Introduction
Chronic ischemic disease, including Buerger's
disease, diabetic foot ulcers and ischemic heart disease, seriously
impair the patients' quality of life. Chronic ischemic diseases are
primarily treated with pharmacological agents, surgery and
endovascular intervention (1).
However, currently available treatment strategies do not achieve
optimal effects when the integrity of the vascular outflow tract is
compromised. In addition, the application of these treatments is
limited to high-risk patients, despite surgical procedures
inevitably increasing vascular damage. Endothelial cells are
involved in the mechanisms underlying vascular injury repair
(2), and transplantation of stem
cells in patients with ischemic disease has been revealed to be
successful (3–5). Mesenchymal stem cells (MSCs) are a
subtype of somatic stem cells that originate from the bone marrow.
MSCs are characterized by multipotent differentiation into various
cell lineages, including osteoblasts, chondrocytes, and adipocytes
(6–8). Asahara et al (9) initially described endothelial
progenitor cells (EPCs), which are the predecessors of endothelial
cells and mainly originate from the bone marrow. EPCs can be
recruited and mobilized in the serum in response to local
stimulation and cell-cell interactions: EPCs differentiate into
endothelial cells to participate in angiogenesis and tissue lesion
repair (10,11). Therapeutic strategies based on
vascular stem cells are currently under research for the treatment
of several clinical conditions (12).
Previous studies from our lab have reported that
MSCs and EPCs adhere to each other in the bone marrow cavity and
in vitro (13,14), and that this mutual adhesion is
important for the biological functions of both cell types. EPCs
have been demonstrated to promote the differentiation of MSCs into
osteoblasts (15); however, the
effects of MSCs on the endothelial differentiation of EPCs have yet
to be elucidated.
Cell differentiation is a result of selective gene
expression. Cell differentiation pathways include extracellular and
intracellular signal transduction, and the role of regulatory
transcription factors is crucial (16,17).
Extracellular signals, including bone morphogenetic protein 2 and
growth factors, interact with cell-surface receptors to initiate
cellular differentiation through the regulation of transcription
factors (18). Previous studies
have suggested that the entire differentiation repertoire of a
given multipotent stem cell may theoretically be specified by a
single determining factor that is located at the top of a
regulatory hierarchy (19,20). Previous research on the interaction
between MSCs and endothelial cells has demonstrated the formation
of microvessel-like structures (21). The interactions between MSCs and
endothelial cells are regulated by paracrine factors, including
vascular endothelial growth factor (VEGF) (22), which is a potent angiogenic factor.
VEGF-induced mobilization of bone marrow-derived EPCs has been
reported to enhance EPC differentiation in vitro and to
potentiate corneal neovascularization in vivo (23).
Therefore, the present study aimed to investigate
whether MSC-derived VEGF may mediate the differentiation of EPCs
into endothelial cells and to explore the regulatory roles of
paracrine pathways in this process. Cluster of differentiation
(CD)31 and von Willebrand Factor (vWF) were used as specific
markers for an endothelial phenotype (24,25).
Materials and methods
Cell source and ethical approval
All experimental protocols used in the present study
were reviewed and approved by the Animal Care and Use Committee of
Shihezi University (Shihezi, China). A total of 24 male C57BL/6J
mice (wild-type; age, 6 weeks; weight, 28–35 g) were purchased from
Xinjiang Medical University (Ürümqi, China; certificate no. SYXK
[Xin] 2010–0001). Mice were maintained in the Animal Facility of
Shihezi University (Shihezi, China) under controlled conditions
(temperature, 20°C; humidity, 55±5%; 12-h light/dark cycles), with
free access to food and water and were used as a cell source. The
technique that was used to harvest and culture all cell types was
the same, except for the materials and the culture media that were
used. All cells used in subsequent experiments were the third
generation.
Isolation and culture of murine bone
marrow MSCs (BMMSCs)
BMMSCs were isolated using the technique reported in
our previous studies (13,14). Briefly, bone marrow cells were
collected from 6-week-old wild-type male C57BL/6 mice euthanized by
cervical dislocation. The cells were cultured in low-glucose
Dulbecco's modified Eagle's medium (LG-DMEM; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 100 U/ml
penicillin (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany),
streptomycin sulfate (100 µg/ml; Sigma-Aldrich; Merck KGaA), and
10% fetal bovine serum (FBS; Hyclone; GE Healthcare Life Sciences,
Logan, UT, USA) at 37°C in a 5% CO2 humidified
incubator. Following 72 h of adhesion, non-adherent cells were
removed, and adherent cells were cultured for an additional 7 days
with a single change of medium on the third day. Adherent cells
were then retrieved by trypsin digestion.
Aliquots of cells (1×106) were incubated
for 20 min at 4°C with phycoerythrin (PE)-conjugated anti-stem
cells antigen (Sca)-1 (cat. no. 108107; dilution, 1:40),
fluorescein isothiocyanate (FITC)-conjugated anti-CD29 (cat. no.
102205; dilution, 1:50), peridinin chlorophyll protein (Per
CP)-conjugated CD45 (cat. no. 202220; dilution, 1:20) and
allophycocyanin (APC)-conjugated anti-CD11b (cat. no. 201809;
dilution, 1:100; all from BioLegend, Inc., San Diego, CA, USA).
Acquisition was performed by fluorescence-activated cell sorting
(FACS) using a FACSAria flow cytometer (BD Biosciences, Franklin
Lakes, NJ, USA), and analysis was performed using FACSDiva software
version 6.1.3 (BD Biosciences). The sorted
CD29+/Sca-1+/CD45−/CD11b−
cells were enriched by further culture in LG-DMEM containing
penicillin, streptomycin sulfate and 10% FBS at 37°C in a 5%
CO2 humidified incubator.
Isolation and characterization of
mouse bone marrow EPCs (BMEPCs)
Bone marrow cells were collected and cultured as
aforementioned. Cell aliquots (1×106) were incubated for
20 min at 4°C with the following anti-mouse antibodies:
APC-conjugated anti-CD11b (dilution, 1:100; BioLegend, Inc.),
FITC-conjugated anti-CD31 (cat. no. 102506; dilution, 1:50;
BioLegend, Inc.), Per CP-conjugated anti-CD144 (cat. no.
46-1441-82; dilution, 1:50; eBioscience; Thermo Fisher Scientific,
Inc.) and PE-conjugated anti-CD133 (cat. no. 141203; dilution,
1:40; BioLegend, Inc.). Acquisition was performed using a FACSAria
flow cytometer, and data were analyzed using FACSDiva software
version 6.1.3. Sorted
CD133+/CD31+/CD144+/CD11b−
cells were enriched by further culture in endothelial growth basal
medium (EBM)-2 (Lonza Group, Ltd., Basel, Switzerland).
Experimental groups and induction
culture conditions
A total of 3 experimental groups were used: i)
Single culture group, where EPCs were seeded in a 6-well plate at a
density of 5×105 cells/plate. This group was used as a
negative control. ii) Co-culture group, where MSCs were plated into
the upper and EPCs into the lower chambers of transwell inserts
(Corning Incorporated, Corning, NY, USA) at a density of
5×105 cells/insert. iii) VEGF group, where EPCs
(5×105 cells) were seeded in a 6-well plate, in medium
supplemented with recombinant VEGF (20 ng/ml; PeproTech, Inc.,
Rocky Hill, NJ, USA). This group was used as a positive control.
The cell culture medium used in the experiments was EBM-2
supplemented with 5% FBS (Hyclone: GE Healthcare Life Sciences),
100 U/ml penicillin (Sigma-Aldrich; Merck KGaA) and 100 µg/ml
streptomycin (Sigma-Aldrich; Merck KGaA). Cells were incubated for
48 h at 37°C in a 5% CO2 humidified incubator. The
alterations in cell morphology were observed under an inverted
microscope (Leica Microsystems GmbH, Wetzlar, Germany).
