA, USA) were added and the plates were incubated for 1
h at 37°C. After washing five times with PBS-T, fluorescent images
were visualized and captured using a DM BL2 fluorescence microscope
(Leica Microsystems GmbH, Wetzlar, Germany).
Antibody affinity for SP70 alanine
scanning mutagenesis peptides determined by peptide-ELISA
assay
All peptides were diluted in bicarbonate buffer to a
final concentration of 2 µg/ml and coated on polystyrene flat
bottom 96-well plates (Corning Inc., Corning, NY, USA) at 4°C
overnight (100 µl/well). Peptide representing the coxsackievirus
A16 VP1 epitope (aa 208–222) was added to each well as a negative
control. Every peptide was respectively coated on different wells
for duplication. The coated wells were washed thrice with PBS-T,
blocked with 1% BSA in PBS (200 µl/well), and incubated 1 h at
37°C. After washing thrice with PBS-T, antibodies diluted in PBS-T
were added to the coated wells (100 µl/well in duplicate) and
incubated for 1 h at 37°C. The plates were washed thrice with
PBS-T, and antibody binding was detected by incubation with
HRP-conjugated goat anti-mouse IgG, goat anti-rat IgG, or goat
anti-rabbit IgG (Santa Cruz Biotechnology) diluted 1:5,000 in PBS-T
(100 µl/well) for 1 h at 37°C. Plates were washed four times in
PBS-T and bound antibodies were detected using 100 µl/well of
freshly prepared O-phenylenediamine (0.4 mg/ml) containing
H2O2 (0.4 mg/ml) in 0.1 M citrate buffer (pH
5.2) (Sigma-Aldrich, St. Louis, MO, USA). After 15 min, the
peroxidase reaction was stopped using 2.0 M
H2SO4 (50 µl/well). The OD was measured at
490 nm on a R680 microplate reader (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). Results were displayed as S/N value, i.e., the
ratio of ODsample to ODnegative control 1.
Negative control 1 was the negative control of the peptide-ELISA
map (PEM) detection; it was used to eliminate the background of the
polypeptide itself. If the average absorbance value of the negative
control 1 was lower than 0.05, the value was fixed at 0.05.
Human blood sample assay
The study using clinical samples was approved by the
Ethics Committee of the Zhejiang Academy of Medical Science, and
conducted according to the principles of the Declaration of
Helsinki. The blood samples were provided by the Ningbo Disease
Prevention and Control Center (Ningbo, China). The written informed
consents were obtained from the patients.
A competitive ELISA was used to measure the affinity
to SP70 peptide The SP70 primary peptides were diluted to 20 µg/ml
with carbonate buffer (pH 9.6), added into 96-well plate (100
µl/well), and incubation overnight at 4°C. The plate was washed
four times and blocked with 1% BSA. The plate was again washed four
times. Positive blood samples were diluted 500 times with PBS-T
containing 1% BSA, and then divided into 18 aliquots. SP70 mutant
peptides (including CA16) was added to each aliquot to reach a
final concentration of 400 µg/ml, followed by incubation overnight
at 4°C. A DMSO control group (DMSO as peptide solvent) was set up.
The next day, the samples were added to a pre-coated 96-well plate
(100 µl/well, in triplicates), and then incubated at 37°C for 1 h.
The plate was washed four times and incubated with goat anti-human
IgGHRP antibody (1:10,000; Santa Cruz Biotechnology) at 37°C for 30
min. The plate was washed four times and added with TMB for
developing. The reaction was stopped with 2 N
H2SO4. The binding activity of the SP70
mutant peptide to the SP70 antibody was evaluated by the S/N ratio.
S was obtained from each peptide-treated serum and N was from the
DMSO treated serum. The binding activity of SP70 mutant peptide to
SP70 antibody was inversely proportional to the ratio (S/N).
Statistical analysis
Data were analyzed using SPSS 19.0 (IBM, Armonk, NY,
USA). Data were expressed as mean ± standard deviation (SD) and
compared using ANOVA with the least significant difference (LSD)
post hoc test. Differences were considered to be statistically
significant when P-values were <0.05.
