Introduction
As one of the most active components of the ethanol
extracts from Sophoraflavescens, oxymatrine (OMT) has been
of primary research interest, in view of its biological activities
and wide range of pharmacological effects, including
anti-inflammatory, anti-allergic, anti-fibrotic and cardiovascular
protective activities (1,2). In particular, it has been extensively
used for the treatment of chronic hepatitis B, and exhibited high
clinical value (3). Previously,
oxymatrine has demonstrated an anti-tumor cell proliferative
effect, and synergistically enhances anti-tumor activity of
oxaliplatin in colon carcinoma (4). Li et al (5) revealed that oxymatrine suppresses
proliferation and facilitates apoptosis of human ovarian cancer
cells, through upregulating microRNA-29b and downregulating matrix
metallopeptidase-2 expression levels. The effects of oxymatrine on
the proliferation and apoptosis of SMMC-7721 human hepatoma
carcinoma cells have additionally been investigated, and have been
proven to exhibit a therapeutic advantage for future
chemoprevention or chemotherapy (6). Anti-proliferative abilities of
oxymatrine have additionally been observed in prostate cancer,
gastric cancer and leukemia cells (1,7,8), and
subsequently introduced oxymatrine as a potential novel therapeutic
in the prevention and treatment of cancer (1).
As one leading cause of cancer-associated mortality
globally, lung cancer predominantly comprises small-cell lung
carcinoma (SCLC) and non-small cell lung carcinoma (NSCLC), which
account for 80–85 and 15–20% of all lung cancer cases, respectively
(9,10). Despite scientific progress,
patients exhibiting NSCLC still experience short survival times and
high mortality (11). Conventional
treatments of lung cancer, including platinum-based chemotherapy
and/or radiation therapy are fairly ineffective, and result in a
host of negative side effects (12). As a result, the requirement for the
discovery of alternative novel drugs and less toxic therapeutic
strategies for the treatment of NSCLC is evident. Previous studies
have suggested that numerous natural extracts and chemically
synthesized compounds exhibit therapeutic efficacy as potential
candidates (13,14). The authors previously identified
several curcumin analogs with anti-proliferative effects on A549
lung cancer cells and studied their molecular mechanisms of
apoptosis, induced by the MHMD [(1E,6E)-4-(furan2yl)
methylene]-1,7-bis(4-hydroxy-3-methoxyphe-nyl) hepta-1,
6-diene-3,5-dione),
IHCH(2E,6E-2-(1H-indol-3-yl)methylene)-6-(4-hydroxy-3-methoxybenzylidene)-cyclohexanone)
and HBC (hydrazinobenzoylcurcumin), which represent possible
therapeutic agents (15–17).
In the present study, the compound oxymatrine was
applied to inhibit proliferation of A549 cells. It was demonstrated
that oxymatrine induced apoptosis of A549 cells using
Hoechst/propidium iodide (PI) staining, terminal deoxynucleotidyl
transferase dUTP nick end labeling assays (TUNEL) analysis and
caspase inhibition experiments. Furthermore, caspases-3, 8 and 9 of
A549 cells were revealed to be activated following treatment with
oxymatrine, which suggested that caspase-dependent extrinsic and
intrinsic mitochondrial pathways of apoptosis were involved in the
anti-proliferative effects of oxymatrine. The present study aimed
to elucidate the anti-proliferative molecular mechanism of
oxymatrine in lung cancer cells, and provide a potential candidate
to act as a future antitumor agent.
Materials and methods
Chemicals and cell culture
Oxymatrine was a kind gift from Professor Min Liu,
(The Xiang'an Hospital of Xiamen University, Xiamen, China). It was
dissolved in PBS solution (stock concentration of 100 mg/ml).
RPMI-1640 medium and newborn calf serum (NCS; Gibco, Thermo Fisher
Scientific, Inc., Waltham, MA, USA) were purchased from Beijing
DingguoChangsheng Biotechnology, Co. Ltd. (Beijing, China). A549
cells were cultured in 1640 medium supplemented with 10% calf serum
and 1% garamycin at 37°C in an incubator in an environment
containing 5% CO2.
Cell proliferation assay of
oxymatrine
A549 cells were seeded into 12-well culture plates
(Corning Incorporated, Corning, NY, USA) at a density of
1×105 cells per well by using Blood Counting chambers,
that would allow the formation of a 90% monolayer following 24 h. A
total of six different final concentrations of oxymatrine (0.1,
0.5, 1.0, 1.5, 2.0 and 2.5 mg/ml) were determined and added into
the wells. The treated cells were then incubated at 37°C for
different time periods. Pictures were taken by light microscope to
detect cell morphological alterations at different time points in
oxymatrine-treated A549 cells.
Cell viability assay
A549 cells were incubated in 96-well plates
(1×104 cells/well) in an incubator containing 5%
CO2 at 37°C. Then, cells were treated with oxymatrine at
different final concentrations for 24 h. A total of 20 µl MTT
[3-(4,5-dimethylthiazolyl-2)-2,5-diphenyltetrazolium bromide, stock
0.1 M] was added into each well and cells were placed in the dark
for 4 h at 37°C. Following aspiration of supernatant, dimethyl
sulfoxide (100 µl/well) was added to the wells, and cells were kept
in the dark at room temperature, on a mixer for 30 min. The
absorbance (OD570) was measured at a wavelength of 495
nm by a microplate spectrophotometer (BioTek China, Beijing, China)
to determine cell viability. All the experiments were repeated in
triplicate. P<0.05 and P<0.01 were considered to indicate a
statistically significant difference.