Immunofluorescence
EPCs (1×105 cells) were washed 3 times in
PBS, fixed in 4% paraformaldehyde for 15 min at room temperature,
and blocked in 3% BSA (Sigma-Aldrich; Merck KGaA) to block
non-specific binding sites at 37°C for 1 h. Cells were then
incubated with the following primary antibodies for 4 h at 37°C:
Rabbit polyclonal anti-CD31 (cat. no. ab28364; 1:50; Abcam,
Cambridge, MA, USA) and goat polyclonal anti-vWF (cat. no. ab11713;
1:100; Abcam). Following washing 3 times with PBS, cells were
incubated with secondary FITC-conjugated goat anti-rabbit
immunoglobulin (Ig)G (cat. no. TA130021; 1:100; OriGene
Technologies, Inc., Beijing, China,) and secondary FITC-conjugated
rabbit anti-goat IgG (cat. no. TA130029; 1:200; OriGene
Technologies, Inc.) at 37°C for 1 h. Cells were then washed and
soaked in ddH2O 3 times and examined under a DM IL Leica
fluorescence microscope (Leica Microsystems GmbH). Experiments were
performed in duplicate and repeated at least 3 times.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Cultured EPCs (1×106 cells) were washed
twice with ice-cold PBS. Total RNA was extracted using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. RNA purity was determined by the ratio
of absorbance at 260 and 280 nm (A260/A280)
using a Nanodrop 2000 spectrophotometer (Thermo Fisher Scientific,
Inc., Wilmington, DE, USA), where a range of 1.8–2.0 was considered
suitable for cDNA synthesis. Total RNA (200 ng) was reverse
transcribed into cDNA using a QuantiTect Reverse Transcription kit
(Qiagen, Inc., Valencia, CA, USA) according to the manufacturer's
protocol. The temperature protocol was as follows: Initial
denaturation at 70°C for 5 min, then 4°C for 2 min, followed by
42°C for 1 h and 70°C for 5 min. qPCR was performed on cDNA using
the QuantiNova SYBR Green PCR kit (Qiagen, Inc.), according to the
manufacturer's protocol. The thermocycling conditions were as
follows: Initial denaturation at 95°C for 3 min, followed by 40
cycles at 95°C for 10 sec and at 55°C for 30 sec. The reaction
included 10 µl SYBR Green PCR mix, 2 µl primers, 3 µl cDNA (500 ng)
and 5 µl DNase/RNase free water to a final reaction volume of 20
µl. The results were analyzed using Bio-Rad CFX96 Manager software
version 3.1 (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Data
were collected after each annealing step. β-actin was used as an
endogenous control to normalize for differences in the amount of
total RNA in each sample. The primer sequences and the sizes of the
amplified fragments were as follows: CD31 (114 bp), forward
5′-CCAACAGAGCCAGCAGTATG-3′, reverse 5′-TGACCACTCCAATGACAACC-3′; vWF
(108 bp), forward 5′-TGCCTCAGTGGGAGAAAGAT-3′, reverse
5′-CAGGTTTGTGCTCTGCTTGA-3′; and β-actin (203 bp), forward
5′-TTCCTTCTTGGGTATGGAAT-3′ and reverse 5′-GAGCAATGATCTTGATCTTC-3′.
Experiments were performed in triplicate and repeated 3 times.
Results were quantified using the 2−∆∆Cq method
(26).
Western blot analysis
Cultured EPCs (1×106 cells) were washed
twice with ice-cold PBS. Total proteins were extracted from EPCs
using radioimmunoprecipitation assay lysis buffer (Thermo Fisher
Scientific, Inc.) containing phenylmethylsulfonyl fluoride (Thermo
Fisher Scientific, Inc.). Protein concentration was measured using
a bicinchoninic acid protein assay kit (Thermo Fisher Scientific,
Inc.). Equal amounts of extracted protein samples (40 µg) were
separated by 6% SDS-PAGE and transferred onto nitrocellulose
membranes. Membranes were blocked by incubation in TBS containing
3% BSA (Sigma-Aldrich; Merck KGaA) for 2 h at room temperature.
Membranes were incubated overnight at 4°C with the following
primary antibodies: polyclonal rabbit anti-CD31 (cat. no. ab28364;
1:500; Abcam) and polyclonal goat anti-vWF (cat. no. ab11713;
1:1,000; Abcam), with gentle agitation. After washing with TBS, the
membrane was incubated for 2 h at room temperature with goat
anti-rabbit (cat. no. ZB-2301; 1:10,000), rabbit anti-goat (cat.
no. ZB-2306; 1:10,000) or goat anti-mouse (cat. no. ZB-2305;
1:20,000; all from OriGene Technologies, Inc.) horseradish
peroxidase-conjugated secondary antibodies. Protein bands were
visualized using enhanced chemiluminescence (GE Healthcare Life
Sciences). β-actin (cat. no. NB600-501; 1:1,000, Novus Biologicals,
LLC, Littleton, CO, USA) was used as an endogenous control for
normalization. Blots were semi-quantified by densitometry using
Gel-Pro Analyzer software version 4.0 (Media Cybernetics, Inc.,
Rockville, MD, USA). Experiments were performed in triplicate and
repeated 3 times.
Detection of VEGF levels using
ELISA
MSCs that were at passage 3 were cultured at a
density of 1×106 cells/plate in serum- and growth
factor-free LG-DMEM without FBS for 24 h at 37°C in an atmosphere
containing 5% CO2. The medium was subsequently
collected, passed through a 0.22 µm filter, and the concentration
of VEGF in the MSC conditioned media (MSCCM) was
measured using an ELISA kit (cat. no. E-EL-M1292c; Elabscience
Biotechnology Co., Ltd., Wuhan, China) according to the
manufacturer's protocol. The absorbance at 450 nm was measured
using a Bio-Rad Model 3550-UV microplate reader (Bio-Rad
Laboratories, Inc.). Experiments were performed in triplicate and
repeated 3 times.
The culture medium of the cells from the 3
experimental groups was also collected, following 48 h of culture,
passed through a 0.22 µm filter, and used for VEGF detection by
ELISA as aforementioned.
VEGF blocking assay
A total of 4 experimental groups were used: i)
LG-DMEM group, where EPCs (5×105) that were at passage 3
were seeded in LG-DMEM without FBS for 48 h at 37°C in a 5%
CO2 humidified incubator; ii) MSCCM group,
where EPCs (5×105) that were at passage 3 were seeded in
MSCCM for 48 h at 37°C in a 5% CO2 humidified
incubator; iii) MSCCM with mouse IgG group, where EPCs
(5×105) that were at passage 3 were seeded in
MSCCM with mouse IgG (cat. no. ZDR-5109; 100 ng/ml;
OriGene Technologies, Inc.) for 48 h at 37°C in a 5% CO2
humidified incubator; iv) MSCCM with anti-VEGF antibody
group, where EPCs (5×105) that were at passage 3 were
seeded in MSCCM with anti-VEGF antibody (cat. no.
ab9570; 100 ng/ml; Abcam) for 48 h at 37°C in a 5% CO2
humidified incubator.
Statistical analysis
Statistical analyses were conducted using SPSS
software version 17.0 (SPSS Inc., Chicago, IL, USA). Data are
expressed as the mean ± standard error of the mean. The statistical
significance of the differences between groups was assessed using
an unpaired Student's t-test for pair-wise comparisons or a one-way
analysis of variance followed by Dunnett's post hoc multiple
comparisons test. P<0.05 was considered to indicate a
statistically significant difference.