Results
Purification of inactivated EV71 and
rEV71-VP1 protein
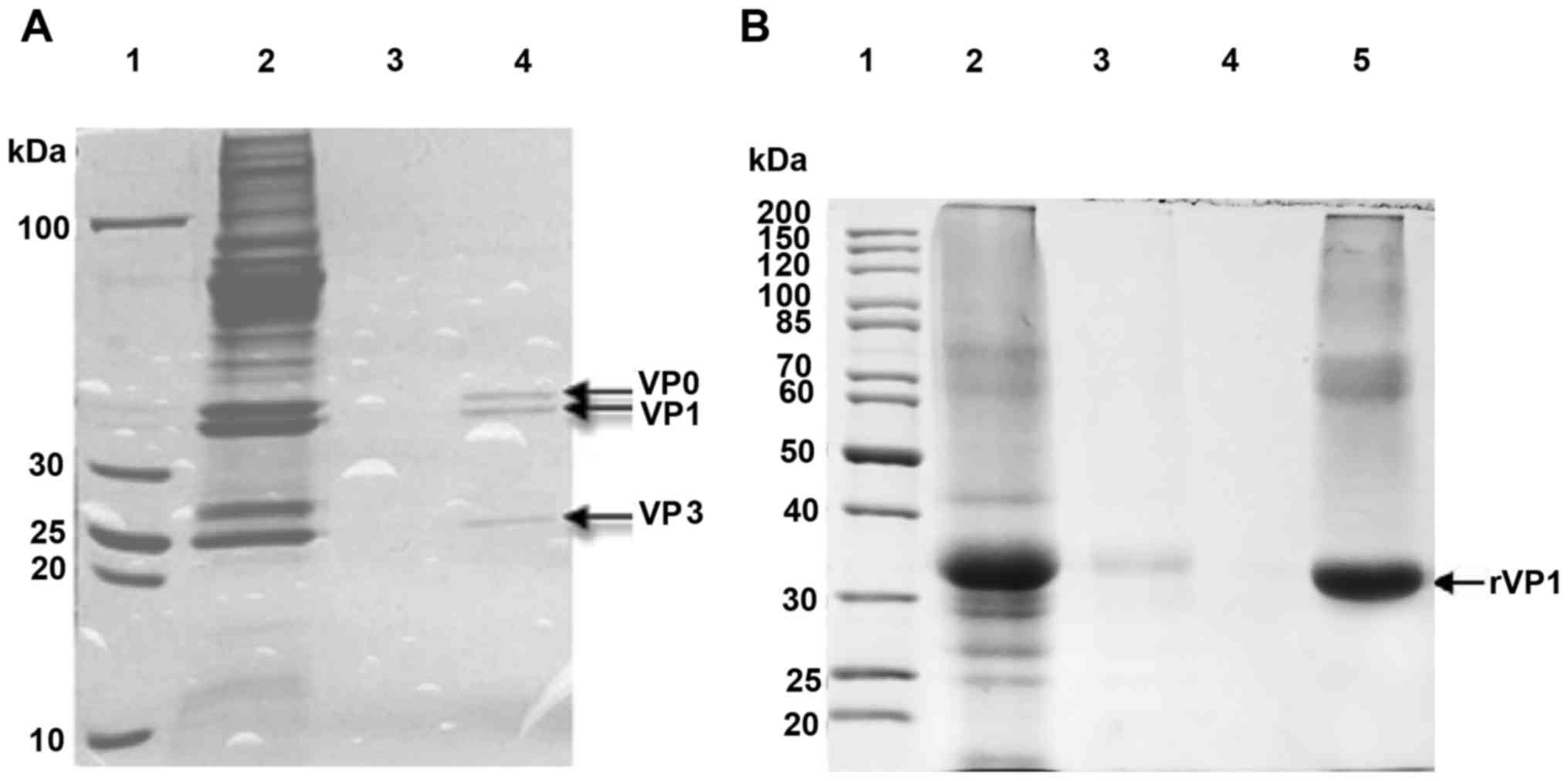
The purification of inactivated EV17 and VP1
recombinant protein immunogens was assessed by SDS-PAGE with
Coomassie Brilliant Blue staining. The SDS-PAGE of purified
(sepharose 6-FF column) inactivated virus revealed three bands,
corresponding to VP0, VP1, and VP3 (Fig. 1A, lane 4). Recombinant VP1 was
purified by HiTrap HP column, and SDS-PAGE revealed a single band
(34 kDa) (Fig. 1B, lane 5), close
to its theoretical molecular weight (33.9 kDa).
