Introduction
Glucocorticoids (GCs) are frequently used clinically
to treat patients with organ transplantation and malignancies, in
addition to diseases, including inflammation, cancer and autoimmune
disorders (1,2). Patients exposed to GC therapy exhibit
osteonecrosis and osteoporosis via the direct responses of
osteoblasts, ultimately reducing bone formation (3,4) and
resulting in changes to cell number and function. It has been
reported that GCs can inhibit osteoblast proliferation and
differentiation, and induce apoptosis (5,6);
however the molecular mechanism underlying the involvement of GCs
in the differentiation and apoptosis of osteoblasts remains to be
fully elucidated. The effect of GCs on osteoblasts involves the
suppression of Wnt/β-catenin signaling (7) and activation of caspase-3, a common
downstream effector of several apoptotic signaling pathways
(8).
Based on similarities in structure and function,
β-ecdysterone, a major component of Chinese herbal medicines, is
likely to have similar protective effects to estrogen, which is the
primary therapeutic strategy for the treatment of osteoporosis
(9). Although β-ecdysterone has
various activities, including anabolic and hepatoprotective
effects, promoting the synthesis of collagen protein, protecting
cells from apoptosis and inducing their proliferation (10–13),
the signaling pathway involved in theβ-ecdysterone-mediated
protective effects remains to be elucidated.
Autophagy is a process of programmed cell death,
which is essential for cell growth, survival, differentiation and
homeostasis (14). Autophagy can
protect cells from apoptosis by removing oxidatively damaged
organelles, and excess autophagy can destroy cellular components.
To date, it has been recognized that estradiol inhibits osteoblast
apoptosis via the promotion of autophagy through the estrogen
receptor (ER)-extracellular signal-regulated protein
kinase-mammalian target of rapamycin (mTOR) pathway (15). GCs have been shown to induce
autophagy in an animal model of osteoporosis through decreasing the
ratio ofmicrotubule-associated protein 1 light chain 3 (LC3)-I to
LC3-II (16). However, whether
autophagy is present in GC-induced osteoblasts, and whether
autophagy is involved in the protective effect of β-ecdysterone
against apoptosis in osteoblasts remain to be elucidated.
The aim of the present study was to investigate
whether β-ecdysterone can improve the dexamethasone (Dex)-induced
effects on osteoblast differentiation and apoptosis, and whether
this is dependent on mTOR signaling pathway-dependent autophagy.
Preliminary investigations were also performed to determine the
potential role of β-ecdysterone in the prevention of
osteoporosis.
Materials and methods
Culture of bone marrow mesenchymal
stem cells (BMSCs) and osteoblasts
The BMSCs were obtained from 4-5-week-old male
C57BL/6 mice purchased from Slacom Experimental Animal Company
(Shanghai, China), as previously described (9). When the cells reached 80% confluence,
they were subcultured until the third passage. The BMSCs were
cultured in DMEM/F12 (Thermo Fisher Scientific, Inc., Waltham, MA,
USA) containing 10% FBS, 100 U/ml penicillin and 100 mg/l
streptomycin (Thermo Fisher Scientific, Inc.) in an incubator at
37°C, 100% humidity and 5% CO2. The induction and
culture of osteoblasts were performed as follows: Briefly, the
BMSCs were seeded in 24-well plates at a density of
5×104 and cultured in modified DMEM/F12 containing 10%
FBS, vitamin C (50 mg/l; Aladdin Shanghai Biochemical Technology
Co., Ltd., Shanghai, China), β-sodium glycerophosphate
(10−3 mol/l; Sigma-Aldrich; Merck Millipore, Darmstadt,
Germany), 100 U/ml penicillin, 100 mg/l streptomycin and Dex
(10−8 mol/l; Sigma-Aldrich; Merck Millipore) at 37°C in
a humidified atmosphere of 5% CO2. After 14 days, the
BMSCs were stained using a BCIP/NBT Alkaline Phosphatase Color
Development kit (cat. no. C3206; Beyotime Institute of
Biotechnology, Shanghai, China) according to the manufacturer's
instructions. The osteoblasts were cultured in DMEM/F12 in the same
manner as the BMSCs. The present study was approved by the ethics
committee of Xiaoshan TCM Hospital (Hangzhou, China).
CCK-8 assay
The osteoblasts were plated in 96-well plates
(5×104 cells/well) and treated with Dex in the absence
or presence of10−5 mol/l alendronate (Ale) or
β-ecdysterone at various concentrations
(10−7−10−5 mol/l) at 37°C. At the end of
treatment (24, 48 and 72 h), 10 µl of CCK-8 solution was added to
each well of the plate. Following incubation at 37°C for 1 h, the
culture medium containing CCK-8 solution was removed. The
absorbance at 450 nm was measured using a spectrophotometric
microplate reader (BioTek Instruments, Inc., Winooski, VT,
USA).
Measurement of alkaline phosphatase
(ALP) activity
The osteoblasts were plated in 24-well plates and
treated with Dex in the absence or presence of Ale (10−5
mol/l) or β-ecdysterone at various concentrations
(10−7−10−5 mol/l). At the end of treatment,
ALP activity was measured using an ALP assay kit according to the
manufacturer's instructions.