Results
MSCs promote the endothelial
differentiation of EPCs
We have previously reported that MSCs and EPCs
adhere to each other in the bone marrow cavity and in vitro
(13,14), and that this mutual adhesion is
important for the biological functions of both cell types. EPCs
have also been reported to promote the differentiation of MSCs into
osteoblasts (15). Therefore, the
effects of MSCs on EPC differentiation were investigated in the
present study. EPCs and MSCs were co-cultured in transwell inserts
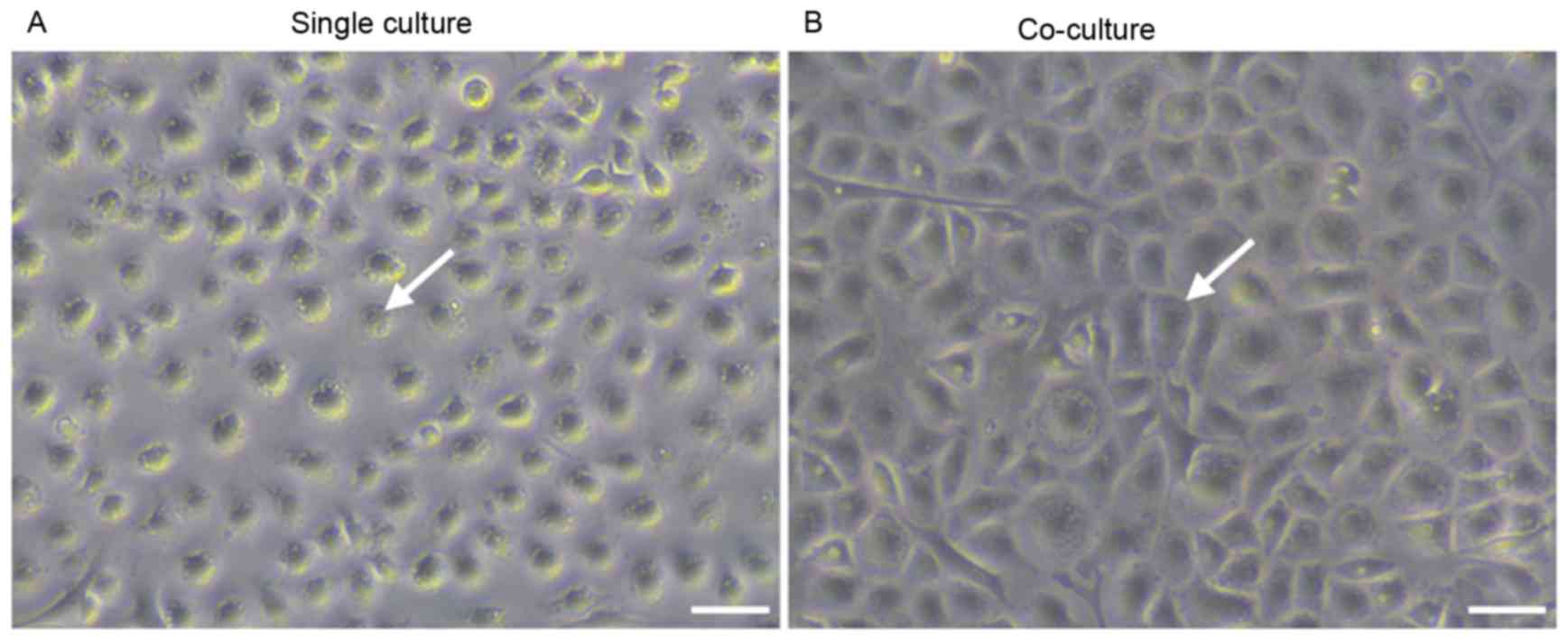
in the absence of direct cellular interactions. Fig. 1 demonstrated the morphological
alterations that were observed in differentiated EPCs. Cells in the
single culture group appeared to be irregularly shaped (Fig. 1A), whereas cells in the co-culture
group displayed a cobblestone endothelial-like appearance (Fig. 1B). These findings suggested that
MSCs may promote the endothelial differentiation of EPCs in
vitro.
VEGF is implicated in EPC
differentiation
Previous studies (21–23)
have suggested a role for VEGF in MSC-promoted EPC differentiation.
EPCs that were cultured in the presence of VEGF (20 ng/ml)
stimulation were used as a positive control in the present study.
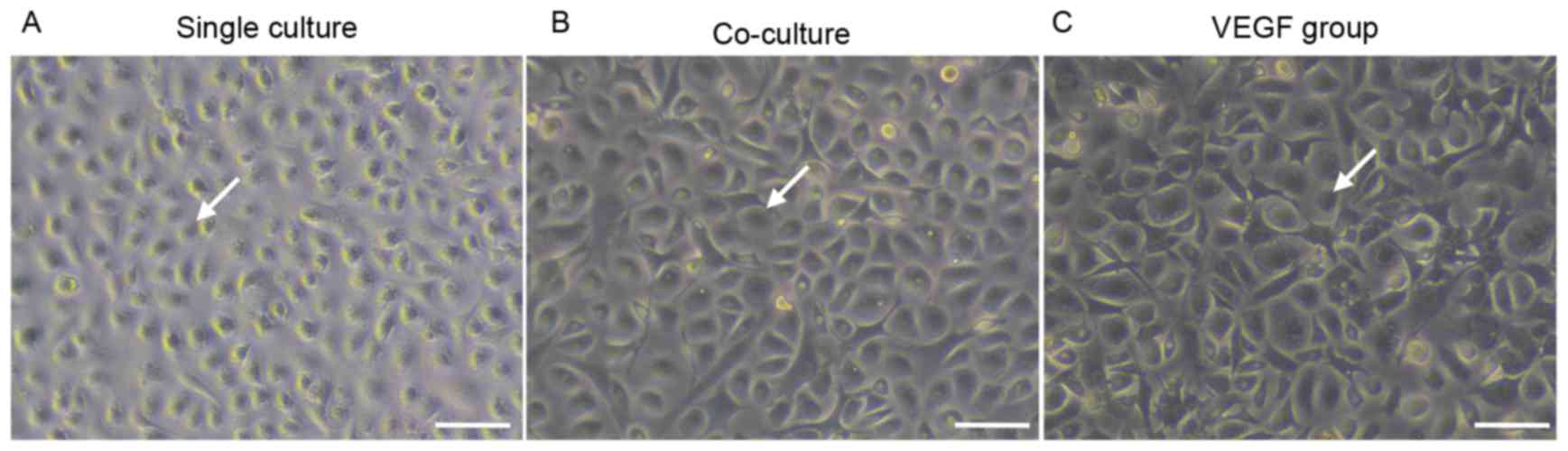
As presented in Fig. 2, following
48 h of culture, EPCs cultured alone appeared to be irregularly
shaped (Fig. 2A), whereas cells in
the co-culture and VEGF groups demonstrated a cobblestone
appearance (Fig. 2B and C).
To assess the phenotypes of the differentiated
cells, the expression of the endothelial cell markers CD31 and vWF
was examined (Fig.3). EPCs in the
single culture group were revealed to weakly express CD31 (Fig. 3A) and vWF (Fig. 3D), whereas EPCs in the co-culture
and VEGF groups highly expressed CD31 (Fig. 3B and C) and vWF (Fig. 3E and F). The expression of specific
endothelial markers was quantified using RT-qPCR and western
blotting. RT-qPCR demonstrated that the mRNA expression levels of
CD31 and vWF were significantly upregulated in the co-culture and
VEGF groups compared with in cells from the single culture group
(Fig. 3G). Similarly, western blot
analysis revealed that CD31 and vWF protein expression was
significantly enhanced in MSC-co-cultured and VEGF-treated cells
compared with in the single culture group (Fig. 3H and I). In addition, the protein
expression levels of CD31 and vWF were significantly increased in
the co-culture group compared with in the VEGF group (Fig. 3H and I). These results suggested
that VEGF may promote the differentiation of EPCs into endothelial
cells in vitro.
