Introduction
Although great efforts have been made to improve the
diagnosis and treatment of tongue cancer, it remains a difficult
disease to cure with a five-year-survival rate of 50% (1). Multiple therapeutics including
surgery, radiotherapy and chemotherapy were applied to advanced
tongue cancer in clinic; nevertheless, little improvements were
achieved in the past few decades (2). Tongue cancer is more common in older
people, but its incidence rate is higher than that of other head
and neck cancers even in young people, greatly due to its various
risk factors, such as certain environment factors, alcohol intake
and even genetic factors (3).
With the rapid development of genome sequencing
technologies, the classic view of the transcriptome landscape has
undergone a fundamental change (4). It was now well established that more
than 90% of the genome can be transcribed with only less than 2%
being subsequently translated, which means that the vast majority
of genome serves as the template for the transcription of noncoding
RNAs (ncRNAs) (5,6). Long noncoding RNAs (lncRNAs) are a
newly emerged class of noncoding RNA containing more than 200
nucleotides that are widely transcribed in the genome (7). Unlike other noncoding RNAs, lncRNAs
involvement in human diseases is largely unclear. Current evidence
has implicated that lncRNAs may widely participate in multiple
intracellular and extracellular activities, including gene
transcription, mRNA splice and tumorigenesis (8). Multiple lncRNAs have been shown to
play significant roles in regulating the process of human tongue
cancer. For instance, the lncRNA MALAT1 was found to interact with
miR-124 and modulate cell growth in human tongue cancer (9). LncRNA HOTTIP was upregulated in human
tongue squamous cell carcinoma and its expression correlated with
tumor sizing and distant metastasis (10).
The epidermal growth factor receptor (EGFR) is a
transmembrane protein, which is a receptor for members of the
epidermal growth factor family (EGF family) of extracellular
protein ligands (11,12). Mutations that lead to EGFR
overexpression or overactivity have been reported in multiple
cancers, including squamous-cell carcinoma of lung (more than 80%
cases) (13), anal cancers
(14) and epithelial tumors of the
head and neck (80–100% cases) (15). Particularly, EGFR played a
prognostic role in the prognosis of tongue cancer (16). Therefore, it is a high priority to
uncover the upstream signaling pathway of EGFR and to identify ways
to decrease the expression of EGFR from the original source.
In the present study, we investigated the role of a
newly-discovered LncRNA, Lnc-EGFR in human tongue cancer. To this
end, a total of 50 tongue cancer patients and four tongue cancer
cell lines were used. Cell proliferation and cell apoptosis were
detected to examine effects of Lnc-EGFR on tongue cancer
proliferation and apoptosis, respectively. Tongue cancer cell lines
UM1 and CAL-27 were transfected with specific shRNAs targeting
Lnc-EGFR (shLnc-EGFR) with or without the presence of recombinant
EGFR protein. Our study is the first to uncover the role of
Lnc-EGFR in tongue cancer. Our data might provide novel clues for
the diagnosis and treatment of tongue cancer patients in
clinic.
Materials and methods
Human tissues
A total of 50 tongue cancer tissues from patients
who were admitted to the Department of Orthodontics, the First
Affiliated Hospital of Harbin Medical University between May 2015
and May 2016 (age range, 30–70 years; mean age, 55 years; male:
female=31: 19) were obtained via surgical resection. Their adjacent
non-cancerous tissues were also dissected from each patient. All
tissues were frozen into liquid nitrogen immediately after
dissection and then stored at −80°C till use. All patients showed
their full intention to participate in the study and written
consent forms were obtained from each patient. The present study
was approved by the ethics committee of The First Affiliated
Hospital of Harbin Medical University.
Cell culture and shRNAs
transfection
Control cells CRL-7421 and tongue cancer cell line
SCC-25 were commercially purchased from American Type Tissue
Collection (ATCC, Massachusetts, USA). Tongue cancer cell lines
HSC-3, UM1 and CAL-27 were purchased from the Cell Bank of Chinese
Academy of Sciences (Shanghai, China). All of the cell lines were
cultured in dulbecco's modified eagle medium (DMEM; Gibco, Grand
Island, NY, USA) supplied with 10% fetal bovine serum (FBS; Gibco).
The cells grew in a 37°C incubator with 5% CO2 and the
culture medium was replaced every other day unless otherwise
stated. The shRNAs against Lnc-EGFR were synthesized by Genepharm.
Co., (Shanghai, China) and the sequences were listed in Table I. The recombinant human EGFR
protein was purchased from Abcam Co., (ab155639; NY, USA) and used
in a final concentration of 10 µM. The transfections were performed
using Lipofectamine 2000 (Invitrogen, NY, USA) according to the
manufactures' instructions in a dose of 2.5 µl for 1.5 µg DNA. Six
h after transfection, the culture medium was replaced with fresh
DMEM supplemented with 10% FBS.
 | Table I.Sequences of the primers used in
reverse transcription-quantitative polymerase chain reaction and
the sequences of shRNAs against Lnc-EGFR. |
Table I.
Sequences of the primers used in
reverse transcription-quantitative polymerase chain reaction and
the sequences of shRNAs against Lnc-EGFR.
| Gene | Primer nucleotide
sequences |
|---|
| Lnc-EGFR |
|
|
Forward |
5′-CAGCAGCCCTGCAATTAAAC-3′ |
|
Reverse |
5′-GGGTCCTCATGTAATGGTAATAGG-3′ |
| EGFR |
|
|
Forward |
5′-AGGCACGAGTAACAAGCTCAC-3′ |
|
Reverse |
5′-ATGAGGACATAACCAGCCACC-3′ |
| GAPDH |
|
|
Forward |
5′-GTGGACATCCGCAAAGAC-3′ |
|
Reverse |
5′-AAAGGGTGTAACGCAACTA-3′ |
| shLnc-EGFR-1 |
5′-GCTCTGCTTTAGTCAGGGT-3′ |
| shLnc-EGFR-2 |
5′-TACATGCCATCCTGGCCAT-3′ |
RNA isolation and RT-PCR
Total RNAs from clinical tissues and cultured cells
were extracted with TriZol® reagent (Takara Bio, Inc.,
Otsu, Japan) in a dilution of 0.5 ml for each well in a 12-well
plate. The RNA quality and quantity were determined by Nanodrop
2000 (ThermoFisher Scientific, Inc., NY, USA). Reverse
transcription (RT) of first-strand cDNAs was performed with
PrimeScript RT Master Mix (Perfect Real Time; Takara) following the
manufacturer's protocol. All PCR reactions were performed in an ABI
PRISM 7900 Real-Time system (Thermo Fisher Scientific, Inc.) with
the SYBR® Premix Ex Taq™ kit (Takara Bio, Inc.). The
thermocycling protocol was shown as follows: Initial denaturation
at 95°C for 2 min, followed by 35 repeats of the three-step cycling
program consisting of 30 sec at 95°C (denaturation), 1 min at 53°C
(primer annealing) and 30 sec at 72°C (elongation), followed by a
final extension step for 10 min at 72°C. The housekeeping gene
GAPDH was included as an internal control. Primer sequences were
listed in Table I. All
quantitative data were normalized to GAPDH using the
2−ΔΔCt method (17).