Specificity of antibodies binding to
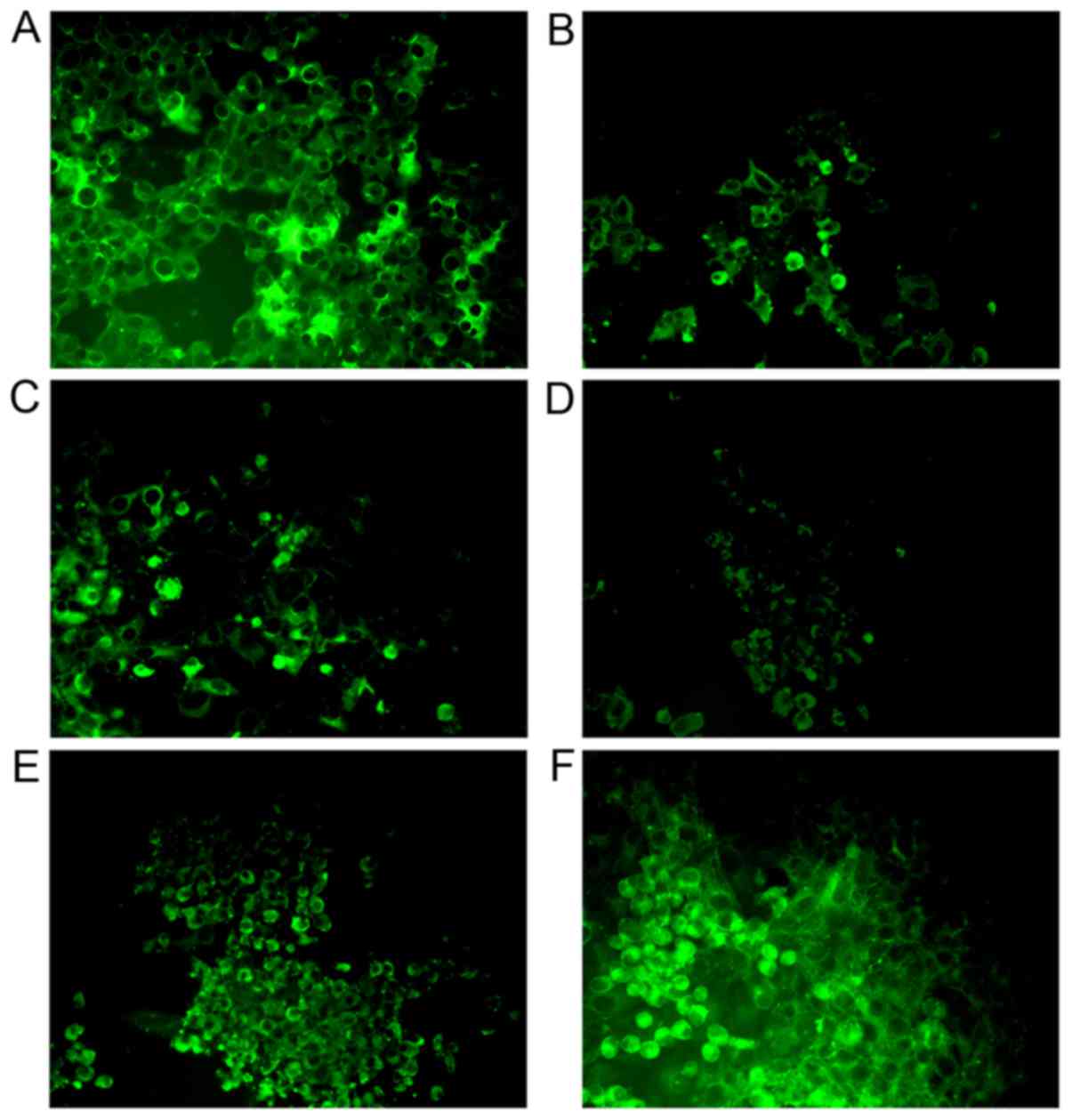
the EV71-infected Vero cells
The specificity of antibodies binding to the
EV71-infected Vero cells was determined by immunofluorescence. All
antibodies could specifically bind to EV71 in Vero cells, but mouse
Mab elicited by SP70 and Pabs from rabbit serum elicited by
inactivated EV71 showed the strongest specificity (Fig. 2).
Reactivity of mouse and rat Mabs
elicited by SP70 to SP70 alanine scanning mutated peptides
Mouse and rat Mabs purified from sub-cloned
hybridoma cells had good neutralizing titers (1:112.36 for mouse
Mab and 1:89.29 for rat Mab) (Table
II). Although triggered by the same antigen, the PEMs of Mabs
isolated from mice and rats were different (Fig. 3). The SP70 epitope binding affinity
of mouse Mab was enhanced by alanine substitution at sites 210,
212, 213, 214 and 221 (vs. wild-type peptide, all P<0.01;
vs. negative control peptide, all P<0.001), weakened by
alanine substitution at sites 208, 209, 211, 220 and 222
(vs. wild-type peptide, all P<0.001; vs. negative
control peptide, all P<0.001), and almost lost by alanine
substitution at sites 215, 216, 217, 218 and 219 (vs.
wild-type peptide, all P<0.001; vs. negative control
peptide, all P>0.05) (Fig.
