Introduction
Osteoporosis is a silent aging process of skeletal
tissues, characterized by reductions in bone mass and the
microarchitectural deterioration of bone tissue, that results in
enhanced bone fragility and, consequently, in increased risk of
fractures (1,2). As the elderly population increases,
the number of patients with osteoporosis is on the rise, and the
prognosis of osteoporosis treatment using pharmacological agents is
frequently affected by the poor bone quality of the elderly
(3). Screening for osteoporosis is
based on the assessment of bone mineral density (BMD) (4); however, the decrease in skeletal
mechanical strength is not only caused by decreases in BMD, but
also by alterations in the bone microstructure, which may be
evaluated using non-invasive micro-computed tomography (CT)
(5,6).
To the best of our knowledge, a number of methods
have been employed to establish reliable osteoporosis models
(7). Glucocorticoid-induced
osteoporosis is the most common form of secondary osteoporosis,
which results in reductions in BMD and an increase in fracture risk
(8,9). Treatment with glucocorticoids leads
to a decrease in the generation of osteoblasts and osteocytes,
accompanied by an elongation in the lifespan of osteoclasts
(10,11), which in turn increases bone
resorption and inhibits the formation of new bone matrix. In the
present study, the prednisolone (PRED)-induced osteoporosis model
was used to study osteoporosis. Increased apoptosis and decreased
autophagy of osteoblasts has been observed following treatment with
a high dose of glucocorticoids in mice (12,13).
A number of mechanisms have been suggested to be involved in
glucocorticoid-induced apoptosis, including the upregulation of the
proapoptotic factors apoptosis regulator BCL2 (Bcl-2)-like protein
11 and Bcl-2 homologous antagonist/killer, and the downregulation
of the pro-survival factor Bcl-xL (14). Notably, the dosage of
glucocorticoid treatment determines the fate of osteocytes: A low
dose of glucocorticoids induces autophagy, whereas a high dose
induces apoptosis (15).
β-Ecdysterone (β-Ecd) is a naturally-occurring
estrogen analog derived from Achyranthes bidentata Blumeand
Cyanotis arachnoidea C.B. Clarke, and has been reported to
stimulate protein synthesis, to promote carbohydrate and lipid
metabolism, to alleviate hyperglycemia and hyperlipidemia, to
modulate immune reactions, and to protect endothelial cells from
apoptosis and induce their proliferation (16–18).
β-Ecd has been revealed to attenuate the
1-methyl-4-phenylpyridinium-induced apoptosis of PC12 cells
(19) and to protect SH-SY5Y cells
against 6-hydroxydopamine-induced apoptosis, by interfering with
p38 mitogen-activated protein kinase-p53 signaling (20). Treatment with β-Ecd additionally
prevented the glucocorticoid-induced reduction in bone formation
rate, the decrease in trabecular bone volume and cortical bone loss
in mice in vivo (9).
The present study aimed to investigate whether β-Ecd
was able to rescue the PRED-induced suppression of bone formation
in rats in vivo. The present results suggested that
treatment with β-Ecd may inhibit bone loss and prevent the
deterioration of bone mechanical properties associated with PRED
use, possibly through the maintenance of bone formation. In
addition, autophagy was examined following treatment with PRED or
β-Ecd, and the present findings suggested that autophagy may be
among the mechanisms underlying the bone-anabolic effects of
β-Ecd.
Materials and methods
Induction of osteoporosis and β-Ecd
treatment
All animal care and experimental procedures in the
present study complied with the protocols approved by the
Institutional Animal Care and Use Committee of Xiao Shan TCM
Hospital (Xiaoshan, China). A total of 24 specific pathogen-free
Sprague-Dawley male rats (age, 4 weeks; weight, 180–220 g) were
purchased from Shanghai SLAC Laboratory Animal Co., Ltd. (Shanghai,
China) and housed in an animal facility at 25°C, with a relative
humidity of 60–70%, under a 12-h light/dark cycle with free access
to food and water. The rats were randomly divided into the 4
following groups (8 rats/groups): i) Control group (untreated
rats); ii) PRED group, where a pellet of PRED (2.5 mg/pellet;
Zhengzhou Lingrui Pharmaceutical Co., Ltd., Xinzheng, China) was
subcutaneously implanted into the back of the rats every day for 4
weeks; iii) PRED + β-Ecd group, where rats received PRED
subcutaneously similar to the PRED group, and were additionally
subcutaneously injected with β-Ecd (5 or 10 mg/kg body weight;
Shanghai Tauto Biotech Co., Ltd., Shanghai, China) 5 times for 4
weeks; and iv) PRED + alendronate (ALN) group, where rats received
PRED subcutaneously similar to the PRED group, and were also
subcutaneously injected with ALN (3 mg/kg body weight; Merck &
Co., Inc., Whitehouse Station, NJ, USA) 5 times for 4 weeks.
Structural analysis using micro-CT and
measurement of BMD
The fifth lumbar vertebrae (L5) were isolated
following 4 weeks of treatment as previously described (21), fixed in 1% formalin overnight at
room temperature and in 70% ethanol for 2 h at room temperature.
The micro-CT apparatus and the SCANCO image processing language
(version 5.08b) software used in the present study were obtained
from Scanco Medical AG (Brüttisellen, Switzerland). The micro-CT
contained a micro X-ray source (current, 85 mA; potential, 70 kV)
directed toward the samples, according to the manufacturer's
instructions. The BMD (g/cm3) of the L5 was measured
using dual-energy C-ray absorptiometry with a DCS-600 Aloka bone
densitometer (Hitachi, Ltd., Tokyo, Japan) using the small-animal
scan mode.
Immunofluorescence
Paraffin-embedded lumbar vertebrae (L1-2) sections
(4–7 µm) from rats (4 rats/group) were fixed in 10% formalin
overnight at room temperature, separately dehydrated by 50, 70, 85,
95 and 100% ethanol for 2 h, deparaffinized by dimethylbenzene for
15 min and separately hydrated by 100, 95, 85 and 75% ethanol for 5
min at room temperature for histological assessment. For antigen
retrieval, the tissue sections were incubated in sodium citrate
buffer (JRDUN Biotechnology, Co., Ltd., Shanghai, China) at 37°C
for 15 min and heated in a microwave oven at 92–98°C for 10–30 min.
Following antigen retrieval, the tissue sections were permeabilized
with 0.1% Triton X-100 in phosphate-buffered saline (PBS) for 20
min at room temperature. Following blocking with 2% bovine serum
albumin (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) in PBS for
1 h at room temperature, the tissue sections were incubated with
anti-Beclin-1 (1:200, cat. no. ab62557; Abcam, Cambridge, MA, USA)
and anti-microtubule-associated protein 1A/1B-light chain (LC)
3I/II (1:200, cat. no. ab128025; Abcam) antibodies at 4°C
overnight, prior to incubation with the corresponding fluorescein
isothiocyanate-labeled goat anti-rabbit immunoglobulin G (IgG)
(H+L) secondary antibodies (1:500, cat. no. A0562; Beyotime
Institute of Biotechnology, Haimen, China) for 1 h at room
temperature. The nuclei were then stained with 1 µg/ml
4′,6-diamidino-2-phenylindole for 5 min at room temperature.