Cell apoptosis assay
To detect apoptosis, an APC Annexin V Apoptosis
Detection kit (cat. no. 88-8007; eBioscience, San Diego, CA, USA)
was used according to the manufacturer's instructions. Briefly, the
osteoblasts were treated with Dex in the presence or absence of Ale
(10−5 mol/l) or β-ecdysterone at various concentrations
(10−7−10−5 mol/l), washed with PBS and
incubated with 195 µl Annexin V-APC and 5 µl propidium iodide for
15 min at 4°C, prior to analysis by flow cytometry (BD Biosciences,
Franklin Lakes, NJ, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
The osteoblasts were cultured in DMEM/F12 and
treated with Dex in the presence or absence of Ale (10−5
mol/l) or β-ecdysterone at various concentrations
(10−7−10−5 mol/l). Total RNA was isolated
from cells using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) and detected by agarose gel electrophoresis. As
the template for PCR, cDNA was synthesized from 5 µg of RNA using
AMV reverse transcriptase (Fermentas; Thermo Fisher Scientific,
Inc.). The RT-qPCR procedure were performed using SYBR®
Green 10X Supermix (Takara Bio, Inc., Otsu, Japan) in a 25 µl total
volume (12.5 µl SYBR® Green 10X SuperMix, 0.5 µl forward
primer, 0.5 µl reverse primer, 9.5 µl ddH2O and 2 µl
cDNA) on a Roche Light Cycler® 480 II system (Roche
Diagnostics, Basel, Switzerland). The primer sequences are shown in
Table I. The gene expression
levels were normalized to the levels of GADPH. The gene expression
levels were calculated using the 2−ΔΔCq method (17).
 | Table I.Primes sequences used in the present
study. |
Table I.
Primes sequences used in the present
study.
| Gene | Sequence |
|---|
| Runx2 |
F:5′-TTTGCCCTCATCCTTCAC-3′ |
| Runx2 |
R:5′-GCTTCTGCTACCACTCTAAC-3′ |
| OCN |
F:5′-GCCCTAAAGCCAAACTCTG-3′ |
| OCN |
R:5′-GCTGCTGTGACATCCATAC-3′ |
| RANKL |
F:5′-GGGCTTTCAAAGTTCAGG-3′ |
| RANKL |
R:5′-GGGCTGTGAGTTTCATAC-3′ |
| Beclin-1 |
F:5′-GCATCCTTTCCCTCTTTC-3′ |
| Beclin-1 |
R:5′-CCACAAGCATCTCATCTC-3′ |
| ATG5 |
F:5′-AACCACCTTGAGTCAGGACAAC-3′ |
| ATG5 |
R:5′-TCGGCTGCATTGCATTTCAC-3′ |
| GAPDH |
F:5′-ATCACTGCCACCCAGAAG-3′ |
| GAPDH |
R:5′-TCCACGACGGACACATTG-3′ |
Protein extraction and western blot
analysis
The osteoblasts were lysed in ice-cold
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology) containing 0.01% protease and phosphatase inhibitor
(Sigma-Aldrich; Merck Millipore), and incubated on ice for 30 min.
The protein concentration was assessed using a bicinchoninic acid
protein assay kit (cat. no. PICPI23223; Thermo Fisher Scientific,
Inc.). The cell lysate was centrifuged at 12,000 × g at 4°C for 10
min, and the proteins in the supernatant were obtained (20–30 µg)
and separated on a 10% SDS-PAGE gel, followed by electrophoretic
transfer onto a polyvinylidene fluoride membrane (EMD Millipore,
Billerica, MA, USA). Following blocking with 5% skim milk, the
membrane was incubated at 4°C overnight with the following rabbit
monoclonal antibodies: Anti-runt-related transcription factor 2
(Runx2; cat. no. ab23981, 1:1,000), anti-osteocalcin (OCN; cat. no.
ab93876, 1:500), anti-RANK ligand (RANKL; cat. no. ab45039, 1:500),
anti-Beclin 1 (cat. no. ab55878, 1:500) and anti-autophagy protein
5 (ATG5; cat. no. ab108327, 1:1,000) (all from Abcam, Cambridge,
MA, USA), anti-B-cell lymphoma 2 (Bcl-2; cat. no. sc-492, 1:100;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA),
anti-Bcl-2-associated X protein (Bax; cat. no. sc-493, 1:100; Santa
Cruz Biotechnology, Inc.), anti-LC3-II (cat. no. 2775, 1:1,000;
Cell Signaling Technology, Inc., Danvers, MA, USA),
anti-phosphorylated (p)-mTOR (cat. no. 2971, 1:1,000), anti-mTOR
(cat. no. 2983, 1:1,000) and anti-GAPDH (cat. no. 5174, 1:1,500)
(all from Cell Signaling Technology, Inc.). The blots were then
incubated for 1 h at 37°C with goat anti-mouse or anti-rabbit
secondary antibody (cat. nos. A0216 and A0208; Beyotime Institute
of Biotechnology) and intensities were measured using enhanced
chemiluminescence (Thermo Fisher Scientific, Inc.). The signal
intensity was determined using ImageJ software version 1.46
(National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
Data are expressed as the mean ± standard deviation.
