Introduction
Cancer is considered to be a process with multiple
stages, and carcinogenicity depends on various factors, including
the extent of oncogene activation and tumor suppressor gene
mutations. In addition carcinogenicity is governed by the duration
of carcinogenic factor exposure (1,2).
Different physiochemical agents are known to induce carcinoma in
the skin of animals. The sequence of processes involved includes
damage to skin, tissue inflammation, formation of hyperplasia,
appearance of papillomas and subsequent squamous cell cancer
(3–5). The induction of papillomasmay be
achieved by the administration of
12-O-tetradecanoylphorbol-13-acetate (TPA) to animal skin, which
causes activation of oncogene transcribing factors, nuclear factor
(NF)-κB and transcription factor AP-1 (AP-1) (6–8).
Following the activation of NF-κB and AP-1, the rate of
inflammatory processes, the initiation of cell proliferation and
the production of papillomas are increased. Studies using models of
skin carcinoma have led to the identification of various factors
involved in the development and progression of skin tumor (3,4). The
use of mouse models of skin carcinoma led to the investigation of
the role of inhibitors of tumor induction and malignant
transformation in skin cancer (5).
The results of previous study revealed that bioactive natural
products isolated from plants may possess potent skin and
epithelial cancer prevention potential (9). The progression of tumors is enhanced
by cellular oxidative stress and inflammatory reactions, which
indicates that the inhibition of these processes is of importance
for the treatment of cancer (10).
Natural products isolated from organisms of marine origin
additionally serve as the source of a number of therapeutically
potent compounds (11). An example
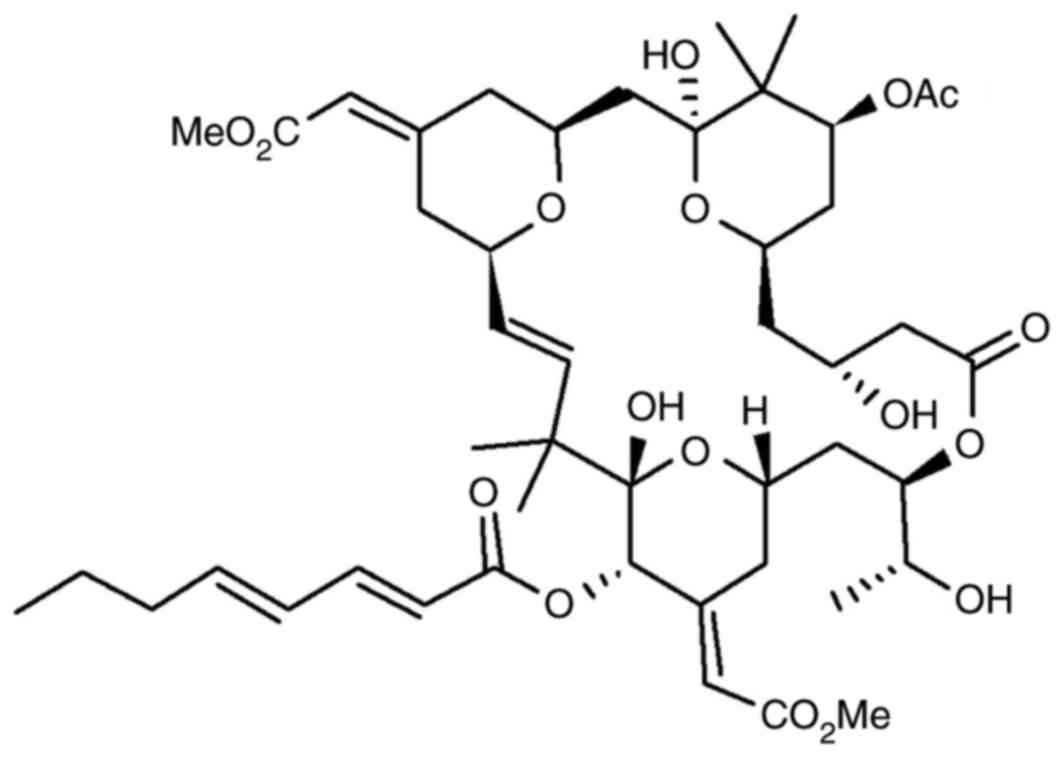
of such a molecule, bryostatin 1 (Fig.
1) is isolated from the invertebrate Bugulaneritina,
which is a member of the Ectoprocta phylum (11). Biological investigation has
revealed that bryostatin 1 aids in the development of the immune
system in addition to causing a promotion of hematopoietic
differentiation and stimulation (12). A previous study demonstrated that
treatment with bryostatin 1 resulted in the inhibition of phorbol
ester-mediated induction of carcinoma growth and proliferation
(13). In ovarian carcinoma,
treatment with bryostatin 1 induced cell growth inhibition via the
induction of apoptotic pathways (11,14).
In hepatic cancer, renal cancer, melanoma and leukaemia cell lines,
exposure to bryostatin 1 inhibited viability and induced apoptosis
(14–16). Studies involving rat models of
carcinoma have revealed that there is direct association between
the in vivo and in vitro anti-tumor potential of
bryostatin 1 (16). The present
study was designed to investigate the tumor inhibitory tendency of
bryostatin 1 in a TPA-induced mouse model of skin cancer. The
results demonstrated that bryostatin 1 inhibited the development
and progression of tumors of skin in mice through the prevention of
inflammation-inducing processes and the quenching of radicals.