Stained sections were observed under a CX41RF fluorescence
microscope (Olympus Corporation, Tokyo, Japan) at ×200
magnification.
Terminal deoxynucleotidyl transferase
deoxyuridine triphosphate nick-end labeling (TUNEL) staining
TUNEL staining of the fifth lumbar vertebrae from
rats (4 rats/group) was performed using the In Situ Cell
Death Detection kit, POD (cat. no. 11684817910; Sigma-Aldrich;
Merck KGaA), according to the manufacturer's instructions. Each
section (4–7 µm) was fixed in 10% formalin overnight and separately
dehydrated by 50, 70, 85, 95 and 100% ethanol for 2 h at room
temperature. Sections were incubated with TUNEL reagent for 1 h at
37°C and stained with DAB for 10 min and hematoxylin for 5 min at
room temperature. Sections were visualized under a fluorescence
microscope (CX41RF; Olympus Corporation, Tokyo, Japan) and the
numbers of TUNEL-positive cells were counted at ×200 magnification
in 30 fields of view/section.
Biochemical analysis
Serum was isolated from rats (4 rats/group) after 4
weeks treatment. Briefly, 10 ml of blood was collected in a serum
separator tube and processed within 1 h. Separation of the serum
was accomplished by centrifugation at 800 × g for 10 min at room
temperature. The serum concentration of calcium and phosphorus, and
the activity of tartrate-resistant acid phosphatase (TRAP) and
alkaline phosphatase (ALP) were measured using a Hitachi 7070
analyzer (Hitachi, Ltd.).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from tibia tissues isolated
and crushed from rats (4 rats/group) after 4 weeks treatment as
previously described (22), using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), according to the manufacturer's instructions. A
total of 1 µg RNA was reverse transcribed to cDNA using a cDNA
synthesis kit (Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. qPCR was performed using SYBR-Green
(Takara Biotechnology Co., Ltd., Dalian, China) on a StepOne
Real-Time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.), with GAPDH as an internal control. The following
thermocycling conditions were used for the PCR: 95°C for 10 min,
followed by 40 cycles of 95°C for 15 sec and 60°C for 45 sec, and a
final extension step of 95°C for 15 sec, 60°C for 1 min, 95°C for
15 sec and 60°C for 15 sec. The primers used in the present study
were as follows: Bone morphogenetic protein 2 (BMP2) forward,
5′-CTGTCCCTACTGATGAGTTTC-3′ and reverse,
5′-CTAACCTGGTGTCCAATAGTC-3′; Beclin-1 forward,
5′-GAGTTGCCGTTGTACTGTTC-3′ and reverse, 5′-TGCCTCCAGTGTCTTCAATC-3′;
Runt-related transcription factor (RUNX) 2 forward,
5′-ACTTCGTCAGCGTCCTATC-3′ and reverse, 5′-CATCAGCGTCAACACCATC-3′;
autophagy protein (ATG) 5 forward, 5′-TGGCTGAGCGAGCATCTGAG-3′ and
reverse, 5′-TGACTGCGGGTGGTTCCATC-3′; receptor activator of nuclear
factor-κB ligand (RANKL) forward, 5′-CACAGCGCTTCTCAGGAGTT-3′ and
reverse, 5′-GATGGTGAGGTGAGCAAACG-3′; and GAPDH forward,
5′-CACCCACTCCTCCACCTTTG-3′ and reverse, 5′-CCACCACCCTGTTGCTGTAG-3′.
Relative gene expression was calculated according to the
comparative Cq method (23) and
the fold-change of target gene expression was normalized to the
internal control GAPDH.
Western blot analysis
The tibia tissue samples obtained from rats (4
rats/group) after 4 weeks of treatment were homogenized in ice-cold
lysis buffer [150 mM NaCl, 0.5% Triton X-100, 50 mM Tris-HCl (pH
7.4), 20 mM EGTA, 1 mM DTT, 1 mM Na3VO4 and
protease inhibitor cocktail tablet] and total cell lysates were
prepared using radioimmunoprecipitation assay lysis buffer
supplemented with protease inhibitor (Beyotime Institute of
Biotechnology). The total protein concentration in each sample was
measured using a Lowry protein assay kit (Bio-Rad Laboratories,
Inc., Hercules, CA, USA). Equal amounts of extracted protein
samples (50 µg) were separated by SDS-PAGE on a 10% gel and
transferred onto polyvinylidene difluoride membranes (Roche
Diagnostics GmbH, Mannheim, Germany). Membranes were blocked in
fat-free milk overnight at 4°C and incubated with primary
antibodies, anti-Runx2 (1:1,000; cat. no. ab76956), anti-RANKL
(1:1,000; cat. no. ab45039), anti-Beclin-1 (1:600; cat. no.
ab55878), anti-ATG5 (1:5,000; cat. no. ab108327), anti-BMP2 (1:800;
cat. no. ab14933), and anti-caspase-3 (1:500; cat. no. ab44976)
(all from Abcam), anti-Bcl-2 (1:400, cat. no. Sc-492; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), anti-LC3 (1:1,000; cat. no.
2775s), and anti-GAPDH (1:1,500; cat. no. 5174) (both from Cell
Signaling Technology, Inc., Danvers, MA, USA), for 2 h at 25°C. The
membranes were subsequently incubated for 1 h at 37°C with
horseradish peroxidase-conjugated IgG secondary antibodies
(1:1,000, cat. nos. A0208, A0181, A0216; Beyotime Institute of
Biotechnology). Protein bands were visualized using Western
Lightning Plus Enhanced Chemiluminescence reagent (PerkinElmer,
Inc., Waltham, MA, USA), and blots were semi-quantified by
densitometry using Quantity One software version 4.62 (Bio-Rad
Laboratories, Inc.).
Statistical analysis
Data are expressed as the mean ± standard deviation
of at least three independent replicates and were analyzed using
unpaired, two-tailed Student's t-test, and one-way analysis of
variance followed by Tukey's post hoc test. Statistical analysis
was performed using GraphPad Prism software version 5.0 (GraphPad
Software, Inc., La Jolla, CA, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
Effects of β-Ecd on PRED-induced
alterations in bone microarchitecture
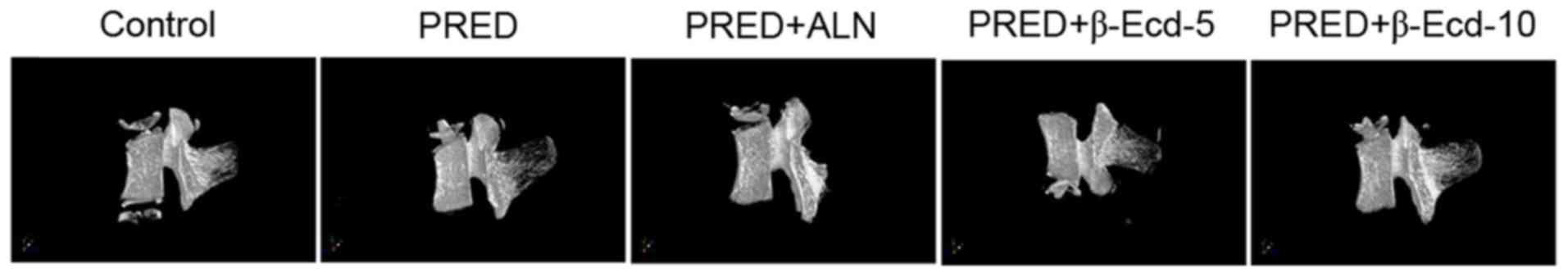
Bone loss was observed in all PRED-treated rats, as
detected using micro-CT analysis (Fig.
