Introduction
Asthma is one of the most common chronic respiratory
diseases in the world. Clear evidence now suggests that asthma is a
heterogeneous and genetically complex disease, with phenotypes
conferred by both genetic susceptibility and environmental
exposure. More than 100 genes have been identified in association
with asthma (1), among which the
orosomucoid-like 3 gene (ORMDL3) and the associated 17q21 locus
have emerged in genome-wide association studies (GWAS) as likely
contributors to the genetic susceptibility and underlying
pathogenesis of asthma. This genetic association has been confirmed
in ethnically diverse populations (2). Although the functions of ORMDL3 are
incompletely understood, Balantic et al (3) have found that the genetic
polymorphism rs4795405 in ORMDL3 is closely related not only to
asthma but also to chronic obstructive pulmonary disease (COPD).
Thus, ORMDL3 may be associated with airway remodeling in
asthma.
Airway remodeling refers to pathological changes in
severe and refractory asthma that include structural changes in the
airway wall caused by chronic inflammation, resulting in persistent
or progressive damage to lung function. The principal features of
airway remodeling are airway epithelial damage, goblet cell
hyperplasia and metaplasia, subepithelial fibrosis and increased
smooth muscle mass. Numerous recent studies have shown that the
airway epithelial cells are the first barrier in the airway and
play an important role in airway remodeling (4,5).
Stimulation by allergens and environmental factors, as well as the
release of inflammatory mediators and cytokines, may induce damage
of the airway epithelial barrier, reduced adhesion among epithelial
cells, down-regulation of epithelial genes, and up-regulation of
interstitial cell genes (6).
Epithelial cells may participate in airway remodeling through
epithelial-mesenchymal transition (EMT) (5).
Miller et al (7,8) have
observed that ORMDL3 is expressed in lung bronchial epithelial
cells after allergen exposure and that it regulates the expression
of metalloproteinases, TGF-β and ADAM-8. ORMDL3 transgenic mice
have increased levels of airway remodeling and exhibit
characteristics similar to those of asthma, including hypertrophy
of airway smooth muscles and increased epithelial fibers and lumen
mucus (7,8). Therefore, we propose that ORMDL3 may
induce bronchial EMT under chronic inflammatory stimulation.
If ORMDL3 is closely related to human asthma, the
expression of ORMDL3 in asthma, the relationship between ORMDL3 and
the severity of airway remodeling in asthma, and the ability of
ORMDL3 to induce EMT in the airway warrant further exploration.
Humans and mice both express three ORMDL family members, and ORMDL3
has 96% homology between these two species, supporting the use of
mice as an appropriate animal model to study the function of ORMDL3
in asthma (9). Thus, in this
study, we observed the trends of ORMDL3 and the EMT-related
indicators E-cadherin (E-cad), fibroblast-specific protein 1
(FSP1), and Vimentin (VIM) at different time points of airway
remodeling in an asthmatic mouse model. Furthermore, an ORMDL3
plasmid was used to transfect the bronchial epithelial cell line
16HBE-14o-, and the invasive ability of 16HBE 14o-cells was
assessed by microscopy. In addition, changes in the expression of
E-cad, VIM, and FSP-1 were assessed to determine whether ORMDL3
induces EMT in bronchial epithelial cells.
Materials and methods
Asthmatic mouse model
A total of 56 specific-pathogen-free female BALB/c
mice (8 weeks old) were purchased from the experimental animal
research center of Shengjing Hospital of China Medical University
(Shenyang, China). All experimental protocols involving animals
were approved by the China Medical University Animal Care Committee
and complied with the guidelines of the China Council on Animal
Care. The animals were randomly assigned to the control group
(n=24), the model group (n=24) and the treatment group (n=8) at the
beginning of the experiment. The mice in the model group were
prepared according to previously described methods using a modified
ovalbumin (OVA; O1641; Sigma-Aldrich, St. Louis, MO, USA)
immunization protocol developed to induce allergic asthma in mice
(10). The mice were initially
intraperitoneally immunized with 20 µg of OVA and 2 mg of aluminum
hydroxide gel (Sigma-Aldrich) in 0.5 ml of PBS on days 0, 7, and
14, as previously reported (11).
Control mice were injected with saline only. From day 16 onward,
the animals were treated every other day for 14 days as follows.
The mice in the model group were placed in a transparent glass
chamber (approximately 20×20×20 cm in volume) connected to an
ultrasonic nebulizer (model 100; Yadu, Shanghai, China) and
subjected to repeated bronchial allergen challenge by inhalation of
OVA (4%) for 30 min/day. The mice in the control group were
challenged with phosphate buffer saline (PBS). The mice in the
dexamethasone treatment group were OVA sensitized and
intraperitoneal injected with 0.2 ml dexamethasone (2 mg/ml) before
OVA challenged. On days 3, 7, and 14 of allergen challenge, 8 mice
from control group and asthma group were anesthetized by
intraperitoneal injection with 5% chloral hydrate and sacrificed.
While all of the treatment group were sacrificed on days 14 of
challenged.
Whole lungs were collected aseptically by chest
opening. The left lungs were fixed in paraformaldehyde (PFA) for
subsequent immunohistochemical staining; the right upper lung lobes
were used for real-time PCR analysis, and the right lower lung
lobes were used for western blot analysis. All specimens were
snap-frozen in liquid nitrogen and stored at −80°C until use.
Lung histopathology
The left lungs were fixed in 4% PFA for 18–24 h,
embedded in paraffin and then routinely processed. Serial 5-µm
tissue sections were stained with hematoxylin and eosin (H&E)
for general histological evaluation. Masson's trichrome staining
was performed for the assessment of collagen particles and airway
remodeling.
Morphometric analysis of the
airway
H&E- and Masson-stained tissue sections were
examined by light microscopy at magnification, ×200, and
morphometric measurements were obtained using Image-Pro Plus 6.0
software (Media Cybernetics, Inc., Rockville, MD, USA) to assess
bronchiolar remodeling and collagen deposition. For H&E-stained
lung sections from each group, the complete cross-section of the
bronchiole without cartilage was selected for analysis. The total
bronchial wall area (Wat, µm2) and the diameter of the
basal membrane perimeter (Pbm, µm) were analyzed for each
bronchiole. Wat/Pbm expresses the wall thickness of the bronchiole
wall. Masson staining was used to assess the deposition of
collagen. Two complete bronchioles were selected from each tissue
section, and the percentage of the area of collagen deposition in
the total stained area was calculated (12,13).
Transmission electron microscopy
After dissection, lung specimens were fixed in 2.5%
glutaraldehyde overnight at 4°C. The tissues were washed in
phosphate buffer (pH 7.4), postfixed in 1% osmium tetroxide in
phosphate buffer, dehydrated in graded ethanol solutions, treated
in propylene oxide and embedded in epoxy-resin embedding media.
Thin transverse random sections (60 nm) were collected on single
copper slot grids coated with parlodion, stained with uranyl
acetate and lead citrate, and observed with a transmission electron
microscope (H-7650; Hitachi Limited, Tokyo, Japan). Images were
documented using Kodak SO163 EM film (Kodak, Rochester, NY,
USA).