3A).
 | Figure 3.Reactivity of mouse and rat monoclonal
antibodies elicited by SP70 to SP70 alanine scanning mutagenesis
peptides. The peptide-ELISA map was created with monoclonal
antibody binding to each peptide. Results were displayed by S/N
value, which is the ratio of ODsample to ODnegative
control 1. Negative control 1 was the negative control of the
peptide-ELISA map detection, used to eliminate the background of
the polypeptide itself. A peptide representing the coxsackievirus
A16 VP1 epitope (aa 208–222) was used a negative control of the
EV71 VP1 SP70 epitope (aa 208–222) (NC). (A) Mouse monoclonal
antibody (diluted 1:5,000). ***P<0.001 vs. NC group;
##P<0.01, ###P<0.001 vs.
wild-type (WT) SP70; ΔΔP<0.01,
ΔΔΔP<0.001 vs. peptide with alanine
substitution at position 6; ▲▲▲P<0.05 vs.
peptide with alanine substitution at position 14. (B) Rat
monoclonal antibody (diluted 1:1,000). *P<0.05, **P<0.01,
***P<0.001 vs. NC group; ##P<0.01,
###P<0.001 vs. WT SP70;
ΔΔP<0.01, ΔΔΔP<0.001
vs. peptide with alanine substitution at position 12;
▲▲▲P<0.05 vs. peptide with alanine
substitution at position 14. Data are shown as mean ± standard
deviation from three vaccinated mice or rats. Black arrows indicate
mutated SP70 peptides with the strongest affinity to monoclonal
antibodies. OD, optical density; aa, amino acid; NC, negative
control. |
| Table II.Neutralization titers of anti-EV71
antibodies. |
Table II.
Neutralization titers of anti-EV71
antibodies.
| Antibody | Antibody type | Source | Immunogen | Neutralization
titers |
|---|
| M-M | Monoclonal | Mouse | KLH-peptide | 112.36 |
| S-M-P1 | Polyclonal | Mouse | rVP1 | 56.11 |
| S-M-P2 | Polyclonal | Mouse | rVP1 | 52.43 |
| S-M-P3 | Polyclonal | Mouse | rVP1 | 50.27 |
| S-M-V1 | Polyclonal | Mouse | Inactivated
virus | 49.48 |
| S-M-V2 | Polyclonal | Mouse | Inactivated
virus | 45.19 |
| S-M-V3 | Polyclonal | Mouse | Inactivated
virus | 42.67 |
| M-R | Monoclonal | Rat | KLH-peptide | 89.29 |
| S-R-P1 | Polyclonal | Rabbit | rVP1 | 28.92 |
| S-R-P2 | Polyclonal | Rabbit | rVP1 | 27.35 |
| S-R-P3 | Polyclonal | Rabbit | rVP1 | 25.24 |
| S-R-V1 | Polyclonal | Rabbit | Inactivated
virus | 75.18 |
| S-R-V2 | Polyclonal | Rabbit | Inactivated
virus | 70.08 |
| S-R-V3 | Polyclonal | Rabbit | Inactivated
virus | 69.26 |
The SP70 epitope binding affinity of rat Mab was
increased by alanine substitution at sites 210, 217, 219 and 221
(vs. wild-type peptide, all P<0.01; vs. negative control
peptide, all P<0.001), weakened by alanine substitution at sites
211, 212, 215, 216, 220 and 222 (vs. wild-type peptide, all
P<0.001; vs. negative control peptide, all P<0.05), and
almost lost by alanine substitution at sites 208, 209, 213, 214 and
218 (vs. wild-type peptide, all P<0.001; vs. negative control
peptide, all P>0.05) (Fig.
3B).
Reactivity of mouse and rabbit serum
Pabs elicited by EV71 rVP1 to SP70 alanine scanning mutated
peptides
Mouse and rabbit serum Pabs elicited by Freund's
complete adjuvant containing rEV71-VP1 protein were collected.
Neutralizing activities of all serum Pabs were assessed by
micro-neutralization assay (Table
II). Rabbit serum Pabs neutralized rEV71-VP1 protein at titers
of 25.24 to 28.92, and mouse serum Pabs neutralized rEV71-VP1
protein at titers of 50.27 to 56.11.