1 and Table I). In the PRED
group, the BMD values of L5 were significantly decreased compared
with in the control group. Micro-CT evaluation revealed that the
2-D model bone surface density (BS/TV) of the trabecular bones was
significantly decreased in the PRED group compared with in control
rats (29.1% decrease; Table I). 3D
model analysis demonstrated that the trabecular number (Tb.N) was
significantly lower following PRED treatment compared with in
control rats (27.3% decrease; Table
I). In addition, a significantly increased trabecular plate
separation (Tb.Sp) was revealed in the PRED treatment group
compared with in the control group (59.8% increase; Table I). The trabecular bone tissue
volume density (BV/TV) was revealed to be 30.4% lower following
treatment with PRED. However, no significant difference was
detected in the trabecular thickness (Tb.Th) and structure model
index (SMI) between PRED-treated and control rats. Notably,
treatment with 10 mg/kg β-Ecd significantly increased the levels of
BMD, BS/TV, BV/TV and Tb.N, whereas it decreased Tb.Sp and SMI
compared with the PRED group; treatment with 3 mg/kg ALN produced
similar results (Table I).
Additionally, treatment with 5 mg/kg β-Ecd significantly increased
the levels of BMD, BS/TV, BV/TV and Tb.N, whereas it decreased
Tb.Th and Tb.Sp levels compared with the PRED group (Table I).
 | Table I.Alterations in BMD and
microarchitectural parameters of the fifth lumbar vertebra of rats
in the various treatment groups. |
Table I.
Alterations in BMD and
microarchitectural parameters of the fifth lumbar vertebra of rats
in the various treatment groups.
|
| Group |
|---|
|
|
|
|---|
| Parameters | Control | PRED | PRED + ALN | PRED + β-Ecd, 5
mg/kg | PRED + β-Ecd, 10
mg/kg |
|---|
| BMD,
g/cm3 |
0.299±0.007 |
0.150±0.031a |
0.259±0.015c |
0.229±0.006b |
0.263±0.006c |
| BS/TV,
mm−1 |
3.78±0.212 |
2.68±0.213a |
3.81±0.139c |
3.74±0.238c |
3.74±0.046c |
| BV/TV, % |
35.68±0.379 |
24.83±1.627a |
32.07±0.558c |
30.49±0.761c |
32.46±0.001c |
| Tb.N,
mm−1 |
1.313±0.049 |
0.954±0.079a |
1.275±0.058c |
1.250±0.064c |
1.276±0.009c |
| Tb.Th, mm |
0.272±0.007 |
0.260±0.005 |
0.252±0.007 |
0.244±0.006b |
0.254±0.002 |
| Tb.Sp, mm |
1.037±0.048 |
1.657±0.155a |
1.067±0.011c |
1.122±0.071c |
1.061±0.030c |
| Structure model
index |
−3.90±0.345 |
−4.32±0.344 |
−3.21±0.261b |
−3.36±0.543 |
−3.48±0.193c |
Effects of β-Ecd on PRED-induced
alterations in biochemical indices
In the present study, treatment with PRED was
demonstrated to significantly increase the serum levels of calcium
and phosphorus by 69.2 and 58.2%, respectively, and enhance the
serum activity of ALP and TRAP by 52.8 and 12.9%, respectively,
compared with the control group (Fig.
2). However, treatment with ALN or 10 mg/kg β-Ecd significantly
decreased the serum calcium and phosphorus levels and suppressed
the serum activity of ALP and TRAP. Conversely, treatment with 5
mg/kg β-Ecd was revealed to attenuate the PRED-induced increase in
ALP and TRAP levels; however, it had no effect on the serum levels
of calcium and phosphorus.
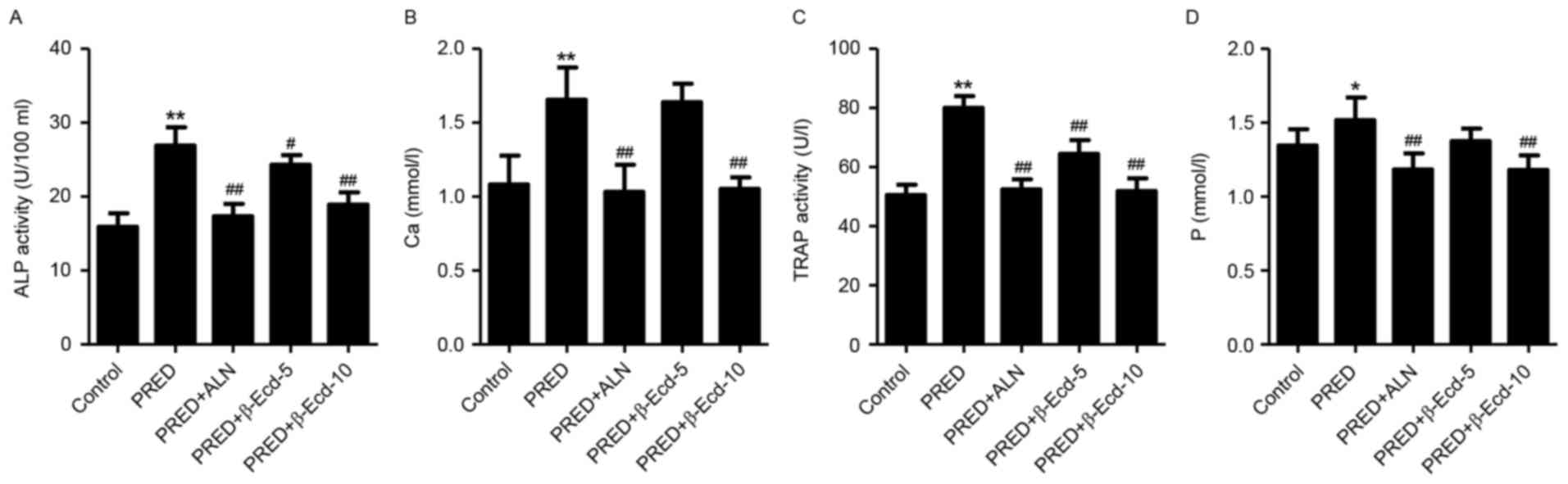 | Figure 2.Biochemical parameters in rats from
various experimental groups. The serum activity or expression of
(A) ALP, (B) Ca, (C) TRAP and (D) P were detected using a
biochemical analyzer. Control, untreated rats; PRED, rats received
PRED for 4 weeks; PRED + ALN, rats received PRED and ALN for 4
weeks; PRED + β-Ecd, rats received PRED and β-Ecd (5 or 10 mg/kg)
for 4 weeks. Data are expressed as the mean ± standard deviation.