Western blot analysis
Briefly, total proteins from human tissues and cells
were collected by lysis buffer (RIPA, Beyotime, Nantong, China) on
ice and quantified using Bio-Rad protein assay reagent (Beyotime).
Equal amounts of protein (40 µg) were loaded onto 12% sodium
dodecyl sulfate-polyacrylamide gel (SDS-PAGE) and transferred to a
0.22 µm nitrocellulose membrane (NC, Millipore, MA, USA). The
membrane was blocked for 1 h with 5% skimmed milk at room
temperature and then incubated with primary antibodies overnight at
4°C. The primary antibodies against EGFR (SAB5500096, 1:1,000) was
purchased from Sigma Co. (NY, USA) and the primary antibody against
Tublin was from Santa Cruz Biotech (Santa Cruz, CA, USA). After
washing with TBST for 4 times (8 min each time), the membrane was
then incubated with secondary goat anti-rabbit antibody (sc-2004;
Santa Cruz Biotech) for 1 h at 37°C with a dilution of 1:1,000.
Finally, the proteins were quantified using ECL Prime Western
Blotting Detection reagent (GE Healthcare, Parsippany, NJ, USA) and
an ImageQuant LAS 4000 Mini Biomolecular Imager (GE
Healthcare).
Colony formation assay
UM1 and CAL-27 cells were transfected with
shLnc-EGFR or control shRNAs (shNC) with or without the presence of
EGFR recombinant protein in six-well plates with a density of 200
cells/well, during which the culture medium was not changed. After
2 weeks in 37°C incubator, the cell colonies that contained more
than 50 cells were counted by staining with crystal violet (0.5%)
for 10 min at room temperature and observed under a light
microscope with a magnification of 200 (Nikon, Japan).
Cell proliferation assay
Both UM1 and CAL-27 cells were seeded in a 96-well
plate at a concentration of 1,000 cells/well. After incubation for
24 h, cells were transfected with shLnc-EGFR or control shRNAs
(shNC) with or without the presence of EGFR recombinant protein.
Cell viability was monitored in a consecutive 5 days with a
CellTiter 96 AQueous Non-Radioactive Cell Proliferation kit
(Promega Corporation, Madison, WI, USA) as per the manufacturer's
protocols. The cell viability was determined by collecting the
absorbance at 490 nm using a microplate reader (Tecan, Männedorf,
Switzerland).
Flow cytometric analysis of cell
apoptosis
The annexin V/PI assay was performed as per the
manufacturer's instructions (Invitrogen). Briefly, UM1 and CAL-27
cells were plated into 6-well plates and transfected with
shLnc-EGFR or control shRNAs (shNC) with or without the presence of
EGFR recombinant protein for 48 h. Afterwards, cells were washed
with pre-cold PBS, trypsinized and re-suspended in 100 µl of
binding buffer with 2.5 µl FITC conjugated annexin-v and 1 µl PI
(100 µg/ml). Afterwards, cells were incubated at room temperature
for 15 min in darkness. A total of at least 10, 000 cells were
collected and calculated by flow cytometry for both cell lines (BD
Biosciences, San Diego, CA, USA).
Determination of caspase
activities
The activities of caspase-3, caspase-8 and caspase-9
were determined by the caspase activity kits (Beyotime) based on
the instructions. Briefly, cells were transfected with shRNAs for
48 h. Afterwards, cell lysates were collected by low speed
centrifuge (1,000 g, 5 min, 4°C). An equal amount of 10 µl proteins
from each sample was added into 96-well plates and mixed with an
aliquot of 80 µl reaction buffer supplied with caspase substrates
(2 mM). After incubated at 37°C for 4 h, caspase activities were
determined by the TECAN reader at an absorbance wavelength of 450
nm.
Immunofluorescence staining
Briefly, cells were cultured on a coverslip in
six-well plates at a density of 10,000 cells/well and then fixed
with cold acetone on ice for 20 min. After washed with PBS, cells
were blocked with normal goat serum for 10 min supplied with 0.1%
Triton X-10 and then incubated with primary antibody against EGFR
(1:250) at 37°C for 1 h and then overnight at 4°C. Secondary
antibodies (Dylight 549) were purchased from Dylight (Abcam) and
used in a dilution of 1:1,000 at 37°C for 1 h. The photos were
taken with a Nikon camera.
Statistical analysis
All experiments were repeated at least three times
in triplicate to obtain reproducible results. All data were
presented as mean ± standard deviation (SD). Student's t-test
analysis was used for the comparison between two groups with
Microsoft Excel 2007. Two-way ANOVA analysis was included for the
comparisons among three or more groups. Data were analyzed with
GraphPad Prism 6 (GraphPad Software Inc., San Diego, CA, USA). Any
value of P<0.05 was considered statistically significant.
