Introduction
Cigarette smoking is an independent risk factor for
cardiovascular disease that can lead to intravascular thrombosis.
It is associated with significantly increased rates of acute
coronary syndrome and sudden cardiac death (1). Autopsy and clinical studies also
demonstrated that smoking is critical to arterial thrombosis via a
direct toxic effect on the injured endothelial cells, resulting in
alterations of the hemostatic system and an increase in platelet
reactivity that is relevant for the pathogenic effect of
atherothrombosis (2,3). Thrombosis is affected by the
anticoagulation and hemostatic systems. However, the mechanism by
which cigarette smoking causes thrombosis via the anticoagulant
system is not clear.
Thrombomodulin (TM) expressed on the surface of the
endothelial cells is critical for anticoagulation (4). Its major function is to form a
complex with thrombin at a 1:1 high affinity, which activates
protein C. Activated protein C is a natural anticoagulant that
interacts with the endothelial protein C receptor (EPCR) to enhance
the function of anticoagulation system. Functional deficiency of
the anticoagulation system could enhance thrombosis formation. As
observed, the endothelium-specific loss of TM in mice led to
spontaneous and fatal thrombosis in their arterial and venous
circulation (5). For human beings,
the downregulation of endothelial TM or EPCR expression in certain
pathologic conditions, including meningococcal sepsis and graft
rejection, may result in thrombotic complications (6,7).
Thus, it was hypothesized that cigarette smoking could reduce the
essential antithrombotic functions of endothelial cells by
inhibiting TM or EPCR expression.
Cigarette smoke comprises >5,600 distinct
components (8). Hence, it is
important to identify which specific components might be
responsible for the observed effects. Nicotine is an indispensable
constituent of cigarette smoke. Studies have demonstrated that
nicotine serves as a catalyst for endothelial and smooth muscle
cell proliferation, and angiogenesis, vascular inflammation and
enhancement of atherogenesis (9–12).
Therefore, it was hypothesized that nicotine may also contribute to
the development of thrombosis by inhibiting TM or EPCR
expression.
To evaluate these hypotheses, a novel study was
performed to examine the effects of cigarette smoke extract (CSE)
and nicotine on TM and EPCR expression in human umbilical vein
endothelial cells (HUVECs) in vitro.
Materials and methods
Materials
Nicotine was purchased from Sigma-Aldrich; Merck
KGaA (Darmstadt, Germany). The monoclonal antibodies for TM (cat
no. ab6980) and EPCR (cat no. ab56689) were obtained from Abcam
(Shanghai, China). Goat polyclonal anti-mouse immunoglobulin G
(IgG)-fluorescein isothiocyanate (FITC) (cat no. ab6785) was
purchased from Abcam. A fluorescent quantitation kit was purchased
from Takara Biotechnology Co., Ltd. (Dalian, China). Tumor necrosis
factor (TNF)-α was purchased from ProSpec (East Brunswick, NJ,
USA). The reagents collagenase I, medium 200 (M200) and low serum
growth supplement (LSGS) were bought from Gibco; Thermo Fisher
Scientific, Inc. (Waltham, MA, USA). Fetal bovine serum was
purchased from Hyclone; GE Healthcare (Chicago, IL, USA). TRIzol
reagent was purchased from Invitrogen; Thermo Fisher Scientific,
Inc. (Waltham, MA, USA). Factor VIII-related antibody (cat no.
ZA-0111) was purchased from Zhongshan Goldenbridge Biotechnology
Co., Ltd. (Beijing, China). Goat polyclonal anti-rabbit
immunoglobulin G (IgG)-streptavidin horseradish peroxidase (HRP;
cat no. 12–348) was purchased from EMD Millipore (Billerica, MA,
USA). Other reagents used in the experiments were all of analytical
grade.
CSE preparation
CSE was prepared using a previously reported method
with slight modification (13,14).
Two cigarettes [each containing 1.1 mg nicotine, 13 mg carbon
monoxide and 12 mg tar (without filters; Hongmei brand, Hongta
Tobacco Group, Yuxi, China)] were smoked consecutively through a
tea infusion using a 50-ml injection syringe with a constant
airflow. The smoke was bubbled through 50 ml phosphate
buffered-saline (PBS, 10 mM Na2HPO4, 1.8 mM
KH2PO4, 2.7 mM KCl, 140 mM NaCl, pH=7.4) controlled by a three-way
cock mimicking the gas fluid pathway during the smoking process in
the human lung. The obtained CSE solution was filtered through a
0.22-µm filter to remove bacterial and large particles. The pH of
the CSE-PBS buffer was adjusted to a final value of 7.4. The
concentration of this obtained stock solution was defined as 100%
CSE, and the solution was diluted using PBS to a final
concentration required in the experiment. The CSE solution was
prepared immediately prior to each experiment.
Cell culture
HUVECs were isolated using an enzymatic technique
(15). Umbilical vein was obtained
from the umbilical cord of a healthy baby born by caesarean section
in the Department of Gynecology and Obstetrics, the General
Hospital of Chinese People's Armed Police Forces (Beijing, China).
Mothers agreed and provided signed consent forms. Approval was
obtained from the hospital ethics committee. The veins were flushed
with PBS, filled with 0.1% collagenase I, clamped at either end,
and incubated in 95% air/5% CO2 at 37°C for 15–20 min.