*P<0.05, **P<0.01 vs. control; #P<0.05,
##P<0.01 vs. PRED. ALP, alkaline phosphatase; TRAP,
tartrate-resistant acid phosphatase; Ca, calcium; P, phosphorus;
PRED, prednisolone; ALN, alendronate; β-Ecd, β-ecdysterone. |
Effects of β-Ecd on
PRED-inducedalterations inautophagy and apoptosis
As presented in Fig.
3, immunofluorescence staining demonstrated that treatment with
PRED markedly downregulated the expression of Beclin-1 and LC3I/II
compared with the control group. However, ALN and β-Ecd
administration appeared to inhibit the PRED-induced decreases in
Beclin-1 and LC3I/II expression. In addition, TUNEL-positive cells
in PRED-treated rats were markedly increased compared with control
rats, whereas treatment with ALN and β-Ecd was revealed to markedly
reduce the number of apoptotic cells in PRED-treated rats (Fig. 3). These findings suggested that
β-Ecd may inhibit the PRED-induced alterations in autophagy and
apoptosis in osteoporotic rats.
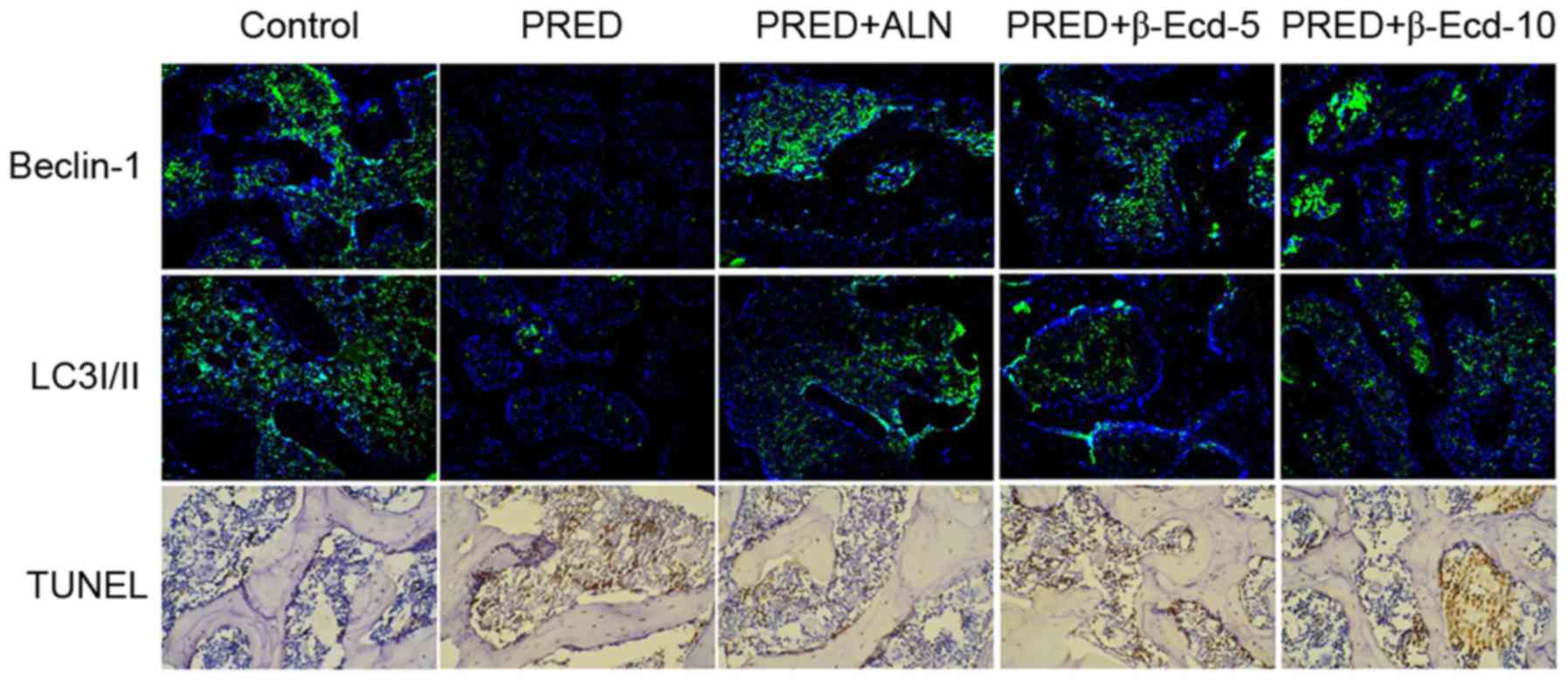 | Figure 3.Immunofluorescence and TUNEL staining
in rats from various experimental groups. The protein expression of
Beclin-1 and LC3I/II was detected using immunofluorescence
staining. Apoptosis was assessed using TUNEL staining.
Magnification, ×200. Control, untreated rats; PRED, rats received
PRED for 4 weeks; PRED + ALN, rats received PRED and ALN for 4
weeks; PRED + β-Ecd, rats received PRED and β-Ecd (5 or 10 mg/kg)
for 4 weeks. TUNEL, terminal deoxynucleotidyl transferase
deoxyuridine triphosphate nick-end labeling; LC3I/II,
microtubule-associated protein 1A/1B-light chain 3I/II; PRED,
prednisolone; ALN, alendronate; β-Ecd, β-ecdysterone. |
Effects of β-Ecd on PRED-induced
alterations in RUNX2, RANKL and BMP2 expression
In order to investigate the effects of PRED and
β-Ecd on osteoblast differentiation, the expression of
differentiation-associated markers, including BMP2, RANKL and
RUNX2, was assessed. Treatment with PRED was revealed to
significantly suppress the mRNA expression of RUNX2 and BMP2,
whereas it upregulated RANKL mRNA expression (Fig. 4A-C). Notably, treatment with ALN or
β-Ecd significantly prevented the PRED-induced downregulation in
RUNX2 and BMP2, and the upregulation in RANKL mRNA expression in
osteoporotic rats (Fig. 4A-C).
Similar results regarding protein expression were obtained
following western blot analysis (Fig.
4D).