One-way analysis of variance, followed by an unpaired two-tailed
t-test was used for statistical analysis (GraphPad Prism 5
software; GraphPad Software, Inc., La Jolla, CA, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
Effect of β-ecdysterone on Dex-induced
osteoblast differentiation
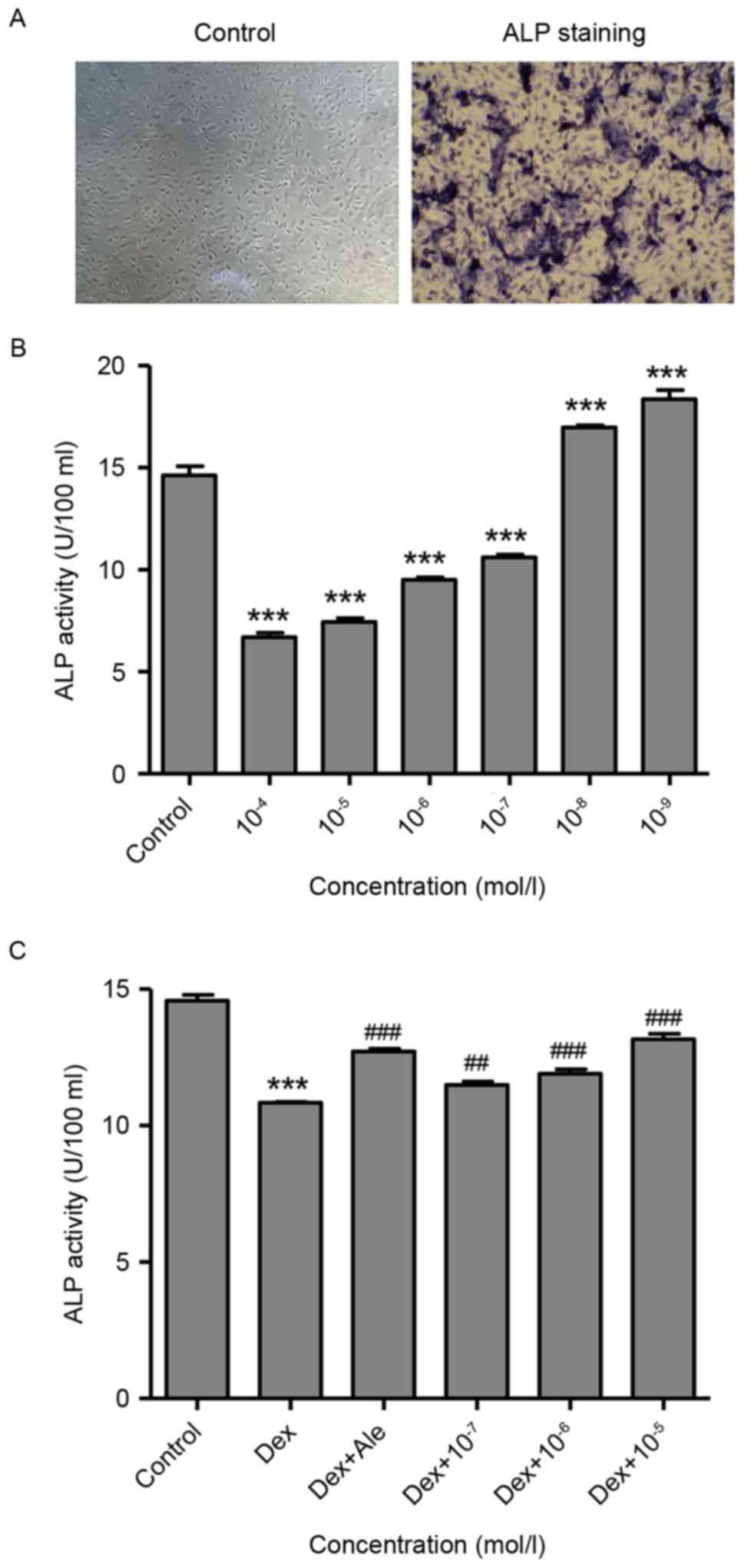
At 14 days post-induction, osteoblasts development
was observed, evidenced by purple staining present in the
extracellular matrix, which was measured using an inverted phase
contrast microscope (magnification, ×100; Fig. 1A). This suggested that the
osteoblast had been successfully induced from the BMSCs, which were
then used in the following experiments.
To investigate the effect of Dex on the activity of
ALP, which is an early marker of osteogenic differentiation, the
osteoblasts were treated with various concentrations of Dex
(10−9−10−4 mol/l) for 72 h, following which
the activity of ALP was measured. As shown in Fig. 1B, Dex significantly stimulated the
activity of ALP at lower concentrations
(10−9−10−8 mol/l) and suppressed the activity
of ALP at higher concentrations (10−7−10−4
mol/l), compared with the untreated control osteoblasts, and these
effects were dose-dependent. However, the cells treated with
β-ecdysterone at concentrations of 10−7−10−5
mol/l significantly stimulated the activity of ALP, compared with
the cells treated with 10−6 mol/l of Dex alone, which
was similar to activity in the cells treated with 10−5
mol/l of alendronate (Ale), which has long been used as a drug for
the treatment of osteoporosis and was included as a positive
control (Fig. 1C).