Fluorescence microscopy analysis
A549 cells were incubated in 6-well plates
(3×105 cells/well) and incubated in an environment
containing 5% CO2. Oxymatrine (1.5 mg/ml) was used to
treat cells in exponential growth period for 24, 48 and 72 h,
respectively. Following immobilization with 4% paraformaldehyde for
10 min at room temperature, A549 cells were stained with Hoechest
33342 (10 µg/ml) and PI, (10 µg/ml) for 10 min in 37°C incubator
successively. Following washing three times with PBS, the treated
cells were sealed with glycerol (50% in PBS) and then observed
under the fluorescence microscope (Nikon TE2000-U or Nikon 80i;
Nikon Corporation, Tokyo, Japan).
For the mitochondrial membrane potential assay, the
green JC-1 (GenView SA, Lausanne, Switzerland) signals were
measured at Ex 485 nm/Em 528 nm, whereas the red signals were
detected at Ex 540 nm/Em 590 nm. The absorbance was determined by a
microplate spectrophotometer (BioTek China).
For caspase inhibition analysis, A549 cells were
firstly incubated with 50 µM z-VAD-FMK (Beyotime Institute of
Biotechnology, Haimen, China) for 30 min. Then, the A549 cells were
treated with oxymatrine (1.5 mg/ml) for another 48 h. Finally, the
fluorescence microscope analysis was carried out in accordance with
the aforementioned procedure.
TUNEL analysis
A549 cells (3×105 cells/well) were
cultured under the aforementioned conditions, and oxymatrine (final
concentration of 1.5 mg/ml) was incubated with cells for 48 h.
Then, apoptotic cells were analyzed according to the TUNEL kit
manufacturer's protocol (DeadEND™ Colorimetric Apoptosis
Detection System; Promega Corporation, Madison, WI, USA).
Flow cytometry analysis
A549 cells (3×105 cells/ml) were seeded
into a 30-mm cell culture dish and treated with oxymatrine (1 mM)
for 24, 48 and 72 h, respectively. The A549 cells were fixed (70%
ethanol) for 2 h at 4°C, washed with PBS, permeabilized (0.1%
Triton X-100) and stained (PI-FITC) in accordance with the
procedure described in Zhou et al (17). Then, cell samples were employed for
the flow cytometry assay. Data were collected and analyzed by the
CellQuest Prosoftware (version 6.0; FACS Calibur; BD Biosciences,
Franklin Lakes, NJ, USA).
Caspase activity detection
Mock-treated or oxymatrine-treated (1.5 mg/ml) A549
cells (48 h) were harvested by scraping, washed with PBS, frozen
for 30 min and centrifuged (16,000 × g for 15 min at 4°C). The
supernatants were assayed by a plate reader (Model 550 Microplate
Reader; Bio-Rad Laboratories, Inc., Hercules, CA) at a wavelength
of 405 nm, for the activities of caspase-3, 8, 9 in the presence of
20 µM Ac-DEVD-pNA, Ac-IETD-pNA and Ac-LEHD-pNA (Beyotime Institute
of Biotechnology, Haimen, China), respectively. The experiments
were carried out in triplicate.
Wound healing assay
The migratory activity of A549 cells was determined
using a wound healing assay, as previously described (17). Briefly, one pipette tip was used to
make a scratch wound across each well in a 12-well plate. Following
washing and removing any loosely held cells, oxymatrine (1.5 mg/ml)
was added into each well for 24, 48 and 72 h, respectively. An
inverted microscope was used to track the migration process of A549
cells.