Results
Expression of EGFR and long noncoding
RNA Lnc-EGFR were upregulated in human tongue cancer
In the present study, a total of 50 tongue cancer
patients were involved and the transcript level of Lnc-EGFR was
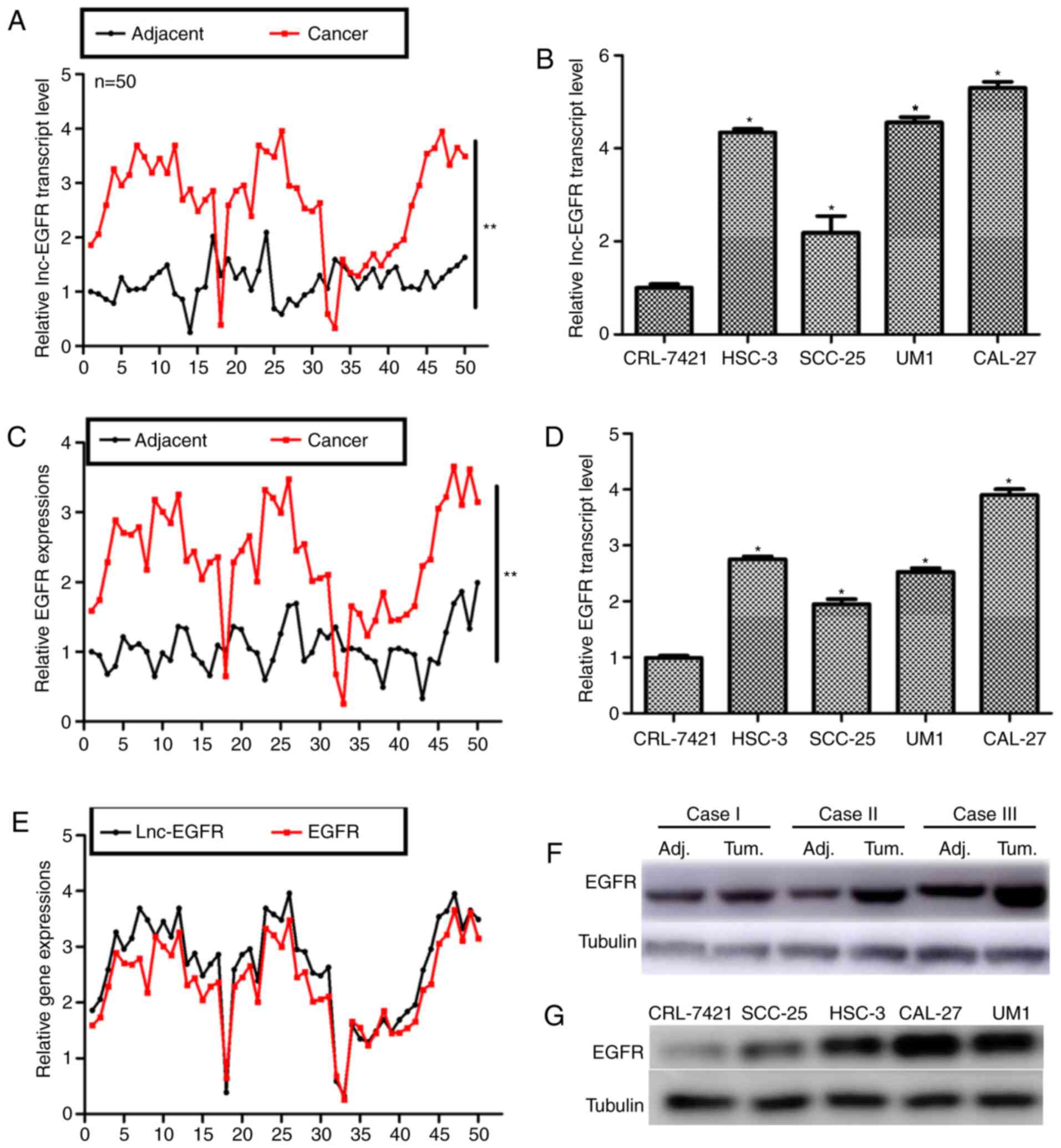
assessed. As shown in Fig. 1A,
only 3 of the 50 tongue cancer patients showed lower expression of
Lnc-EGFR as compared with the adjacent non-cancerous tissues
(P<0.01). Next, four tongue cancer cell lines were cultured and
subjected for RT-PCR analysis. All of the four tongue cancer cell
lines exhibited higher transcript levels of Lnc-EGFR as compared
with the control CRL-7421 cell. Particularly, UM1 and CAL-27 showed
the highest expressions of Lnc-EGFR (Fig. 1B). Thus, these two cell lines were
selected for subsequent functional analysis. Clinical
characteristics were also analyzed in Table II. The patients were divided into
two categories based on the expression of Lnc-EGFR: high level of
Lnc-EGFR (n=16) denotes those with transcript levels of Lnc-EGFR
higher than its median and low level of Lnc-EGFR (n=34) denotes
those with transcript level of Lnc-EGFR lower than the median
level. Among the examined variables (age, sex, tumor size, lymph
node metastasis, distant metastasis and TNM stage), the expression
of Lnc-EGFR was only associated with tongue tumor size
(P<0.001). Therefore, the effects of Lnc-EGFR on cell
proliferation and apoptosis were thereafter explored.
| Table II.Association of Lnc-EGFR with clinical
variables among 50 tongue cancer patients. |
Table II.
Association of Lnc-EGFR with clinical
variables among 50 tongue cancer patients.
|
|
| Expression of
Lnc-EGFR |
|
|---|
|
|
|
|
|
|---|
| Variable | Numbers | Low (n=34) | High (n=16) | P-value |
|---|
| Age (year) |
|
|
| 0.47 |
|
<40 | 8 | 7 | 1 |
|
|
40-50 | 16 | 10 | 6 |
|
|
>50 | 26 | 17 | 9 |
|
| Sex |
|
|
| 0.228 |
| Male | 31 | 19 | 12 |
|
|
Female | 19 | 15 | 4 |
|
| Tumor size (T) |
|
|
|
<0.001a |
| T1 and T2
(≤4 cm) | 39 | 32 | 7 |
|
| T3 and T4
(>4 cm or any size with distant metastasis) | 11 | 2 | 9 |
|
| Lymph node metastasis
(N) |
|
|
| 0.191 |
| N0 | 35 | 26 | 9 |
|
| N1 or
above | 15 | 8 | 7 |
|
| Distant metastasis
(M) |
|
|
| 0.486 |
| M0 | 38 | 27 | 11 |
|
| M1 | 12 | 7 | 5 |
|
| TNM stage |
|
|
| 0.103 |
| I/II | 34 | 26 | 8 |
|
|
III/IV | 16 | 8 | 8 |
|
Expression of EGFR was also detected with RT-PCR and
western blot analysis, since Lnc-EGFR was predicted as a potential
enhancer of EGFR (18). It was
shown in Fig. 1C and D, the
transcription levels of EGFR were significantly increased in both
clinical tongue cancer tissues and in cultured tongue cancer cells.