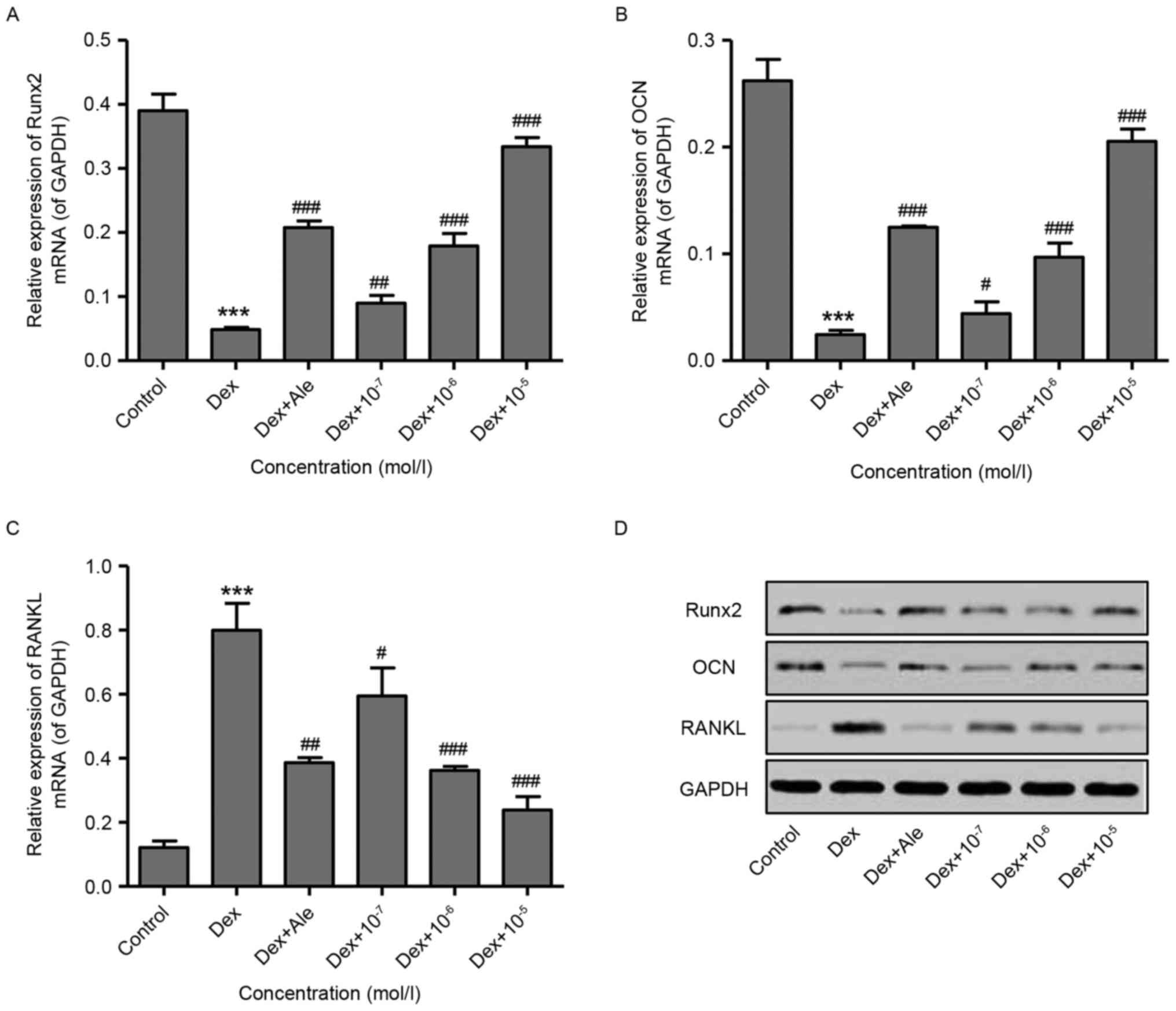
To confirm the effect of β-ecdysterone on the
induction of osteogenic differentiation, RT-qPCR and western blot
analyses were performed to examine gene expression. The results
showed that 10−6 mol/l of Dex significantly decreased
the expression levels of Runx2 and OCN, but increased the
expression of RANKL at the mRNA and protein levels (Fig. 2A-D). However, treatment of the
cells with β-ecdysterone at various concentrations
(10−7−10−5 mol/l) dose-dependently reversed
the changes in the expression of Runx2, OCN and RANKL induced by
10−6 mol/l of Dex. This was similar to the effects of
10−5 mol/l Ale on the osteoblasts (Fig. 2A-D). These results indicated that
β-ecdysterone reversed the Dex-induced suppression of osteoblast
differentiation.
Effect of β-ecdysterone on Dex-induced
osteoblast proliferation
The present study also examined the effects of
β-ecdysterone on Dex-induced osteoblast proliferation using a CCK-8
assay. The results showed that Dex treatment for 24 h markedly
increased cell proliferation at lower concentrations
(10−9−10−8 mol/l; Fig. 3A) and suppressed cell proliferation
at higher concentrations (10−6−10−4 mol/l)
after 48 and 72 h treatment, compared with the control (Fig. 3B and C). With the exception of the
results over 24 h (Fig. 3D),
significant increases in cell proliferation were observed in the
cells treated with β-ecdysterone at various concentrations
(10−7−10−5 mol/l) for 48 and 72 h (Fig. 3E and F), compared with the cells
treated with 10−6 mol/l of Dexalone. This was similar to
the effect of treatment with 10−5 mol/l Ale.