Materials and methods
Chemicals and reagents
Bryostatin 1, benzo[α]pyrene and TPA were supplied
by Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
Animals
A total of 48 female 7-week old BALB/c nude mice
(12–20 g) were obtained from the Animal Experiment Center of
Beijing, China. The animals were housed in an animal facility
center under conditions free of pathogens with 12 hourly light and
dark cycle at 24°C with 30–70% humidity. All the mice were provided
with access to food and water ad libitum. The animal
experimental protocol was approved by the Animal Ethical Committee
of the Tongji Medical College of Huazhong University of Sciences
and Technology (Wuhan, China).
Treatment strategy
The day prior to the administration of bryostatin 1,
the animals were shaved on the upper side of the body to remove all
hair. The bryostatin 1 (30 µM) was administered 10 min prior to the
administration of TPA (7.2 mg). All animal protocols used in the
present study were approved by the Committee of Experimental Animal
Administration of the Second Military Medical University Laboratory
(Shanghai, China).
Analysis of bryostatin 1 radical
inhibition activity
For the determination of the radical inhibition
activity of bryostatin 1, 2,2-diphenyl-1-picrylhydrazyl (DPPH) was
used. From a 10-mM stock solution of DPPH in methyl alcohol, 30 µl
was added to a tube containing various concentrations of bryostatin
1 (5, 10, 15, 20, 25 and 30 µM). Subsequently, 3 ml methyl alcohol
was added to the tube, which was allowed to stand for 30 min. A
total of 1 ml distilled water and 3 ml toluene was added to the
tube, followed by centrifugation at 2,500 × g for 15 min at room
temperature. The absorbance of the tube was measured three times
independently at a wavelength of 517 nm.
In vivo radical inhibition activity of
bryostatin 1
The animals were shaved on the upper side of the
body one day prior to the start of the experiment. The following
day, 200 µl 2-propanone was applied to the body surface, followed
by the administration of 5 nmol TPA two times with an intervening
gap of 24 h. The administration of 5 nmol TPA was performed either
subsequent to pretreatment with bryostatin 1 or without bryostatin
1. Sacrifice of the mice was performed using pentobarbital sodium
(3%), 2 h subsequent to the administration of the second TPA dose.
The production of H2O2 induced by the
administration of TPA, and its inhibition by bryostatin 1, was
subsequently determined. For this purpose, the mouse epidermal
layers were extracted and treated with a buffer solution consisting
of potassium hydrogen phosphate (50 mM) and NaN3 (10
mM). The layers were subjected to homogenization using a Polytron
homogenizer (Kinematica AG, Lucerne, Switzerland) followed by 15
min centrifugation at (2,500 × g) at 4°C. A 0.5-ml quantity of the
collected supernatant was transferred to glass tubes, treated with
phenol red (0.2 mg/ml) and subsequently subjected to incubation for
15 min at 37°C. 1N (normal) solution of sodium hydroxide (100 µl)
was added to the tubes, followed by absorbance measurements at 612
nm. DMSO was used as control.
Skin myeloperoxidase (MPO) activity
determination
Skin tissues, following extraction, were subjected
to homogenization and treatment with a 0.5% solution of
C16H33N (CH3)3 Br in 50
mMK2HPO4 buffer using the Polytron
homogenizer. Following homogenization, the samples were centrifuged
for 20 min at 4°C at 2,000 × g. A mixture of 4-aminoantipyrine (25
mM), phenol (2%) and hydrogen peroxide (1.7 mM) was transferred to
a glass tube. A total of 0.5 ml tissue suspension was added to the
tube and mixed thoroughly. For each tube, the absorbance was
measured at a wavelength of 460 nm. The activity of MPO in the skin
tissue samples was expressed as units/g.
Determination of edema in mouse ear
tissue
The mice in the untreated group were treated with
TPA (7.2 mg) solution in 2-propanone into the right ear (6
mice/group). In the treatment group, mice were given bryostatin 1
prior to the application of TPA solution in 2-propanone, whereas
those in the normal control group received normal saline. Following
8 h of TPA administration, the mice were sacrificed using
pentobarbital sodium (3%) to extract an 8-mm ear-punch for
biopsies. The samples were weighed to record the increased weight
in the untreated group.
Analysis of ornithine decarboxylase
activity
The extracted skin samples from the mice were kept
in water for 45 sec at 60°C and subsequently transferred toice-cold
water. The epidermal layer of the skin was treated with 50 mM
KH2PO4 buffer mixed with 2 mM dithiothreitol
and 0.1 mM ethylene diamine tetra acetate. Homogenization of the
epidermal layer was performed using the Polytron instrument,
followed by centrifugation for 30 min at 12,000 × g at 4°C. The
activity of ornithine decarboxylase in the supernatant was
determined by monitoring the evolution of
14CO2 using liquid scintillation counting
(17).
Analysis of cyclooxygenase-2 (COX-2)
activity
The epidermal layer was lysed using
radioimmunoprecipitation assay buffer (Sigma-Aldrich, Merck KGaA)
and a DC™ protein assay (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) was used for determination of the protein
concentration in the samples. The extracted proteins (40 µg/lane)
were loaded and separated in 15% SDS-PAGE gel and then transferred
to polyvinylidene fluoride membranes. The non-specific sites in the
membranes were blocked for 15 min at room temperature via treatment
with non-fat dried milk (5%) in TBS with Tween-20 (TBS-T) buffer.