PEMs of mouse serum Pabs elicited by rEV71-VP1
protein presented the same trend as mouse Mab elicited by SP70
(Fig. 4A). The ability of SP70
epitope binding to mouse serum Pabs was enhanced by mutations at
sites 213 and 221 (vs. wild-type peptide, both P<0.001).
Mutations at sites 210, 212 and 214 did not influence the affinity
(vs. wild-type peptide, all P>0.05; vs. negative control
peptide, all P<0.001). Mutation at site 209 weakened the
affinity (vs. wild-type peptide, P<0.05; vs. negative control
peptide, P<0.05). Mutations at the other sites led to loss of
affinity (vs. wild-type peptide, all P<0.05; vs. negative
control peptide, all P>0.05). Meanwhile, mutations at sites 214,
215 and 217 led to loss of recognition by the rabbit serum Pabs
elicited by rEV71-VP1 protein (vs. wild-type peptide; all
P<0.05; vs. negative control peptide, all P>0.05), while most
other mutations did not influence antibody binding (vs. wild-type
peptide, all P>0.05) (Fig.
4B).
Reactivity of mouse and rabbit serum Pabs elicited
by inactivated EV71 to SP70 alanine scanning mutated peptides.
Mouse and rabbit serum Pabs elicited by Freund's complete adjuvant
containing inactivated EV71 were collected. Neutralizing activities
of all serum Pabs were assessed by micro-neutralization assay
(Table II). Rabbit serum Pabs
neutralized inactivated EV71 at titers of 69.26 to 75.18, and mouse
serum Pabs neutralized rEV71-VP1 protein at titers of 42.67 to
49.48.
Mouse serum antibody elicited by inactivated EV71
(Fig. 5A) bound to wild-type SP70,
but lost its affinity for mutated peptides (vs. wild-type peptide,
all P<0.001; vs. negative control peptide, all P>0.05).
Conversely, rabbit serum antibody elicited by inactivated EV71
robustly recognized SP70 mutants (vs. wild-type peptide, all
P>0.05; vs. negative control peptide, all P<0.05) (Fig. 5B).
Human blood sample assay
Table III shows
five blood samples from EV71-infected patients had neutralizing
activity against SP70. Compared with the DMSO group, wild-type
peptide and peptides with mutations at the sites 209, 211, 214,
215, 216, 217, 219 and 221 had a higher reactivity with SP70
antibody in the serum (all P<0.05). Compared with the wild-type,
mutations at the sites 209, 219 and 221 of SP70 lead to increased
affinity with the serum antibodies (all P<0.05) (Table IV).
| Table III.Neutralizing activity of blood
samples from EV71-infected patients. |
Table III.
Neutralizing activity of blood
samples from EV71-infected patients.
|
| EV71 PCR | Neutralizing
activity |
|---|
| YY-164 | Positive | >1,024 |
| YY-142 | Positive |
1,024 |
| YY-148 | Positive |
768 |
| YY-15 | Positive |
1,024 |
| YY-123 | Positive | >1,024 |
| Table IV.Reactivity of blood samples from
EV71-infected patients to mutated SP70 peptides. |
Table IV.
Reactivity of blood samples from
EV71-infected patients to mutated SP70 peptides.