Immunohistochemistry
ORMDL3 and FSP-1 protein expression in the lung was
detected by immunohistochemistry. Lung sections (5 µm) were cut,
blocked with peroxide and non-immune animal serum, and incubated
sequentially with anti-ORMDL3 antibody (dilution, 1:200; LS-B 9583;
LifeSpan BioSciences, Inc., Seattle, WA, USA) or FSP-1 antibody
(dilution, 1:200; 16105-1-AP; Proteintech, Wuhan, China),
biotin-labeled secondary antibody, and streptomycin anti-biotin
peroxidase. Finally, the sections were stained with DAB,
counterstained with hematoxylin, dehydrated, cleared in xylene, and
fixed. Negative staining controls were generated by replacing the
primary antibody with PBS. A laser scanning confocal microscope
(MTC-600; Bio-Rad, Hercules, CA, USA) was used for image
acquisition, and the deposition of brown particles in the cytoplasm
indicated a positive result.
Cell culture and transfection
Immortalized SV-40 virus-transformed 16HBE 14o-human
bronchial epithelial cells were purchased from the cell line
resource of XiangYa School of Medicine. The cells were cultured in
RPMI 1640 medium (SH30809.01B; HyClone; GE Healthcare, Logan, UT,
USA) containing 12% fetal bovine serum (FBS; SH30084.03; HyClone;
GE Healthcare), penicillin (100 U/ml), and streptomycin (100 µg/ml)
at 37°C in a humidified 5% CO2 atmosphere. 16HBE-14o
cells were efficiently transfected with either the
GV230-ORMDL3-eGFP plasmid, which encodes the full-length ORMDL3
cDNA with a terminal GFP tag, or empty vector containing the GFP
tag (eGFP-control) as a negative control (GeneChem, Shanghai,
China) using Lipofectamine 2000 (11668019; Invitrogen, Carlsbad,
CA, USA) according to the manufacturer's instructions to generate
the GFP-ORMDL3 group and GFP-vector group, respectively.
Transwell analysis
To study the role of ORMDL3 in 16HBE 14o-migration,
Transwell analysis was conducted after harvesting the transfected
cells with trypsin and resuspending them (1.0×106
cells/ml) in serum-free growth medium. The two groups of cells were
added to the upper chamber. After incubation for 24 h at 37°C, the
membranes were removed, the cells on the upper side were scraped
off, and the cells that migrated to the lower side of the membrane
were fixed with 4% polyoxymethylene. The number of cells was
counted in 5 random fields under magnification, ×20, and the mean
was calculated.
Immunocytochemistry
The two groups of 16HBE 14o-cells were cultured on
coverslips, and protein expression (E-cad, FSP-1) was detected by
immunocytochemistry. After the cells reached confluency, they were
fixed in 4% polyphosphate formaldehyde, blocked with peroxide and
non-immune animal serum, and incubated sequentially with primary
antibody (anti-E-cad; dilution, 1:50; ab11512, Abcam, Cambridge,
UK; anti-FSP-1; dilution, 1:50; Santa Cruz, CA, USA) at 4°C. Rat
IgG (for E-cad) and Goat IgG (for FSP-1) were used as controls, and
Cy3-conjugated donkey anti-goat IgG (dilution, 1:200; Proteintech)
and goat anti-rat IgG (dilution, 1:200; Santa Cruz) were used as
the secondary antibodies. The slides were washed, mounted in medium
containing 4,6-diamidino-2-phenylindole (DAPI) and visualized by
confocal microscopy (×400) at ambient temperature.
Western blot analysis
Total protein extracted from lung tissue and
16HBE-14o cells was quantified using a BCA protein assay kit. A
20-µl aliquot of protein was then loaded onto a 12% SDS-PAGE gel
and transferred onto polyvinylidene difluoride membranes
(Millipore, Billerica, MA, USA). The membranes were blocked with 5%
skimmed milk for 1 h and then incubated with primary antibody
(dilution, 1:800, ORMDL3; dilution, 1:5,000, E-cad; dilution,
1:1,000 FPS-1; dilution, 1:200 VIM; and dilution, 1:200, GAPDH)
diluted in PBS overnight at 4°C. After washing in Tris-buffered
saline + 1% Tween-20 (TBST), the membranes were incubated for 2 h
with horseradish peroxidase-conjugated secondary antibody
(dilution, 1:500) and then imaged using enhanced chemiluminescence
reagents. The density of the protein bands was analyzed using
ImageJ software. GAPDH was used as an internal control.
Real-time PCR
Total RNA was isolated following the manufacturer's
instructions using TRIzol reagent (Invitrogen, Carlsbad, CA, USA)
and RNAiso™ Plus reagent (Takara, Dalian, China) and
quantified using a spectrophotometer. Following quantification, 2
µg of RNA was reverse-transcribed into cDNA, and real-time
quantitative PCR assays were conducted using an ABI PRISM 7500
Real-time PCR System (Applied Biosystems, Foster City, CA, USA) and
the SYBR PrimeScript™ RT-PCR kit reagent (Takara,
Dalian, China). The PCR conditions for NK-1R were 40 cycles of
denaturation at 95°C for 5 sec and annealing and extension at 60°C
for 30 sec. Target mRNA levels were normalized to those of GAPDH.
The oligonucleotide primers used are presented in Table I.
 | Table I.Primer sequences used for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primer sequences used for reverse
transcription-quantitative polymerase chain reaction.
| Gene name | Species | Primer sequences
(5′-3′) |
|---|
| GAPDH | Human | F:
GAGAAGGCTGGGGCTCATTT |
|
|
| R:
AGTGATGGCATGGACTGTGG |
| ORMDL3 | Human | F:
GAGCATCCCGTTTGTGAGTGTC |
|
|
| R:
CCATCTGCTCCCAGTGGGTTA |
| E-cad | Human | F:
GAGTGCCAACTGGACCATTCAGTA |
|
|
| R:
AGTCACCCACCTCTAAGGCCATC |
| VIM | Human | F:
GCAGGAGGCAGAAGAATGGT |
|
|
| R:
ACGAAGGTGACGAGCCATTT |
| FSP-1 | Human | F:
GGGTGACAAGTTCAAGCTCAACAA |
|
|
| R:
ATCATGGCGATGCAGGACAG |
| GAPDH | Mouse | F:
CTACCCCCAATGTGTCCGTC |
|
|
| R:
GGGATAGGGCCTCTCTTGCT |
| ORMDL3 | Mouse | F:
CCCTGTGGGTTTGAACTCCTG |
|
|
| R:
GTCGAAGTGAACCCGCTTCTG |
| E-cad | Mouse | F:
ATTGCAAGTTCCTGCCATCCTC |
|
|
| R:
CACATTGTCCCGGGTATCATCA |
| VIM | Mouse | F:
CCTATGTGACCCGGTCCTCG |
|
|
| R:
AAGGTCAAGACGTGCCAGAG |
| FSP-1 | Mouse | F:
GGAGGCCCTGGATGTAATTGTG |
|
|
| R:
CAACTTCATTGTCCCTGTTGCTG |
Statistical analysis
SPSS 17 software was used for statistical analysis.
Inter-group comparisons were performed using the t-test, multiple
group comparisons were conducted using one-way analysis of variance
(ANOVA), and correlation analysis was performed using Pearson's
test. All data are expressed as the mean (X) ± standard deviation
(SD). P<0.05 was considered to indicate a statistically
significant difference.
Results
Airway remodeling increases with the
development of asthma
The development of airway remodeling was assessed in
lung tissues stained with H&E and Masson's trichrome. We
observed the level of peribronchial cellular infiltration and
evaluated bronchial wall thickness and lung collagen content in all
experimental mice. Representative sections of each group were
stained with either H&E (Fig.
1A-C) or Masson's trichrome (Fig.