Effect of β-ecdysterone on Dex-induced
osteoblast apoptosis
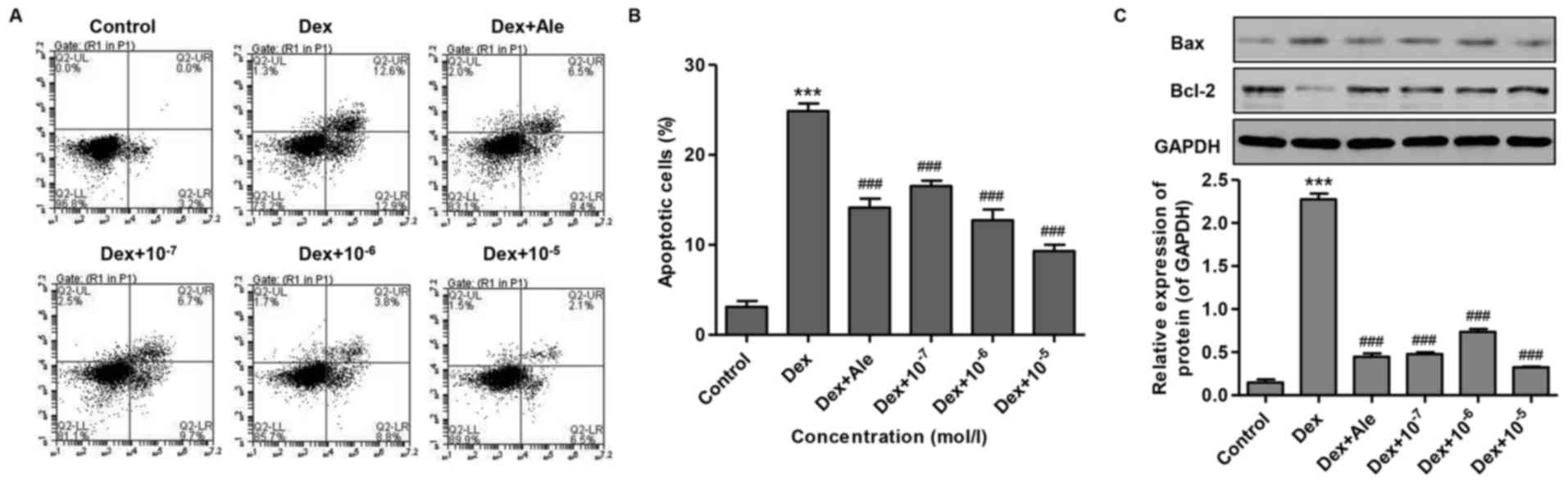
To investigate the effect of β-ecdysterone on
Dex-induced osteoblast apoptosis, the osteoblasts were treated with
10−6 mol/l of Dex for 72 h, and cell apoptosis was
measured. As shown in Fig. 4A and
B, Dex increased cell apoptosis 7-fold compared with that in
the control. However, the cells treated with β-ecdysterone at
various concentrations (10−7−10−5 mol/l)
exhibited significantly reduced cell apoptosis, compared with the
cells treated with 10−6 mol/l of Dex alone, which was
similar to the effect of treatment with 10−5 mol/l of
Ale.
To confirm the inducive effect of β-ecdysterone on
osteogenic apoptosis, western blot analysis was performed to
examine protein expression levels. The results showed that
10−6 mol/l of Dex significantly increased the ratio of
Bax/Bcl-2, compared with that in the control (Fig. 4C). However, in the cells treated
with β-ecdysterone at various concentrations
(10−7−10−5 mol/l), the alteration of the
Bax/Bcl-2 ratio induced by 10−6 mol/l of Dex was
reversed in a dose-dependent manner, which was similar to the
effect of treatment with 10−5 mol/l of Ale (Fig. 4C). These results suggested that
β-ecdysterone reversed the effect of Dex-induced osteoblast
apoptosis.
