Introduction
Worldwide, ovarian cancer is one of the most common
types of human gynecological tumor, and the morbidity and mortality
rate is increased compared with other gynecological malignancies
(1,2). A number of factors may lead to
tumorigenesis in ovarian cancer, demonstrating the complex
pathology of the disease (3). A
previous clinical study indicated that the incidence of ovarian
cancer is increasing worldwide (4). Currently, tumorectomy, radiotherapy
and chemotherapy are the primary therapies for patients with
ovarian cancer in the clinic. However, ineffective treatments and
treatment noncompliance frequently contribute to a worsening of
symptoms due to the apoptosis resistance of ovarian malignant cells
in patients with cancer (5).
Although novel treatments to improve clinical medicine have been
investigated, the improvements have had little effect on the
survival rate of patients with ovarian cancer (6).
Apoptosis resistance is important for tumor growth
and aggressiveness, and is caused by regulatory disorders of the
apoptotic signaling pathway (7).
Numerous molecules and proteins have been identified to be
associated with the biochemical processes underlying apoptosis
resistance, and may regulate the apoptotic signaling pathway in
various tumor cells (8–10). Cellular studies and clinical data
have suggested a direct association between the expression of the
anti-apoptotic gene survivin and the apoptotic susceptibility of
human ovarian cancer cells (11–13).
Resistance to radiotherapy and chemotherapy is the principal
challenge for the treatment of recurrent ovarian cancer, and
frequently causes varying degrees of immune system damage,
treatment failure and tumor metastasis. Therefore, understanding
the underlying mechanisms of radioresistance and chemoresistance in
ovarian cancer may contribute to the inhibition of apoptosis
resistance and the benefits of cancer therapy, in addition to the
development of innovative systemic therapies.
Apoptosis resistance, particularly increased
epithelial-mesenchymal transition (EMT), remains an intractable
clinical problem in the treatment of ovarian cancer (14). A previous study revealed that EMT
may affect the cell cycle, differentiation, survival and apoptosis,
regulated by transforming growth factor (TGF)-β in tumor cells
(15). In addition, TGF-β
upregulation may suppress mothers against decapentaplegic homolog
(Smad) and non-Smad signaling in mammary epithelial cells, leading
to EMT and the inhibition of growth arrest and apoptosis (16). Additionally, Chorna et al
(17) demonstrated that TGF-β
production may act as a natural immunosupressor, which is regarded
as an anti-apoptotic protein for doxorubicin. Research has
indicated that the liver kinase B1 (LKB1)-salt-inducible kinase 1
(SIK1) signaling pathway is associated with lung cancer cell
growth, and previous results have suggested that attenuating
LKB1-SIK1 may promote tumor invasion via upregulation of TGF-β
production in non-small cell lung cancer cells (18). However, the signaling pathway and
molecular mechanisms of LKB1-SIK1 has not been investigated in
ovarian cancer.
In the present study, to determine the role of
LKB1-SIK1 in ovarian cancer, the activity and expression levels of
LKB1-SIK1 were analyzed in ovarian cancer tissues with normal
adjacent tissues as the control. Migratory and invasive capacities
were evaluated using Transwell. The association between LKB1-SIK1,
EMP, apoptosis resistance and ovarian cancer cell growth was
investigated using western blotting, small interfering RNA (siRNA),
protein overexpression and immunofluorescence.
Materials and methods
Ethics statement
The present clinical investigation (no.
HMCH2010072508) was performed in strict accordance with the
recommendations in the Guide for Haidian Maternal and Child
Healthcare Center (Beijing, China) between May 2010 and October
2015. A total of 12 female patients with ovarian fibroma were
required to review trial protocols and amendments, and to provide
written informed consent. The present study was approved by the
ethics committee of Haidian Maternal and Child Healthcare Center.
The demographic and clinical pathological characteristics of the
patients are summarized in Table
I.
 | Table I.Characteristics of patients with
ovarian cancer. |
Table I.
Characteristics of patients with
ovarian cancer.
| Characteristic | Value |
|---|
| Patient no. | 12 |
| Age range,
years | 32.4–53.8 |
| Cancer type | Ovarian
fibroma |
| Tumor stage, n |
|
| I | 8 |
| II | 3 |
|
III | 1 |
| IV | 0 |
| History of
cancer | None |
| History of
allergy | None |
| Prior
treatment | None |
Cell culture and reagents
Ovarian cancer cells (1×107) were
isolated from patients with ovarian cancer using tumor cell
separation methods (19). Ovarian
cancer tissues and adjacent normal tissues were washed with PBS.