The collagenase solution was subsequently decanted into a sterile
tube and centrifuged at 300 × g for 10 min at 37°C to produce an
enteric-coated pellet. The pellet was then resuspended in the low
serum endothelial cell medium (M200 + LSGS) in an atmosphere of 95%
air/5% CO2 at 37°C. The medium was replaced after 24 h,
and subsequently every 48 h; HUVECs were plated onto gelatin-coated
35-mm dishes (2×105 cells/ml) and incubated overnight to
produce a confluent monolayer of cells. The cells were used for the
experiment at passages 2–3. Factor VIII-related antibody was used
to identify endothelial cells by immunocytochemistry staining
(16). HUVECs were fixed in 4%
paraformaldehyde solution for 30 min at 4°C, washed in 0.1 M
glycine (2×5 min, 22°C), incubated with 3% hydrogen peroxide (5
min, 22°C), rinsed in 0.01 M PBS (6.8 mM
Na2HPO4, 2.6 mM
NaH2PO4, pH 7.2), and incubated with
heat-inactivated goat serum (1:20 in 0.01 M PBS, 1 h, 22°C). Cells
were incubated with Factor VIII-related antibody (1:100 dilution),
overnight at 22°C in a humidified chamber. Cells were washed in
0.01 M PBS (6×5 min, 22°C), incubated with goat polyclonal
anti-rabbit immunoglobulin G (IgG)-streptavidin HRP (1:200 in 0.01
M PBS; 1 h, 22°C). After a further 3×5 min washes in 0.01 M PBS,
cells were incubated with 0.1 M acetate buffer (pH 5.3; 3 min,
22°C), and 3-amino-9-ethylcarbazole solution for 3 min at 22°C for
detection of vWF. Cells were rinsed in distilled water. Coverslips
were mounted onto microscope slides using glycerol.
Photomicrographs were obtained using a Nikon DS-Fi2 camera
connected to a Nikon Eclipse Ti microscope.
The HUVECs were seeded into 6-well plates
(2×105 cells/ml). The cells were then grown in complete
medium for 48 h to 90% confluence, with a change of the medium, and
incubated with 0.5, 1, 3.5 or 5% CSE or 10-3 to 10-9 mol/l nicotine
for 0, 6, 10, 12 or 24 h. Control cells received only the fresh
medium. All experiments were performed in triplicate.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
The cells were washed with cold PBS twice, and total
RNA was extracted using TRIzol reagent. Briefly, the cells were
harvested and treated with 0.5 ml TRIzol reagent for 5 min at room
temperature and then centrifuged at 12,000 × g for 15 min at 4°C.
The aqueous portion containing RNA was transferred to another
RNAase-free Eppendorf tube, and 0.5 ml isopropanol was added and
incubated for 10 min at room temperature, followed by precipitating
total RNA by centrifugation at 7,500 × g at 4°C for 10 min. The
pellet was then washed with 75% ethanol twice and dried at room
temperature on ice for 5 min; total RNA pellet was dissolved in 20
µl H2O. The RNA concentration was measured at A260/A280
using the NanoVue spectrophotometer. cDNA was obtained from reverse
transcription of 1 µg total RNA in a 20-µl reaction volume using an
iScrip cDNA Synthesis kit (Bio-Rad Laboratories, Inc., Hercules,
CA, USA).
The primers for TM and β-actin were designed using
Beacon Designer 2.1 software (Premier Biosoft International, Palo
Alto, CA, USA) and synthesized using Sangon Biotech, Co., Ltd.
(Shanghai, China). The designed oligonucleotide sequences were as
follows: TM: 5′-CCCAACACCCAGGCTAGCT-3′ (forward),
5′-CGTCGATGTCCGTGCAGAT-3′ (reverse); EPCR: 5′-ATTGCTGCCGATACTGCT-3′
(forward), 5′-AGAGGAAAGGCCAAGGTC-3′ (reverse); β-actin:
5′-TCACCAACTGGGACGACA-3′ (forward), 5′-ACAGCCTGGATAGCAACG-3′
(reverse). qPCR was performed using a fluorescent quantitation kit
(Takara Biotechnology, Co., Ltd.). Briefly, 1 µl of the reaction
product of the RT reaction was used in a 25-µl PCR reaction. The
annealing temperature and primer concentration were adapted for
each gene. The amplification product was initially incubated at
95°C for 10 sec at 20°C, followed by 40 cycles of denaturation at
95°C for 5 sec, annealing at 45°C for 10 sec, extension at 72°C for
10 sec, and elongation at 80°C for 0.1 sec, using an PCR detection
system. Data were analyzed using LightCycler 2.0 software (Applied
Biosystems; Thermo Fisher Scientific, Inc.) and presented as the
relative amount of mRNA using the formula 2−∆∆Cq, which
stands for the difference in the quantitation cycle (Cq) between a
gene of interest and the housekeeping gene β-actin (17). The Cq is the point at which sample
fluorescence rises above the background level. Each sample was
measured in triplicate. mRNA (no reverse transcription) or
H2O (no DNA samples) were included as negative
controls.
Flow cytometry
The cells were harvested and isolated by papain and
centrifuged at 500 × g for 5 min at 10°C twice. They were then
incubated with anti-TM and monoclonal anti-EPCR antibodies (1:100
dilution) for 30 min at 4°C, washed twice with the PBS buffer, and
incubated with FITC-conjugated goat anti-mouse IgG (1:100 dilution)
in the PBS buffer at 4°C for 30 min. The cells were washed again
with the PBS buffer, and cell-bound fluorescence was determined by
flow cytometry, with 30,000 events/sample counted using a FACS
calibur cytofluorometer (BD Biosciences, Franklin Lakes, NJ, USA).
The data were analyzed using WinMDI 2.9 software (Scripps Research
Institute, La Jolla, CA, USA).