Effect of β-ecdysterone on Dex-induced
osteoblast autophagy
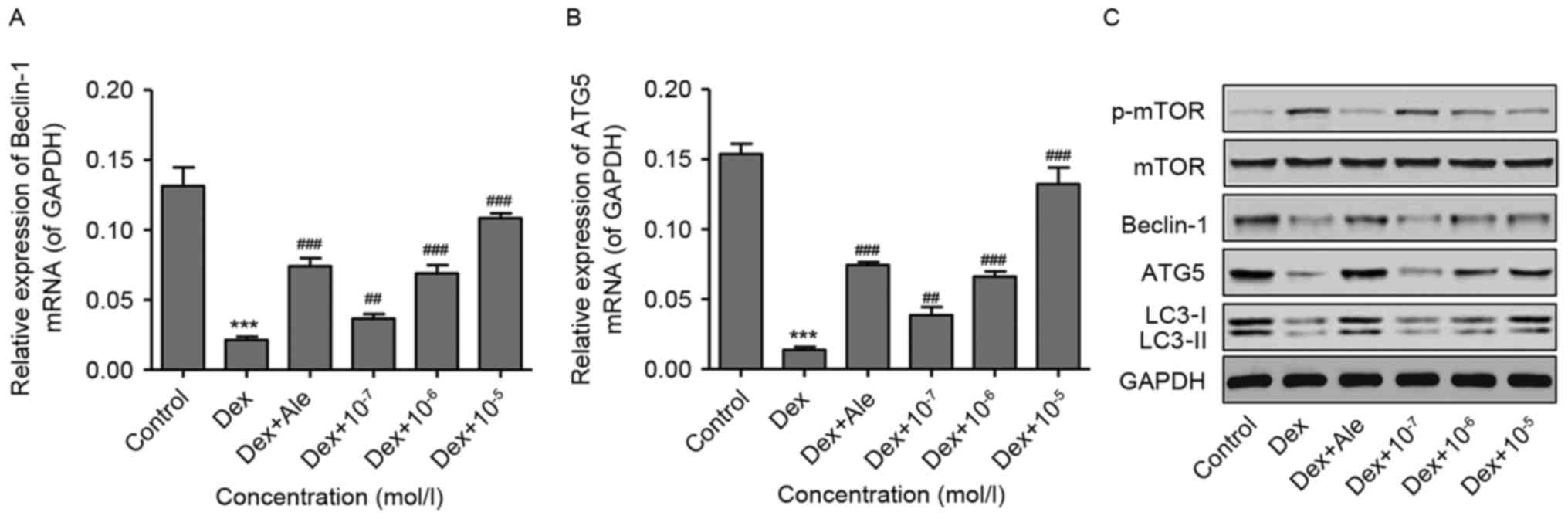
To investigate the effect of β-ecdysterone on
Dex-induced osteoblast autophagy, the osteoblasts were treated with
10−6 mol/l of Dex for 48 h, following which the
expression of autophagy-related genes were measured using RT-qPCR
and western blot analyses. As shown in Fig. 5A-C, 10−6 mol/l of Dex
significantly activated p-mTOR, and decreased the expression levels
of Beclin-1, ATG5 and LC3-II, compared with those in the control.
However, in the cells treated with β-ecdysterone at various
concentrations (10−7−10−5 mol/l), the changes
in the expression levels of p-mTOR, Beclin-1, ATG5 and LC3-II
induced by 10−6 mol/l of Dex were reversed in a
dose-dependent manner, which was similar to the results in the
osteoblasts treated with 10−5 mol/l of Ale. These
results suggested that β-ecdysterone reversed Dex-induced
osteoblast autophagy, possibly through inactivation of the TOR
signaling pathway.
Discussion
All diseases involving bone loss have a common
pattern, in that osteoclasts are specifically responsible for bone
resorption and, when osteoporosis occurs, osteoblast activity is
overcome by osteoclast activity. In the present study, it was
demonstrated that Dex inhibited the osteogenic differentiation,
proliferation and autophagy of osteoblasts in vitro, and
induced apoptosis. The protective effect of β-ecdysterone on
Dex-induced osteoporosis was also shown. These findings provided
the first evidence, to the best of our knowledge, that
β-ecdysterone isolated from traditional Chinese medicine can induce
osteogenic differentiation and autophagy, and this effect is
dependent on the inactivation of the mTOR signaling pathway.
During differentiation in vitro, osteoblast
phenotypic markers appear in the following order: Formation of type
I collagen matrix, expression of ALP and then Runx2, secretion of
OCN, and mineralization of bone nodules. Runx2 is an important
transcription factor, which controls several subsequent osteogenic
differentiation-related gene expression events, and mice with
Runx2-knockout have severe bone defects (18). OCN is another marker expressed
later during differentiation (19). RANKL is a membrane-bound tumor
necrosis factor receptor, which is expressed on osteoblast
precursor cells and recognize RANK on the osteoclast surface
through direct cell-cell interaction (20). The results of the present study
showed that Dex at higher concentrations inhibited osteoblast
differentiation through decreasing the activity of ALP, reducing
the expression of Runx2 and OCN, and increasing the expression of
RANKL, which was in accordance with the results of other studies
(21). However, higher
concentrations of Dex promoted osteoblast differentiation with
increased ALP activity and expression of Runx2 (22). These findings suggest that the
effects of Dex on impairing or favoring osteoblastic
differentiation appear to be dependent on its concentration and
experimental conditions. β-ecdysterone reversed the effect of Dex
on osteoblast differentiation and proliferation in a dose-dependent
manner, which was similar to previous findings that β-ecdysterone
stimulates the activity of ALP, and induce the expression of Runx2
and OCN (9).
Apoptosis and autophagy are two important programmed
mechanisms to control cell death and survival. Autophagy can either
delay the occurrence of apoptosis (23) or promote apoptosis (24). The balance between apoptosis and
autophagy is critical to the cellular homeostasis. In the present
study, it was found that Dex induced osteoblast apoptosis and
inhibited autophagy, with an increased Bax/Bcl-2 ratio and
decreased expression of autophagy-related markers, including
Beclin-1, ATG5 and LC3-II. In partial agreement with these
findings, there are other studies that a number of apoptotic cells
and increased autophagic activity are generated in response to Dex,
suggesting that autophagy is a self-protective response to GCs
(16,25). Previous studies have shown that the
overexpression of Beclin-1 in HeLa cells results in an increased
number of autophagosome (26), and
that Beclin-1 deficiency in mice reduces autolysosome production
(27). This indicated that the
expression of Beclin-1 accompanies autophagy. The deletion of ATG5
has been found to reduce the protein expression of LC3-II in
cortical bone of mice, indicating decreased autophagic activity
(28).