1E-G). As shown in Fig. 1A, no
inflammation, mucosal edema or epithelial lesions were observed in
the control group. Mild inflammation, mucosal edema and epithelial
lesions were observed in the asthma group at 3 days, whereas
sensitization to OVA at 7 days and 14 days was associated with
predominantly moderate to severe inflammation, mucosal edema and
epithelial lesions that included interstitial infiltrates and large
lymphoid aggregates (Fig. 1B).
Bronchial wall thickness was quantitatively estimated based on
H&E staining (Fig. 1D) and was
observed to increase with the duration of OVA exposure
(P<0.01).
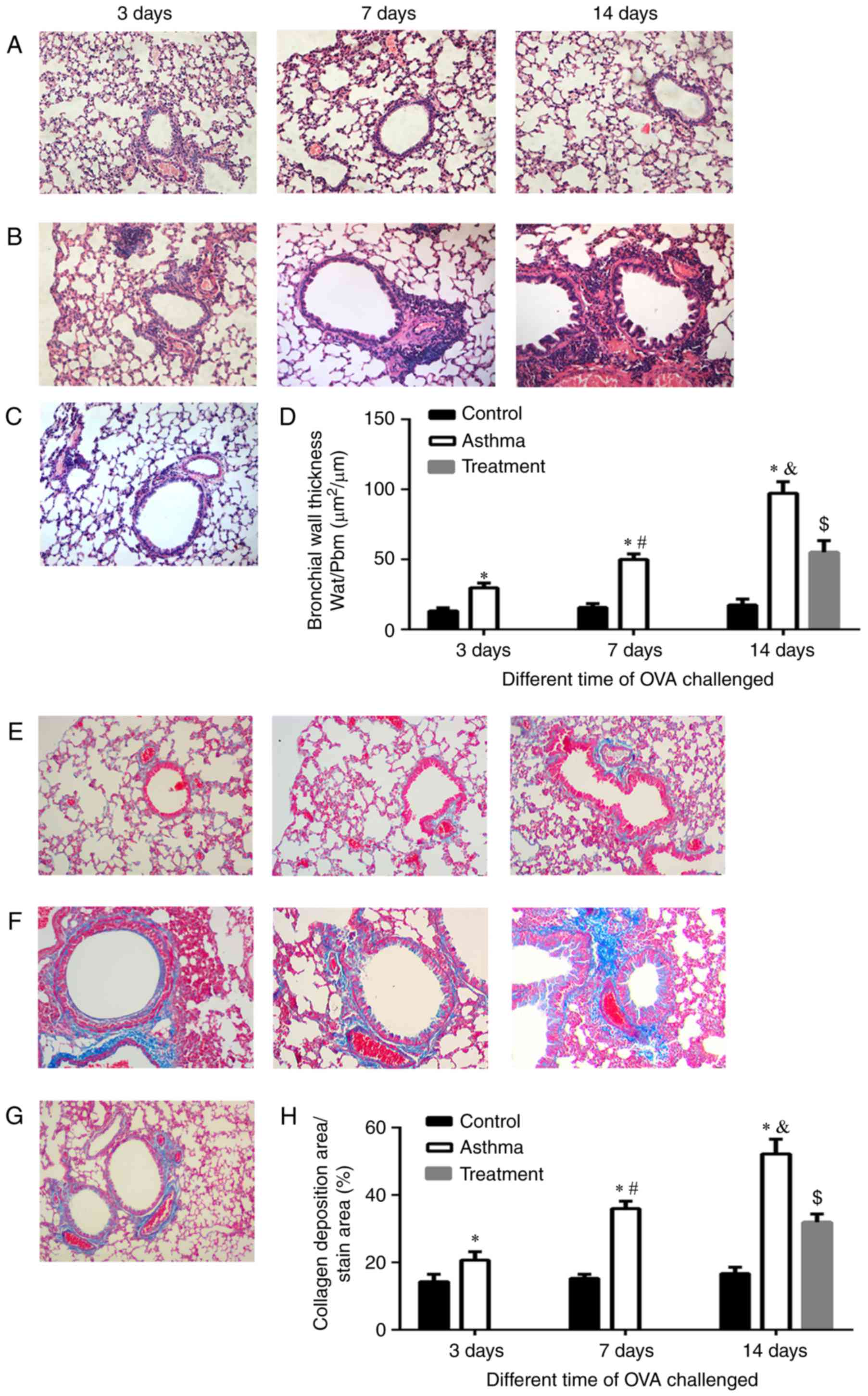 | Figure 1.Airway remodeling increased with the
development of asthma. H&E staining of paraffin sections of
lung tissues from BALB/c mice in the (A) control, (B) asthma and
(C) treatment groups with different durations of exposure to OVA
(magnification, ×200). Masson staining of lung tissues from BALB/c
mice in the (E) control, (F) asthma and (G) treatment groups with
different durations of exposure to OVA (magnification, ×200).
Bronchial wall thickness (Wat/Pbm) was assessed by (D) H&E
staining, and the percentage of the area of collagen deposition was
assessed by (H) Masson staining. Wat/Pbm, the ratio of the total
bronchial wall area (Wat, µm2) to the diameter of the
basal membrane perimeter (Pbm), represents the thickness of the
bronchial wall and was used to evaluate the severity of airway
remodeling. The percentage of the area of collagen deposition was
used to determine the severity of fibrosis in airway remodeling.
*P<0.01 vs. control; #P<0.01 vs. asthma group at 3
days; &P<0.01 vs. asthma group at 7 days;
$P<0.01 vs. treatment group. H&E, hematoxylin and
eosin; OVA, ovalbumin. |
As the duration of OVA exposure increased, the
peribronchial trichrome-stained area and lung collagen levels were
increased at 3, 7 and 14 days (Fig.
1F) of asthma group compared with those of the control group
(Fig. 1E). The collagen content
was quantitatively estimated (Fig.
1H) by Masson staining and was observed to increase with an
increasing duration of OVA exposure (P<0.01). While in the
treatment group, bronchial wall thickness was reduced and the
collagen content reduced compared with the asthmatic model group;
however, it was still higher than the control group, respectively
(Fig. 1C, D, G and H)
(P<0.01).