Incubation with primary antibodies against COX-2 (1:500; cat. no.
H-62; Santa Cruz Biotechnology Inc., Dallas, TX, USA) and α-tubulin
(1:500; cat. no. TU-02; Santa Cruz Biotechnology Inc.) for 1 h at
4°C was followed by washing with 4X TBS-T buffer. Membrane
incubation was performed with horseradish peroxidase-conjugated
anti-mouse immunoglobulin (1:10,000, cat. no. sc2005; Santa Cruz
Biotechnology Inc.) for 1 h at 4°C. Complexes formed by the
interaction between proteins and antibodies were analyzed using an
enhanced chemiluminescence detection system (GE Healthcare
Bio-Sciences, Pittsburgh, PA, USA).
Histological analysis of skin
Following administration of TPA, either with or
without pretreatment with bryostatin 1, the mice were sacrificed to
extract skin samples. The samples were fixed in 10% neutral
buffered formalin at 4°C overnight. Then, samples were embedded in
paraffin and sectioned at a thickness of 4 µm. The samples were
subjected to staining using hematoxylin and eosin for 10 min at
30°C. Examination of the skin samples was performed using light
microscope for the determination of epidermal thickness (indicators
of hyperplasia) and infiltration of polymorpho nuclear (PMN)
leukocytes (magnification, ×100). For each section, six different
randomly selected regions were observed.
Analysis of skin tumor formation
The mice were shaved on the upper side of the body
and treated with 2-propanone the following day. For tumor
initiation, benzo[α]pyrene dissolved in 2-propanone was applied to
the skin of the mice and the animals were kept under sterile
conditions for 7 days. Subsequently, the mice were administered
TPA, either without or with prior treatment with bryostatin 1.
Following three months, the number of skin tumors in each mouse
were counted with maximum size of 4 mm.
Statistical analysis
Data are presented as the mean ± standard error of
the mean of three independent experiments. Statistical analysis was
evaluated by one way analysis of variance followed by Bonferroni
multiple-comparison test unless otherwise stated. All data analyses
were performed using SPSS software package (version 17.0; SPSS,
Inc., Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
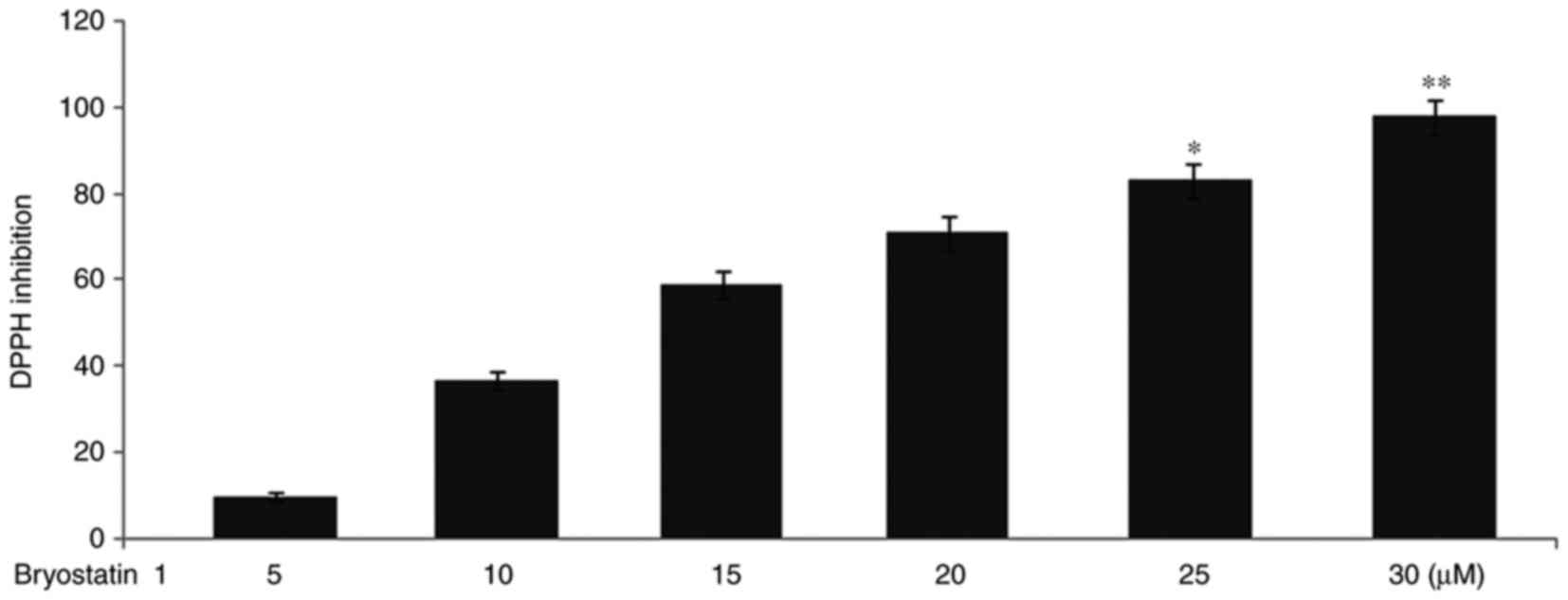
Bryostatin1 inhibits DPPH radical
activity
The radical inhibition potential of various doses of
bryostatin 1 was investigated against DPPH bleach in vitro.