Statistical analysis
One-way analysis of variance followed by LSD and
Tukey post-hoc tests was used to evaluate differences between
groups. All data were analyzed by SPSS software (version 19.0; IBM
Corp., Armonk, NY, USA) and expressed as the mean ± standard error.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Oxymatrine induces an
anti-proliferative effect in A549 cells
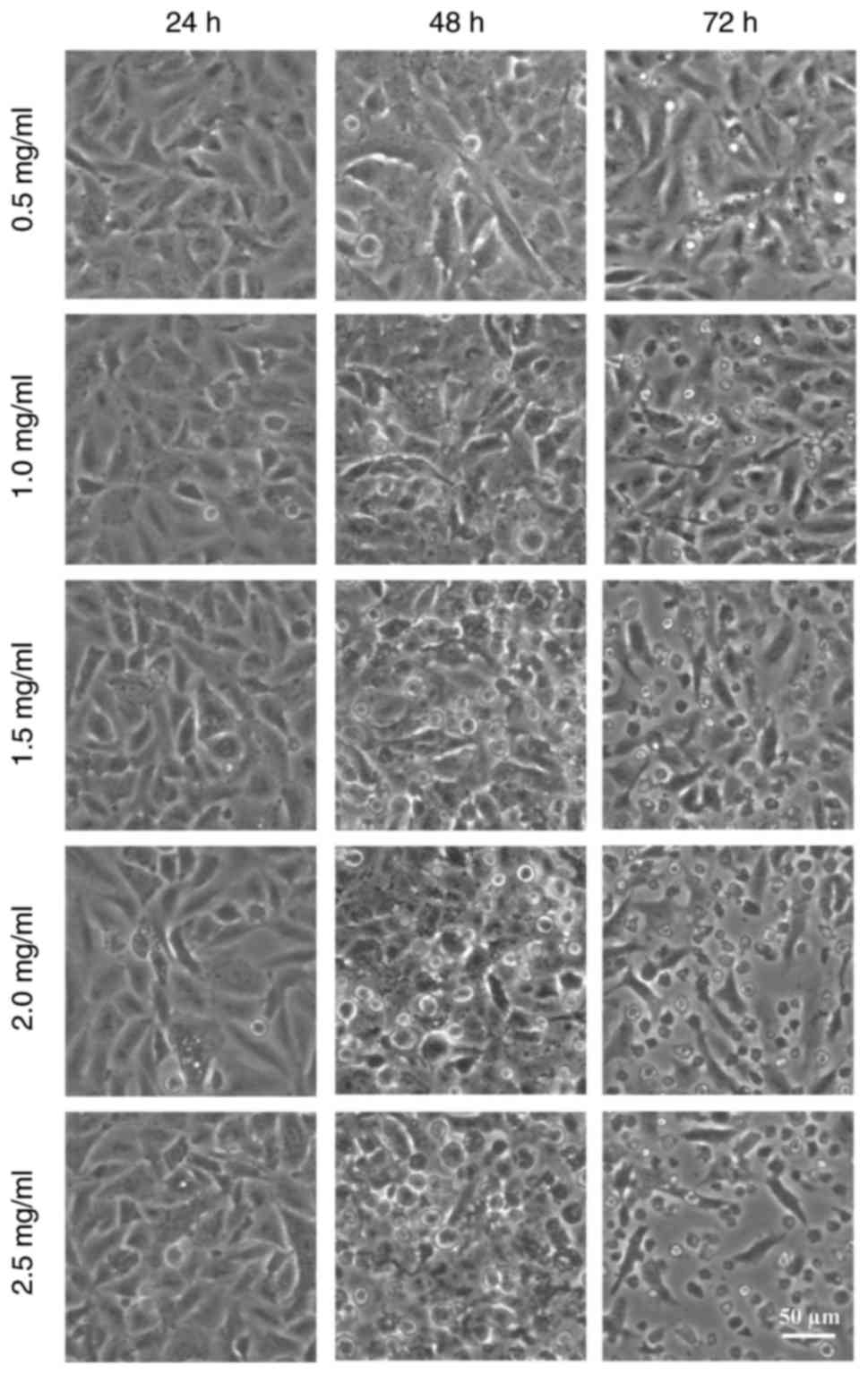
Microscope observation analysis indicated that A549
cells became rounded and granular following treatment with
oxymatrine at different concentrations (Fig. 1). At 24 h, the treated cells did
not appear to exhibit any properties characteristic of apoptosis.
However, over the increasing time period, the treated A549 cells
detached from the monolayer and floated in the cell medium, and
this effect appeared to be enhanced with increased concentrations
of oxymatrine.
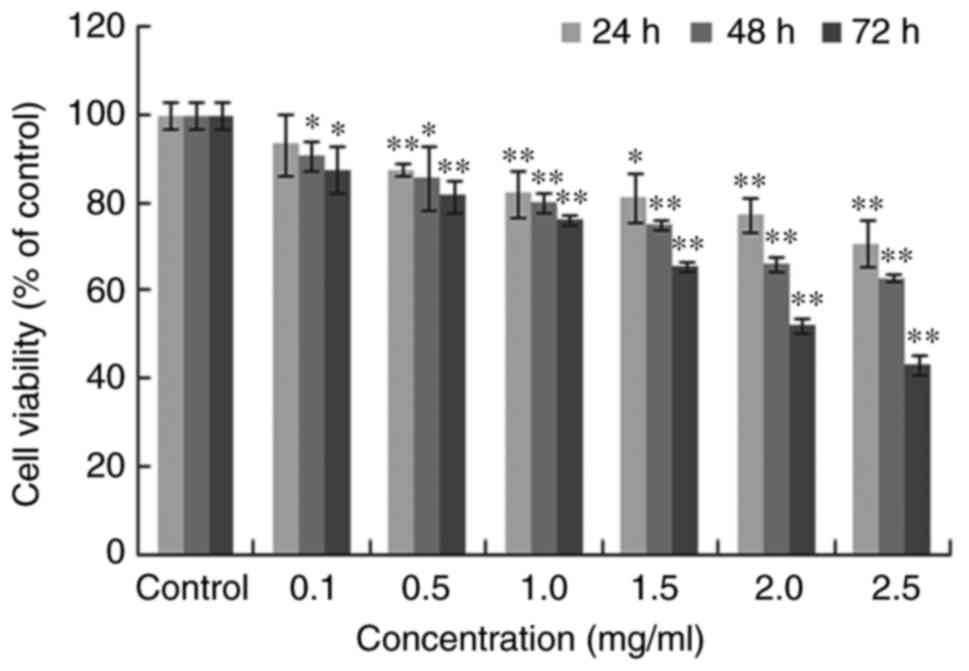
Furthermore, MTT analysis was applied to identify
the A549 cell viability following treatment. The experiments
demonstrated that oxymatrine inhibited A549 cell growth in a time-
and dose-dependent manner (Fig.