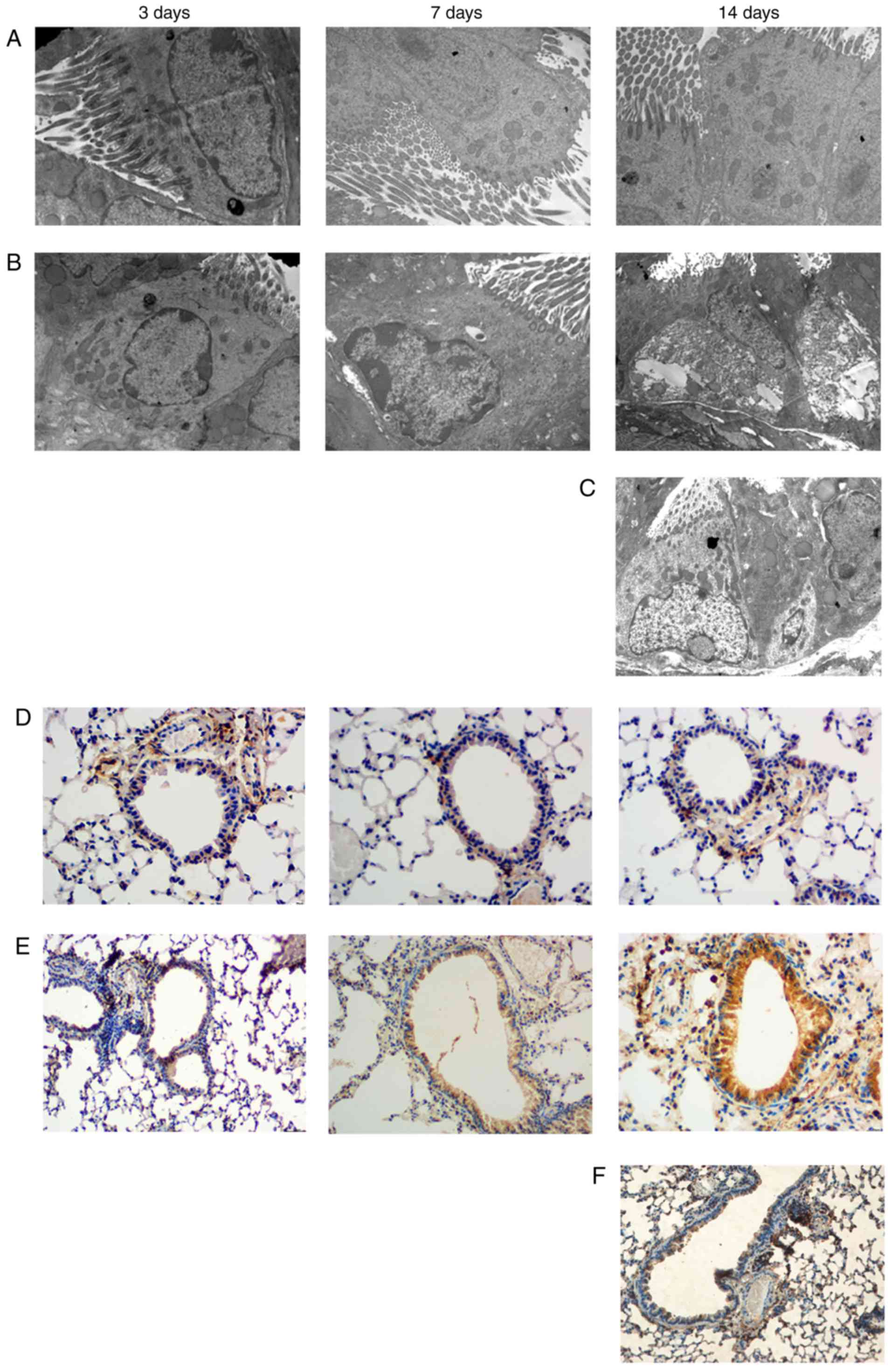
EMT is exacerbated with the
development of asthma
Morphological changes in bronchiolar epithelial
cells were assessed by transmission electron microscopy. The
control group (Fig. 2A) exhibited
normal bronchial epithelial cells without edema, neatly arranged
cilia, no or rare cytoplasmic vacuoles, regularly shaped nuclei,
mitochondria, endoplasmic reticulum dilation, thin subepithelial
basal lamina, few collagen fibers, and intercellular tight linking.
At 3 days, the asthma group exhibited disordered cilia and a
slightly expanded endoplasmic reticulum. At 7 days, in addition to
disordered cilia, the asthma group exhibited obvious dilation of
mitochondria and the endoplasmic reticulum, basal lamina
thickening, and collagen deposition in clusters. At 14 days, the
asthma group exhibited shedding of the ciliated epithelium,
undetached ciliated epithelial cells, significantly increased
bronchial epithelial cells, mitochondrial swelling, endoplasmic
reticulum expansion, increased cytoplasmic vacuoles, globally
increased secretory cells, intercellular tight junction gaps, and
increased basal lamina collagen fibers (Fig. 2B). However, in the treatment group,
the damage of organelles and ciliate epithelium was alleviated by
asthma group (Fig. 2C).
E-cad, FSP1, and VIM were evaluated as EMT-related
indicators. FSP-1 is a fibroblast marker that indicates EMT when
expressed in epithelial cells. Immunohistochemistryrevealed that
FSP-1 expression was nearly absent in bronchial epithelial cells in
the control group. However, some FSP-1 was observed on the
bronchial epithelium in the asthma group at 3 days, and obvious
FSP-1 staining on the bronchial epithelium was observed in the
asthma group at 7 days, with even greater staining at 14 days
(Fig. 2E). While FSP-1 expression
decrease in the treatment group.
Western blotting (Fig.
3A) revealed that OVA challenge resulted in decreased E-cad
expression and increased VIM and FSP-1 expression in the asthma
group compared with the control group, consistent with EMT
progression.
Real-time PCR demonstrated that the mRNA expression
of E-cad (Fig. 3C), FSP-1
(Fig. 3D), and VIM (Fig. 3E) did not differ significantly
between the asthma group at 3 days and the control group
(P>0.05). At 7 and 14 days, E-cad expression (Fig. 3C) was significantly lower and FSP-1
(Fig. 3D) and VIM expression
(Fig. 3E) was greater in the
asthma group than in the control group (P<0.01).
The treatment group signifcantly increased the mRNA
and protein expression of E-cad, and decreased the mRNA and protein
expression of VIM and FSP-1 compared with asthma group (Fig. 3B-E) (P<0.01).
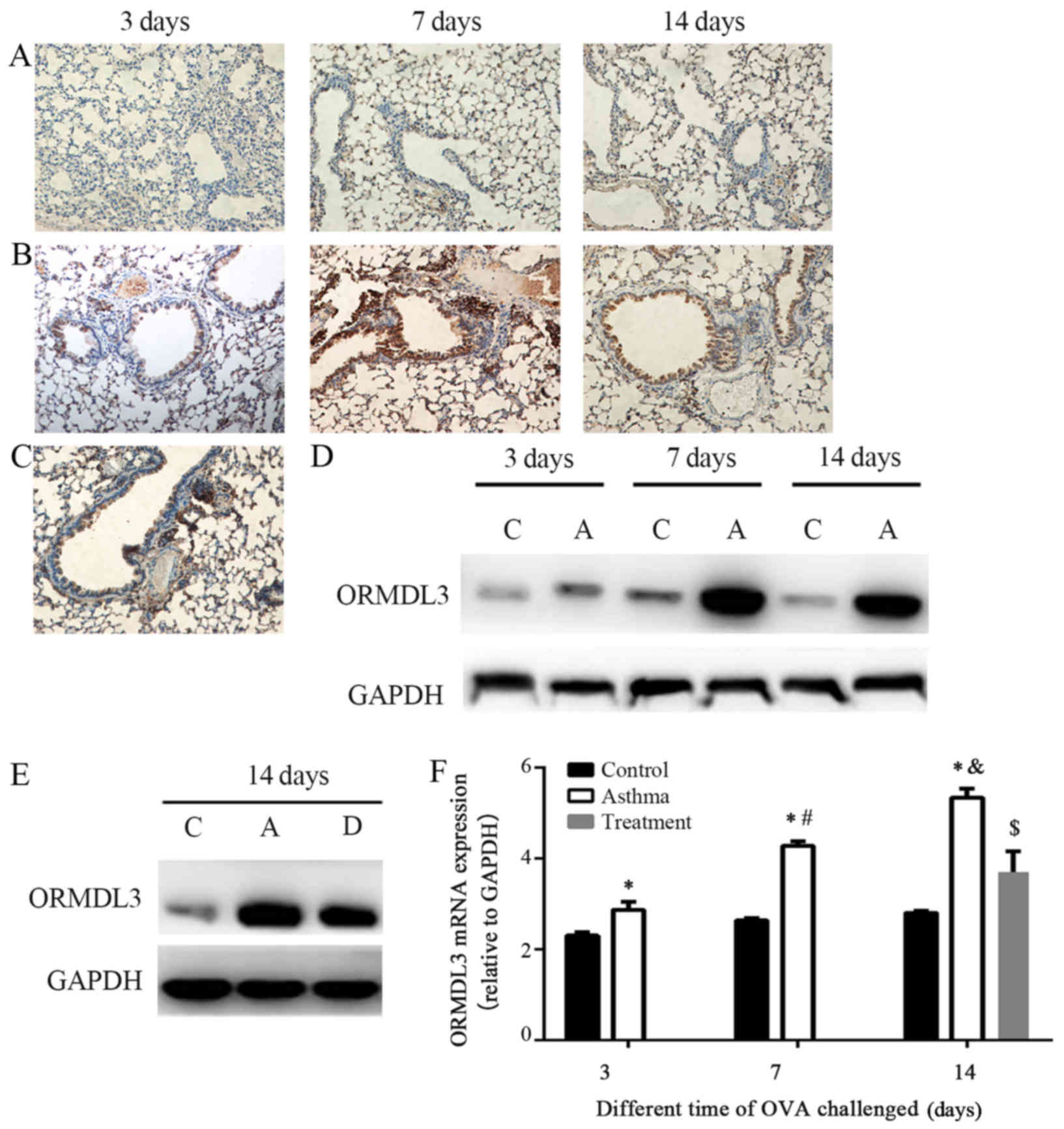
ORMDL3 expression in the bronchial
epithelium is associated with airway remodeling and EMT progression
in vivo
We examined the location of the ORMDL3 protein by
immunohistochemical staining of lung tissue from the control and
asthma groups exposed to OVA over time. After OVA challenge, ORMDL3
was expressed in both the bronchial epithelium and lymphocytes
around blood vessels in the asthma group (Fig. 4B), alleviated in treatment group
(Fig. 4C), but was poorly
expressed in the control group (Fig.