|
|
|
|
| 95% CI |
|
|---|
|
|
|
|
|
|
|
|---|
| Control | Peptide | Mean (I-J) | Standard error | Lower limit | Upper limit | P-value |
|---|
| DMSO | Wild-type | 29.2 | 6.7 | 15.8 | 42.5 | <0.001 |
|
| 1 |
1.8 | 6.7 | −11.5 | 15.2 |
0.783 |
|
| 2 | 55.4 | 6.7 | 42.1 | 68.8 | <0.001 |
|
| 3 | 18.5 | 6.7 |
5.2 | 31.8 |
0.007 |
|
| 4 | 43.9 | 6.7 | 30.6 | 57.2 | <0.001 |
|
| 5 |
9.2 | 6.7 | −4.1 | 22.6 |
0.171 |
|
| 6 |
9.6 | 6.7 | −3.8 | 22.9 |
0.156 |
|
| 7 | 21.1 | 6.7 |
7.8 | 34.4 |
0.002 |
|
| 8 | 45.2 | 6.7 | 31.8 | 58.5 | <0.001 |
|
| 9 | 22.4 | 6.7 |
9.1 | 35.7 |
0.001 |
|
| 10 | 32.6 | 6.7 | 19.3 | 45.9 | <0.001 |
|
| 11 | 11.6 | 6.7 | −1.8 | 24.9 |
0.088 |
|
| 12 | 54.6 | 6.7 | 41.3 | 67.9 | <0.001 |
|
| 13 |
4.7 | 6.7 | −8.6 | 18.0 |
0.484 |
|
| 14 | 56.2 | 6.7 | 42.9 | 69.5 | <0.001 |
|
| 15 |
8.4 | 6.7 | −4.9 | 21.7 |
0.212 |
|
| NC | 18.7 | 6.7 |
5.4 | 32.0 |
0.007 |
| Wild-type | 1 | −27.3 | 6.7 | −40.6 | −14.0 | <0.001 |
|
| 2 | 26.3 | 6.7 | 13.0 | 39.6 | <0.001 |
|
| 3 | −10.7 | 6.7 | −24.0 |
2.6 |
0.114 |
|
| 4 | 14.7 | 6.7 |
1.4 | 28.0 |
0.031 |
|
| 5 | −19.9 | 6.7 | −33.2 | −6.6 |
0.004 |
|
| 6 | −19.6 | 6.7 | −32.9 | −6.3 |
0.005 |
|
| 7 | −8.1 | 6.7 | −21.4 |
5.3 |
0.232 |
|
| 8 | 16.0 | 6.7 |
2.7 | 29.3 |
0.019 |
|
| 9 | −6.7 | 6.7 | −20.1 |
6.6 |
0.316 |
|
| 10 |
3.4 | 6.7 | −9.9 | 16.8 |
0.610 |
|
| 11 | −17.6 | 6.7 | −30.9 | −4.3 |
0.010 |
|
| 12 | 25.4 | 6.7 | 12.1 | 38.8 | <0.001 |
|
| 13 | −24.5 | 6.7 | −37.8 | −11.1 | <0.001 |
|
| 14 | 27.0 | 6.7 | 13.7 | 40.4 | <0.001 |
|
| 15 | −20.7 | 6.7 | −34.1 | −7.4 |
0.003 |
|
| NC | −10.5 | 6.7 | −23.8 |
2.9 |
0.122 |
|
| DMSO | −29.2 | 6.7 | −42.5 | −15.8 | <0.001 |
Discussion
The EV71 epitope SP70 is the target of EV71
neutralizing antibodies (17).
Mice can produce neutralizing antibodies against EV71 after
vaccination with a synthetic SP70 peptide and suckling Balb/c mice
can fight EV71 infections by passive transfer of anti-SP70
antibodies (18). In the present
study, mice, rats, and rabbits were inoculated with inactivated
EV71, rEV71-VP1 protein, or KLH-conjugated EV71 SP70 peptide. All
immunization protocols generated EV71 neutralizing antibodies, with
titers ranging from 1:25.24 (serum Pabs of rabbit elicited by
rEV71-VP1 protein) to 112.36 (Mab of mouse elicited by SP70).
Immunofluorescence showed that all antibodies could specifically
bind to EV71 in Vero cells, but mouse Mab elicited by SP70 and Pabs
from rabbit serum elicited by inactivated EV71 showed the strongest
specificity.
Previous studies showed that changing a single AA
can change the affinity of the peptide to antibodies. Indeed,
Wunderlich et al (19)
showed the affinity of antibodies to the PA protein of influenza A
virus polymerase can be modulated by substituting single aa.
Similar observations were made when substituting single aa in the
hemagglutinin protein of the influenza virus (20) and in the capsid protein of porcine
circovirus 2 (21). In the present
study, an alanine scanning mutagenesis protocol was used to
synthesize fifteen SP70 mutated peptides. The affinity of
EV71-specific antibodies to these mutated peptides was assessed by
ELISA. The capacity of these antibodies to recognize SP70 mutants
differed among species. Mouse serum antibody elicited by
inactivated EV71 bound to wild-type SP70, but lost its affinity for
mutated peptides. Conversely, rabbit serum antibody elicited by
inactivated EV71 robustly recognized SP70 mutants. These
observations indicate that rabbit serum Pabs elicited by
inactivated EV71 were more flexible, recognizing a wider range of
SP70 variants. Meanwhile, mutations at sites 214, 215 and 217 led
to loss of recognition by the rabbit serum Pabs elicited by the
rEV71-VP1 protein, indicating that in rabbits the robustness of the
SP70-directed antibody response was weakened by vaccination with
only the rEV71-VP1 protein, rather than the whole virus. Two Mabs
from mice and rats, elicited by the same SP70 antigen, exhibited
reduced affinity for different mutant peptides, indicating that the
key antigenic SP70 residues differed between these animals. Liu
et al also observed differences in VP1-directed responses
among species following immunization with inactivated EV71
(22). These observations indicate
that the B cell repertoire to key antigenic residues in SP70
differs among species, as inoculation of rabbits, rats, and mice
elicited antibodies directed to different segments of the SP70
epitope.