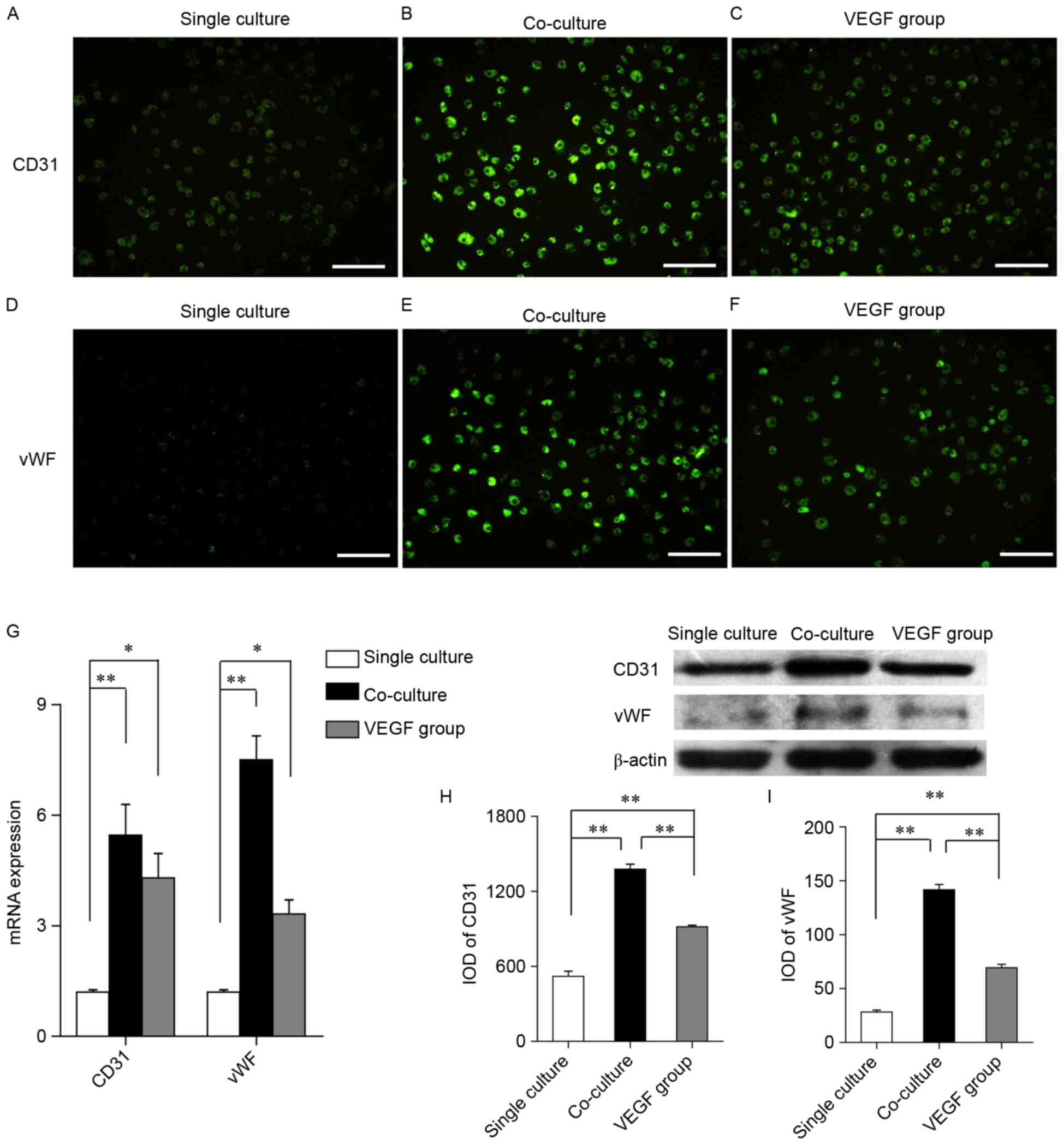 | Figure 3.VEGF and MSCs upregulate the
expression of endothelial cell differentiation markers in EPCs
in vitro. (A-C) Immunofluorescence staining demonstrating
CD31 expression in the (A) single culture, (B) co-culture and (C)
VEGF groups. (D-F) Immunofluorescence staining demonstrating vWF
expression in the (D) single culture, (E) co-culture and (F) VEGF
groups. Scale bar, 50 µm. (G) Reverse transcription-quantitative
polymerase chain reaction was used to assess the mRNA expression of
CD31 and vWF in the single culture, co-culture and VEGF groups. (H
and I) Western blotting revealed (H) CD31 and (I) vWF protein
expression. Data are expressed as the mean ± standard error of the
mean. *P<0.05 and **P<0.01, as indicated. VEGF, vascular
endothelial growth factor; MSC, mesenchymal stem cell; EPC,
endothelial progenitor cell; CD, cluster of differentiation; vWF,
von Willebrand factor; IOD, integrated optical density. |
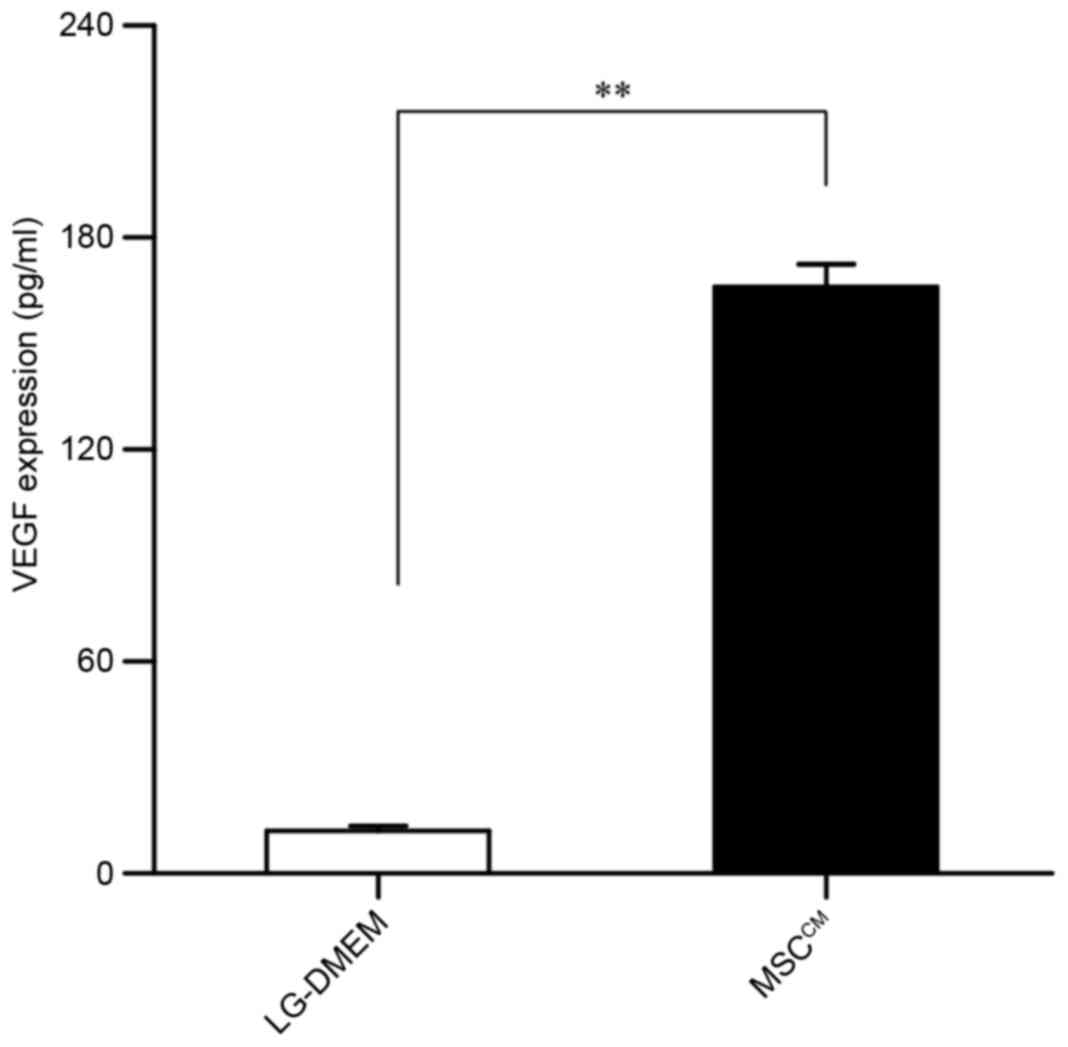
MSCs secrete VEGF
On the basis of the aforementioned findings, we
investigated whether MSCs may secrete VEGF, which may mediate EPC
differentiation. VEGF levels in MSCCM were measured
using ELISA. As presented in Fig.