4A).
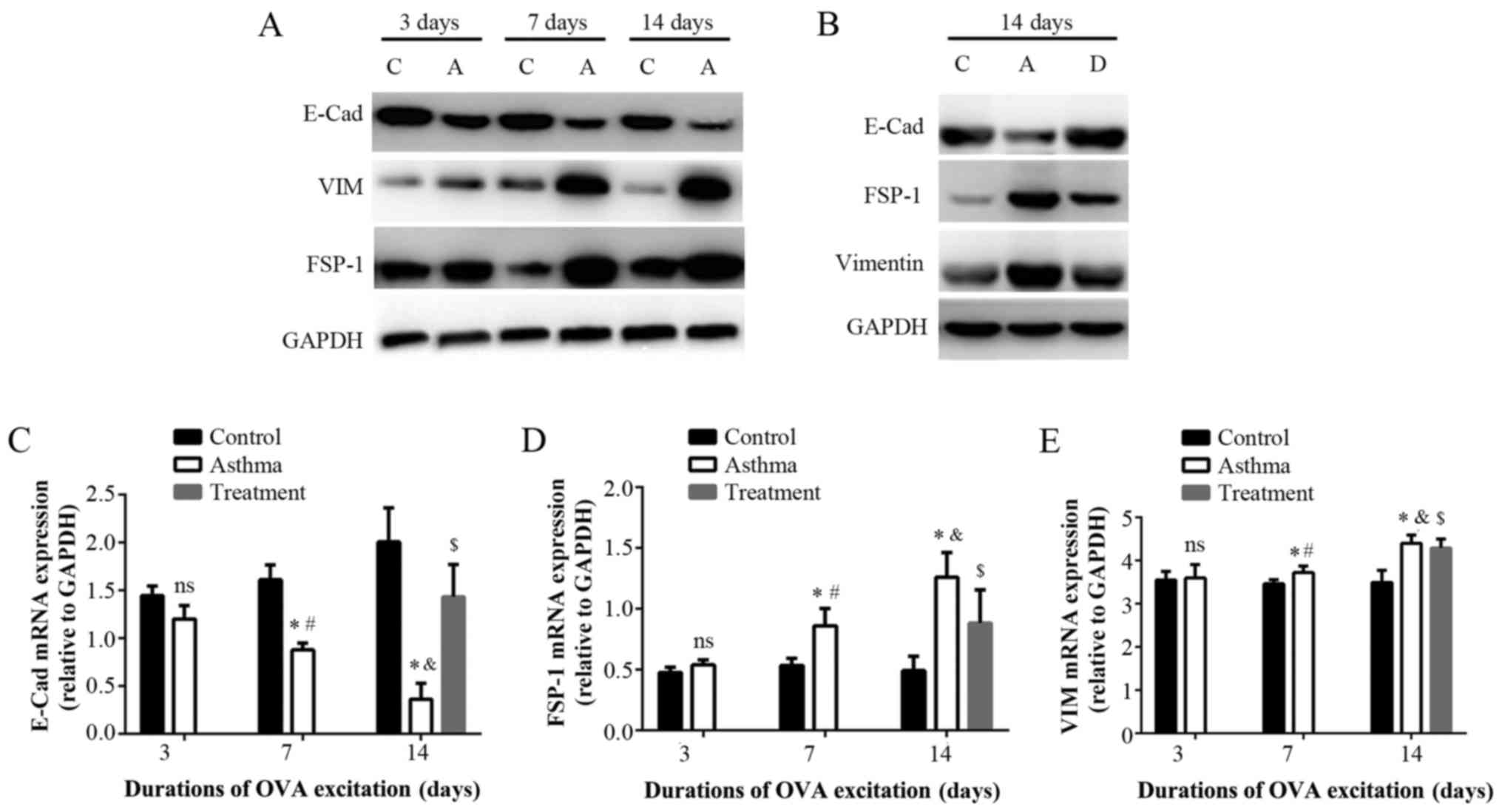 | Figure 4.Protein and mRNA expression of
epithelial-mesenchymal transition associated indicators induced by
the development of asthma. (A) Western blotting of the relative
protein expression of E-cad, VIM and FSP-1 in the lung tissues of
the control and asthma groups at different time points. (B) Western
blotting of the relative protein expression of E-cad, VIM and FSP-1
in the lung tissues of the control, asthma and treatment groups
following 14 days exposure to OVA. Reverse
transcription-quantitative polymerase chain reaction of the
relative mRNA expression of (C) E-cad, (D) FSP-1 and (E) VIM in the
lung tissues of the different groups. *P<0.01 vs. control;
#P<0.01 vs. asthma group at 3 days;
&P<0.01 vs. asthma group at 7 days;
$P<0.01 vs. treatment group. C, control group; A,
asthma group; D, dexamethasone treatment group; E-cad, E-cadherin;
VIM, vimentin; FSP-1, fibroblast-specific protein 1; OVA,
ovalbumin. |
We also examined ORMDL3 protein and mRNA levels in
lung tissue. Compared with the control group, ORMDL3 protein and
mRNA expression was significantly increased in lung tissues from
the asthma group upon OVA challenge (Fig. 4D, F; P<0.05). the treatment
group signifcantly reduced the mRNA and protein expression of
ORMDL3 (Fig. 4E, F) (P<0.01).
The expression of ORMDL3 mRNA was positively related to the
expression of FSP-1 mRNA and VIM mRNA, bronchiolar wall thickness,
and the percentage of collagen deposition but negatively related to
the expression of E-cad mRNA (as indicated by Pearson's correlation
coefficient, P<0.01) (Table
II).
| Table II.Pearson correlation analysis of
Orosomucoid-like 3 with E-cad, FPS-1, VIM, Wat/Pbm and the
percentage of collagen. |
Table II.