2). When the treatment concentration of oxymatrine increased,
the A549 cell viability continuously decreased. Overall, the
aforementioned results suggested that oxymatrine had an
anti-proliferative effect on the A549 lung cancer cells.
Oxymatrine induces apoptosis in A549
cells
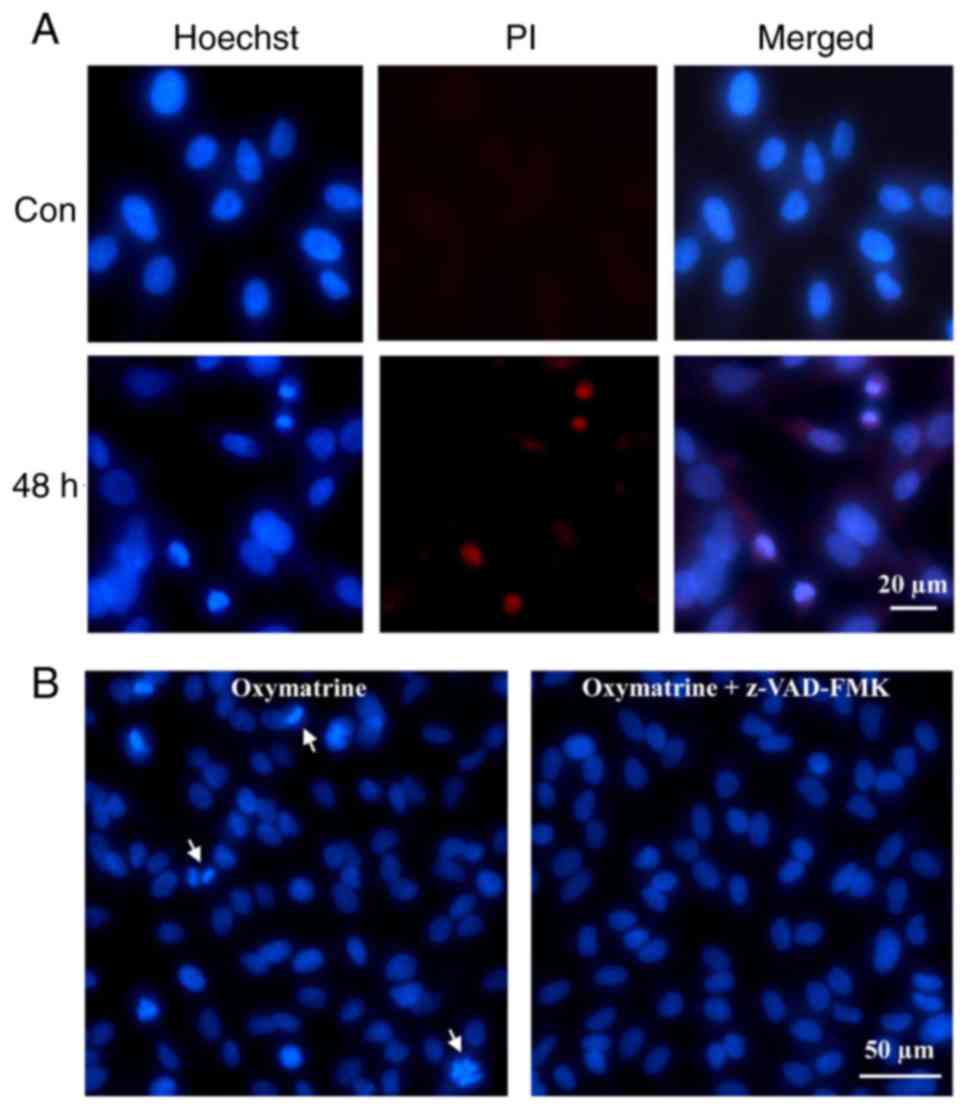
To further identify the apoptosis mechanism of A549
induced by oxymatrine, Hoechst/PI dual-staining assay was employed
to detect possible cell nuclei alterations. Generally, Hoechst
enters into the normal cytomembrane and stains the nucleolus blue,
whereas PI is a nucleic acid dye that only has access to the
damaged cytomembrane and stains nuclei red, therefore is often used
for assessment of the late apoptotic cells. Hoechst 33342 staining
exhibited apparent nuclear fragmentation at 48 h (Fig. 3A), whereas the nuclei of control
cells remained intact. The red fluorescence of PI was additionally
detected, suggesting that oxymatrine induce apoptosis of A549
cells.
Given that apoptosis is often caspase-dependent, the
caspase inhibitor z-VAD-FMK was used to pre-incubate the A549
cells, and then fluorescence observation analysis revealed that
z-VAD-FMK, in part, inhibited the fragmentation of A549 cell nuclei
(Fig. 3B). This suggested that the
oxymatrine-induced apoptosis of A549 cells was
caspase-dependent.