β-ecdysterone has previously been shown to mediate
the neuroprotective effect in SH-SY5Y cells through preventing
mitochondria-dependent apoptosis via inhibiting the activation of
p53, which was limited within concentrations of 1–10 µM (13). In agreement, the results of the
present study showed that β-ecdysterone at different concentrations
(10−7−10−5 mol/l) reduced osteoblastic
apoptosis in a dose-dependent manner. In addition, a sustained
state of cellular stress, including exposure to a high dose or
chronic GC treatment, can result in failure of the induction of
autophagy. Based on the results of the present study, it was
hypothesized that osteoblasts initially responded to the
Dex-induced stress by increasing the number of cells undergoing
autophagy, which was in accordance with a previous report that
β-ecdysterone prevented the GC-induced increase in autophagy of
BMSCs and whole bone (29).
It has been reported that mTOR is a negative
regulator of autophagy and that the inhibition of p-mTOR has been
used widely as an indicator of activation of the autophagy pathway
(30,31). AMPK controls the osteogenic
differentiation of human mesenchymal stem cells through early mTOR
inhibition-mediated autophagy and late activation of the Akt/mTOR
signaling axis (32). The results
of the present study showed that Dex dose-dependently increased the
expression of p-mTOR, which was reversed by β-ecdysterone
treatment. This suggested that β-ecdysteron may act upon the
autophagic pathway via an mTOR-dependent mechanism.
In conclusion, the present study demonstrated that
Dex inhibited proliferation, differentiation and autophagy, and
induced apoptosis of osteoblasts. β-ecdysterone significantly
reversed Dex-inducedapoptosis through inducing autophagy via mTOR
signaling. These results provide pre-clinical support for further
assessment of the ability of β-ecdysterone treatment to improve
skeletal fragility resulting from excess GC.
Acknowledgements
This study was supported by the Science and
Technology Project from Traditional Chinese Medicine of Zhejiang
Province (grant no. 2015ZA174) and the Social Development Major
Scientific and Technological Projects in Xiaoshan District of
Hangzhou City (grant no. 2014207).
References
|
1
|
Whittier X and Saag KG:
Glucocorticoid-induced Osteoporosis. Rheum Dis Clin North Am.
42:177–189. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Buehring B, Viswanathan R, Binkley N and
Busse W: Glucocorticoid-induced osteoporosis: An update on effects
and management. J Allergy Clin Immunol. 132:1019–1030. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fukushima W, Fujioka M, Kubo T, Tamakoshi
A, Nagai M and Hirota Y: Nationwide epidemiologic survey of
idiopathic osteonecrosis of the femoral head. Clin Orthop Relat
Res. 468:2715–2724. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Compston J: Management of
glucocorticoid-induced osteoporosis. Nat Rev Rheumatol. 6:82–88.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yun SI, Yoon HY, Jeong SY and Chung YS:
Glucocorticoid induces apoptosis of osteoblast cells through the
activation of glycogen synthase kinase 3beta. J Bone Miner Metab.
27:140–148. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li H, Qian W, Weng X, Wu Z, Li H, Zhuang
Q, Feng B and Bian Y: Glucocorticoid receptor and sequential P53
activation by dexamethasone mediates apoptosis and cell cycle
arrest of osteoblastic MC3T3-E1 cells. PLoS One. 7:e370302012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ohnaka K, Tanabe M, Kawate H, Nawata H and
Takayanagi R: Glucocorticoid suppresses the canonical Wnt signal in
cultured human osteoblasts. Biochem Biophys Res Commun.
329:177–181. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
O'Brien CA, Jia D, Plotkin LI, Bellido T,
Powers CC, Stewart SA, Manolagas SC and Weinstein RS:
Glucocorticoids act directly on osteoblasts and osteocytes to
induce their apoptosis and reduce bone formation and strength.
Endocrinology. 145:1835–1841. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gao L, Cai G and Shi X: Beta-ecdysterone
induces osteogenic differentiation in mouse mesenchymal stem cells
and relieves osteoporosis. Biol Pharm Bull. 31:2245–2249. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cahlíková L, Macáková K, Chlebek J,
Host'álková A, Kulhánková A and Opletal L: Ecdysterone and its
activity on some degenerative diseases. Nat Prod Commun. 6:707–718.