Pearson correlation analysis of
Orosomucoid-like 3 with E-cad, FPS-1, VIM, Wat/Pbm and the
percentage of collagen.
| Analysis | E-cad | FSP-1 | VIM | Wat/Pbm | Collagen (%) |
|---|
| Correlation
index | −0.959 | 0.971 | 0.961 | 0.977 | 0.972 |
| P-value | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
Transfection of bronchial epithelial
cell line 16HBE 14o- with ORMDL3 induces EMT in vitro
Because our in vivo studies demonstrated that
ORMDL3 expression increased in the airway epithelium along with
bronchial EMT development with increasing duration of OVA allergen
challenge, we examined whether ORMDL3 regulates the migration of
airway epithelial cell-like mesenchymal cells and the expression of
the EMT indicators FSP-1, VIM and E-cad in the human bronchial cell
line 16HBE 14o-. 16HBE14o- cells were transfected with GFP-vector
or GFP-ORMDL3, and the indicated protein levels were determined by
western blot analysis (Fig. 5A).
ORMDL3 expression was significantly lower in cells expressing
GFP-vector than in those expressing GFP-ORMDL3. We also observed
that cells expressing GFP-ORMDL3 had higher FSP-1 and VIM
expression and lower E-cad expression than those expressing
GFP-vector, as determined by western blotting (Fig. 5A) and real-time PCR (Fig. 5B). Immunofluorescence microscopy
revealed that the transfection of ORMDL3 into 16HBE 14o-cells
resulted in increased FSP-1 expression and decreased E-cad
expression (Fig. 5C, D). The
number of migrating cells was significantly increased among the
cells transfected with GFP-ORMDL3 compared with those transfected
with GFP-vector (Fig. 5E, F)
(568.7±5.7/HP vs. 403.0±9.5/HP).
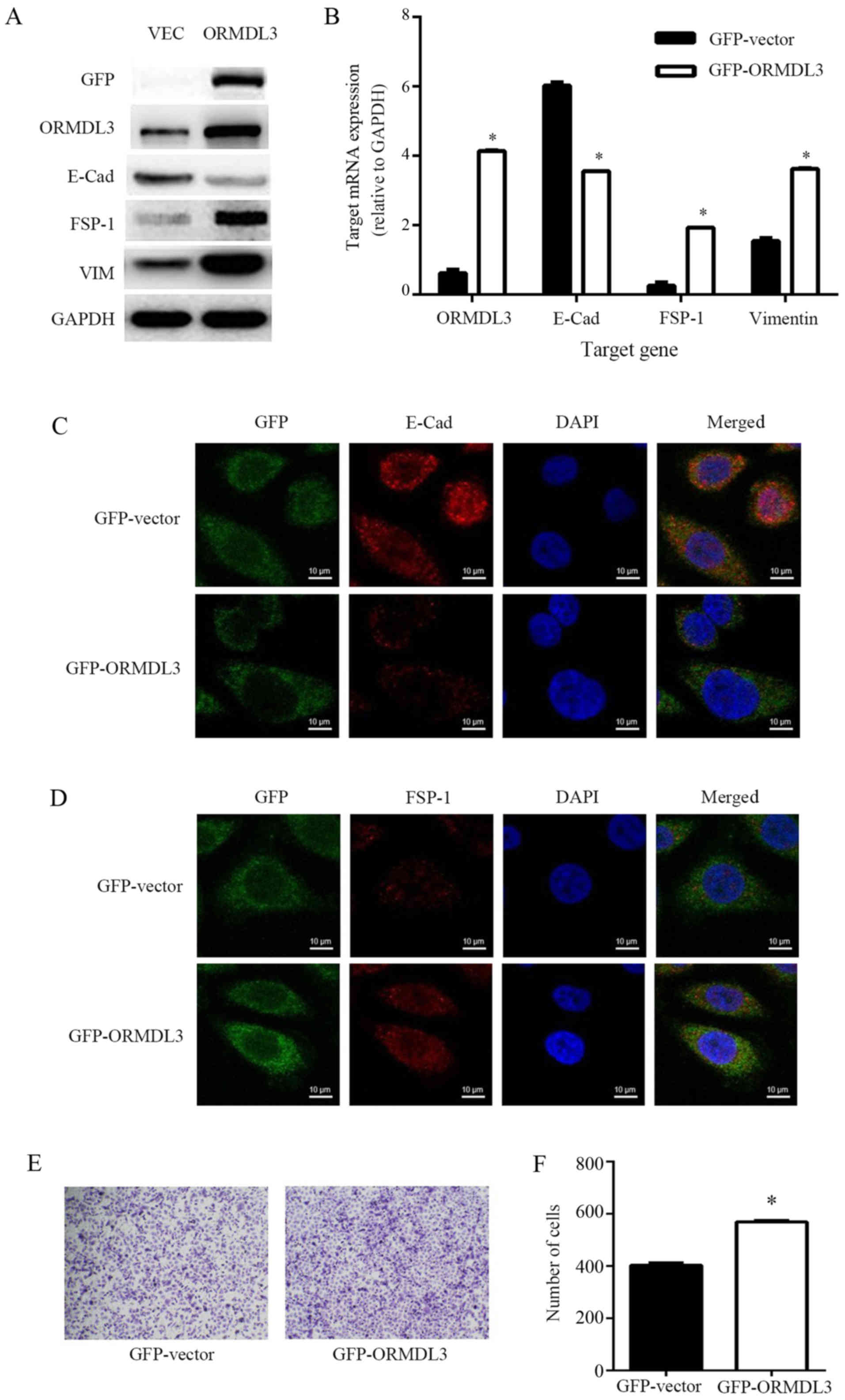 | Figure 5.Transfection of ORMDL3 in the
bronchial epithelial cell line 16HBE 14o- in vitro induced
EMT. (A) The levels of the indicated proteins were determined by
western blot analysis. Relative expression of the EMT indicators
ORMDL3, E-cad, FSP-1 and VIM in 16HBE 14o- cells transfected with
GFP-vector or GFP-ORMDL3 was assessed by western blotting and (B)
reverse transcription-quantitative polymerase chain reaction. (C)
Assessment of E-cad expression in 16 HBE 14o- cells transfected
with GFP-vector or GFP-ORMDL3 by double-label confocal
immunofluorescence microscopy (magnification, ×600). (D) Assessment
of FSP-1 expression in 16HBE 14o-cells transfected with GFP-vector
or GFP-ORMDL3 by confocal immunofluorescence microscopy
(magnification, ×600). (E) Observation of the migration of 16HBE
14o-cells transfected with GFP-vector or GFP-ORMDL3 in a Transwell
chamber using an inverted microscope (magnification, ×100) and (F)
determination of the number of 16HBE-14o cells in the two groups.
*P<0.01 vs. GFP-vector. ORMDL3, orosomucoid-like 3; EMT,
epithelial-mesenchymal transition; E-cad, E-cadherin; VIM,
vimentin; FSP-1, fibroblast-specific protein 1; GFP, green
fluorescent protein; VEC, GFP vector. |
Discussion
ORMDL3, a member of the ORM family, is located on
chromosome 1q. The gene order of this chromosome is highly
conserved from yeasts to vertebrates, and the ORMDL3 gene encodes a
tetraspanin consisting of 153 amino acids located in the
endoplasmic reticulum (9). The
biological function of ORMDL3 remains to be clarified, but a
relationship between ORMDL3 and endoplasmic reticulum stress has
been established. ORMDL3 was discovered by Moffatt et al
(2) in 2007 and is the gene most
definitively associated with pediatric asthma. The correlation
between ORMDL3 and asthma has been confirmed in a number of ethnic
populations (14), and increased
expression of ORMDL3 is observed in more than one third of
asthmatic children younger than 7 years of age (2). Epidemiology investigations related to
the expression of ORMDL3 and pediatric asthma conducted in Beijing,
Hong Kong and Chongqing in China have also suggested that ORMDL3 is
closely associated with asthma in Han Chinese children (15–17).