Further identification of
oxymatrine-induced apoptosis in A549 cells
Fluorescence microscopy analysis was used to study
apoptosis of A549 cells following oxymatrine treatment. In addition
to this, TUNEL and flow cytometry analysis were applied to further
verify the apoptotic characteristics of A549 cells. TUNEL analysis
demonstrated that the apoptotic cells contracted and the nuclei
appeared navy and the perinuclei brown in the oxymatrine-treated
group, whereas the positive signals were not detected in control
cells (Fig. 4).
Furthermore, as a lipophilic and cationic dye, JC-1
aggregates locate in areas with high mitochondrial membrane
potential (∆Ψm) and emits red fluorescence, and JC-1 monomers emit
green fluorescence, and locate in low ∆Ψm cell areas. The ratio of
red/green fluorescence acts as a marker of ∆Ψm alterations in cells
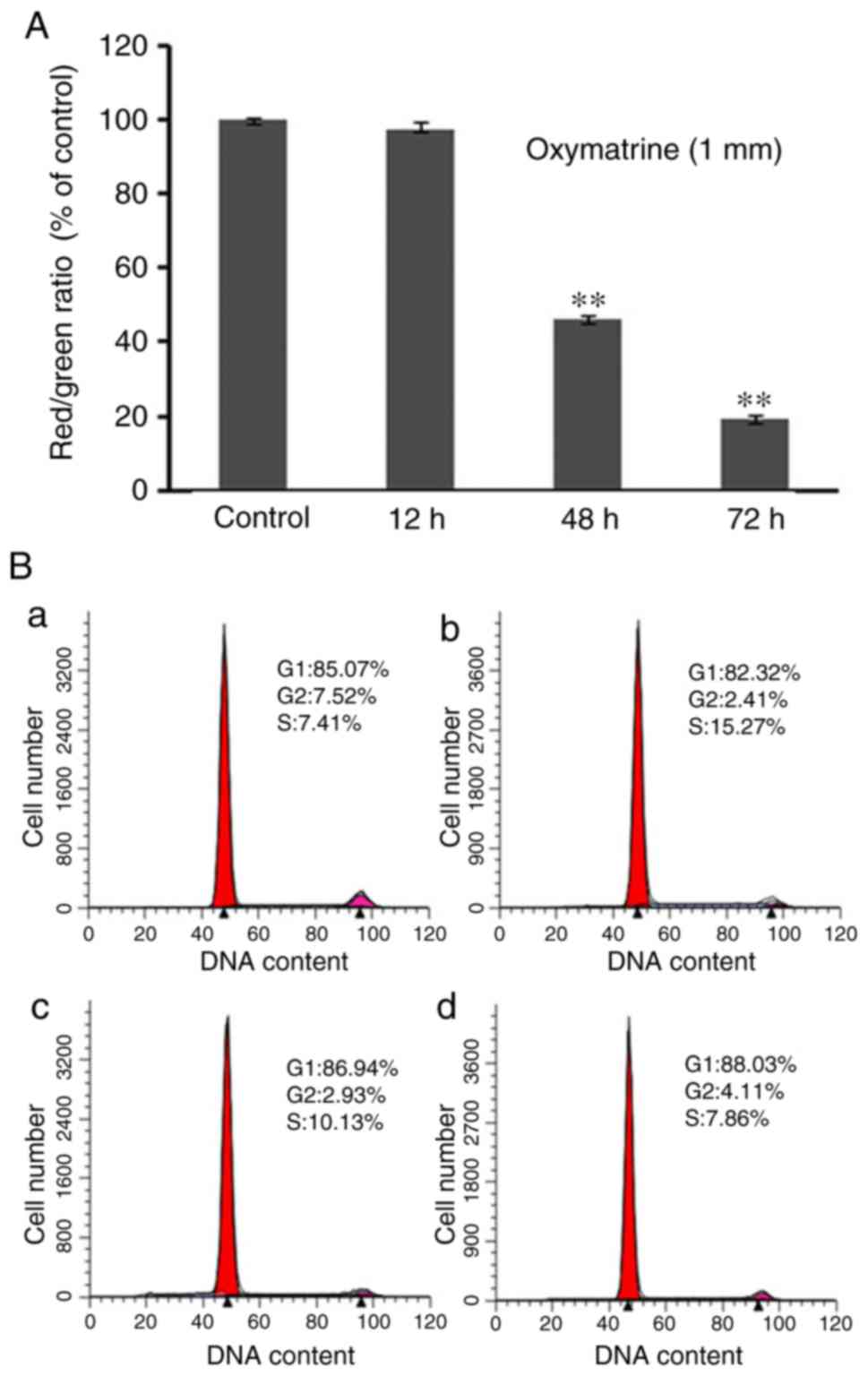
(18). In the present study, JC-1
staining revealed a decrease of red/green ratio following treatment
with oxymatrine for 12, 48 and 72 h, respectively (Fig. 5A). The decrease of ∆Ψm further
verified that oxymatrine induced A549 cell apoptosis.