Statistical analysis
Data are expressed as the mean ± standard deviation
and statistically analyzed using one-way or two-way analysis of
variance as appropriate, followed by the Student-Newman-Keuls
method. SPSS 17.0 software (SPSS, Inc., Chicago, IL, USA) was used.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effects of CSE and nicotine on TM mRNA
expression in HUVECs
More than 95% of the endothelial cell cytoplasm
contained brown granules, confirming that the cultured cells were
HUVECs (data not shown). RT-qPCR was performed to study the effect
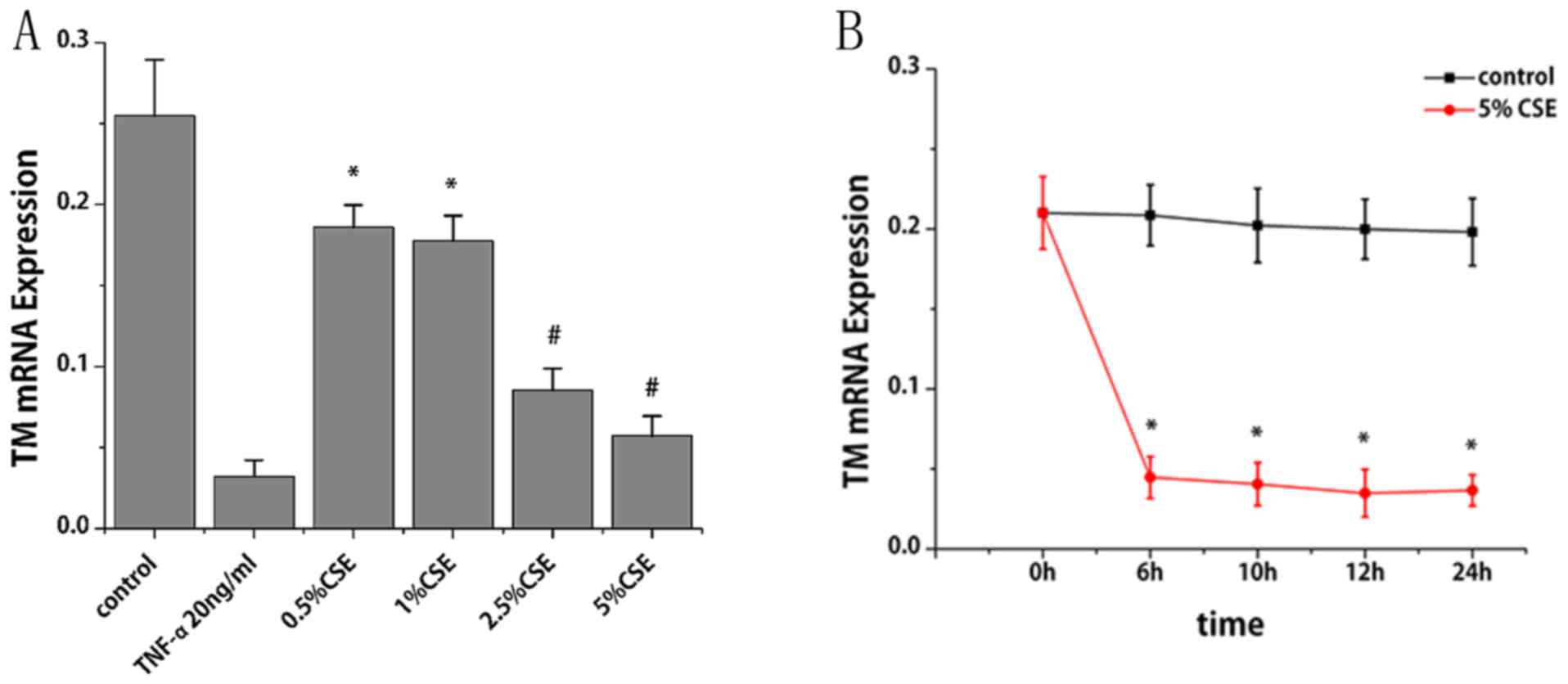
of CSE and nicotine on TM mRNA expression. CSE treatment for 6 h
led to a reduction TM mRNA expression in a dose-dependent manner.
Compared with the control group, TM mRNA levels decreased by 27,
30, 67 and 78% following treatment of CSE at 0.5, 1, 2.5 and 5%,
respectively (P<0.01; Fig. 1A).
With the treatment of CSE (5%), TM mRNA decreased by 79, 80, 83 and
82% at 6, 10, 12 and 24 h, respectively (P<0.001; Fig. 1B). Notably, TM mRNA expression
decreased progressively within 12 h; however, it slightly increased
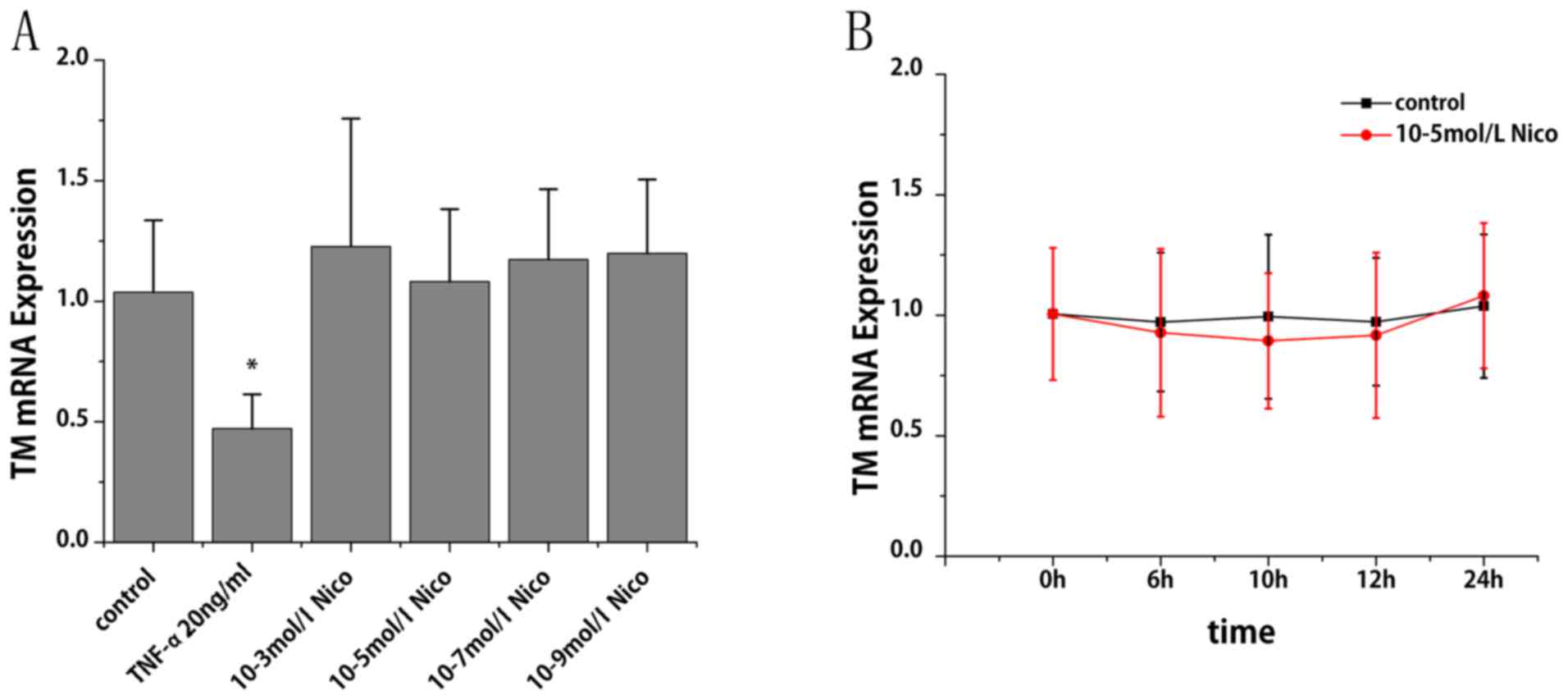
at 24 h compared with at 12 h. The specific constituent of
cigarette smoke that might be responsible for the observed
inhibition was subsequently investigated. As presented in Fig. 2A and B, nicotine at a dose ranging
from 10-3 to 10-9 mol/l did not cause any significant alterations
in TM mRNA expression.