The association of Lnc-EGFR and EGFR was also analyzed in clinical
tissues (Fig. 1E). Furthermore,
the protein level of EGFR was also upregulated in 3 randomly
selected tongue cancer patients as depicted in Fig. 1F. Total proteins were also
extracted from cultured cells and it was shown that the expression
of EGFR was remarkably higher in all of the four tongue cancer cell
lines (Fig. 1G). All of these
results suggested that in parallel to EGFR, Lnc-EGFR was notably
increased in human tongue cancer.
Knockdown of Lnc-EGFR inhibited cell
proliferation in human tongue cancer cells
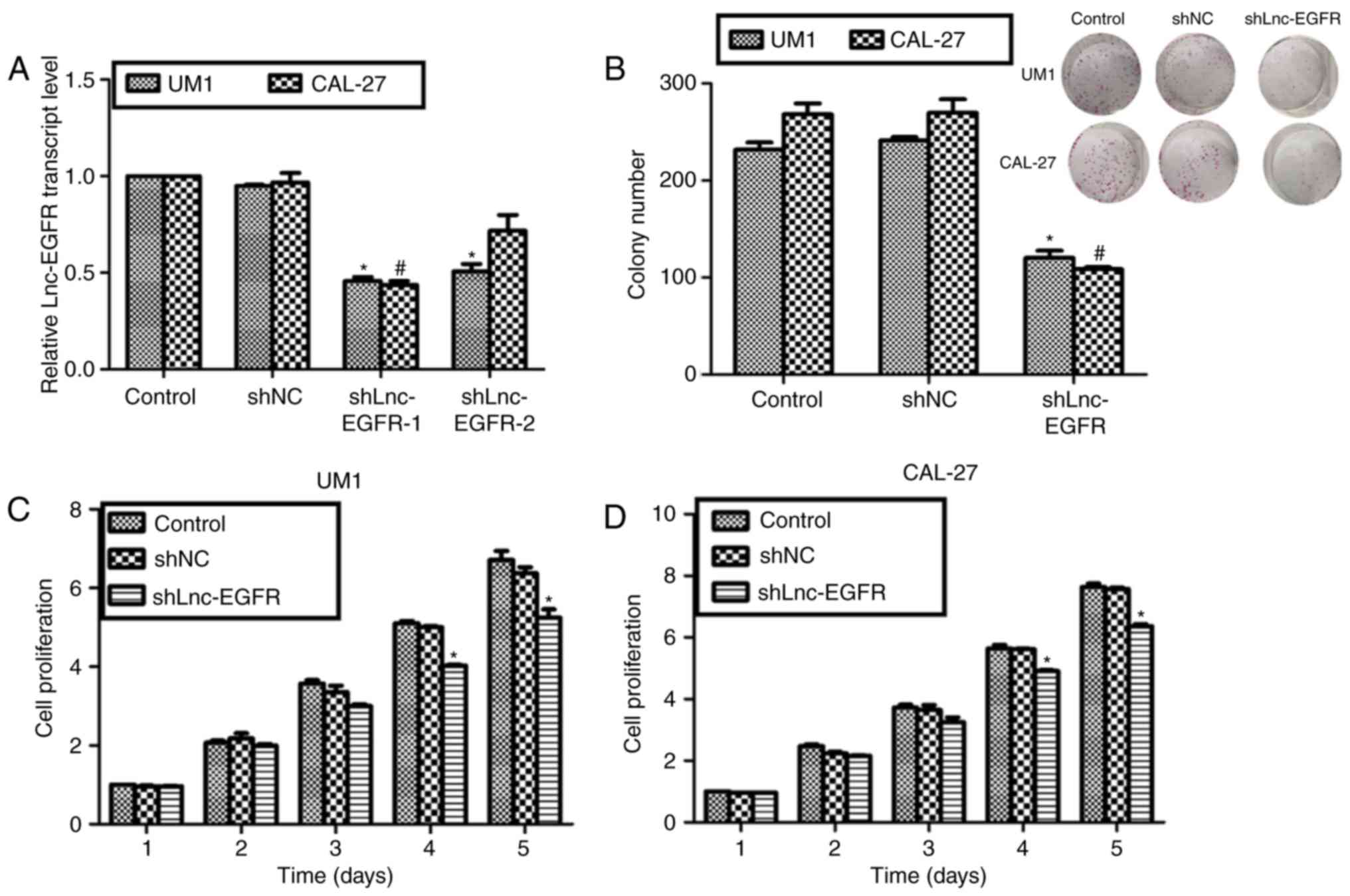
To explore the roles of Lnc-EGFR, two specific
shRNAs against Lnc-EGR were synthesized and transfected into UM1
and CAL-27 cells. The expression of Lnc-EGFR was decreased by more
than 50% in both cell lines upon shLnc-EGFR-1 transfection;
however, shLnc-EGFR-2 was only effective for UM1 cells (Fig. 2A). Thus, shLnc-EGFR-1 was selected
and renamed as shLnc-EGFR. Colony formation assays and cell
proliferation assays were performed to reveal the roles of
Lnc-EGFR. Approximate 230 colonies and 275 colonies were observed
in control UM1 cells and CAL-27 cells, respectively; however, only
an average of 110 colonies in UM1 cells and 105 colonies in CAL-27
cells were counted upon shLnc-EGFR transfection, while control
shRNA caused no effects on both cell lines (Fig. 2B). As for the cell proliferation
assays, there were no notable difference among the three
experimental groups in the former three days for both UM1 and
CAL-27 cells. Interestingly, the proliferative rate of UM1 was
suppressed by 24% on the fourth day and 29% on the fifth day
(Fig. 2C). Likewise, knockdown of
Lnc-EGFR with shLnc-EGFR inhibited cell proliferative rate on the
fourth and fifth day in CAL-27 cells (Fig. 2D). These data suggested that
knockdown of Lnc-EGFR in human tongue cancer suppressed cell
proliferation in vitro.