Mouse serum antibody elicited by inactivated EV71
bound to wild-type SP70, but lost its affinity for mutated
peptides. The mouse serum Pabs elicited by EV71 rVP1 protein
presented the same trend as mouse Mab elicited by SP70. The rVP1
protein may present the SP70 epitope as a linear antigen. Our
observations thus suggest that the structure is also important for
SP70 epitope to elicit robust antibody responses, but this point
needs further study.
Surprisingly, we observed that some antibodies
exhibited enhanced affinity for specific SP70 mutants. The SP70
epitope binding affinity of the mouse Mab was enhanced by alanine
substitution at sites 210, 212, 213, 214 and 221, while the SP70
epitope binding affinity of the rat Mab was enhanced by alanine
substitution at sites 210, 217, 219 and 221. Furthermore, the SP70
epitope binding affinity of Pabs from mouse serum elicited by
rEV71-VP1 was enhanced by alanine substitution at sites 213 and
221. On the other hand, mouse serum antibody elicited by
inactivated EV71 bound to wild-type SP70, but lost affinity for
mutated peptides. These enhancements appear to be influenced by
species and structure of SP70. These observations also suggest that
an optimized SP70 peptide may possess higher immunogenicity in some
species. This peptide antigen has even been used in the production
of recombinant vaccines that have elicited neutralizing responses
in mice (12,23). These observations highlight the
potential role of this peptide as immunogen in EV71 vaccination
that may elicit a protective antibody response to a wide variety of
EV71 subtypes (22). Future work
should investigate the immunogenicity and the capacity of inducing
neutralizing antibody response for these mutant SP70 peptides in
mice or rabbits.
Antibody responses triggered by inactivated EV71,
rEV71-VP1, and KLH-conjugated EV71 SP70 peptide differed among
species in neutralizing capacity and affinity for SP70 mutant
peptides. Mutations in SP70 both reduced and enhanced antibody
affinity and these changes varied among the immunogen and species
used to generate the antibodies. These observations highlight the
potential for optimization of a novel SP70 subunit antigen for EV71
vaccine design.
Acknowledgements
Te present study was supported by Zhejiang Pukang
Biotechnology Co., Ltd. (Hangzhou, China) and they provided the
EV71/H3-TY virus strain. This study was supported by the Zhejiang
Province Science Department (2010C03005), the ‘Qianjiang’ Talent
Project from Zhejiang Province (2012R100081), and the Zhejiang
Province Key Laboratory of Biological Vaccine R&D
(2008F3022).
Glossary
Abbreviations
Abbreviations:
|
aa
|
amino acid
|
|
ATCC
|
American Type Culture Collection
|
|
DMSO
|
dimethyl sulfoxide
|
|
EV71
|
enterovirus 71
|
|
FBS
|
fetal bovine serum
|
|
HFMD
|
hand, foot, and mouth disease
|
|
KLH
|
keyhole limpet hemocyanin
|
|
LSD
|
least significant difference
|
|
MEM
|
minimal essential medium
|
|
OD
|
optical density
|
|
PEMs
|
peptide-ELISA maps
|
References
|
1
|
Sanders SA, Herrero LJ, McPhie K, Chow SS,
Craig ME, Dwyer DE, Rawlinson W and McMinn PC: Molecular
epidemiology of enterovirus 71 over two decades in an Australian
urban community. Arch Virol. 151:1003–1013. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sato C, Syoji M, Ueki Y, Sato Y, Okimura
Y, Saito N, Kikuchi N, Yagi T and Numakura H: Isolation of
enterovirus 71 from patients with hand, foot and mouth disease in a
local epidemic on March 2006, in Miyagi prefecture, Japan. Jpn J
Infect Dis. 59:3482006.PubMed/NCBI
|
|
3
|
Chang LY: Enterovirus 71 in Taiwan.