The focus of research on ORMDL3 has turned from
epidemiology to the mechanism of ORMDL3 in asthma. In this study,
using an asthmatic airway remodeling model prepared by OVA
sensitization and challenge, ORMDL3 expression was observed in
vivo during airway remodeling, and the EMT progression was
detected in the bronchial epithelium. The relationship between
ORMDL3 and EMT in bronchial epithelial cells was further confirmed
in vitro.
Immunohistochemistry indicated that ORMDL3 was
mainly expressed in bronchial epithelial cells in the asthma group
but not in the control group, suggesting that ORMDL3 expression was
associated with epithelial cell injury. Bronchial epithelial cells,
which represent the front line of defense in the airway, are
malleable. Following injury and during self-repair, some cells lose
cell polarity and intercellular junctions, resulting in
cytoskeletal remodeling, the emergence of interstitial cell
phenotypes, and consequently, EMT. In the present study, the most
severely injured portion of the lung tissue was the bronchial
epithelium. Examination of the bronchial epithelium in the asthma
group by light microscopy revealed cell edema, focal damage and
detachment, while transmission electron microscopy indicated
detachment of ciliated epithelial cells and disorganized cilia,
increased cytoplasmic vacuoles, a significant increase in the
secretory granules of goblet cell mucus, and slightly lax
intracellular junctions. These observations indicated that
significant damage and morphological changes occurred in bronchial
epithelial cells in the asthma group.
In this research, changes in EMT-related indicators
(E-cad, FSP-1, and VIM) were observed in lung tissues in the asthma
group. FSP-1 belongs to the family of S100 calcium-binding proteins
and is a typical marker of fibroblasts. It is mainly expressed at
the transitional stage between epithelial cells and fibroblasts.
FSP-1 is commonly used to detect the process of type II EMT because
FSP-1 is present in most epithelial cells with type II EMT at the
early stage of the process of organ fibrosis (18). E-cad is a marker of epithelial
cells, and VIM is a marker of mesenchymal cells.
Immunohistochemistry revealed increased FSP-1 in the bronchiolar
epithelium in the asthma group, suggesting that some bronchiolar
epithelial cells suffer EMT at an early stage. In addition,
decreased E-cad expression and increased VIM expression were
observed in the lung tissues of the asthma group, indicating
early-stage fibrosis. In the asthma group, the expression of E-cad
gradually declined and the expression of FSP-1 and VIM gradually
increased in lung tissues with increasing duration of OVA challenge
(3, 7 and 14 days), indicating aggravation of EMT. Most
importantly, the expression of ORMDL3 also increased with
increasing duration of OVA challenge and was positively correlated
with the degree of severity of airway remodeling and the expression
of EMT-related indicators. When a plasmid expressing ORMDL3 was
transfected into 16HBE-14o cells, E-cad expression decreased, FSP-1
and VIM expression increased, and cellular migration ability was
enhanced, indicating induction of EMT. Therefore, we infer that
ORMDL3 may be involved in the development of airway remodeling in
asthma, consistent with the results of previous studies. Miller
et al (7) have reported
that airway remodeling changes similar to those observed in asthma,
including bronchial smooth muscle thickening, increased fibers
under basilar membranes and mucus in the lumen, appear in the lung
tissues of ORMDL3 transgenic mice. Moreover, the present study
reveals a dynamic role of ORMDL3 in the progression of asthma, as
the level of ORMDL3 expression was correlated with the progression
of asthmatic airway remodeling, and suggests that the
overexpression of ORMDL3 may cause EMT. ORMDL3 might aggravate
asthmatic airway remodeling by causing bronchial epithelial EMT, a
novel aspect of the mechanism of ORMDL3 in asthma.
Yu et al (19) have also reported ORMDL3 was
associated with airway remodeling in asthma via the ERK/MMP-9
pathway. Our study had some similarity to Yu et al (19) about the methods, the purpose and
the results. Two papers both investigated the mechanism of ORMDL3
in airway remodeling in asthma. Yu et al (19) focuses on the study of the
relationship between ORMDL3 and ERK/MMP-9 channels, while our paper
focuses on the study of the relationship between ORMDL3 and EMT.
But Yu et al (19) did not
explain the direct relationship between the ORMDL3 and the
p-ERK/MMP-9 pathway in vivo, nor did it indicate whether
ORMDL3 regulates p-ERK/MMP-9 or p-ERK/MMP-9 regulates ORMDL3.
Otherwise, it is known that Inhaled corticosteroids is the first
choice to treat asthma. Studies has also been demonstrated that
budesonide inhibits the p-ERK/MMP-9 pathway and ORMDL3 (20,21).
p-ERK/MMP-9 pathway is one of the classic airway remodeling
pathways. Thus, budesonide treatment is more likely to directly
inhibit p-ERK/MMP-9, leading to reduction in airway remodeling,
rather than the reduced expression of ORMDL3 affect the p-ERK/MMP-9
pathway resulted in reduction in airway remodeling. There was lack
of evidence to prove the relationship between ORMDL3, p-ERK/MMP-9
pathway and airway remodeling in Yu et al's paper (19). Airway remodeling in asthma includes
smooth muscle cell proliferation and hypertrophy, extracellular
matrix deposition, basement membrane thickening, inflammatory cell
infiltration and gland proliferation, hypertrophy and
epithelial-mesenchymal transition. This study focuses on the effect
of ORMDL3 on airway remodeling in asthma by EMT. This paper shows
the airway remodeling in asthma in different periods, the
expression of ORMDL3 was gradually increased with EMT process
including airway wall thickness and collagen deposition, and the
epithelial cell marker (E-cad negative correlation, mesenchymal
cell markers FSP-1 and Vimentin positive correlation). In addition,
cell experiments in vitro have shown that overexpression of
ORMDL3 could induce epithelial EMT. Thus, the relationship between
ORMDL3 and airway remodeling and EMT was clearly demonstrated. The
similarity of the detection methods (H&E-staining,
Masson-staining and immunohistochemistry), and both of the two
specimens were lung tissues of BALB/c mice, thus, the pictures of
the two articles were similar. But carefully distinguish, there was
difference between the two papers. First of all, the image of color
and the shades of staining are different. Secondly, subjects of Yu
et al (19) were divided
into three groups (asthma group, budesonide treatment group,
control group). In our paper, subjects were also divided into 3
groups too (control group, asthma group and treatment group), and
according to the different time of allergen challenge 3rd, 7th,
14th, control group and asthma group divided into 6 groups (control
group for 3 days, control group for 7 days, control group for 14
days, and asthma group for 3 days, asthma group for 7 days, asthma
group for 14 days). Finally, our purpose was to observe the
relationship of ORMDL3 expression and morphological changes in
different period of airway remodling. And the research content and
results displayed in our pictures are different from Yu et
al (19).
The process of EMT is regulated by a variety of
cytokines and signaling pathways, but the signaling pathway by
which ORMDL3 affects bronchiolar EMT is unknown. Two major
mechanisms of ORMDL3 have been revealed in previous studies: 1)
ORMDL3 co-localizes with sarcoplasmic reticulum Ca-ATPase on
endoplasmic reticulum membranes (SERCA) to inhibit SERCA function,
increase calcium ions in the cytoplasm and reduce calcium irons in
the endoplasmic reticulum, leading to endoplasmic reticulum calcium
imbalance, subsequent endoplasmic reticulum stress, and the
unfolded protein response (22).