Effects of CSE and nicotine on TM
protein expression levels in HUVECs
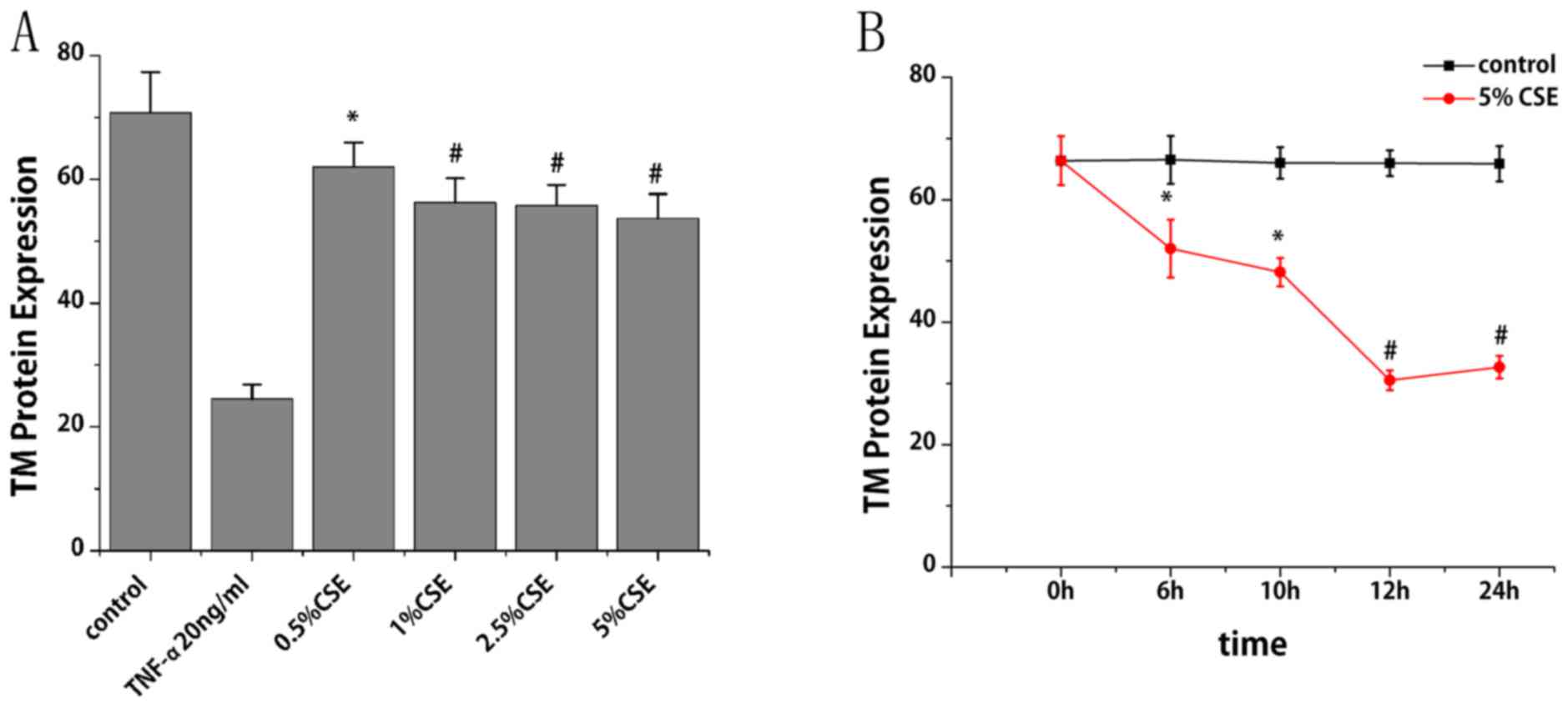
Flow cytometry was performed to study the effect of
CSE on TM protein expression. With the treatment of CSE (0.5, 1,
2.5 and 5%) for 6 h, TM protein expression levels reduced by 12,
21, 21 and 24%, respectively, compared with the control group
(P<0.05; Fig. 3A). With the
treatment of CSE (5%) for different times, TM protein expression
levels decreased by 22, 27, 54 and 50% at 6, 10, 12 and 24 h,
respectively, compared with the control group (P<0.05; Fig. 3B). TM protein expression levels
were also observed to decrease progressively within 12 h, and then
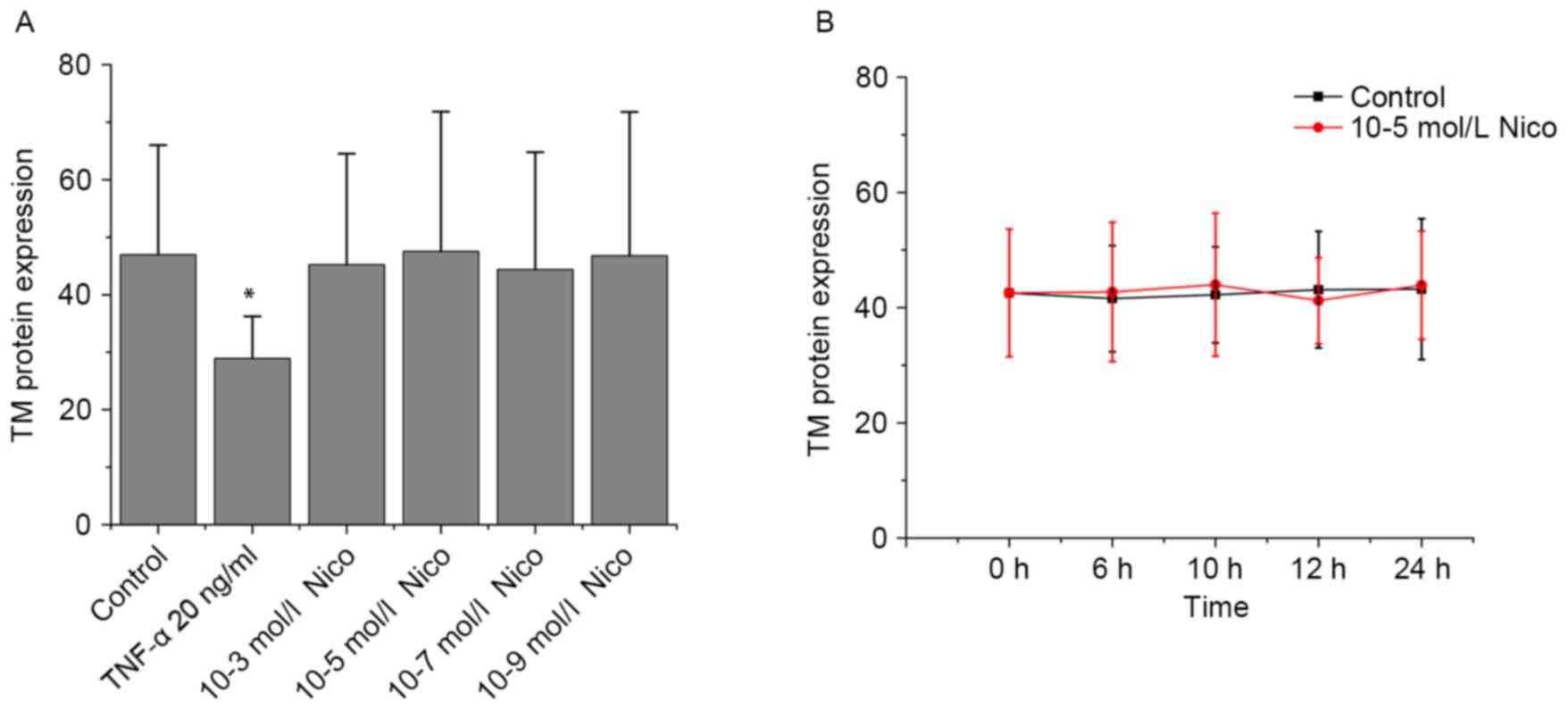
increase slightly at 24 h. As presented in Fig. 4A and B, nicotine did not induce any