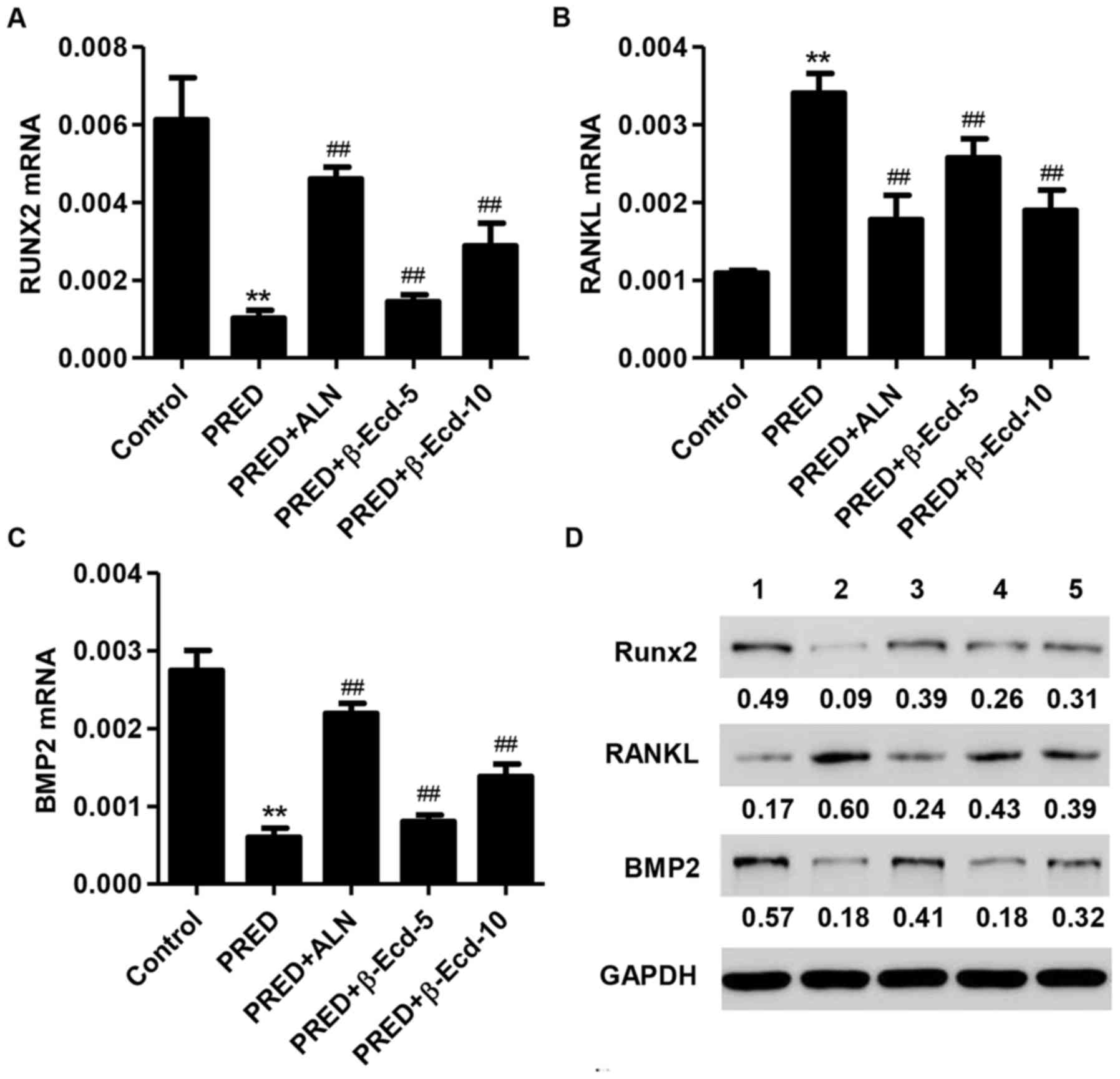 | Figure 4.Expression of RUNX2, RANKL and BMP2 in
rats from the various experimental groups. The mRNA expression
levels of (A) RUNX2, (B) RANKL and (C) BMP2 were measured using the
reverse transcription-quantitative polymerase chain reaction. (D)
The protein expression levels of RUNX2, RANKL and BMP2 were
detected using western blot analysis. Control, untreated rats;
PRED, rats received PRED for 4 weeks; PRED + ALN, rats received
PRED and ALN for 4 weeks; PRED + β-Ecd, rats received PRED and
β-Ecd (5 or 10 mg/kg) for 4 weeks. Data are expressed as the mean ±
standard deviation. **P<0.01 vs. control; ##P<0.01
vs. PRED. RUNX, Runt-related transcription factor; RANKL, receptor
activator of nuclear factor-κB ligand; BMP, bone morphogenetic
protein; PRED, prednisolone; ALN, alendronate; β-Ecd,
β-ecdysterone. |
Effects of β-Ecd on PRED-induced
alterations in Beclin-1, ATG5, LC3I/II, Bcl-2 and caspase-3
expression
The balance between autophagy and apoptosis is
important for the maintenance of cellular homeostasis (24). ATG5 is required for LC3-I
conjugation and LC3-II formation, and it is regarded as a key
marker of autophagosome formation (25), while Beclin-1 has been reported to
serve a role in autophagy (26).
The present results demonstrated that PRED administration
significantly downregulated the mRNA expression of Beclin-1 and
ATG5 in rats, whereas treatment with ALN or β-Ecd significantly
inhibited the PRED-induced decreases in the mRNA expression levels
of Beclin-1 and ATG5 (Fig. 5A and
B). In addition, western blot analysis revealed that PRED
administration markedly suppressed the protein expression of
Beclin-1, ATG5, LC3I/II and Bcl-2, whereas it upregulated the
expression of caspase-3 (Fig. 5C).
Notably, treatment with ALN or β-Ecd appeared to counteract the
PRED-induced alterations in the expression of Beclin-1, ATG5,
LC3I/II, Bcl-2 and caspase-3 in vivo (Fig. 5C).