In addition, to determine the cause of apoptosis,
flow cytometry analysis was applied to study the cell arrest of
oxymatrine-treated A549 cells. The results suggested that the DNA
synthesis decreased in a time dependent manner, and the number of
G0/G1 cells increased prior to 48 h (Fig. 5B). Compared with control cells,
treated groups at different time-points all revealed S phase
arrest. The above mentioned experiments verified that oxymatrine
resulted in A549 cell arrest and induced apoptosis. Future studies
should focus on the specific molecular apoptotic mechanism of A549
cells.
Oxymatrine-induced apoptosis in A549
cells is caspase-dependent
To further investigate whether the apoptosis induced
by oxymatrine was caspase-dependent or not, the oxymatrine-treated
(48 and 72 h) or untreated control A549 cells were harvested by
scraping. The supernatants of these cell lysates were detected for
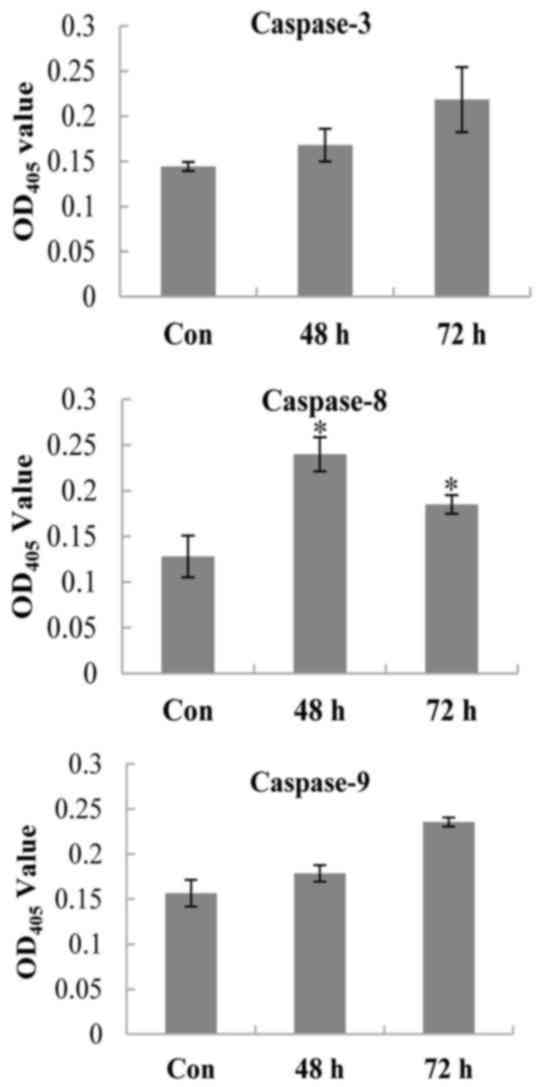
the activities of several key caspases in the process of apoptosis.
The results demonstrated that the absorbance (OD405nm,
n=3) of oxymatrine-treated cell supernatants all increased compared
with the control cells (P<0.05; Fig. 6) which suggested that caspases-3, 8
and 9 were activated following oxymatrine treatment in A549 cells,
and the induced apoptosis was caspase-dependent.
Oxymatrine inhibits migration of A549
cells
Due to their proliferative abilities, a further
property of malignant tumor cells is the ability to migrate
(19). The present study performed
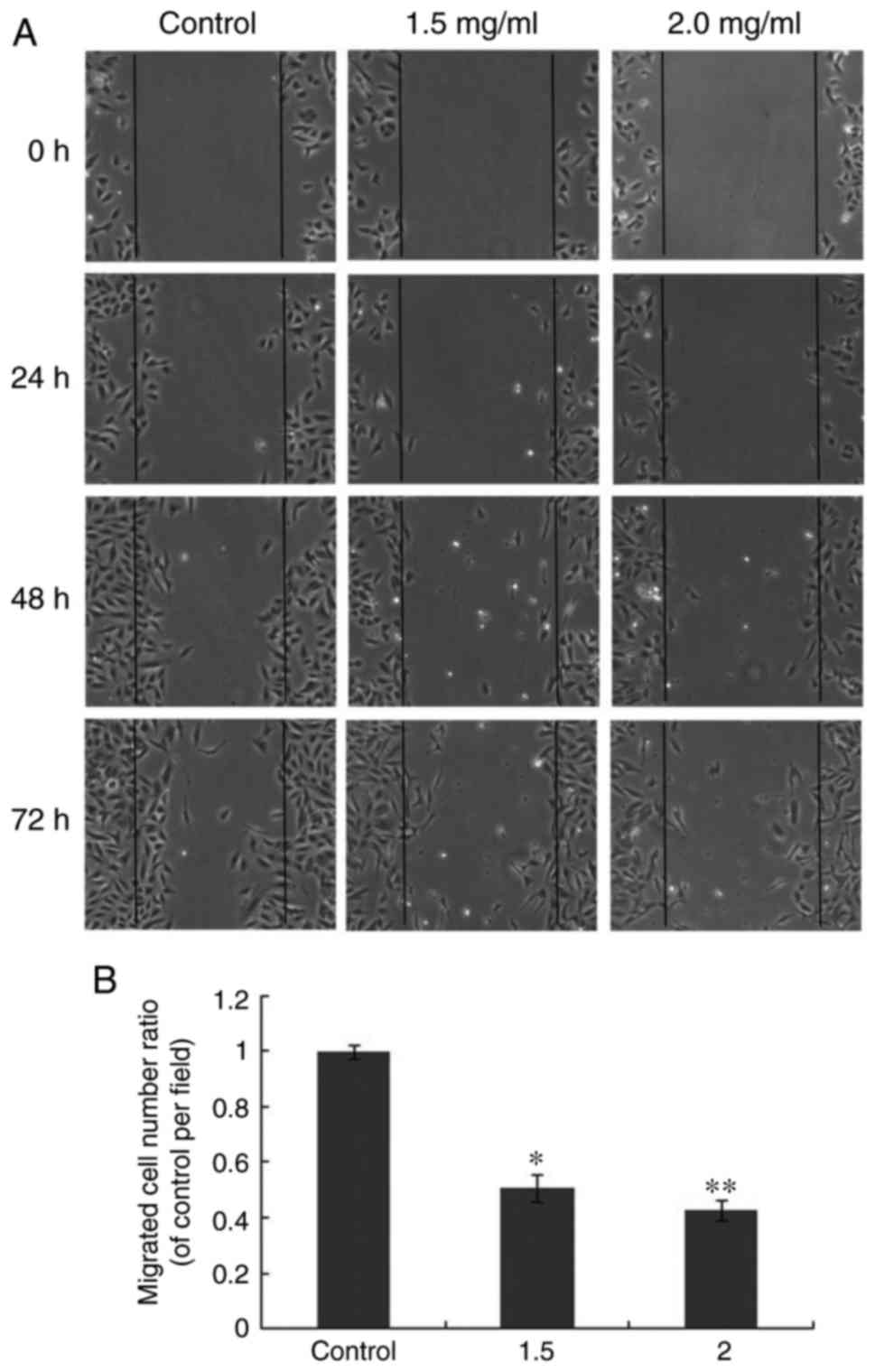
the wounding healing assay to examine the anti-migratory abilities
of oxymatrine. The results demonstrated that following treatment
with oxymatrine (1.5 and 2.0 mg/ml) for different time periods, a
decreased number of A549 cells migrated to the blank field
(Fig. 7A). Increased
concentrations of oxymatrine resulted in a smaller migrated cell
number ratio (Fig. 7B). This
suggested that oxymatrine inhibited migration of A549 cells. Future
investigations should focus on factors involved in the migration of
A549 cells following oxymatrine administration.
Discussion
The present study firstly applied traditional
techniques, including light and fluorescence microscope
observation, MTT analysis and TUNEL assay to detect the apoptotic
characteristics of A549 cells following oxymatrine treatment for
different time-periods, verifying that oxymatrine inhibited
proliferation of A549 cells by inducing cell apoptosis.
Furthermore, JC-1 staining was applied to monitor the decrease of
∆Ψm in oxymatrine-induced A549 cell apoptosis. Flow cytometry was
used to analyze the cell arrest of A549 cells following oxymatrine