significant alterations in TM protein expression.
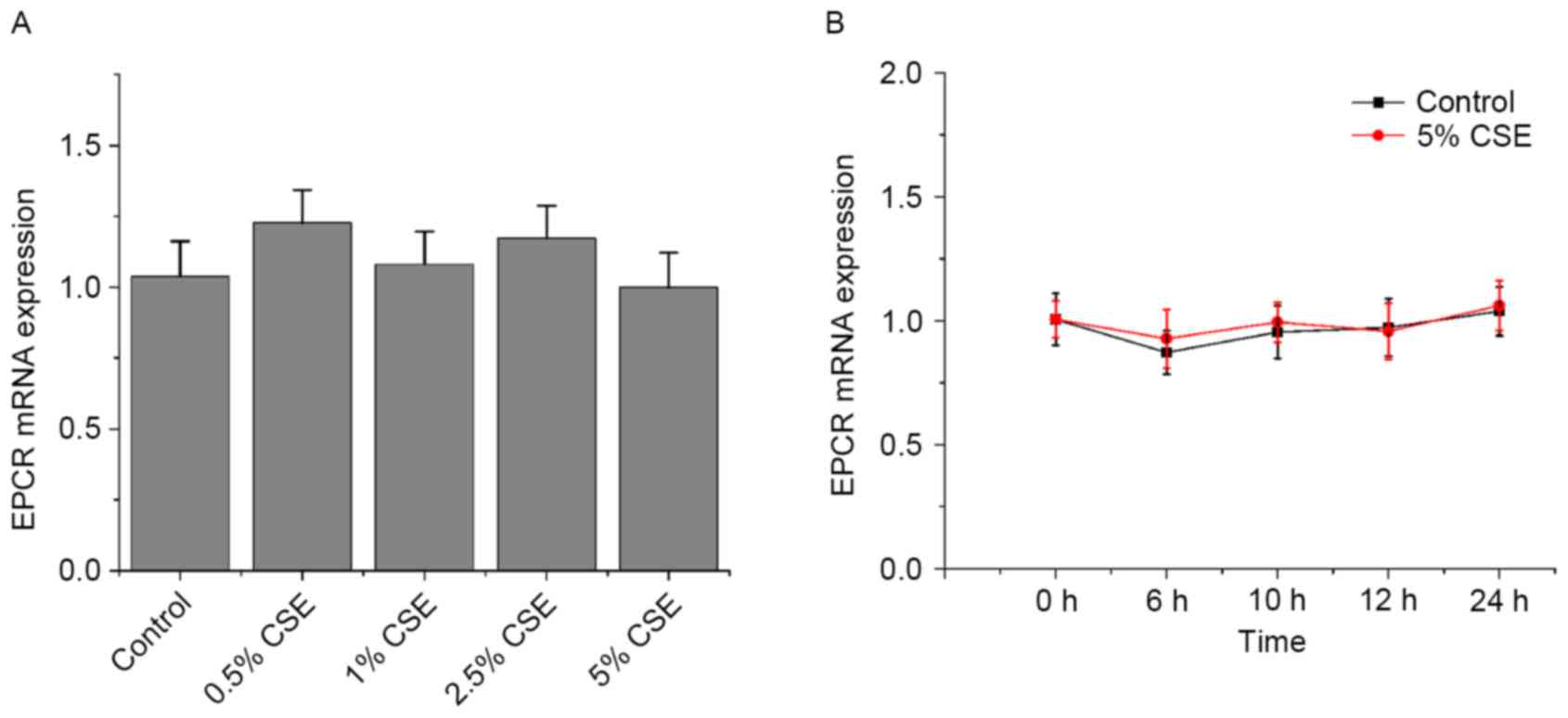
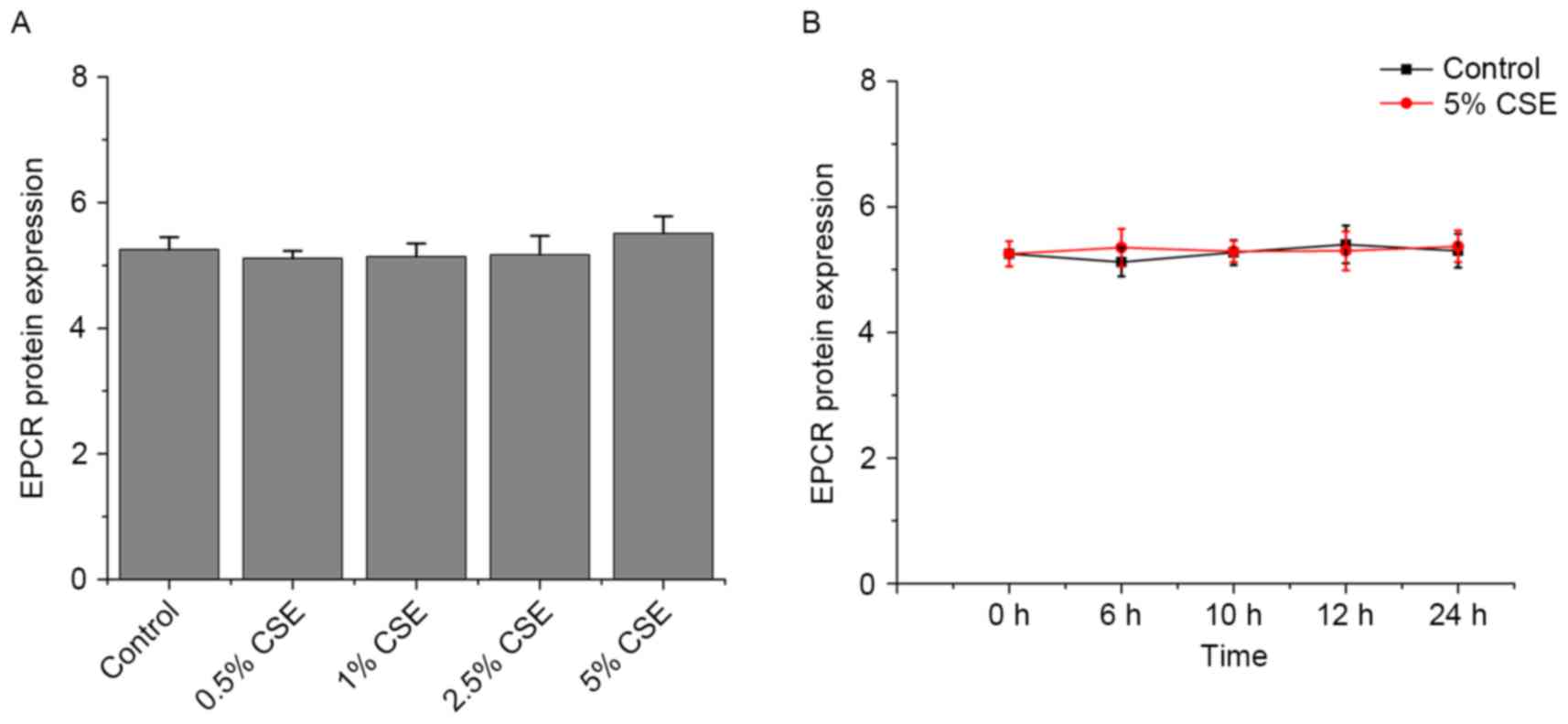
Effects of CSE and nicotine on EPCR
expression in HUVECs
As presented in Figs.
5 and 6, CSE and nicotine had
no effect on EPCR expression.
Discussion
The present study demonstrated that CSE
significantly decreased TM expression in a dose- and time-dependent
manner in HUVECs. Both mRNA and protein expression levels were
significantly reduced in CSE-treated HUVECs. The data from this
study suggested a novel molecular mechanism of cigarette
smoking-associated thrombosis by the decreased expression of TM.
However, CSE had no effect on EPCR.
Previous studies on the control of TM expression
have been performed in HUVECs, but studies on the influence of
cigarette smoke on TM and EPCR expression are lacking. Previous
studies have demonstrated that the expression of TM is inhibited by
inflammatory and procoagulant molecules and environmental factors,
including interleukin (IL)-1β, lipopolysaccharide, TNF-α,
C-reactive protein, oxidized low-density lipoprotein, transforming
growth factor (TGF)-β1, TGF-β2, irradiation and shear (18–20).
TM expression was increased by anti-inflammatory and anticoagulant
molecules and environmental factors, including retinoic acid,
cyclic adenosine monophosphate, IL-4, prostacyclin, okadaic acid,
curcumin, vascular endothelial growth factor, prostaglandin E1,
statins, histamine and heat shock (21–23).