4, VEGF levels were significantly upregulated in
MSCCM compared with in serum- and growth factor-free
LG-DMEM. A previous study from our group (15) demonstrated that EPCs secreted VEGF;
in combination with the present results, it may be hypothesized
that the MSC-promoted EPC differentiation may be related to VEGF
secretion by MSCs.
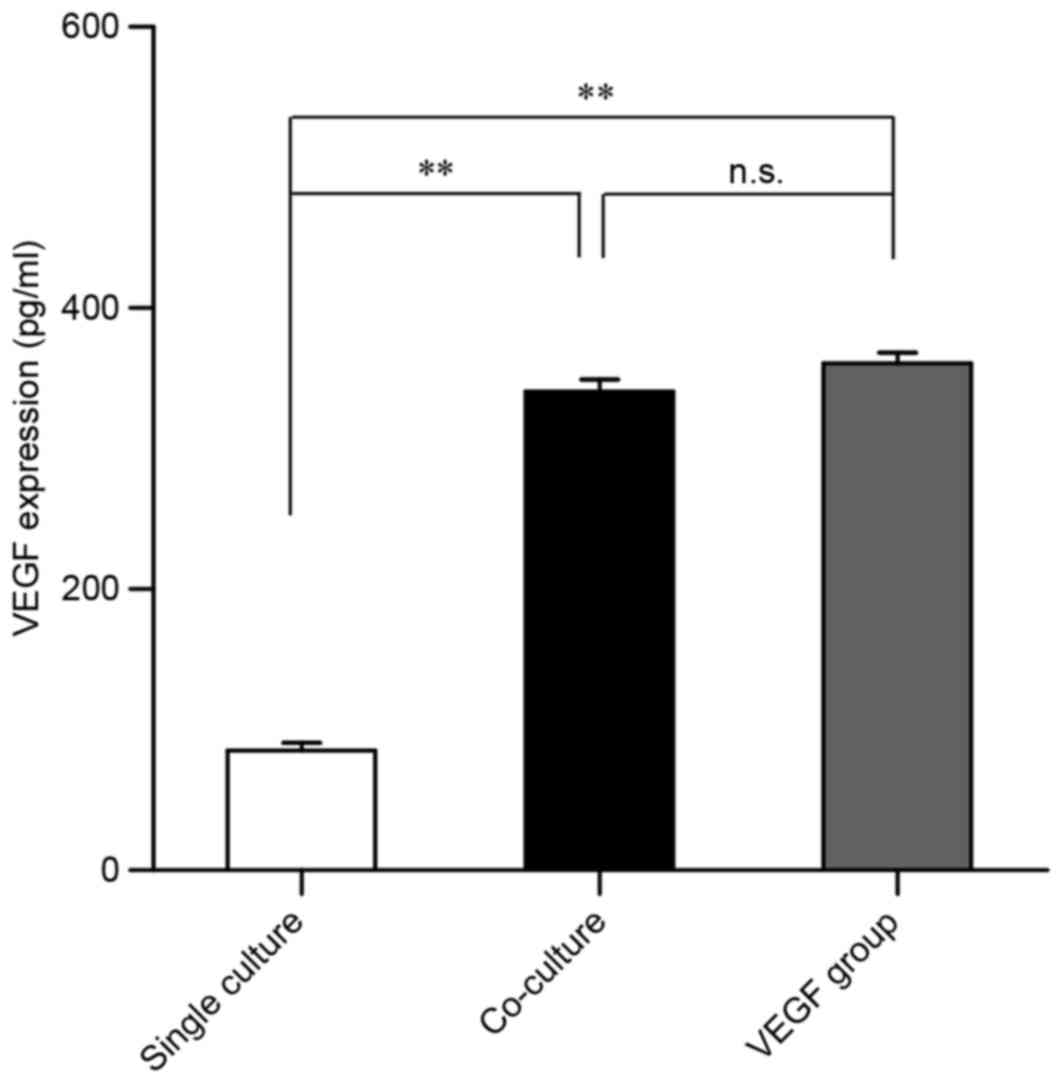
VEGF levels are increased in
co-culture conditioned media
Based on the aforementioned findings, the culture
medium from each of the 3 experimental groups was used to measure
VEGF levels via ELISA. Compared with the single culture group, VEGF
levels were significantly upregulated in conditioned media form the
co-culture and VEGF groups (Fig.
5). No statistically significant differences in VEGF levels
were detected between the co-culture and VEGF groups (Fig. 5). These results also suggested that
MSC-secreted VEGF may mediate the endothelial differentiation of
EPCs in vitro.
Anti-VEGF inhibits the differentiation
of EPCs
To further investigate whether MSC-secreted VEGF may
promote the endothelial differentiation of EPCs, cells were treated
with a VEGF neutralizing antibody or mouse IgG (Fig. 6). EPCs were cultured in LG-DMEM,
MSCCM, MSCCM with mouse IgG, and
MSCCM with anti-VEGF antibody for 48 h. The expression
of CD31 (Fig. 6A-C) and vWF
(Fig. 6E-G) appeared to be
enhanced in the MSCCM groups compared with in the
LG-DMEM group. However, EPCs cultured in MSCCM with
anti-VEGF antibody were demonstrated to weakly express CD31
(Fig. 6D) and vWF (Fig. 6H) compared with EPCs cultured in
MSCCM and MSCCM with IgG. RT-qPCR revealed
that CD31 and vWF mRNA expression levels were significantly
upregulated in the MSCCM group compared with in the
LG-DMEM group (Fig. 6I and J),
whereas treatment with IgG did not appear to affect CD31 and vWF
mRNA expression. However, CD31 and vWF mRNA expression levels were
significantly downregulated in EPCs cultured in MSCCM
and treated with the anti-VEGF antibody (Fig. 6I and J). Western blot analysis
revealed similar effects in CD31 and vWF protein expression
(Fig. 6K and L). The present
results demonstrated that anti-VEGF significantly inhibited the
protein expression of CD31 and vWF in EPCs. These findings
suggested that VEGF may promote the in vitro differentiation
of EPCs into endothelial cells.