treatment. The results of the aforementioned experiments
demonstrated an apoptotic effect of oxymatrine in A549 cells.
Finally, the caspase 3, 8 and 9 activities of oxymatrine-treated
A549 cells were detected, suggesting that all caspases were
activated following oxymatrine treatment at differing
concentrations. Caspases (particularly caspase-3) are critical in
regulating apoptosis and inflammation and caspase-dependent cell
apoptosis primarily results from extrinsic and intrinsic apoptotic
pathways. Caspase-8, caspase-9 and caspase-12 are the specific
caspases in the extrinsic and intrinsic apoptotic pathways
(20). The present study currently
analyzed the activities of the aforementioned three caspases in
oxymatrine-treated A549 cells, and results indicated increased
activity for all, compared with controls. It was therefore
suggested that oxymatrine induced apoptosis of A549 cells via
extrinsic and intrinsic pathways. The present study firstly
verified the apoptotic modes of oxymatrine-induced A549 lung
cancer, and following work should focus on the specific
apoptosis-associated cellular proteins, or other factors involved
in the anti-proliferative process.
Cell migration during cancer progression determines
the capacity of tumor cells to escape from the primary tumors and
invade adjacent tissues to finally form metastases, and cancer
cells modify their migration mechanisms in response to different
conditions, which makes cancer therapeutics designed to target
adhesion receptors or proteases in slowing tumor progression in
clinical trials inefficient (21).
Therefore, research into the cellular and molecular basis of these
different migration programs and developing novel chemical drugs
may lead to novel potential treatment strategies. The present study
applied the wound healing assay to assess the effect of oxymatrine
in A549 cells, and it was demonstrated that oxymatrine inhibited
the migration of A549 cells in vitro. However, whether its
anti-migration effect in lung cancer cells exists in vivo is
still not known, and future work should be carried out to
investigate the specific mechanism underlying A549 cell
dissemination.
Oxymatrine is a traditional Chinese medicine that
has been demonstrated to induce anti-cancer effects (1). The present study revealed that
oxymatrine inhibited proliferation of A549 cells. One previous
study also suggested that oxymatrine inhibits cell proliferation by
inducing apoptosis in human lung cancer A549 cells, however
specific apoptosis pathways remain to be elucidated (22).
In conclusion, the present study further
demonstrated that oxymatrine could induce A549 lung cancer cell
apoptosis via the extrinsic and intrinsic pathways.