With the increase in dosage of bryostatin1, a reduction was
observed in the DPPH radical activity. The DPPH radical potential
was reduced to 10, 37, 59, 71, 83 and 98%, respectively, at 5, 10,
15, 20, 25 and 30 µM doses of bryostatin 1 (Fig. 2).
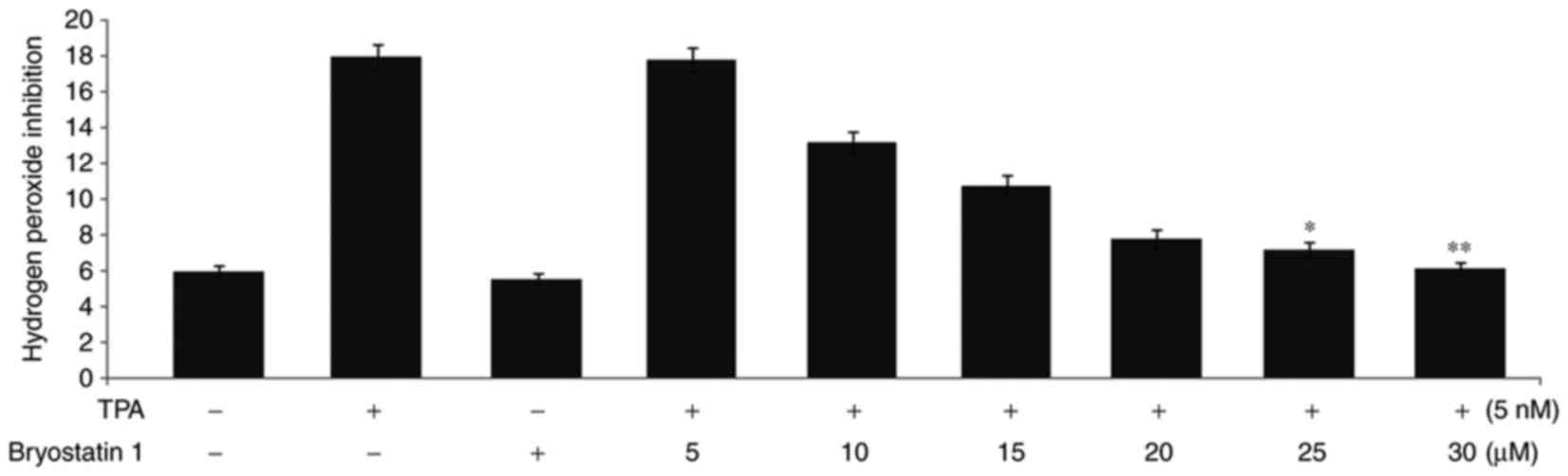
Pretreatment with Bryostatin1 prevents
the TPA-mediated increase in the level of
H2O2 and the activity of MPO
Administration of TPA to the mice at a concentration
of 5 nmol twice, with an intervening gap of 24 h, led to a 3-fold
increase in the H2O2 level in the epidermal
layer compared with normal mice (Fig.
3). However, treatment of the mice with bryostatin1 prior to
TPA administration prevented the increase in the level of
H2O2 in the mouse epidermal layer. The
H2O2 inhibition level was observed to be 3,
2.2, 1.8, 1.3, 1.2 and 1-fold higher in the mice following
treatment with 5, 10, 15, 20, 25 and 30 µM, respectively,
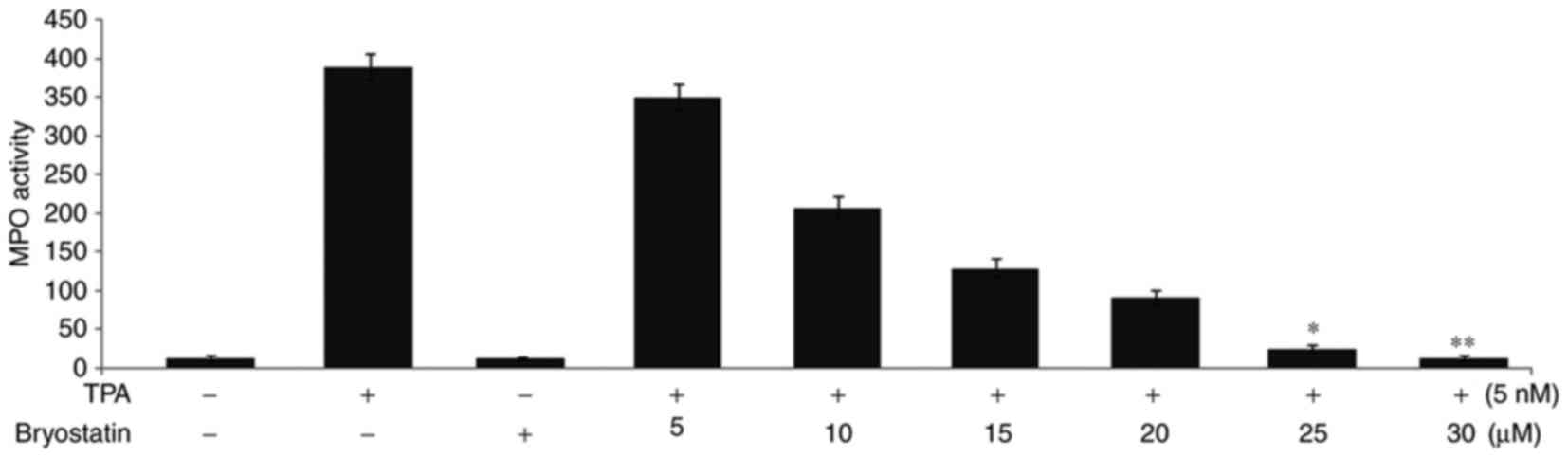
bryostatin 1. Administration of TPA to the mice at a concentration
of 5 nmol twice, with an intervening gap of 24 h, led to a 30-fold
increase in MPO activity compared with normal mice (Fig. 4). Treatment with bryostatin 1 prior
to TPA administration at doses of 5, 10, 15, 20, 25 and 30 µM,
respectively, reduced the level of MPO activity in a dose dependent
manner.