Knockdown of Lnc-EGFR increased cell
apoptosis in human tongue cancer in vitro
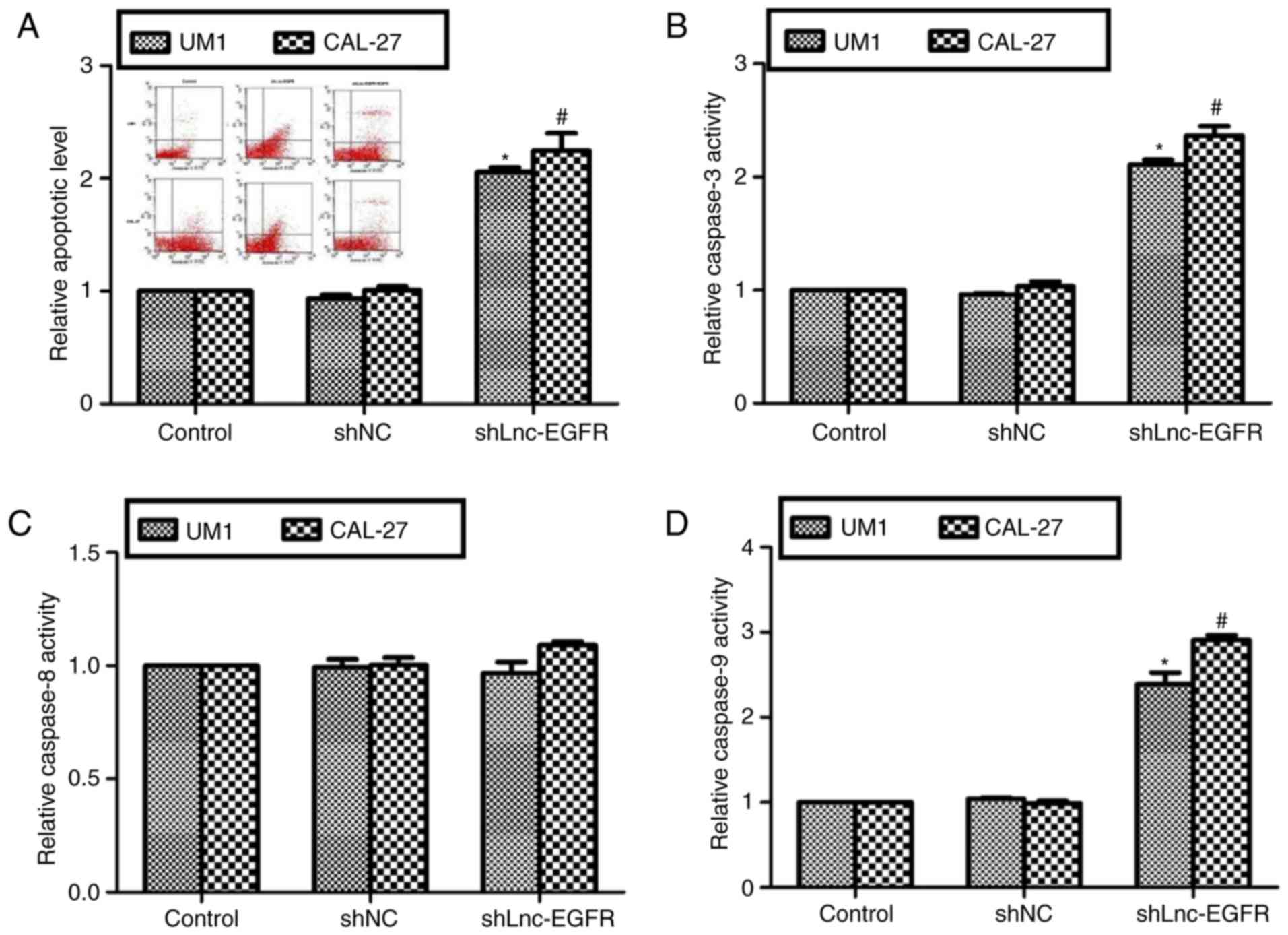
Next, cell apoptotic rates were assessed in UM1 and
CAL-27 cells. As shown in Fig. 3A,
upon knockdown of Lnc-EGFR for 72 h, the apoptotic rate was
increased to 2-fold in UM1 cells and 2.2-fold in CAL-27 cells. Cell
apoptosis have two classical signal pathways: intrinsic pathway
(caspase-3 and caspase-9) and extrinsic pathway (capsase-8)
(19). It was shown in Fig. 3B, the relative caspase-3 activities
were remarkably increased in both cell lines when Lnc-EGFR was
knocked down with shLnc-EGFR. However, the relative caspase-8
activities remained unchanged upon transfection of shLnc-EGFR
(Fig. 3C). Meanwhile, the
activities of caspase-9 were increased by more than 1-fold in both
cell lines (Fig. 3D). All of above
observations suggested that Lnc-EGFR suppressed cell apoptosis
through intrinsic pathway in human tongue cancer in
vitro.