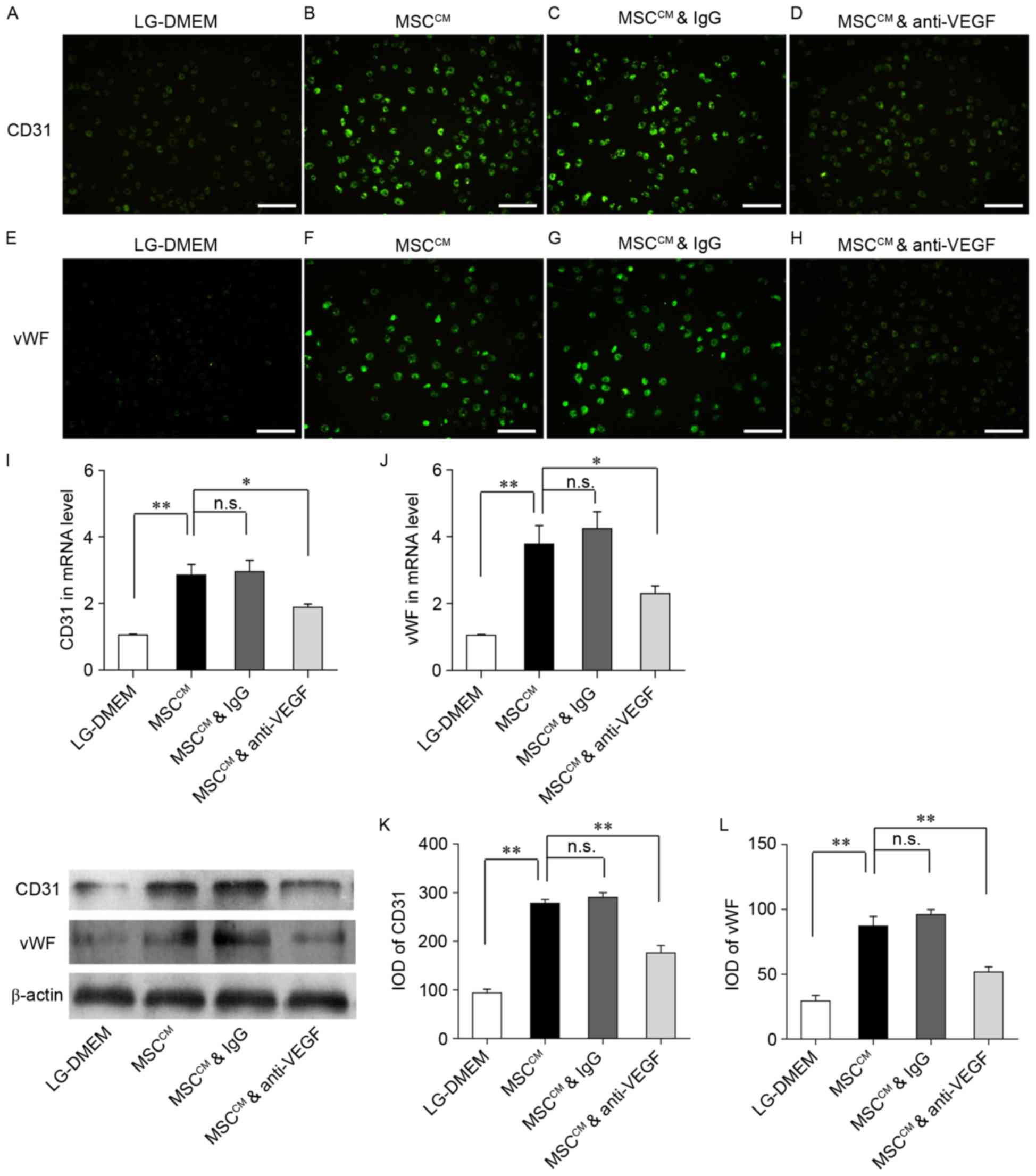 | Figure 6.Anti-VEGF neutralizing antibody
inhibits the expression of endothelial markers in differentiating
EPCs in vitro. A VEGF neutralizing antibody (100 ng/ml) was
added to MSCCM. MSCCM with mouse IgG (100
ng/ml) was used as the control. EPCs were cultured in LG-DMEM,
MSCCM, MSCCM with IgG and MSCCM
with anti-VEGF antibody for 48 h. (A-D) Immunofluorescence staining
demonstrating CD31 expression in the (A) LG-DMEM, (B)
MSCCM, (C) MSCCM with IgG and (D)
MSCCM with anti-VEGF groups. (E-H) Immunofluorescence
staining demonstrating vWF expression in the (E) LG-DMEM, (F)
MSCCM, (G) MSCCM with IgG and (H)
MSCCM with anti-VEGF groups. Scale bar, 50 µm. (I and J)
Reverse transcription-quantitative polymerase chain reaction was
used to assess the expression of CD31 and vWF mRNA in the LG-DMEM,
MSCCM, MSCCM with IgG and MSCCM
with anti-VEGF groups. (K and L) Western blotting demonstrated the
protein expression levels of (K) CD31 and (L) vWF in the LG-DMEM,
MSCCM, MSCCM with IgG and MSCCM
with anti-VEGF groups. Data are expressed as the mean ± standard
error of the mean. *P<0.05, **P<0.01, as indicated. VEGF,
vascular endothelial growth factor; EPC, endothelial progenitor
cell; MSC, mesenchymal stem cell; MSCCM, MSC conditioned
media; Ig, immunoglobulin; LG-DMEM, low-glucose Dulbecco's modified
Eagle's medium; CD, cluster of differentiation; vWF, von Willebrand
factor; n.s., not significant; IOD, integrated optical density. |
Discussion
Cell differentiation is a highly significant process
for living organisms, and it results in the optimized formation of
various cell populations with specialized biological functions.
VEGF is a cytokine that has been implicated in numerous cell
differentiation pathways and regulatory mechanisms: VEGF is
critical for determining the fate of differentiating cells and
controlling the differentiation process (21,27,28).
The various cellular functions of VEGF result from its ability to
initiate a diverse, complex and integrated network of signaling
pathways through its main receptor, the kinase insert domain
receptor: VEGF can stimulate cell differentiation, proliferation,
migration and survival (29,30).
Previous studies have reported that VEGF was critical for the
differentiation of endothelial cells, and that nitric oxide was an
important effector of the biological actions of VEGF (31–33).
In addition, VEGF has been reported to induce the differentiation
of mouse multipotent adult progenitor cells into endothelial cells
(34,35), through a mitogen-activated protein
kinase/extracellular signal-regulated kinase 1/2 signaling
pathway-mediated mechanism (36).
These findings suggested that VEGF may influence cell
differentiation through complex interactions and various signal
transduction pathways.
The differentiation of EPCs into endothelial cells
has been suggested to be critical for endothelial repair (2). Stem/progenitor cell differentiation
is closely related to the cellular microenvironment (19,20),
and VEGF may be implicated in the differentiation mechanisms. In
the present study, MSCs were demonstrated to regulate the
endothelial differentiation of EPCs, through the secretion of
paracrine factors, including VEGF. EPCs were co-cultured with MSCs
for 48 h in vitro and the observed morphological alterations
suggested that MSCs may be implicated in the differentiation of
EPCs into endothelial cells. Phase-contrast microscopy was used to
examine the effects of VEGF on EPC morphology: Cells in the single
culture group appeared to be irregularly shaped, whereas cells in
the co-culture and VEGF groups displayed a cobblestone
endothelial-like appearance.
To assess the phenotypes of differentiated cells,
the expression of the endothelial cell markers CD31 and vWF
(24,25) was examined. Immunofluorescence,
RT-qPCR, and western blotting results revealed that CD31 and vWF
expression levels were significantly upregulated in the co-culture
and VEGF groups compared with the single culture group, thus
suggesting that VEGF may have similar effects to the presence of
MSCs on EPC differentiation. Furthermore, MSCs were demonstrated to
secrete VEGF, in accordance with our previous report (15).
VEGF levels in the conditioned media form the 3
experimental groups were also measured: VEGF levels were
significantly increased in the co-culture group compared with in
the single culture group. To further investigate the roles of VEGF
during EPC differentiation, MSCCM were neutralized with
an anti-VEGF antibody. Anti-VEGF was demonstrated to inhibit the
effects of secreted VEGF in MSCCM on the endothelial
differentiation of EPCs. These findings suggested that VEGF may
promote EPC differentiation to endothelial cells in
vitro.