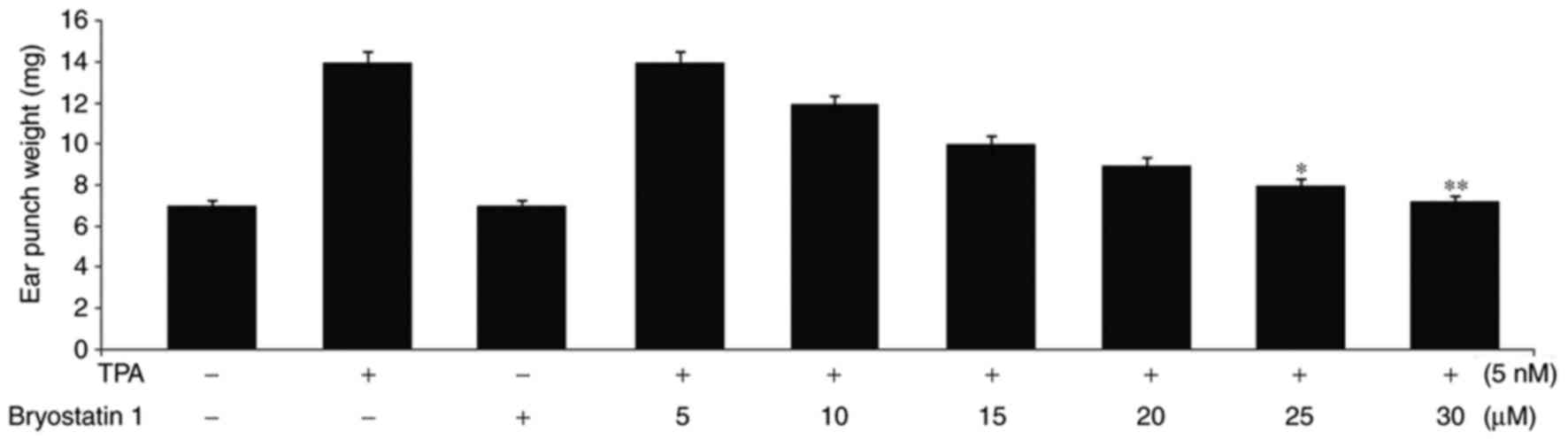
Bryostatin 1 prevents edema of the ear
and skin inflammation caused by TPA administration in mice
In mice administered with 5 nmol doses of TPA twice,
with an intervening gap of 24 h, edema formation was observed in
the ear. The mean weight of the extracted edema masses from the
mouse ears was recorded to be 14 mg. However, in mice pretreated
with bryostatin 1 at doses of 5, 10, 15, 20, 25 and 30 µM, the
edema ear masses were recorded to be 14, 12, 10, 9, 8 and 7.2 mg,
respectively (Fig. 5).