Lnc-EGFR promoted cell proliferation
through EGFR in human tongue cancer cells
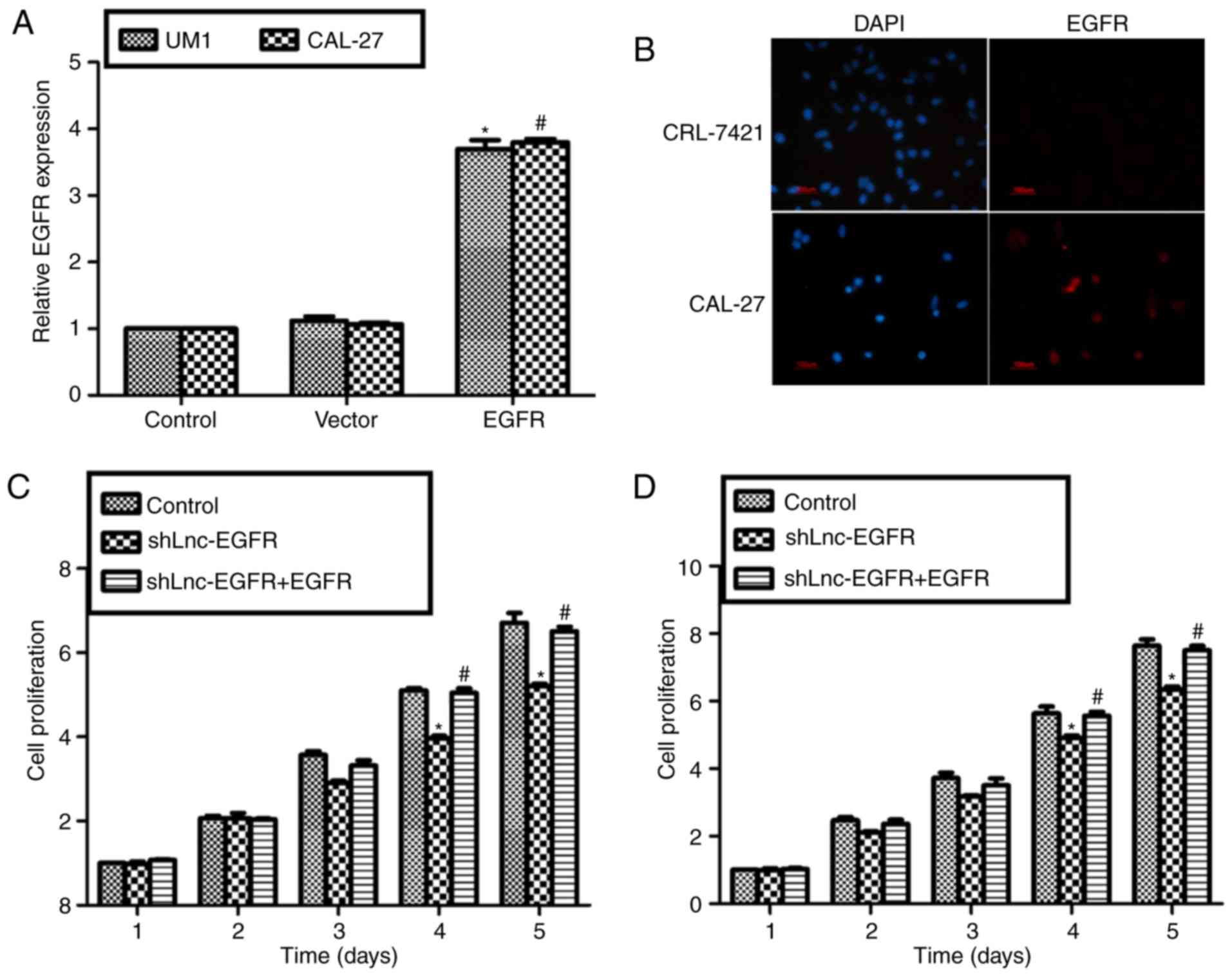
Both UM1 and CAL-27 cells showed elevated expression
of EGFR when recombinant protein EGFR were added into both cells
lines (Fig. 4A).
Immunofluorescence staining showed that there was almost no EGFR
expression in the control cell line CRL-7421 and the expression of
EGFR was obviously upregulated in CAL-27 cells (Fig. 4B). Similarly, although cell
proliferative rate was decreased upon transfection with shLnc-EGFR
in UM1 cells (Fig. 4C) and CAL-27
cells (Fig. 4D), the cell
proliferative capacity was recovered to basic level when shLnc-EGFR
and recombinant EGFR protein co-treated each cell line (Fig. 4C and D). Altogether with Fig. 2, our findings indicated Lnc-EGFR
increased cell proliferation through EGFR in human tongue cancer
cell lines UM1 and CAL-27.
Lnc-EGFR suppressed cell apoptosis
through EGFR in human tongue cancer cells
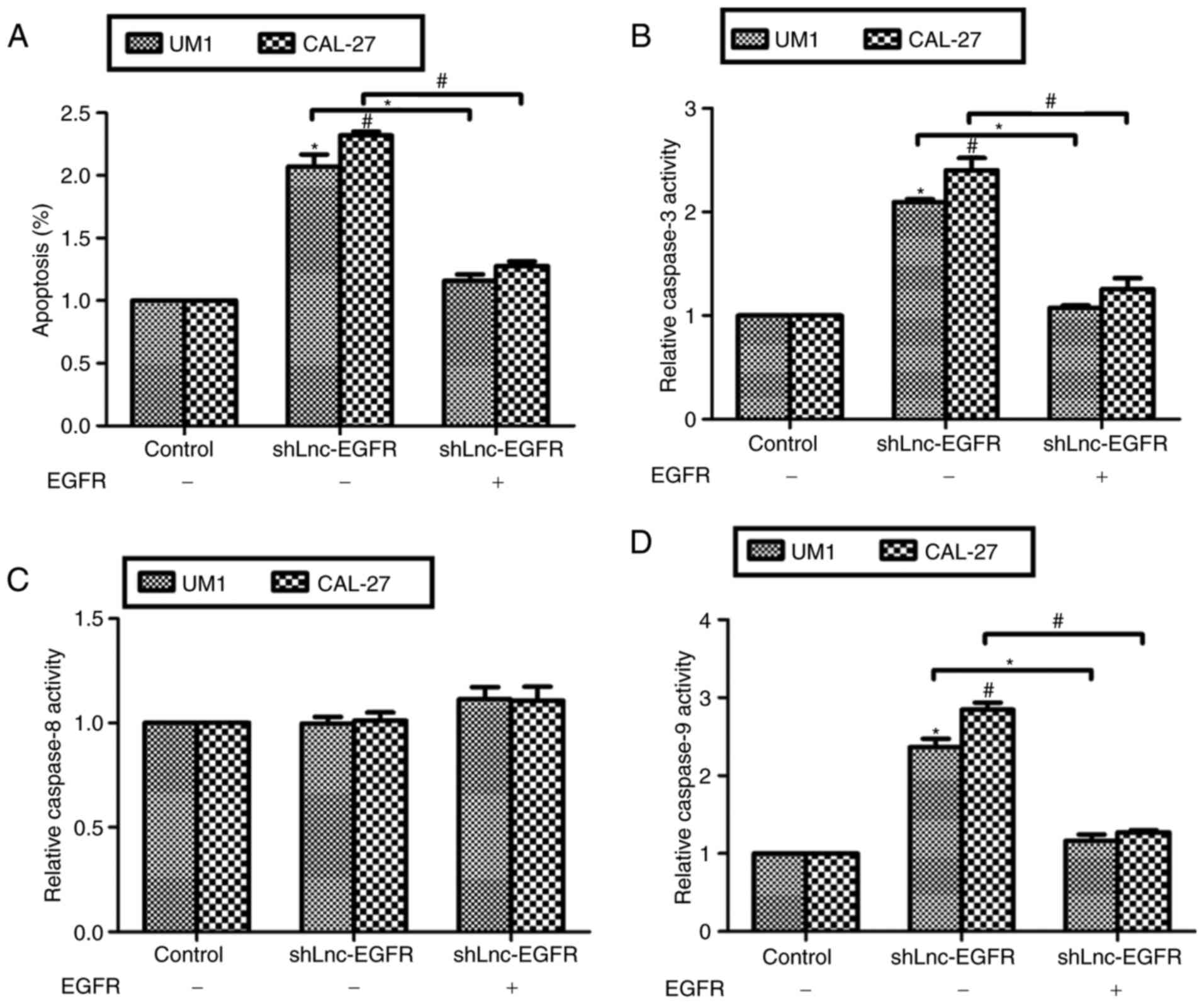
We also examined the effects of EGFR re-expression
on Lnc-EGFR knockdown-mediated cell apoptosis. As shown in Fig. 5, transfection of shLnc-EGFR
increased cell apoptosis to 2-fold in both cell lines, while the
cell apoptotic rate was decreased to the normal level when cells
were co-treated with shLnc-EGFR and recombinant EGFR protein.