VEGF may be among the several cytokines that
influence EPC differentiation. VEGF expression was not revealed to
be significantly different between the co-culture and VEGF groups;
however, the expression of the endothelial cell markers CD31 and
vWF was significantly increased in the co-culture group compared
with in the VEGF group. In addition, blocking the influence of VEGF
in MSCCM was revealed to diminish the effects of
MSCCM on EPC differentiation. EPC differentiation was
not abolished, possible due to the presence of other cytokines
secreted by MSCs in the medium, which may exert synergistic effects
on EPC differentiation. Guo et al (37) reported that EPCs derived from
CD34+ cells could differentiate into endothelial-like
cells, and that differentiation was induced by the addition of
basic fibroblast growth factor or platelet-derived growth
factor-BB. Previous studies have also demonstrated that VEGF was
critical for the in vitro differentiation of EPCs or
hematopoietic stem cells into endothelial cells (38–40).
These findings suggested that several MSC-released cytokines may
have additive or synergistic effects on EPC differentiation
pathways. Therefore, compared with single cytokine approaches,
therapeutic strategies based on the combination of MSCs with EPCs
may be more effective in enhancing tissue repair in humans.
Several kinds of seed cells have been used in
applications of tissue engineering to promote angiogenesis in
ischemic diseases. MSCs and EPCs have favorable biological
characteristics that make them suitable for use in tissue
engineering applications: They are able to migrate to injured
tissues, and they are able to differentiate into various cell
phenotypes according to the type of tissue in which they reside
(41). In addition, MSCs
demonstrate potent self-renewal properties, and can secrete various
cytokines that may exert a greater or synergetic impact on the
biological function of EPCs compared with one type of cytokine.
Furthermore, MSCs are more easily available compared with cytokines
(42,43). Therefore, therapeutic approaches
based on the co-culture of EPCs with MSCs may have potential to be
successfully applied in tissue engineering and regenerative
medicine strategies.
In conclusion, the results of the present study
suggested that the presence of MSCs may enhance the in vitro
endothelial differentiation of EPCs, through a paracrine mechanism
that may involve the secretion of cytokines, including VEGF,
instead of direct cell-cell contacts. Further studies are required
to explore the molecular mechanisms that mediate EPC
differentiation and to elucidate the exact roles of MSCs in the
differentiation processes. As endothelial cells can participate in
endothelial repair and angiogenesis following ischemia, the
findings of the present study may promote the understanding of
tissue repair mechanisms, and may lead to the development of novel
strategies for therapeutic interventions aimed at ischemic
diseases.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 31271458), the
Science and Technology Program of Xinjiang Production and
Construction Corps (grant no. 2014AB047), the Scientific Research
Foundation for the Returned Overseas Chinese Scholars, the Ministry
of Human Resources and Social Security of the People's Republic of
China (grant no. RSLX201201) and the Shihezi University Youth
Science and Technology Research and Development Program, Basis and
Application Research Project (grant no. 20142RKXYQ20).
References
|
1
|
Lenti M, Cieri E, De Rango P, Pozzilli P,
Coscarella C, Bertoglio C, Troiani R and Cao P: Endovascular
treatment of long lesions of the superficial femoral artery:
Results from a multicenter registry of a spiral, covered
polytetrafluoroethylene stent. J Vasc Surg. 45:32–39. 2007.
View Article : Google Scholar
|
|
2
|
Vasa M, Fichtlscherer S, Aicher A, Adler
K, Urbich C, Martin H, Zeiher AM and Dimmeler S: Number and
migratory activity of circulating endothelial progenitor cells
inversely correlate with risk factors for coronary artery disease.
Circ Res. 89:E1–E7. 2001. View Article : Google Scholar
|
|
3
|
Tateishi-Yuyama E, Matsubara H, Murohara
T, Ikeda U, Shintani S, Masaki H, Amano K, Kishimoto Y, Yoshimoto
K, Akashi H, et al: Therapeutic angiogenesis for patients with limb
ischaemia by autologous transplantation of bone-marrow cells: A
pilot study and a randomised controlled trial. Lancet. 360:427–435.
2002. View Article : Google Scholar
|
|
4
|
Kim SW, Han H, Chae GT, Lee SH, Bo S, Yoon
JH, Lee YS, Lee KS, Park HK and Kang KS: Successful stem cell
therapy using umbilical cord blood-derived multipotent stem cells
for Buerger's disease and ischemic limb disease animal model. Stem
Cells. 24:1620–1626. 2006. View Article : Google Scholar
|
|
5
|
Saigawa T, Kato K, Ozawa T, Toba K,
Makiyama Y, Minagawa S, Hashimoto S, Furukawa T, Nakamura Y, Hanawa
H, et al: Clinical application of bone marrow implantation in
patients with arteriosclerosis obliterans, and the association
between efficacy and the number of implanted bone marrow cells.
Circ J. 68:1189–1193. 2004. View Article : Google Scholar
|
|
6
|
Friedenstein AJ, Chailakhjan RK and
Lalykina KS: The development of fibroblast colonies in monolayer
cultures of guinea-pig bone marrow and spleen cells. Cell Tissue
Kinet. 3:393–403. 1970.
|
|
7
|
Bianco P, Robey PG and Simmons PJ:
Mesenchymal stem cells: Revisiting history, concepts and assays.
Cell Stem Cell. 2:313–319. 2008. View Article : Google Scholar :
|
|
8
|
Wu X, Pang L, Lei W, Lu W, Li J, Li Z,
Frassica FJ, Chen X, Wan M and Cao X: Inhibition of Sca-1-positive
skeletal stem cell recruitment by alendronate blunts the anabolic
effects of parathyroid hormone on bone remodeling. Cell Stem Cell.
7:571–580. 2010. View Article : Google Scholar :
|
|
9
|
Asahara T, Murohara T, Sullivan A, Silver
M, van der Zee R, Li T, Witzenbichler B, Schatteman G and Isner JM:
Isolation of putative progenitor endothelial cells for
angiogenesis. Science. 275:964–967. 1997. View Article : Google Scholar
|
|
10
|
Ribatti D: The discovery of endothelial
progenitor cells. An historical review. Leuk Res. 31:439–444. 2007.
View Article : Google Scholar
|
|
11
|
Cao X, Wu X, Frassica D, Yu B, Pang L,
Xian L, Wan M, Lei W, Armour M, Tryggestad E, Wong J, Wen CY, Lu WW
and Frassica FJ: Irradiation induces bone injury by damaging bone
marrow microenvironment for stem cells. Proc Natl Acad Sci USA.
108:1609–1614. 2011. View Article : Google Scholar :
|
|
12
|
Geng YJ, Yang YJ, Casscells SW and
Willerson JT: Vascular stem cells: A new concept in the
pathogenesis of atherosclerosis and interventions for coronary
heart disease. Future Cardiol. 2:585–592. 2006. View Article : Google Scholar
|
|
13
|
Zhang H, Xian L, Lin Z, Yang C, Zhang M,
Feng W, Peng X, Chen X and Wu X: Endothelial progenitor cells as a
possible component of stem cell niche to promote self-renewal of
mesenchymal stem cells. Mol Cell Biochem. 397:235–243. 2014.
View Article : Google Scholar
|
|
14
|
Xia J, Zhang H, Gao X, Guo J, Hou J, Wang
X, Wang S, Yang T, Zhang X, Ge Q, et al: E-cadherin-mediated
contact of endothelial progenitor cells with mesenchymal stem cells
through β-catenin signaling. Cell Biol Int. 40:407–418. 2016.
View Article : Google Scholar
|
|
15
|
Zhang M, Zhang H, Feng W, Wang Y, Yin S,
Chen X and Wu X: Endothelial progenitor cells promote osteogenic
differentiation of marrow stromal cells in a paracrine manner.
Zhonghua Yi Xue Za Zhi. 95:1253–1257. 2015.(In Chinese).
|
|
16
|
Bost F, Caron L, Marchetti I, Dani C, Le
Marchand-Brustel Y and Binétruy B: Retinoic acid activation of the
ERK pathway is required for embryonic stem cell commitment into the
adipocyte lineage. Biochem J. 361:621–627. 2002. View Article : Google Scholar :
|
|
17
|
Tashiro K, Kawabata K, Sakurai H, Kurachi
S, Sakurai F, Yamanishi K and Mizuguchi H: Efficient adenovirus
vector-mediated PPAR gamma gene transfer into mouse embryoid bodies
promotes adipocyte differentiation. J Gene Med. 10:498–507. 2008.