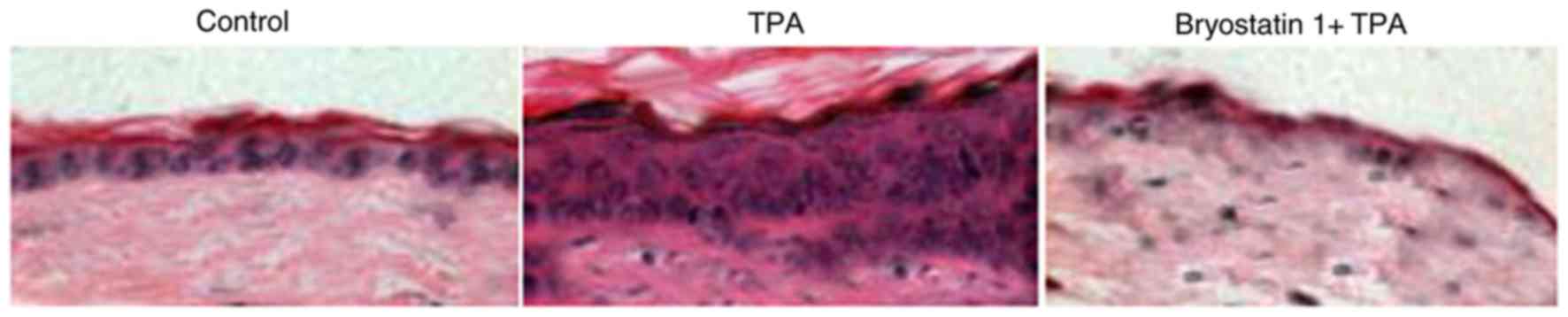
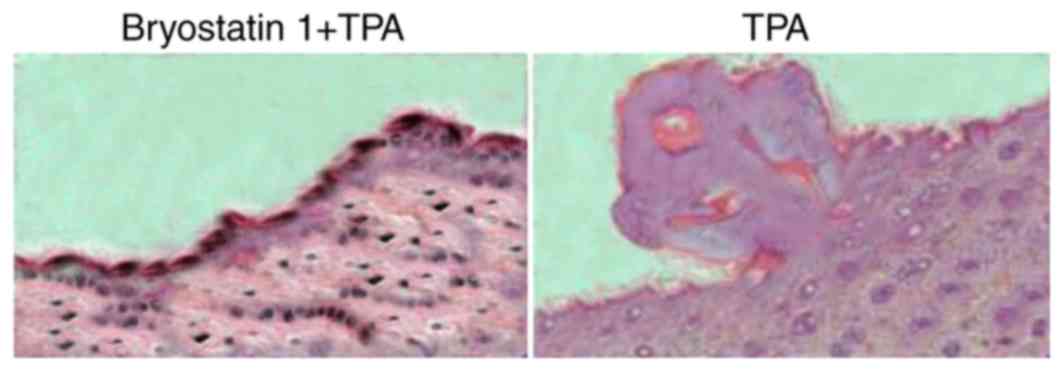
Administration of TPA to the mice for one week at a 5-nmoldose
caused hyperplasia in the epidermal layer and aggregation of
inflammatory cells leading to increased skin thickness. Treatment
of the mice with bryostatin 1 prior to TPA administration markedly
prevented inflammation of the skin by inhibiting hyperplasia in the
epidermal layer and the aggregation of inflammatory cells (Fig. 6). At a 30-µM concentration of
bryostatin 1, inflammation of the skin in the mice was completely
prevented.
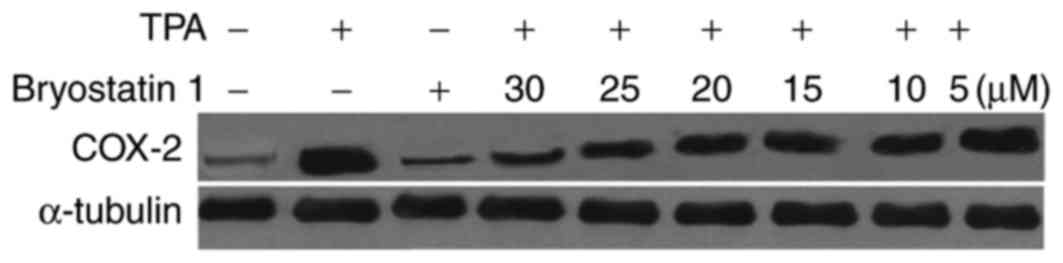
Bryostatin 1 inhibits the TPA-mediated
increased level of COX-2
The mice administered with TPA exhibited marked
increased levels of the protein COX-2 compared with the normal
group. However, treatment of the mice with bryostatin 1 prior to
TPA administration at a dosage of 30 µM inhibited the TPA-mediated
increase in the level of COX-2 (Fig.
7).
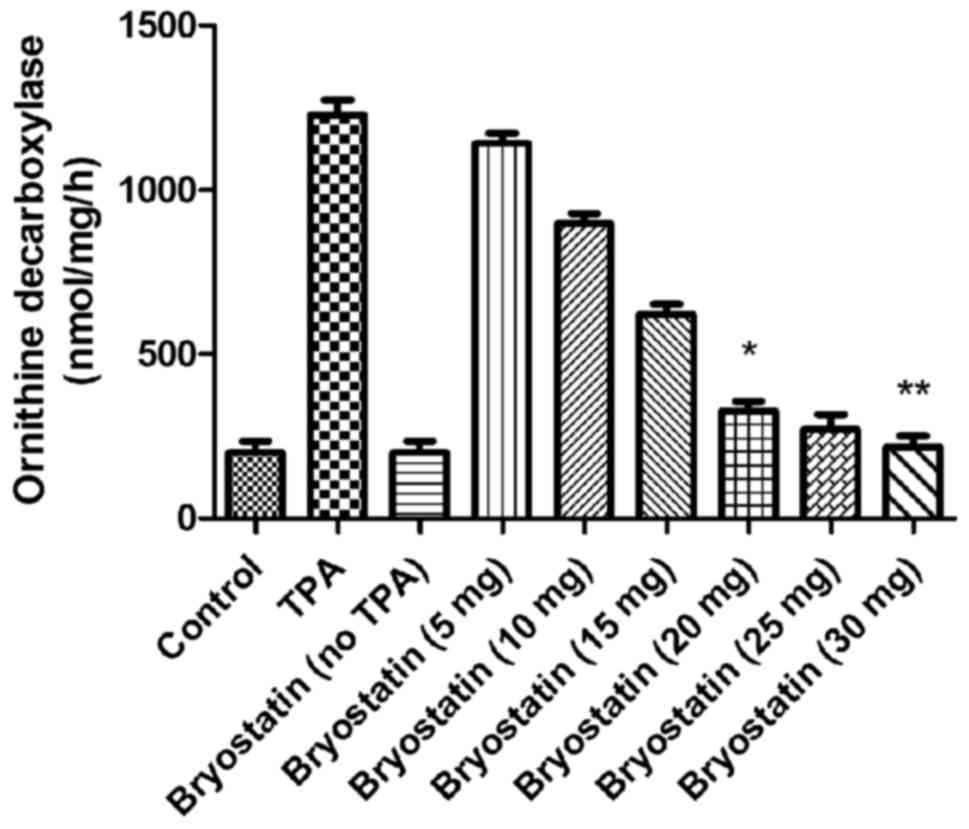
Bryostatin 1 inhibits the TPA-mediated
increased activity of ornithine decarboxylase
The activity of ornithine decarboxylase was
increased markedly in TPA-administered mice compared with the
control. However, when mice were treated with bryostatin 1 prior to
TPA administration at a dosage of 30 µM, the activity of ornithine
decarboxylase in the epidermal layer was significantly inhibited.