Similarly, the relative activities of caspase-3 (Fig. 5B) and caspase-9 (Fig. 5D) were increased by knockdown of
Lnc-EGFR and recovered by co-treated with EGFR recombinant protein
in both UM1 and CAL-27 cells. However, the caspase-8 activity
remained unchanged despite any treatment (Fig. 5C). These data suggested that
Lnc-EGFR suppressed cell apoptosis through positive regulation of
EGFR in human tongue cancer cells in vitro.
Discussion
This is a preliminary study on the role of Lnc-EGFR
in human tongue cancer. In this study, we demonstrated the relative
transcript level of Lnc-EGFR was upregulated in clinical tongue
cancer tissues and in cultured tongue cancer cells, which was
consistent with that of EGFR. Knockdown of Lnc-EGFR inhibited
colony formation and cell proliferative rates in tongue cancer
cells UM1 and CAL-27, and increased cell apoptosis by enhancing the
activities of caspase-3 and caspase-9, but not caspase-8. More
interestingly, re-expression of EGFR in Lnc-EGFR-depleted UM1 and
CAL-27 cells rescued Lnc-EGFR depletion-mediated inhibition of cell
proliferative ability and promotion of cell apoptotic capacity.
These results suggested that the oncogenic potential of Lnc-EGFR
was achieved by upregulating the expression of EGFR.
Lnc-EGFR is a newly identified long noncoding RNA
using high-throughput screening in Treg cells in hepatocellular
carcinoma (18). Lnc-EGFR
upregulation in Tregs correlates positively with the tumor size and
expression of EGFR/Foxp3, but negatively with IFN-g expression in
patients and xenografted mouse models. Lnc-EGFR stimulates Treg
differentiation, suppresses CTL activity and promotes HCC growth in
an EGFR-dependent manner (18).
Consistent with this pioneer study, we also showed that Lnc-EGFR
was significantly increased in tongue cancer tissues. Altogether,
the results by others and us might suggested that Lnc-EGFR is a
novel oncogene gene. Lnc-EGFR might play a wide range of critical
roles in solid tumors. However, it remains scarce that how Lnc-EGFR
functions in solid tumors. We provided evidence that Lnc-EGFR
positively regulated the expression of EGFR, an important member of
the receptor tyrosine kinase family. Previously, it has been
indicated that Lnc-EGFR specifically binds to EGFR and blocks its
interaction with and ubiquitination by c-CBL, stabilizing it and
augmenting activation of itself and its downstream AP-1/NF-AT1
axis, which in turn elicits EGFR expression (18). Hence, we speculated that Lnc-EGFR
might directly bind to EGFR and regulate its expression in human
tongue cancer. However, it merits further investigation of this
detailed interaction and other possible mechanisms underlying
Lnc-EGFR functions remains to be further elucidated in tongue
cancer.
The induction of apoptosis is a good basis for
anticancer treatment and a valuable guide to predict tumor response
after anticancer therapies are monitored (20). Two major pathways are well-known to
be involved in the initiation of apoptosis: the
mitochondria-induced intrinsic pathway and the death
receptor-mediated extrinsic pathway (21). In the intrinsic pathway, once
cytochrome c is released, it binds with apoptotic protease
activating factor-1 (Apaf-1) and ATP, which then bind to
pro-caspase-9 to create a protein complex known as an apoptosome.
The apoptosome cleaves the pro-caspase to its active form of
caspase-9, which in turn activates the effector caspase-3 (22). In the extrinsic pathway, the Fas
(first apoptosis signal) receptor binds to the Fas ligand (FasL)
and results in the formation of the death-inducing signaling
complex (DISC), which contains the FADD, caspase-8 and caspase-10
(23). Therefore, we examined the
relative activities of caspase-3, caspase-8 and caspase-9 and
demonstrated that only intrinsic pathway was involved in
Lnc-EGFR-regulated cell apoptosis in human tongue cancer.
In total, our study was the first one to identify
the role of Lnc-EGFR in human tongue cancer and suggested that
Lnc-EGFR functioned by positive regulation of EGFR. Lnc-EGFR was
shown to promote cell proliferation and inhibit cell apoptosis
in vitro and this phenomenon could be reversed by decreasing
EGFR. Due to the key role of EGFR in tumorigenesis, this study
suggested the diagnostic value of Lnc-EGFR for tongue cancer and
might provide novel insights into the development of therapeutic
strategies for treatment of tongue cancer.
References
|
1
|
Leemans CR, Braakhuis BJ and Brakenhoff
RH: The molecular biology of head and neck cancer. Nat Rev Cancer.