Cigarette smoking is associated with oxidative stress, increased
blood thrombogenicity and an inflammatory response. In humans,
cigarette smoking is associated with increased levels of multiple
inflammatory and procoagulant molecules, such as TNF-α, peripheral
leukocytes, C-reactive protein, IL-6 and homocysteine (24–26).
Furthermore, cigarette smoke can directly provide free radicals to
induce oxidative stress. The present study examined the damaging
effect of cigarette smoke on TM expression. It was earlier observed
that CSE could notably reduce the binding probability of TM and
thrombin (27). EPCR can be
downregulated at the transcriptional level by shear stress and
inflammation to kinases, including IL-1α and TNF-α (28,29).
However, CSE had no effect on EPCR.
Cigarette smoke comprises >5,600 distinct
components, of which >200 have been identified as carcinogens
and respiratory toxins. The present study investigated whether
nicotine was responsible for inhibiting TM and EPCR expression.
Although nicotine is the well-recognized component of cigarette
smoke, it had no effect on TM and EPCR expression in HUVECs. Cornel
et al (30) observed that
Korean teenage and adult smokers had 0.27 and 0.5 µM plasma
nicotine concentrations, respectively. Al Mutairi et al
(31) reported a nicotine
concentration of up to 13 µM in Kuwaiti smokers. However, the
highest concentration tested in this study was ~three times greater
than the reported blood levels in smokers. Nicotine still did not
induce any change in TM and EPCR expression in HUVECs. Previous
studies have demonstrated that the effect of nicotine on
thrombohemostatic factors, including platelets, fibrinogen, tissue
plasminogen activator and plasminogen activator inhibitor-1, is
small and probably serves only a minor role in atherothrombotic
events directly (32–34).
The present study demonstrated following CSE
treatment, TM expression decreased progressively within 12 h;
however, it increased slightly at 24 h. It is possible that the
effect of CSE on TM expression reached saturation after 12 h, or a
part of soluble substances in CSE volatilized over time, weakening
the effect of CSE on TM expression. In addition, the decrease in TM
mRNA expression was more obvious than the decrease in TM protein
expression at 6 h. It is possible that CSE acts at the
transcriptional level, leading to the downregulation of TM protein.
The present study had some limitations. The insoluble and volatile
harmful ingredients in tobacco were not included in the study, and
HUVECs were the only cell model used.
In conclusion, the present study demonstrated that
exposure to CSE (except nicotine) significantly decreased the
expression of TM in HUVECs in a concentration-dependent manner. The
results of the present study support a noevl molecular mechanism of
cigarette smoking-associated thrombosis by the decreased expression
of TM. Further studies are required to investigate the distinct
components in CSE responsible for decreasing TM expression and its
associated consequences.
Acknowledgements
The present study was supported by the Nature
Science Foundation of China (grant no. 81370315).
References
|
1
|
Dunbar A, Gotsis W and Frishman W:
Second-hand tobacco smoke and cardiovascular disease risk: An
epidemiological review. Cardiol Rev. 21:94–100. 2014. View Article : Google Scholar
|
|
2
|
Szpak D, Grochowalski A, Chrzaszcz R,
Florek E, Jawień W and Undas A: Tobacco smoke exposure and
endothelial dysfunction in patients with advanced coronary artery
disease. Pol Arch Med Wewn. 123:474–481. 2013.
|
|
3
|
Barua RS and Ambrose JA: Mechanisms of
coronary thrombosis in cigarette smoke exposure. Arterioscler
Thromb Vasc Biol. 33:1460–1467. 2013. View Article : Google Scholar
|
|
4
|
Ito T and Maruyama I: Thrombomodulin:
Protectorate God of the vasculature in thrombosis and inflammation.
J Thromb Haemost. 9 Suppl 1:S168–S173. 2011. View Article : Google Scholar
|
|
5
|
Isermann B, Hendrickson SB, Zogg M, Wing
M, Cummiskey M, Kisanuki YY, Yanagisawa M and Weiler H:
Endothelium-specific loss of murine thrombomodulin disrupts the
protein C anticoagulant pathway and causes juvenile-onset
thrombosis. J Clin Invest. 108:537–546. 2001. View Article : Google Scholar :
|
|
6
|
Faust SN, Levin M, Harrison OB, Goldin RD,
Lockhart MS, Kondaveeti S, Laszik Z, Esmon CT and Heyderman RS:
Dysfunction of endothelial protein C activation in severe
meningococcal sepsis. N Engl J Med. 345:408–416. 2001. View Article : Google Scholar
|
|
7
|
Iino S, Abeyama K, Kawahara KI, Aikou T
and Maruyama I: Thrombomodulin expression on Langerhans' islet: Can
endogenous ‘anticoagulant on demand’ overcome detrimental
thrombotic complications in clinical islet transplantation. J
Thromb Haemost. 2(833–844): 20042011.
|
|
8
|
Perfett TA and Rodgman A: The complexity
of tobacco and tobacco smoke. Beitr Tabakforsch Int. 24:215–232.
2011.
|
|
9
|
Li JM, Cui TX, Shiuchi T, Liu HW, Min LJ,
Okumura M, Jinno T, Wu L, Iwai M and Horiuchi M: Nicotine enhances
angiotensin II-induced mitogenic response in vascular smooth muscle
cells and fibroblasts. Arterioscler Thromb Vasc Biol. 24:80–84.
2004. View Article : Google Scholar
|
|
10
|
Lee J and Cooke JP: Nicotine and
pathological angiogenesis. Life Sci. 91:1058–1064. 2012. View Article : Google Scholar :
|
|
11
|
Heeschen C, Weis M and Cooke JP: Nicotine
promotes arteriogenesis. J Am Coll Cardiol. 41:489–496. 2003.
View Article : Google Scholar
|
|
12
|
Lau PP, Li L, Merched AJ, Zhang AL, Ko KW
and Chan L: Nicotine induces proinflammatory responses in
macrophages and the aorta leading to acceleration of
atherosclerosis in low-density lipoprotein receptor(−/-) mice.