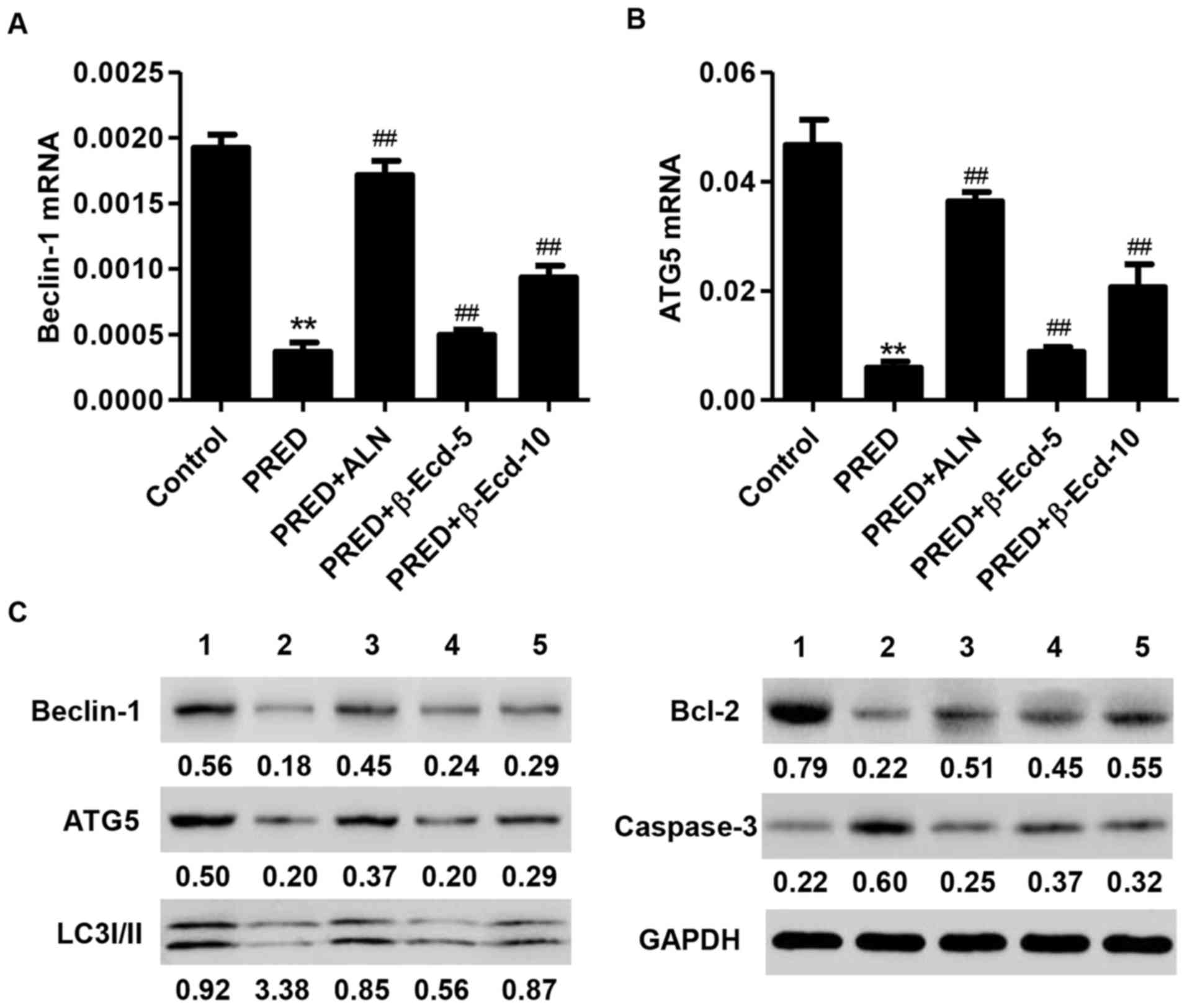 | Figure 5.Expression of Beclin-1, ATG5, LC3I/II,
Bcl-2 and caspase-3 in rats from various experimental groups. The
mRNA expression levels of (A) Beclin-1 and (B) ATG5 were measured
using reverse transcription-quantitative polymerase chain reaction.
(C) The protein expression levels of Beclin-1, ATG5, LC3I/II, Bcl-2
and caspase-3 were detected using western blot analysis. Control,
untreated rats; PRED, rats received PRED for 4 weeks; PRED + ALN,
rats received PRED and ALN for 4 weeks; PRED + β-Ecd, rats received
PRED and β-Ecd (5 or 10 mg/kg) for 4 weeks. Data are expressed as
the mean ± standard deviation. **P<0.01 vs. control;
##P<0.01 vs. PRED. ATG, autophagy protein; LC3I/II,
microtubule-associated protein 1A/1B-light chain 3I/II; Bcl, B-cell
lymphoma; PRED, prednisolone; ALN, alendronate; β-Ecd,
β-ecdysterone. |
Discussion
In the present study, the decreases in BMD and the
deterioration of trabecular microarchitecture were demonstrated to
reflect bone fragility and bone mass loss in PRED-treated rats. In
addition, the alterations in biochemical parameters, including ALP
and TRAP activity, calcium and phosphorus levels, and in the
expression of autophagy and apoptosis-associated factors, suggested
that bone absorption was enhanced, whereas the activity of
osteoblastic cells and the formation of new bone were suppressed in
PRED-treated rats.
The PRED-induced osteoporosis model has been used in
a number of previous studies: Huang et al (4) reported a significant decrease in BMD
and in the serum levels of calcium, phosphorus and osteocalcin in
PRED-induced osteoporotic rats. PRED treatment has additionally
been reported to decrease cortical bone mineral contents, cortical
thickness, the stress/strain index and mandibular volume, whereas
it did not produce marked alterations in trabecular structural
parameters (27). Similarly, the
findings of the present study demonstrated that PRED induced a
significant decrease in BMD, in the serum levels of calcium and
phosphorus, and in the serum ALP and TRAP activity in rats, thus
indicating high rates of bone turnover following treatment with
PRED. To the best of our knowledge, this is the first time that a
decrease in volumetric BMD was revealed to be accompanied by the
deterioration of trabecular microarchitecture, as measured by
micro-CT in rats in vivo. Previous studies have reported
that the maintenance of trabecular structure is a critical factor
for the strength of the lumbar vertebra (28,29).
In accordance with the present results, previous reports have
demonstrated that the BMD was correlated with bone
microarchitectural parameters (30,31).
In the present study, treatment with PRED was revealed to decrease
BS/TV, BV/TV and Tb.N, whereas it increased Tb.Sp values in the
trabecular bones of rats in vivo; Tb.Th and SMI values
remained unaltered in PRED-treated rats. The present findings
suggested that BS/TV, BV/TV, Tb.N and Tb.Sp may have potential as
sensitive biomarkers for the early detection of alterations in
trabecular structure, indicative of osteoporosis development.
A previous study used a combination of linkage and
association analysis, and linked BMP2 expression to a phenotype of
low BMD combined with a high fracture risk (32). BMP2 is produced and secreted by
osteoblasts, and has been reported to exert independent effects on
the regulation of osteoblast proliferation and mineralization
(33). Osteoblasts are involved in
bone formation and, through the production of RANKL, are able to
modulate the formation and differentiation of osteoclasts; RANKL
provides a signal to osteoclast progenitors through RANK, leading
to the activation of osteoclast differentiation and function
(34). Overexpression of RUNX2 in
transgenic mice resulted in an osteoporotic phenotype, thus
suggesting that RUNX2 may be involved in genetic processes
associated with osteoporosis (35). In the present study, PRED treatment
was revealed to suppress the expression of RUNX2 and BMP2, whereas
it potentiated the expression of RANKL. In accordance with the
present findings, previous studies have reported decreased BMP2 and
increased RANKL expression in glucocorticoid-induced osteoporosis
models in vivo (36) and
in vitro (37).
A number of mechanisms have been suggested to
underlie glucocorticoid-induced osteoblast apoptosis: PRED
administration (2.1 mg/kg) has been reported to enhance osteoblast
apoptosis in mouse vertebrae, and increase the frequency of
osteocyte apoptosis in metaphysical cortical bone, thus resulting
in a decrease in vertebral cancellous bone due to reduced bone
formation (38). In accordance
with this previous study, the present results demonstrated that
PRED induced cell apoptosis, via suppressing the expression of the
antiapoptotic factor Bcl-2 and promoting the activation of the
proapoptotic caspase-3. In addition, PRED administration was
revealed to inhibit the expression of autophagosome-regulatory
proteins, including Beclin-1, ATG5 and LC3I/II in tibia tissue
samples of rats (39); in
agreement with the present results, transfection of osteoblasts
with Beclin-1-targeting small interfering-RNA effectively inhibited
autophagosome formation in vitro (39). However, a study by Piemontese et
al (40) reported
contradictory results, as PRED was demonstrated to increase the
autophagic flux in osteocyte-enriched mouse bones, as measured
using LC3 conversion. The variations in the experimental animals
and drug dosages that were used may explain the contradictory
findings in the study by Piemontese et al (40). A previous study revealed that
dexamethasone, in contrast to PRED, induced osteocyte apoptosis,
resulted in an increase in autophagy markers and promoted the
accumulation of autophagosome vacuoles in vitro and in
vivo, thus promoting the onset of osteocyte autophagy (10).