View Article : Google Scholar
|
|
18
|
Shah NM, Groves AK and Anderson DJ:
Alternative neural crest cell fates are instructively promoted by
TGFbeta superfamily members. Cell. 85:331–343. 1996. View Article : Google Scholar
|
|
19
|
Cepko CL, Austin CP, Yang X, Alexiades M
and Ezzeddine D: Cell fate determination in the vertebrate retina.
Proc Natl Acad Sci USA. 93:589–595. 1996. View Article : Google Scholar :
|
|
20
|
Morrison SJ, Shah NM and Anderson DJ:
Regulatory mechanisms in stem cell biology. Cell. 88:287–298. 1997.
View Article : Google Scholar
|
|
21
|
Li H, Daculsi R, Grellier M, Bareille R,
Bourget C, Remy M and Amedee J: The role of vascular actors in two
dimensional dialogue of human bone marrow stromal cell and
endothelial cell for inducing self-assembled network. PLoS One.
6:e167672011. View Article : Google Scholar :
|
|
22
|
Mayer H, Bertram H, Lindenmaier W, Korff
T, Weber H and Weich H: Vascular endothelial growth factor (VEGF-A)
expression in human mesenchymal stem cells: Autocrine and paracrine
role on osteoblastic and endothelial differentiation. J Cell
Biochem. 95:827–839. 2005. View Article : Google Scholar
|
|
23
|
Asahara T, Takahashi T, Masuda H, Kalka C,
Chen D, Iwaguro H, Inai Y, Silver M and Isner JM: VEGF contributes
to postnatal neovascularization by mobilizing bone marrow-derived
endothelial progenitor cells. EMBO J. 18:3964–3972. 1999.
View Article : Google Scholar :
|
|
24
|
Newman PJ, Berndt MC, Gorski J, White GC
II, Lyman S, Paddock C and Muller WA: PECAM-1 (CD31) cloning and
relation to adhesion molecules of the immunoglobulin gene
superfamily. Science. 247:1219–1222. 1990. View Article : Google Scholar
|
|
25
|
da Silva Lopes M and Cutler DF: von
Willebrand factor multimerization and the polarity of secretory
pathways in endothelial cells. Blood. 128:277–285. 2016. View Article : Google Scholar :
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(Delta Delta C (T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
27
|
Oswald J, Boxberger S, Jørgensen B,
Feldmann S, Ehninger G, Bornhäuser M and Werner C: Mesenchymal stem
cells can be differentiated into endothelial cells in vitro. Stem
Cells. 22:377–384. 2004. View Article : Google Scholar
|
|
28
|
Zaniboni A, Bernardini C, Bertocchi M,
Zannoni A, Bianchi F, Avallone G, Mangano C, Sarli G, Calzà L,
Bacci ML and Forni M: In vitro differentiation of porcine aortic
vascular precursor cells to endothelial and vascular smooth muscle
cells. Am J Physiol Cell Physiol. 309:C320–C331. 2015. View Article : Google Scholar
|
|
29
|
Zachary I: VEGF signalling: Integration
and multi-tasking in endothelial cell biology. Biochem Soc Trans.
31:1171–1177. 2003. View Article : Google Scholar
|
|
30
|
Gupta K, Ramakrishnan S, Browne PV,
Solovey A and Hebbel RP: A novel technique for culture of human
dermal microvascular endothelial cells under either serum-free or
serum-supplemented conditions: Isolation by panning and stimulation
with vascular endothelial growth factor. Exp Cell Res. 230:244–251.
1997. View Article : Google Scholar
|
|
31
|
Fong GH, Rossant J, Gertsenstein M and
Breitman ML: Role of the Flt-1 receptor tyrosine kinase in
regulating the assembly of vascular endothelium. Nature. 376:66–70.
1995. View Article : Google Scholar
|
|
32
|
Risau W: Mechanisms of angiogenesis.
Nature. 386:671–674. 1997. View Article : Google Scholar
|
|
33
|
Ziche M, Morbidelli L, Choudhuri R, Zhang
HT, Donnini S, Granger HJ and Bicknell R: Nitric oxide synthase
lies downstream from vascular endothelial growth factor-induced but
not basic fibroblast growth factor-induced angiogenesis. J Clin
Invest. 99:2625–2634. 1997. View Article : Google Scholar :
|
|
34
|
Jiang Y, Vaessen B, Lenvik T, Blackstad M,
Reyes M and Verfaillie CM: Multipotent progenitor cells can be
isolated from postnatal murine bone marrow, muscle, and brain. Exp
Hematol. 30:896–904. 2002. View Article : Google Scholar
|
|
35
|
Liu Z, Jiang Y, Hao H, Gupta K, Xu J, Chu
L, McFalls E, Zweier J, Verfaillie C and Bache RJ: Endothelial
nitric oxide synthase is dynamically expressed during bone marrow
stem cell differentiation into endothelial cells. Am J Physiol
Heart Circ Physiol. 293:H1760–H1765. 2007. View Article : Google Scholar
|
|
36
|
Xu J, Liu X, Jiang Y, Chu L, Hao H, Liua
Z, Verfaillie C, Zweier J, Gupta K and Liu Z: MAPK/ERK signalling
mediates VEGF-induced bone marrow stem cell differentiation into
endothelial cell. J Cell Mol Med. 12:2395–2406. 2008. View Article : Google Scholar :
|
|
37
|
Guo S, Cheng Y, Ma Y and Yang X:
Endothelial progenitor cells derived from CD34+ cells
form cooperative vascular networks. Cell Physiol Biochem.
26:679–688. 2010. View Article : Google Scholar
|
|
38
|
Gehling UM, Ergün S, Schumacher U, Wagener
C, Pantel K, Otte M, Schuch G, Schafhausen P, Mende T, Kilic N, et
al: In vitro differentiation of endothelial cells from
AC133-positive progenitor cells. Blood. 95:3106–3112. 2000.
|
|
39
|
Quirici N, Soligo D, Caneva L, Servida F,
Bossolasco P and Deliliers GL: Differentiation and expansion of
endothelial cells from human bone marrow CD133(+) cells. Br J
Haematol. 115:186–194. 2001. View Article : Google Scholar
|
|
40
|
Reyes M, Lund T, Lenvik T, Aguiar D,
Koodie L and Verfaillie CM: Purification and ex vivo expansion of
postnatal human marrow mesodermal progenitor cells. Blood.
98:2615–2625. 2001. View Article : Google Scholar
|
|
41
|
Horwitz EM, Gordon PL, Koo WK, Marx JC,
Neel MD, McNall RY, Muul L and Hofmann T: Isolated allogeneic bone
marrow-derived mesenchymal cells engraft and stimulate growth in
children with osteogenesis imperfecta: Implications for cell
therapy of bone. Proc Natl Acad Sci USA. 99:8932–8937. 2002.
View Article : Google Scholar :
|
|
42
|
Glaser DE, Turner WS, Madfis N, Wong L,
Zamora J, White N, Reyes S, Burns AB, Gopinathan A and McCloskey
KE: Multifactorial Optimizations for Directing Endothelial Fate
from Stem Cells. PLoS One. 11:e01666632016. View Article : Google Scholar :
|
|
43
|
Kinnaird T, Stabile E, Burnett MS, Lee CW,
Barr S, Fuchs S and Epstein SE: Marrow-derived stromal cells
express genes encoding a broad spectrum of arteriogenic cytokines
and promote in vitro and in vivo arteriogenesis through paracrine
mechanisms. Circ Res. 94:678–685. 2004. View Article : Google Scholar
|