2011.PubMed/NCBI
|
|
11
|
Syrov VN, Khushbaktova ZA and Nabiev AN:
An experimental study of the hepatoprotective properties of
phytoecdysteroids and nerobol in carbon tetrachloride-induced liver
lesion. Eksp Klin Farmakol. 55:61–65. 1992.(In Russian). PubMed/NCBI
|
|
12
|
Zou Y, Wang R, Guo H and Dong M:
Phytoestrogen β-ecdysterone protects PC12 cells against
MPP+−induced neurotoxicity in vitro: Involvement of
PI3K-Nrf2-regulated pathway. Toxicol Sci. 147:28–38. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pan Z, Niu Y, Liang Y, Zhang X and Dong M:
β-Ecdysterone protects SH-SY5Y cells against
6-hydroxydopamine-induced apoptosis via mitochondria-dependent
mechanism: Involvement of p38MAPK-p53 signaling pathway. Neurotox
Res. 30:453–466. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang Z and Klionsky DJ: Eaten alive: A
history of macroautophagy. Nat Cell Biol. 12:814–822. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang YH, Chen K, Li B, Chen JW, Zheng XF,
Wang YR, Jiang SD and Jiang LS: Estradiol inhibits osteoblast
apoptosis via promotion of autophagy through the ER-ERK-mTOR
pathway. Apoptosis. 18:1363–1375. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xia X, Kar R, Gluhak-Heinrich J, Yao W,
Lane NE, Bonewald LF, Biswas SK, Lo WK and Jiang JX:
Glucocorticoid-induced autophagy in osteocytes. J Bone Miner Res.
25:2479–2488. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Komori T: Regulation of osteoblast
differentiation by Runx2. Adv Exp Med Biol. 658:43–49. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Choo MK, Yeo H and Zayzafoon M: NFATc1
mediates HDAC-dependent transcriptional repression of osteocalcin
expression during osteoblast differentiation. Bone. 45:579–589.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Boyce BF and Xing L: Functions of
RANKL/RANK/OPG in bone modeling and remodeling. Arch Biochem
Biophys. 473:139–146. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhu FB, Wang JY, Zhang YL, Quan RF, Yue
ZS, Zeng LR, Zheng WJ, Hou Q, Yan SG and Hu YG: Curculigoside
regulates proliferation, differentiation, and pro-inflammatory
cytokines levels in dexamethasone-induced rat calvarial
osteoblasts. Int J Clin Exp Med. 8:12337–12346. 2015.PubMed/NCBI
|
|
22
|
Hamidouche Z, Haÿ E, Vaudin P, Charbord P,
Schüle R, Marie PJ and Fromigué O: FHL2 mediates
dexamethasone-induced mesenchymal cell differentiation into
osteoblasts by activating Wnt/beta-catenin signaling-dependent
Runx2 expression. FASEB J. 22:3813–3822. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Degenhardt K, Mathew R, Beaudoin B, Bray
K, Anderson D, Chen G, Mukherjee C, Shi Y, Gélinas C, Fan Y, et al:
Autophagy promotes tumor cell survival and restricts necrosis,
inflammation, and tumorigenesis. Cancer Cell. 10:51–64. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Canu N, Tufi R, Serafino AL, Amadoro G,
Ciotti MT and Calissano P: Role of the autophagic-lysosomal system
on low potassium-induced apoptosis in cultured cerebellar granule
cells. J Neurochem. 92:1228–1242. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Conradie MM, de Wet H, Kotze DD, Burrin
JM, Hough FS and Hulley PA: Vanadate prevents
glucocorticoid-induced apoptosis of osteoblasts in vitro and
osteocytes in vivo. J Endocrinol. 195:229–240. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Luo S and Rubinsztein DC: Apoptosis blocks
Beclin 1-dependent autophagosome synthesis: An effect rescued by
Bcl-xL. Cell Death Differ. 17:268–277. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Qu X, Yu J, Bhagat G, Hibshoosh H, Troxel
A, Rosen J, Eskelinen EL, Mizushima N, Ohsumi Y, Cattoretti G and
Levine B: Promotion of tumorigenesis by heterozygous disruption of
the beclin 1 autophagy gene. J Clin Invest. 112:1809–1820. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nollet M, Santucci-Darmanin S, Breuil V,
Al-Sahlanee R, Cros C, Topi M, Momier D, Samson M, Pagnotta S,
Cailleteau L, et al: Autophagy in osteoblasts is involved in
mineralization and bone homeostasis. Autophagy. 10:1965–1977. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dai W, Jiang L, Lay YA, Chen H, Jin G,
Zhang H, Kot A, Ritchie RO, Lane NE and Yao W: Prevention of
glucocorticoid induced bone changes with beta-ecdysone. Bone.
74:48–57. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wander SA, Hennessy BT and Slingerland JM:
Next-generation mTOR inhibitors in clinical oncology: How pathway
complexity informs therapeutic strategy. J Clin Invest.
121:1231–1241. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fiorini C, Menegazzi M, Padroni C, Dando
I, Dalla Pozza E, Gregorelli A, Costanzo C, Palmieri M and
Donadelli M: Autophagy induced by p53-reactivating molecules
protects pancreatic cancer cells from apoptosis. Apoptosis.
18:337–346. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pantovic A, Krstic A, Janjetovic K, Kocic
J, Harhaji-Trajkovic L, Bugarskib D and Trajkovica V: Coordinated
time-dependent modulation of AMPK/Akt/mTOR signaling and autophagy
controls osteogenic differentiation of human mesenchymal stem
cellsc. Bone. 52:524–531. 2013. View Article : Google Scholar : PubMed/NCBI
|