The present results suggested that β-Ecd may inhibit
PRED-induced bone loss and apoptosis, and enhance autophagy in rats
in vivo. These effects were similar to ALN, which is a
bisphosphonate used in the treatment of osteoporosis as it inhibits
bone resorption by interfering with the activity of osteoclasts;
ALN was used as a positive control in the present study (41). In accordance with the present
results, a previous study reported that β-Ecd induced osteogenic
differentiation in mouse mesenchymal stem cells and attenuated the
development of osteoporosis through an endoplasmic
reticulum-associated signaling pathway (16). Additionally, β-Ecd has been
demonstrated to prevent glucocorticoid-induced alterations in bone
formation, bone cell viability and bone mass, and to prevent the
glucocorticoid-induced increase in autophagy in bone marrow stromal
cells and whole bone tissue (9).
These data generated from an in vitro system may not be
necessarily and comprehensively consistent with the effects of
β-Ecd on autophagy in vivo.
In conclusion, the results of the present study
suggested that β-Ecd may prevent PRED-induced osteoporosis in rats
in vivo through the inhibition of bone loss and apoptosis
and through the induction of autophagy. Therefore, β-Ecd may in the
future be used for the development of treatment strategies for
osteoporosis in humans.
Acknowledgements
The present study was supported by the Science and
Technology Project from Traditional Chinese Medicine of Zhejiang
Province (grant no. 2015ZA174), and the Social Development Major
Scientific and Technological Projects in Xiaoshan District of
Hangzhou City (grant no. 2014207).
References
|
1
|
Emkey GR and Epstein S: Secondary
osteoporosis: Pathophysiology & diagnosis. Best Pract Res Clin
Endocrinol Metab. 28:911–935. 2014. View Article : Google Scholar
|
|
2
|
Shiraishi A, Higashi S, Masaki T, Saito M,
Ito M, Ikeda S and Nakamura T: A comparison of alfacalcidol and
menatetrenone for the treatment of bone loss in an ovariectomized
rat model of osteoporosis. Calcif Tissue Int. 71:69–79. 2002.
View Article : Google Scholar
|
|
3
|
Eriksson F, Mattsson P and Larsson S: The
effect of augmentation with resorbable or conventional bone cement
on the holding strength for femoral neck fracture devices. J Orthop
Trauma. 16:302–310. 2002. View Article : Google Scholar
|
|
4
|
Huang Y, Bo Y, Wu X, Wang Q, Qin F, Zhao L
and Xiong Z: An intergated serum and urinary metabonomic research
based on UPLC-MS and therapeutic effects of Gushudan on
prednisolone-induced osteoporosis rats. J Chromatogr B Analyt
Technol Biomed Life Sci. 1027:119–130. 2016. View Article : Google Scholar
|
|
5
|
Siu WS, Qin L, Cheung WH and Leung KS: A
study of trabecular bones in ovariectomized goats with
micro-computed tomography and peripheral quantitative computed
tomography. Bone. 35:21–26. 2004. View Article : Google Scholar
|
|
6
|
Lill CA, Fluegel AK and Schneider E: Sheep
model for fracture treatment in osteoporotic bone: A pilot study
about different induction regimens. J Oorthop Trauma. 14:559–566.
2000. View Article : Google Scholar
|
|
7
|
Wang Z and Zhao J: Research progress in
osteoporotic models. Chin J Osteoporos. 2012.
|
|
8
|
Silverman SL and Lane NE:
Glucocorticoid-induced osteoporosis. Curr Osteoporos Rep. 7:23–26.
2009. View Article : Google Scholar
|
|
9
|
Dai W, Jiang L, Lay YA, Chen H, Jin G,
Zhang H, Kot A, Ritchie RO, Lane NE and Yao W: Prevention of
glucocorticoid induced bone changes with beta-ecdysone. Bone.
74:48–57. 2015. View Article : Google Scholar :
|
|
10
|
Xia X, Kar R, Gluhak-Heinrich J, Yao W,
Lane NE, Bonewald LF, Biswas SK, Lo WK and Jiang JX:
Glucocorticoid-induced autophagy in osteocytes. J Bone Miner Res.
25:2479–2488. 2010. View
Article : Google Scholar :
|
|
11
|
Canalis E, Mazziotti G, Giustina A and
Bilezikian JP: Glucocorticoid-induced osteoporosis: Pathophysiology
and therapy. Osteoporos Int. 18:1319–1328. 2007. View Article : Google Scholar
|
|
12
|
Rauch A, Seitz S, Baschant U, Schilling
AF, Illing A, Stride B, Kirilov M, Mandic V, Takacz A,
Schmidt-Ullrich R, et al: Glucocorticoids suppress bone formation
by attenuating osteoblast differentiation via the monomeric
glucocorticoid receptor. Cell Metab. 11:517–531. 2010. View Article : Google Scholar
|
|
13
|
Schober A, Parlato R, Huber K, Kinscherf
R, Hartleben B, Huber TB, Schütz G and Unsicker K: Cell loss and
autophagy in the extra-adrenal chromaffin organ of Zuckerkandl are
regulated by glucocorticoid signalling. J Neuroendocrinol.
25:34–47. 2013. View Article : Google Scholar
|
|
14
|
Chang JK, Li CJ, Liao HJ, Wang CK, Wang GJ
and Ho ML: Anti-inflammatory drugs suppress proliferation and
induce apoptosis through altering expressions of cell cycle
regulators and pro-apoptotic factors in cultured human osteoblasts.
Toxicology. 258:148–156. 2009. View Article : Google Scholar
|
|
15
|
Jia J, Yao W, Guan M, Dai W, Shahnazari M,
Kar R, Bonewald L, Jiang JX and Lane NE: Glucocorticoid dose
determines osteocyte cell fate. FASEB J. 25:3366–3376. 2011.
View Article : Google Scholar :
|
|
16
|
Gao L, Cai G and Shi X: Beta-ecdysterone
induces osteogenic differentiation in mouse mesenchymal stem cells
and relieves osteoporosis. Biol Pharm Bull. 31:2245–2249. 2008.
View Article : Google Scholar
|
|
17
|
Omanakuttan A, Bose C, Pandurangan N,
Kumar GB, Banerji A and Nair BG: Nitric Oxide and ERK mediates
regulation of cellular processes by Ecdysterone. Exp Cell Res.
346:167–175. 2016. View Article : Google Scholar
|
|
18
|
Zhang X, Xu X, Xu T and Qin S:
β-Ecdysterone suppresses interleukin-1beta-induced apoptosis and
inflammation in rat chondrocytes via inhibition of NF-kappaB
signaling pathway. Drug Dev Res. 75:195–201. 2014.
|
|
19
|
Zou Y, Wang R, Guo H and Dong M:
Phytoestrogen β-ecdysterone protects PC12 cells against
MPP+−induced neurotoxicity in vitro: Involvement of
PI3K-Nrf2-regulated pathway. Toxicol Sci. 147:28–38. 2015.