Pediatr Neonatol. 49:103–112. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tee KK, Takebe Y and Kamarulzaman A:
Emerging and re-emerging viruses in Malaysia, 1997–2007. Int J
Infect Dis. 13:307–318. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lum LC, Wong KT, Lam SK, Chua KB and Goh
AY: Neurogenic pulmonary oedema and enterovirus 71
encephalomyelitis. Lancet. 352:13911998. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mao LX, Wu B, Bao WX, Han FA, Xu L, Ge QJ,
Yang J, Yuan ZH, Miao CH, Huang XX, et al: Epidemiology of hand,
foot, and mouth disease and genotype characterization of
enterovirus 71 in Jiangsu, China. J Clin Virol. 49:100–104. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang Y, Tan XJ, Wang HY, Yan DM, Zhu SL,
Wang DY, Ji F, Wang XJ, Gao YJ, Chen L, et al: An outbreak of hand,
foot and mouth disease associated with subgenotype C4 of human
enterovirus 71 in Shandong, China. J Clin Virol. 44:262–267. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chan KP, Goh KT, Chong CY, Teo ES, Lau G
and Ling AE: Epidemic hand, foot and mouth disease caused by human
enterovirus 71, Singapore. Emerg Infect Dis. 9:78–85. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chang LY, Lin TY, Huang YC, Tsao KC, Shih
SR, Kuo ML, Ning HC, Chung PW and Kang CM: Comparison of
enterovirus 71 and coxsackie-virus A16 clinical illnesses during
the Taiwan enterovirus epidemic, 1998. Pediatr Infect Dis J.
18:1092–1096. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang SM, Liu CC, Tseng HW, Wang JR, Huang
CC, Chen YJ, Yang YJ, Lin SJ and Yeh TF: Clinical spectrum of
enterovirus 71 infection in children in southern Taiwan, with an
emphasis on neurological complications. Clin Infect Dis.
29:184–190. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xu J, Qian Y, Wang S, Serrano JM, Li W,
Huang Z and Lu S: EV71: An emerging infectious disease vaccine
target in the Far East? Vaccine. 28:3516–3521. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Foo DG, Alonso S, Chow VT and Poh CL:
Passive protection against lethal enterovirus 71 infection in
newborn mice by neutralizing antibodies elicited by a synthetic
peptide. Microbes Infect. 9:1299–1306. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Foo DG, Alonso S, Phoon MC, Ramachandran
NP, Chow VT and Poh CL: Identification of neutralizing linear
epitopes from the VP1 capsid protein of enterovirus 71 using
synthetic peptides. Virus Res. 125:61–68. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee MS and Chang LY: Development of
enterovirus 71 vaccines. Expert Rev Vaccines. 9:149–156. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Brown BA and Pallansch MA: Complete
nucleotide sequence of enterovirus 71 is distinct from poliovirus.
Virus Res. 39:195–205. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lim XF, Jia Q, Chow VT and Kwang J:
Characterization of a novel monoclonal antibody reactive against
the N-terminal region of enterovirus 71 VP1 capsid protein. J Virol
Methods. 188:76–82. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yokoyama WM: Production of monoclonal
antibodies. Curr Protoc Cell Biol Chapter. 16:Unit 16 1. 2001.
View Article : Google Scholar
|
|
18
|
Yokoyama WM, Christensen M, Dos Santos G,
Miller D, Ho J, Wu T, Dziegelewski M and Neethling FA: Production
of monoclonal antibodies. Curr Protoc Immunol. 102:Unit 2.5. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wunderlich K, Juozapaitis M, Ranadheera C,
Kessler U, Martin A, Eisel J, Beutling U, Frank R and Schwemmle M:
Identification of high-affinity PB1-derived peptides with enhanced
affinity to the PA protein of influenza A virus polymerase.
Antimicrob Agents Chemother. 55:696–702. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ping J, Li C, Deng G, Jiang Y, Tian G,
Zhang S, Bu Z and Chen H: Single-amino-acid mutation in the HA
alters the recognition of H9N2 influenza virus by a monoclonal
antibody. Biochem Biophys Res Commun. 371:168–171. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Saha D, Lefebvre DJ, Ooms K, Huang L,
Delputte PL, Van Doorsselaere J and Nauwynck HJ: Single amino acid
mutations in the capsid switch the neutralization phenotype of
porcine circovirus 2. J Gen Virol. 93:1548–1555. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu CC, Chou AH, Lien SP, Lin HY, Liu SJ,
Chang JY, Guo MS, Chow YH, Yang WS, Chang KH, et al: Identification
and characterization of a cross-neutralization epitope of
enterovirus 71. Vaccine. 29:4362–4372. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Plevka P, Perera R, Cardosa J, Kuhn RJ and
Rossmann MG: Crystal structure of human enterovirus 71. Science.
336:12742012. View Article : Google Scholar : PubMed/NCBI
|