Arterioscler Thromb Vasc Biol. 26:143–149. 2006. View Article : Google Scholar
|
|
13
|
Chen HW, Chien ML, Chaung YH, Lii CK and
Wang TS: Extracts from cigarette smoke induce DNA damage and cell
adhesion molecule expression through different pathways. Chem Biol
Interact. 150:233–241. 2004. View Article : Google Scholar
|
|
14
|
Su Y, Han W, Giraldo C, De Li Y and Block
ER: Effect of cigarette smoke extract on nitric oxide synthase in
pulmonary artery endothelial cells. Am J Respir Cell Mol Biol.
19:819–825. 1998. View Article : Google Scholar
|
|
15
|
Li Y, Shi XL, Liu HL, Yi SQ, Zhang XJ and
Fang XH: Study of the effect of atorvastatin on the interaction
between ICAM-1 and CD11b by live-cell single-molecule force
spectroscopy. Science China (Chemistry). 53:752–758. 2010.
View Article : Google Scholar
|
|
16
|
Bürgin-Maunder CS, Brooks PR and Russell
FD: Omega-3 fatty acids modulate weibel-palade body degranulation
and actin cytoskeleton rearrangement in PMA-stimulated human
umbilical vein endothelial cells. Mar Drugs. 11:4435–4450. 2013.
View Article : Google Scholar :
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
18
|
Nan B, Lin P, Lumsden AB, Yao Q and Chen
C: Effects of TNF-alpha and curcumin on the expression of
thrombomodulin and endothelial protein C receptor in human
endothelial cells. Thromb Res. 115:417–426. 2005. View Article : Google Scholar
|
|
19
|
Nan B, Yang H, Yan S, Lin PH, Lumsden AB,
Yao Q and Chen C: C-reactive protein decreases expression of
thrombomodulin and endothelial protein C receptor in human
endothelial cells. Surgery. 138:212–222. 2005. View Article : Google Scholar
|
|
20
|
Conway EM: Thrombomodulin and its role in
inflammation. Semin Immunopathol. 34:107–125. 2012. View Article : Google Scholar
|
|
21
|
Masamura K, Oida K, Kanehara H, Suzuki J,
Horie S, Ishii H and Miyamori I: Pitavastatin-induced
thrombomodulin expression by endothelial cells acts via inhibition
of small G proteins of the Rho family. Arterioscler Thromb Vasc
Biol. 23:512–517. 2003. View Article : Google Scholar
|
|
22
|
Weiler-Guettler H, Yu K, Soff G, Gudas LJ
and Rosenberg RD: Thrombomodulin gene regulation by cAMP and
retinoic acid in F9 embryonal carcinoma cells. Proc Natl Acad Sci
USA. 89:2155–2159. 1992. View Article : Google Scholar :
|
|
23
|
Morser J: Thrombomodulin links coagulation
to inflammation and immunity. Curr Drug Targets. 13:421–431. 2012.
View Article : Google Scholar
|
|
24
|
Tracy RP, Psaty BM, Macy E, Bovill EG,
Cushman M, Cornell ES and Kuller LH: Lifetime smoking exposure
affects the association of C-reactive protein with cardiovascular
disease risk factors and subclinical disease in healthy elderly
subjects. Arterioscler Thromb Vasc Biol. 17:2167–2176. 1997.
View Article : Google Scholar
|
|
25
|
Bermudez EA, Rifai N, Buring JE, Manson JE
and Ridker PM: Relation between markers of systemic vascular
inflammation and smoking in women. Am J Cardiol. 89:1117–1119.
2002. View Article : Google Scholar
|
|
26
|
Price JF, Mowbray PI, Lee AJ, Rumley A,
Lowe GD and Fowkes FG: Relationship between smoking and
cardiovascular risk factors in the development of peripheral
arterial disease and coronary artery disease: Edinburgh Artery
Study. Eur Heart J. 20:344–353. 1999. View Article : Google Scholar
|
|
27
|
Wei Y, Zhang X, Xu L, Yi S, Li Y, Fang X
and Liu H: The effect of cigarette smoke extract on
thrombomodulin-thrombin binding: An atomic force microscopy study.
Sci China Life Sci. 55:891–897. 2012. View Article : Google Scholar
|
|
28
|
Fukudome K and Esmon CT: Identification,
cloning, and regulation of a novel endothelial cell protein
C/activated protein C receptor. J Biol Chem. 269:26486–26491.
1994.
|
|
29
|
Jun P, Huangqing C, Xiaoheng L, Ruheng L
and Xiaohong Z: Effects of shear stress on protein C activation,
EPCR expression and TM expression in endothelial cells. Sheng Wu Yi
Xue Gong Cheng Xue Za Zhi. 26:303–309. 2009.
|
|
30
|
Cornel JH, Becker RC, Goodman SG, Husted
S, Katus H, Santoso A, Steg G, Storey RF, Vintila M, Sun JL, et al:
Prior smoking status, clinical outcomes, and the comparison of
ticagrelor with clopidogrel in acute coronary syndromes-insights
from the PLATelet inhibition and patient Outcomes (PLATO) trial. Am
Heart J. 164:334–342.e1. 2012. View Article : Google Scholar
|
|
31
|
Al Mutairi SS, Shihab-Eldeen AA,
Mojiminiyi OA and Anwar AA: Comparative analysis of the effects of
hubble-bubble (Sheesha) and cigarette smoking on respiratory and
metabolic parameters in hubble-bubble and cigarette smokers.
Respirology. 11:449–455. 2006. View Article : Google Scholar
|
|
32
|
Ambrose JA and Barua R: The
pathophysiology of cigarette smoking and cardiovascular disease: An
update. J Am Coll Cardiol. 43:1731–1737. 2004. View Article : Google Scholar
|
|
33
|
Smith CJ and Fischer TH: Particulate and
vapor phase constituents of cigarette mainstream smoke and risk of
myocardial infarction. Atherosclerosis. 158:257–267. 2011.
View Article : Google Scholar
|
|
34
|
Yang YM and Liu GT: Damaging effect of
cigarette smoke extract on primary cultured human umbilical vein
endothelial cells and its mechanism. Biomed Environ Sci.
17:121–134. 2004.
|