At 5, 10, 15, 20, 25 and 30 µM doses of bryostatin 1, the activity
level of ornithine decarboxylase was reduced to 1,150, 900, 630,
340, 290 and 240, respectively, compared with 1,200 in TPA group
(Fig. 8).
Bryostatin 1 inhibits TPA-mediated
increase in progression of tumor in mouse
Examination of the mice 3 months subsequent to the
administration of TPA demonstrated the appearance of papillomas in
all animals. In the mice treated with bryostatin 1 at 30-µM doses
prior to the administration of TPA, the appearance of the
papillomas was 20% at 2 months. Determination of papilloma
multiplicity revealed 5.2 papillomas on average in all the mice
administered with TPA. In the mice pretreated with bryostatin 1 at
30-µM doses prior to TPA administration, the mean papilloma number
in each animal was observed to be 0.4 (Fig. 9).
Discussion
The present study demonstrated the effect of
bryostatin 1 on the progression of tumors of the skin in
TPA-induced model skin cancer. The results demonstrated that
bryostatin 1 in habited the progression of skin tumor in mice by
suppressing the activity of ornithine decarboxylase and reducing
the level of COX-2. A previous study revealed that damage to
tissues caused by the onset of inflammatory reactions is associated
with the progress of carcinoma (18). The transduction of inflammatory
signals leads to an aggregation of immune cells and subsequent
expression of radicals at the site of inflammation, which in turn
destroy cellular components, including chromatin and membranes
(19). Therefore, radical species
serve an important role in the development and progression of
cancer. In view of theses finding, it is evident that compounds
which are able to quench radicals and inhibit inflammatory
reactions may be of value for the treatment of carcinoma. The
present study illustrated that bryostatin 1 exhibitedanti-oxidative
potential in vitro and in vivo. Bryostatin 1
inhibited the radical activity of DPPH completely at a 30-µM dose
in vitro. In vivo, pretreatment with bryostatin 1
prevented the formation of H2O2 in the
pidermal layer of TPA-mediated skin tumor model mice. The
expression of myeloperoxidase in the epidermal layer of skin tumor
model mice represents the production of radicals (20). The results of the present study
demonstrated that pretreatment with bryostatin 1 inhibited the
expression of myeloperoxidase in TPA-treated mice. It has been
reported that prostaglandin formation from arachidonic acid is
catalyzed by the enzyme COX (21).
It is known that COX-2 expression is enhanced by various factors,
including toxic secretions from bacteria, tumor promoting agents
and pro-inflammatory cytokines (22,23).
An increased level of COX-2 leads to the onset of inflammatory
reactions and promotes tumor progression (24). Therefore, it has been proposed that
diminishing the expression of COX-2 maybe of therapeutic value for
the prevention of inflammation (24). The present study demonstrated that
mice administered with TPA exhibited marked increased level of the
protein COX-2. However, treatment of the mouse with bryostatin1
prior to TPA administration inhibited the TPA-mediated increase in
the level of COX-2. Inflammation of tissues results in the
promotion of ornithine decarboxylase activity, which subsequently
serves an important role in the progression of skin tumors in mice
(25). Ornithine decarboxylase, on
activation, catalyzes the formation of polyamine in cells which
promotes the development of tumors (25). The results of the present study
demonstrated marked increase in the activity of ornithine
decarboxylase in the TPA-treated mice. However, treatment with
bryostatin 1 prior to TPA administration in habited he activity of
ornithine decarboxylase in the epidermal layer. Compounds derived
from plants are being evaluated for therapeutic value against
various disorders, with the aim of developing novel and efficient
treatment strategies (26). The
results of the present study revealed that bryostatin 1 reduced the
number of papillomas in each mouse in addition to the number of
mice with papillomas. Therefore, bryostatin 1 exhibited the
potential to act as a chemo preventive agent against skin tumor
development.