11:9–22. 2011. View Article : Google Scholar
|
|
2
|
Hasegawa H, Kusumi Y, Asakawa T, Maeda M,
Oinuma T, Furusaka T, Oshima T and Esumi M: Expression of von
hippel-lindau tumor suppressor protein (pVHL) characteristic of
tongue cancer and proliferative lesions in tongue epithelium. BMC
Cancer. 17:3812017. View Article : Google Scholar :
|
|
3
|
Scully C, Field JK and Tanzawa H: Genetic
aberrations in oral or head and neck squamous cell carcinoma 3:
Clinico-pathological applications. Oral Oncol. 36:404–413. 2000.
View Article : Google Scholar
|
|
4
|
de Hoon M, Shin JW and Carninci P:
Paradigm shifts in genomics through the FANTOM projects. Mamm
Genome. 26:391–402. 2015. View Article : Google Scholar :
|
|
5
|
Xin Y, Li Z, Shen J, Chan MT and Wu WK:
CCAT1: A pivotal oncogenic long non-coding RNA in human cancers.
Cell Prolif. 49:255–260. 2016. View Article : Google Scholar
|
|
6
|
Guo X and Hua Y: CCAT1: An oncogenic long
noncoding RNA in human cancers. J Cancer Res Clin Oncol.
143:555–562. 2017. View Article : Google Scholar
|
|
7
|
Yang YT, Wang YF, Lai JY, Shen SY, Wang F,
Kong J, Zhang W and Yang HY: Long non-coding RNA UCA1 contributes
to the progression of oral squamous cell carcinoma by regulating
the WNT/β-catenin signaling pathway. Cancer Sci. 107:1581–1589.
2016. View Article : Google Scholar :
|
|
8
|
Wang L, Ye S, Wang J, Gu Z, Zhang Y, Zhang
C and Ma X: HuR stabilizes lnc-Sox5 mRNA to promote tongue
carcinogenesis. Biochemistry (Mosc). 82:438–445. 2017. View Article : Google Scholar
|
|
9
|
Zhang TH, Liang LZ, Liu XL, Wu JN, Su K,
Chen JY, Zheng QY, Huang HZ and Liao GQ: Long non-coding RNA MALAT1
interacts with miR-124 and modulates tongue cancer growth by
targeting JAG1. Oncol Rep. 37:2087–2094. 2017. View Article : Google Scholar
|
|
10
|
Zhang H, Zhao L, Wang YX, Xi M, Liu SL and
Luo LL: Long non-coding RNA HOTTIP is correlated with progression
and prognosis in tongue squamous cell carcinoma. Tumour Biol.
36:8805–8809. 2015. View Article : Google Scholar
|
|
11
|
Arteaga CL: Overview of epidermal growth
factor receptor biology and its role as a therapeutic target in
human neoplasia. Semin Oncol. 29 5 Suppl 14:S3–S9. 2002. View Article : Google Scholar
|
|
12
|
Herbst RS: Review of epidermal growth
factor receptor biology. Int J Radiat Oncol Biol Phys. 59 2
Suppl:S21–S26. 2004. View Article : Google Scholar
|
|
13
|
Gao JW, Zhan P, Qiu XY and Song Y:
Erlotinib-based doublet targeted therapy versus erlotinib alone in
previously treated advanced non-small-cell lung cancer: A
meta-analysis from 24 randomized controlled trials. Oncotarget.
32:2017.
|
|
14
|
Walker F, Abramowitz L, Benabderrahmane D,
Duval X, Descatoire V, Hénin D, Lehy T and Aparicio T: Growth
factor receptor expression in anal squamous lesions: Modifications
associated with oncogenic human papillomavirus and human
immunodeficiency virus. Hum Pathol. 40:1517–1527. 2009. View Article : Google Scholar
|
|
15
|
Alterio D, Marvaso G, Maffini F, Gandini
S, Chiocca S, Ferrari A, Preda L, Rocca MC, Lepanto D, Fodor C, et
al: Role of EGFR as prognostic factor in head and neck cancer
patients treated with surgery and postoperative radiotherapy:
Proposal of a new approach behind the EGFR overexpression. Med
Oncol. 34:1072017. View Article : Google Scholar
|
|
16
|
Jonsson Lindell E, Nylander K, Hallén L
and Laurell G: Immunohistochemical analysis of EGFR and hyaluronan
in tongue cancer and the development of regional recurrence in
patients initially diagnosed N0. Acta Otolaryngol. 137:877–882.
2017. View Article : Google Scholar
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
18
|
Jiang R, Tang J, Chen Y, Deng L, Ji J, Xie
Y, Wang K, Jia W, Chu WM and Sun B: The long noncoding RNA lnc-EGFR
stimulates T-regulatory cells differentiation thus promoting
hepatocellular carcinoma immune evasion. Nat Commun. 8:151292017.
View Article : Google Scholar :
|
|
19
|
Islam S, Qi W, Morales C, Cooke L, Spier
C, Weterings E and Mahadevan D: Disruption of aneuploidy and
senescence induced by aurora inhibition promotes intrinsic
apoptosis in double hit or double expressor diffuse large b-cell
lymphomas. Mol Cancer Ther. doi: 10.1158/1535-7163. 2017.
View Article : Google Scholar
|
|
20
|
Li HH, Su JH, Chiu CC, Lin JJ, Yang ZY,
Hwang WI, Chen YK, Lo YH and Wu YJ: Proteomic investigation of the
sinulariolide-treated melanoma cells A375: Effects on the cell
apoptosis through mitochondrial-related pathway and activation of
caspase cascade. Mar Drugs. 11:2625–2642. 2013. View Article : Google Scholar :
|
|
21
|
Liu JY, Liu Z, Wang DM, Li MM, Wang SX,
Wang R, Chen JP, Wang YF and Yang DP: Induction of apoptosis in
K562 cells by dicyclohexylammonium salt of hyperforin through a
mitochondrial-related pathway. Chem Biol Interact. 190:91–101.
2011. View Article : Google Scholar
|
|
22
|
Kerr JF, Wyllie AH and Currie AR:
Apoptosis: A basic biological phenomenon with wide-ranging
implications in tissue kinetics. Br J Cancer. 26:239–257. 1972.
View Article : Google Scholar :
|
|
23
|
Wajant H: The Fas signaling pathway: More
than a paradigm. Science. 296:1635–1636. 2002. View Article : Google Scholar
|