View Article : Google Scholar
|
|
20
|
Pan Z, Niu Y, Liang Y, Zhang X and Dong M:
β-Ecdysterone protects SH-SY5Y cells against
6-hydroxydopamine-induced apoptosis via mitochondria-dependent
mechanism: Involvement of p38(MAPK)-p53 signaling pathway. Neurotox
Res. 30:453–466. 2016. View Article : Google Scholar
|
|
21
|
Ominsky MS, Li X, Asuncion FJ, Barrero M,
Warmington KS, Dwyer D, Stolina M, Geng Z, Grisanti M, Tan HL, et
al: RANKL inhibition with osteoprotegerin increases bone strength
by improving cortical and trabecular bone architecture in
ovariectomized rats. J Bone Miner Res. 23:672–682. 2008. View Article : Google Scholar
|
|
22
|
Houlihan DD, Mabuchi Y, Morikawa S, Niibe
K, Araki D, Suzuki S, Okano H and Matsuzaki Y: Isolation of mouse
mesenchymal stem cells on the basis of expression of Sca-1 and
PDGFR-α. Nat Protoc. 7:2103–2111. 2012. View Article : Google Scholar
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
24
|
Walsh CM and Edinger AL: The complex
interplay between autophagy, apoptosis, and necrotic signals
promotes T-cell homeostasis. Immunol Rev. 236:95–109. 2010.
View Article : Google Scholar :
|
|
25
|
Lee HK, Lund JM, Ramanathan B, Mizushima N
and Iwasaki A: Autophagy-dependent viral recognition by
plasmacytoid dendritic cells. Science. 315:1398–1401. 2007.
View Article : Google Scholar
|
|
26
|
Yue Z, Jin S, Yang C, Levine AJ and Heintz
N: Beclin 1, an autophagy gene essential for early embryonic
development, is a haploinsufficient tumor suppressor. Proc Natl
Acad Sci USA. 100:15077–15082. 2003. View Article : Google Scholar :
|
|
27
|
Lawrence T: The nuclear factor NF-kappaB
pathway in inflammation. Cold Spring Harb Perspect Biol.
1:a0016512009. View Article : Google Scholar :
|
|
28
|
McDonnell P, McHugh PE and O'Mahoney D:
Vertebral osteoporosis and trabecular bone quality. Ann Biomed Eng.
35:170–189. 2007. View Article : Google Scholar
|
|
29
|
Roux JP, Wegrzyn J, Boutroy S, Bouxsein
ML, Hans D and Chapurlat R: The predictive value of trabecular bone
score (TBS) on whole lumbar vertebrae mechanics: An ex vivo study.
Osteoporos Int. 24:2455–2460. 2013. View Article : Google Scholar
|
|
30
|
Chen P, Miller PD, Recker R, Resch H, Rana
A, Pavo I and Sipos AA: Increases in BMD correlate with
improvements in bone microarchitecture with teriparatide treatment
in postmenopausal women with osteoporosis. J Bone Miner Res.
22:1173–1180. 2007. View Article : Google Scholar
|
|
31
|
Jehle S, Hulter HN and Krapf R: Effect of
potassium citrate on bone density, microarchitecture, and fracture
risk in healthy older adults without osteoporosis: A randomized
controlled trial. J Clin Endocrinol Metab. 98:207–217. 2013.
View Article : Google Scholar
|
|
32
|
Styrkarsdottir U, Cazier JB, Kong A,
Rolfsson O, Larsen H, Bjarnadottir E, Johannsdottir VD,
Sigurdardottir MS, Bagger Y, Christiansen C, et al: Linkage of
osteoporosis to chromosome 20p12 and association to BMP2. PLoS
Biol. 1:E692003. View Article : Google Scholar :
|
|
33
|
Mundy C, Gannon M and Popoff SN:
Connective tissue growth factor (CTGF/CCN2) negatively regulates
BMP-2 induced osteoblast differentiation and signaling. J Cell
Physiol. 229:672–681. 2014. View Article : Google Scholar :
|
|
34
|
Yamaguchi Y, Sakai E, Sakamoto H, Fumimoto
R, Fukuma Y, Nishishita K, Okamoto K and Tsukuba T: Inhibitory
effects of tert-butylhydroquinone on osteoclast differentiation via
up-regulation of heme oxygenase-1 and down-regulation of HMGB1
release and NFATc1 expression. J Appl Toxicol. 34:49–56. 2014.
View Article : Google Scholar
|
|
35
|
Bustamante M, Nogués X, Agueda L, Jurado
S, Wesselius A, Cáceres E, Carreras R, Ciria M, Mellibovsky L,
Balcells S, et al: Promoter 2 −1025 T/C polymorphism in the RUNX2
gene is associated with femoral neck bmd in Spanish postmenopausal
women. Calcif Tissue Int. 81:327–332. 2007. View Article : Google Scholar
|
|
36
|
Zhu FB, Wang JY, Zhang YL, Quan RF, Yue
ZS, Zeng LR, Zheng WJ, Hou Q, Yan SG and Hu YG: Curculigoside
regulates proliferation, differentiation, and pro-inflammatory
cytokines levels in dexamethasone-induced rat calvarial
osteoblasts. Int J Clin Exp Med. 8:12337–12346. 2015.
|
|
37
|
Wang FS, Ko JY, Weng LH, Yeh DW, Ke HJ and
Wu SL: Inhibition of glycogen synthase kinase-3beta attenuates
glucocorticoid-induced bone loss. Life Sci. 85:685–692. 2009.
View Article : Google Scholar
|
|
38
|
Weinstein RS, Jilka RL, Parfitt AM and
Manolagas SC: Inhibition of osteoblastogenesis and promotion of
apoptosis of osteoblasts and osteocytes by glucocorticoids.
Potential mechanisms of their deleterious effects on bone. J Clin
Invest. 102:274–282. 1998. View
Article : Google Scholar :
|
|
39
|
Liu W, Dai N, Wang Y, Xu C, Zhao H, Xia P,
Gu J, Liu X, Bian J, Yuan Y, et al: Role of autophagy in
cadmium-induced apoptosis of primary rat osteoblasts. Sci Rep.
6:204042016. View Article : Google Scholar :
|
|
40
|
Piemontese M, Onal M, Xiong J, Wang Y,
Almeida M, Thostenson JD, Weinstein RS, Manolagas SC and O'Brien
CA: Suppression of autophagy in osteocytes does not modify the
adverse effects of glucocorticoids on cortical bone. Bone.
75:18–26. 2015. View Article : Google Scholar :
|
|
41
|
Wells GA, Cranney A, Peterson J, Boucher
M, Shea B, Robinson V, Coyle D and Tugwell P: Alendronate for the
primary and secondary prevention of osteoporotic fractures in
postmenopausal women. Cochrane Database Syst Rev.
1:CD0011552008.
|