2) ORMDL3 combines with serine palmitoyltransferase to form the
SPOTS complex and decrease the enzymatic activity of serine
palmitoyltransferase. As this enzyme catalyzes the rate-limiting
step in sphingolipid production, formation of the complex with
ORMDL3 could negatively regulate sphingolipid production (23). EMT occurs when lung epithelial
cells are subjected to the endoplasmic reticulum stress response
(24), whereas abnormal
sphingolipids cause mast cell degranulation, bronchial
hyperreactivity and the migration of immune cells (25). However, whether ORMDL3 causes
bronchiolar epithelial EMT changes via the endoplasmic reticulum
stress reaction and inhibition of sphingomyelin production remains
to be further verified.
In summary, this study has revealed that ORMDL3 is
associated closely with asthmatic airway remodeling and the
progression of bronchiolar EMT and may cause bronchial epithelial
EMT. A further understanding of the role of the dynamic changes in
ORMDL3 in the development of asthma may guide clinical
applications. ORMDL3 may be useful as an index to monitor asthmatic
airway remodeling or its severity and is likely to be a new
precision medicine therapeutic target for asthma, especially for
refractory cases.
Acknowledgements
We would like to acknowledge the ‘Clinical Ability
Construction Projects of Liaoning Province’ (serial no.
LNCCC-C02-2015) for the financial support of this study.
References
|
1
|
Anderson GP: Endotyping asthma: New
insights into key pathogenic mechanisms in a complex, heterogeneous
disease. Lancet. 372:1107–1119. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Moffatt MF, Kabesch M, Liang L, Dixon AL,
Strachan D, Heath S, Depner M, von Berg A, Bufe A, Rietschel E, et
al: Genetic variants regulating ORMDL3 expression contribute to the
risk of childhood asthma. Nature. 448:470–473. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Balantic M, Rijavec M, Flezar M, Camlek T,
Hudoklin I, Kosnik M, Korosec P and Suskovic S: A polymorphism in
ORMDL3 is associated not only with asthma without rhinitis but also
with chronic obstructive pulmonary disease. J Investig Allergol
Clin Immunol. 23:256–261. 2013.PubMed/NCBI
|
|
4
|
Holgate ST: The sentinel role of the
airway epithelium in asthma pathogenesis. Immunol Rev. 242:205–219.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hackett TL: Epithelial-mesenchymal
transition in the pathophysiology of airway remodelling in asthma.
Curr Opin Allergy Clin Immunol. 12:53–59. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Johnson JR, Roos A, Berg T, Nord M and
Fuxe J: Chronic respiratory aeroallergen exposure in mice induces
epithelial-mesenchymal transition in the large airways. PLoS One.
6:e161752011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Miller M, Rosenthal P, Beppu A, Mueller
JL, Hoffman HM, Tam AB, Doherty TA, McGeough MD, Pena CA, Suzukawa
M, et al: ORMDL3 transgenic mice have increased airway remodeling
and airway responsiveness characteristic of asthma. J Immunol.
192:3475–3487. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Miller M, Tam AB, Cho JY, Doherty TA, Pham
A, Khorram N, Rosenthal P, Mueller JL, Hoffman HM, Suzukawa M, et
al: ORMDL3 is an inducible lung epithelial gene regulating
metalloproteases, chemokines, OAS, and ATF6. Proc Natl Acad Sci
USA. 109:16648–16653. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hjelmqvist L, Tuson M, Marfany G, Herrero
E, Balcells S and Gonzàlez-Duarte R: ORMDL proteins are a conserved
new family of endoplasmic reticulum membrane proteins. Genome Biol.
3:RESEARCH00272002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Meyer-Martin H, Reuter S and Taube C:
Mouse models of allergic airway disease. Methods Mol Biol.
1193:127–141. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shin YS, Takeda K and Gelfand EW:
Understanding asthma using animal models. Allergy Asthma Immunol
Res. 1:10–18. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
James AL, Hogg JC, Dunn LA and Paré PD:
The use of the internal perimeter to compare airway size and to
calculate smooth muscle shortening. Am Rev Respir Dis. 138:136–139.
1988. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Palmans E, Kips JC and Pauwels RA:
Prolonged allergen exposure induces structural airway changes in
sensitized rats. Am J Respir Crit Care Med. 161:627–635. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Galanter J, Choudhry S, Eng C, Nazario S,
Rodríguez-Santana JR, Casal J, Torres-Palacios A, Salas J, Chapela
R, Watson HG, et al: ORMDL3 gene is associated with asthma in three
ethnically diverse populations. Am J Respir Crit Care Med.
177:1194–1200. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Leung TF, Sy HY, Ng MC, Chan IH, Wong GW,
Tang NL, Waye MM and Lam CW: Asthma and atopy are associated with
chromosome 17q21 markers in Chinese children. Allergy. 64:621–628.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yu X, Yu C, Ren Z, Deng Y, Song J, Zhang H
and Zhou H: Genetic variants of 17q21 are associated with
childhood-onset asthma and related phenotypes in a northeastern Han
Chinese population: A case-control study. Tissue Antigens.
83:330–336. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhe J, Xin C, Qiang W and Hong L: Study of
Association between ORMDL3 expression level, life-style characters,
indoor air qualifies and childhood asthma in Beijing. J Med Res.
39:22–25. 2010.
|
|
18
|
Donato R, Cannon BR, Sorci G, Riuzzi F,
Hsu K, Weber DJ and Geczy CL: Functions of S100 proteins. Curr Mol
Med. 13:24–57. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yu F, Sun Y, Yu J, Ding Z, Wang J, Zhang
L, Zhang T, Bai Y and Wang Y: ORMDL3 is associated with airway
remodeling in asthma via the ERK/MMP-9 pathway. Mol Med Rep.
15:2969–2976. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rae MT, Price D, Harlow CR, Critchley HO
and Hillier SG: Glucocorticoid receptor-mediated regulation of MMP9
gene expression in human ovarian surface epithelial cells. Fertil
Steril. 92:703–708. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zou LP, Zhang X, Zhang Y, Xu XJ and Wang
TF: Down-regulatory effects of budesonide on expression of STAT6
and ORMDL3 in lung tissues of asthmatic mice. Zhongguo Dang Dai Er
Ke Za Zhi. 16:198–202. 2014.(In Chinese). PubMed/NCBI
|
|
22
|
Cantero-Recasens G, Fandos C,
Rubio-Moscardo F, Valverde MA and Vicente R: The asthma-associated
ORMDL3 gene product regulates endoplasmic reticulum-mediated
calcium signaling and cellular stress. Hum Mol Genet. 19:111–121.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Siow D, Sunkara M, Dunn TM, Morris AJ and
Wattenberg B: ORMDL/serine palmitoyltransferase stoichiometry
determines effects of ORMDL3 expression on sphingolipid
biosynthesis. J Lipid Res. 56:898–908. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tanjore H, Cheng DS, Degryse AL, Zoz DF,
Abdolrasulnia R, Lawson WE and Blackwell TS: Alveolar epithelial
cells undergo epithelial-to-mesenchymal transition in response to
endoplasmic reticulum stress. J Biol Chem. 286:30972–30980. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Breslow DK, Collins SR, Bodenmiller B,
Aebersold R, Simons K, Shevchenko A, Ejsing CS and Weissman JS: Orm
family proteins mediate sphingolipid homeostasis. Nature.
463:1048–1053. 2010. View Article : Google Scholar : PubMed/NCBI
|