Acknowledgements
The present study was supported, in part, by the
Science and Technology Planning Project of Henan Province, China
(grant no. 172102410082), the Foundation of Henan Educational
Committee, China (grant no. 16A180027), the Training Program for
Youth Backbone Teacher of Henan University of Technology (grant no.
001170) and the Foundation for University Key Teacher by the Henan
Province, China (grant no. 2015GGJS-041).
References
|
1
|
Liu Y, Xu Y, Ji W, Li X, Sun B, Gao Q and
Su C: Anti-tumor activities of matrine and oxymatrine: Literature
review. Tumor Biol. 35:5111–5119. 2014. View Article : Google Scholar
|
|
2
|
Wang CY, Bai XY and Wang CH: Traditional
Chinese medicine: A treasured natural resource of anticancer drug
research and development. Am J Chin Med. 42:543–559. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
He M, Wu Y, Wang M, Chen W and Jiang J:
Meta-analysis of the clinical value of oxymatrine on sustained
virological response in chronic hepatitis B. Ann Hepatol.
15:482–491. 2016.PubMed/NCBI
|
|
4
|
Liu Y, Bi T, Wang Z, Wu G, Qian L, Gao Q
and Shen G: Oxymatrine synergistically enhances antitumor activity
of oxaliplatin in colon carcinoma through PI3K/AKT/mTOR pathway.
Apoptosis. 21:1398–1407. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li J, Jiang K and Zhao F: Oxymatrine
suppresses proliferation and facilitates apoptosis of human ovarian
cancer cells through upregulating microRNA-29b and downregulating
matrix metalloproteinase-2 expression. Mol Med Rep. 12:5369–5374.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu Y, Bi T, Dai W, Wang G, Qian L, Gao Q
and Shen G: Effects of oxymatrine on the proliferation and
apoptosis of human hepatoma carcinoma cells. Technol Cancer Res
Treat. 15:487–497. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guo B, Zhang T, Su J, Wang K and Li X:
Oxymatrine targets EGFR(p-Tyr845) and inhibits EGFR-related
signaling pathways to suppress the proliferation and invasion of
gastric cancer cells. Cancer Chemother Pharmacol. 75:353–363. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu C, Huang W, Guo Y, Xia P, Sun X, Pan X
and Hu W: Oxymatrine inhibits the proliferation of prostate cancer
cells in vitro and in vivo. Mol Med Rep. 11:4129–4134. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Field JK and Duffy SW: Lung cancer
screening: The way forward. Br J Cancer. 99:557–562. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pore MM, Hiltermann TJ and Kruyt FA:
Targeting apoptosis pathways in lung cancer. Cancer Lett.
332:359–368. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Heist RS and Engelman JA: SnapShot:
Non-small cell lung cancer. Cancer Cell. 21:4482012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Azzoli CG, Temin S, Aliff T, Baker S Jr,
Brahmer J, Johnson DH, Laskin JL, Masters G, Milton D, Nordquist L,
et al: 2011 focused update of 2009 American Society of Clinical
Oncology clinical practice guideline update on chemotherapy for
stage IV non-small-cell lung cancer. J Clin Oncol. 29:3825–3831.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mehta HJ, Patel V and Sadikot RT: Curcumin
and lung cancer-a review. Target Oncol. 9:295–310. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Feng Y, Zhou J and Jiang Y: Resveratrol in
lung cancer-a systematic review. J BUON. 21:950–953.
2016.PubMed/NCBI
|
|
15
|
Zhou GZ, Xu SL, Sun GC and Chen XB: Novel
curcumin analogue IHCH exhibits potent anti-proliferative effects
by inducing autophagy in A549 lung cancer cells. Mol Med Rep.
10:441–446. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou GZ, Zhang SN, Zhang L, Sun GC and
Chen XB: A synthetic curcumin derivative hydrazinobenzoylcurcumin
induces autophagy in A549 lung cancer cells. Pharm Biol.
52:111–116. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou GZ, Cao FK and Du SW: The apoptotic
pathways in the curcumin analog MHMD-induced lung cancer cell death
and the essential role of actin polymerization during apoptosis.
Biomed Pharmacoth. 71:128–134. 2015. View Article : Google Scholar
|
|
18
|
Gamal-Eldeen AM, Hamdy NA, Abdel-Aziz HA,
El-Hussieny EA and Fakhr IM: Induction of intrinsic apoptosis
pathway in colon cancer HCT-116 cells by novel
2-substituted-5,6,7,8-tetrahydronaphthalene derivatives. Eur J Med
Chem. 77:323–333. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Polacheck WJ, Zervantonakis IK and Kamm
RD: Tumor cell migration in complex microenvironments. Cell Mol
Life Sci. 70:1335–1356. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fulda S and Debatin KM: Extrinsic versus
intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene.
25:4798–4811. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Friedl P and Wolf K: Tumor-cell invasion
and migration: Diversity and escape mechanisms. Nat Rev Cancer.
3:362–374. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang B, Han Q and Zhu Y: Oxymatrine
inhibited cell proliferation by inducing apoptosis in human lung
cancer A549 cells. Biomed Mater Eng. 26 Suppl 1:S165–S172.
2015.PubMed/NCBI
|