In conclusion, bryostatin 1 inhibited the
development and progression of tumors of the skin in mice through
the prevention of inflammation-inducing processes and the quenching
of radicals. Therefore, bryostatin 1 may be suggested to exert an
important role in the treatment of skin tumor.
References
|
1
|
Bishop JM: Molecular themes in on
cogenesis. Cell. 64:235–248. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Knudson AG: Hereditary predisposition to
cancer. Ann NY Acad Sci. 833:58–67. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Boutwell RK: Some biological aspects of
skin carcinogenisis. Prog Exp Tumor Res. 4:207–250. 1964.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Slaga TJ: Cancer: Etiology, mechanisms,
and prevention – a summary. Carcinog Compr Surv. 5:243–262.
1980.PubMed/NCBI
|
|
5
|
DiGiovanni J: Multistage carcinogenesis in
mouse skin. Pharmacol Ther. 54:63–128. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim EJ, Park H, Kim J and Park JH:
3,3′-diindolylmethane suppresses
12-O-tetradecanoylphorbol-13-acetate-induced inflammation and tumor
promotion in mouse skin via the downregulation of inflammatory
mediators. Mol Carcinog. 49:672–683. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Budunova IV, Perez P, Vaden VR, Spiegelman
VS, Slaga TJ and Jorcano JL: Increased expression of p50-NF-kappaB
and constitutive activation of NF-kappaB transcription factors
during mouse skin carcinogenesis. Oncogene. 18:7423–7431. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Przybyszewski J, Yaktine AL, Duysen E,
Blackwood D, Wang W, Au A and Birt DF: Inhibition of phorbol
ester-induced AP-1-DNA binding, c-Jun protein and c-jun mRNA by
dietary energy restriction is reversed by adrenalectomy in SENCAR
mouse epidermis. Carcinogenesis. 22:1421–1427. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Walaszek Z, Hanausek M and Slaga TJ:
Mechanisms of chemoprevention. Chest. 125 5 Suppl:128–133. 2004.
View Article : Google Scholar
|
|
10
|
De Flora S and Ferguson LR: Overview of
mechanisms of cancer chemopreventive agents. Mutat Res. 591:8–15.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pettit GR, Herald CL, Doubek DL, Herald
DL, Arnold E and Clardy J: Isolation and structure of bryostatin 1.
J Am Chem Soc. 104:6846–6848. 1982. View Article : Google Scholar
|
|
12
|
May WS and Sensenbrenner LL: Stimulation
of stem cell growth by the bryostatins. US Patent 5358711 A. Filed
September 10, 1993; issued November 23, 1994.
|
|
13
|
Hennings H, Blumberg PM, Pettit GR, Herald
CL, Shores R and Yuspa SH: Bryostatin 1, an activator of protein
kinase C, inhibits tumour promotion by phorbol esters in SENCAR
mouse skin. Carcinogenesis. 8:1343–1346. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dale IL and Gescher A: Effects of
activators of protein kinase C, including bryostatins 1 and 2 on
the growth of A549 human lung carcinoma cells. Int J Cancer.
43:158–163. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Krafft AS, William F, Pettit GR and Lilly
MB: Varied differentiation responses of human leukaemias to
bryostatin 1. Cancer Res. 49:1287–1293. 1989.PubMed/NCBI
|
|
16
|
Hornung RL, Pearson JW, Beckwith M and
Longo DL: Preclinical evaluation of bryostatin1 as an anticancer
agent against several murine tumour cell lines: in vitro versus in
vivo activity. Cancer Res. 52:101–107. 1992.PubMed/NCBI
|
|
17
|
Morgan MD: Oxidized polyamines and the
growth of human vascular endothelial cells. Prevention of cytotoxic
effects by selective acetylation. Biochem J. 242:347–352. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hussain SP, Hofseth LJ and Harris CC:
Radical cause of cancer. Nat Rev Cancer. 3:276–285. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Prescott SM and Fitzpatrick FA:
Cyclooxygenase-2 and carcinogenesis. Biochim Biophys Acta.
1470:M69–M78. 2000.PubMed/NCBI
|
|
20
|
Trush MA, Egner PA and Kensler TW:
Myeloperoxidase as a biomarker of skin irritation and inflammation.
Food Chem Toxicol. 32:143–147. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hamberg M and Samuelsson B: On the
mechanism of the biosynthesis of prostaglandins E-1 and F-1-alpha.
J Biol Chem. 242:5336–5343. 1967.PubMed/NCBI
|
|
22
|
Gately S: The contributions of
cyclooxygenase-2 to tumor angiogenesis. Cancer Metastasis Rev.
19:19–27. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Marton LJ and Pegg AE: Polyamines as
targets for therapeutic intervention. Annu Rev Pharmacol Toxicol.
35:55–91. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chun KS, Keum YS, Han SS, Song YS, Kim SH
and Surh YJ: Curcumin inhibits phorbol ester-induced expression of
cyclooxygenase-2 in mouse skin through suppression of extracellular
signal-regulated kinase activity and NF-kappaB activation.
Carcinogenesis. 24:1515–1524. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Coussens LM and Werb Z: Inflammation and
cancer. Nature. 420:860–867. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Surh YJ: Molecular mechanisms of chemo
preventive effects of selected dietary and medicinal phenolic
substances. Mutat Res. 428:305–327. 1999. View Article : Google Scholar : PubMed/NCBI
|