Tumor tissue was cut into pieces and separated into individual
cells using minimum essential medium (MEM) containing 5% pancreatin
and penicillin/streptomycin (2 mM) (both from Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Cells were maintained for 12 h
at 37°C in the presence of 5% CO2. Subsequently, cells
were filtered, collected and identified by microscopic
investigation (20). Ovarian
cancer cells were cultured in MEM (Gibco; Thermo Fisher Scientific,
Inc.) supplemented with 10% fetal bovine serum (Invitrogen; Thermo
Fisher Scientific, Inc.). Cells were cultured in a humidified
atmosphere containing 5% CO2 at 37°C in a cell culture
incubator (Gibco; Thermo Fisher Scientific, Inc.).
Western blotting
Ovarian cancer cells transfected with siRNA or the
eukaryotic expression vector for LKB1 were homogenized in lysate
buffer containing protease-inhibitor (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) and were centrifuged at 8,000 × g at 4°C for 10
min. The supernatant was used for analysis of the total protein
using a bicinchoninic protein assay kit (Gibco; Thermo Fisher
Scientific, Inc.). Protein samples (20 µg) were separated on 12%
sodium dodecyl sulfate polyacrylamide gels and transferred onto
polyvinylidene fluoride membranes (EMD Millipore, Billerica, MA,
USA) as previously described (21). Protein was blocked with 5% bovine
serum albumin reagent (Roche Diagnostics, Basel, Switzerland) for 1
h at 37°C. For western blotting, primary goat anti-human antibodies
against TGF-β (cat. no. ab31013), zinc-finger protein SNAI1 (cat.
no. ab53519), Snai2 (cat. no. ab187109), Twist-related protein 1
(cat. no. ab50887), zinc finger E-box-binding homeobox 1 (ZEB1;
cat. no. ab71286), E-cadherin (cat. no. ab76319), vimentin (VIM;
cat. no. ab137321), VEGF (cat. no. ab27278), angiotensin-1 (Ang-1;
cat. no. ab53951) and β-actin (cat. no. ab8226) (all 1:500
dilution; Abcam, Cambridge, UK), were incubated overnight at 4°C,
followed by incubation with horseradish peroxidase-conjugated
polyclonal anti-rabbit immunoglobulin G antibody (1:10,000, cat.
no. HAF008; R&D Systems, Inc., Minneapolis, MN, USA) for 1 h at
room temperature. A Ventana Benchmark automated staining system was
used for analyzing protein expression (Olympus BX51; Olympus,
Tokyo, Japan).
siRNA transfection
Ovarian cancer cells (1×106) were
transfected with HiPerFect reagent (Qiagen GmbH, Hilden, Germany),
according to the manufacturer's instructions (22). siRNA-TGF-β (100 pmol), siRNA-LKB1
(100 pmol) and siRNA-vector (100 pmol) were transfected into
ovarian cancer cells for 24 h at 37°C. The sequences of
siRNA-TGF-β, siRNA-LKB1 and siRNA-vector were designed and are
listed in Table II. siRNA
oligonucleotide pools containing three sequences targeting TGF-β or
LKB1 were purchased from Eurogentec, Ltd. (Liège, Belgium).
| Table II.Sequences of siRNAs. |
Table II.
Sequences of siRNAs.
| Name | Sense | Antisense |
|---|
| TGF-β |
5′-GGATACCAACTATTGCTTCAGCTCC-3′ |
5′-AGGCTCCAAATATAGGGGCAGGGTC-3′ |
| LKB1 |
5′-CTAGCTCAGACCGTTAGACGCCAGGACGGGCTGTCAGGCTGGCGCCTTTT-3′ |
5′-AAAAGGCGCCAGCCTGACAGCCCGTCCTGGCGTCTAACGGTCTGAGCTAG-3′ |
| Vector |
5′-AGAGGGAAATCGTGCGTGAC-3′ |
5′-CAATAGTGATGACCTGGCCGT-3′ |
Endogenous expression of LKB1
In order to establish stable ovarian cancer cells
with endogenous LKB1 expression, the eukaryotic expression vector
pCMVp-NEO-BAN (Takara Biotechnology Co., Ltd., Dalian, China) was
used to construct recombinant plasmids. LKB1 was cloned, sequenced
and recombined into pCMVp-NEO-BAN to construct pCMVp-NEO-LKB1
(pLKB1). The recombinant plasmid pCMVp-NEO-LKB1 was subsequently
transfected into ovarian cancer cells. pLKB1 (1.0 µg) or pvector
(1.0 µg) was transfected into cultured ovarian cancer cells
(5×106) using Lipofectamine 2000 (Sigma-Aldrich; Merck
KGaA), according to the manufacturer's instructions. Stable
LKB1-overexpressing ovarian cancer cells were selected by G418
screening (23). After 48 h
transfection, LKB1-overexpressing ovarian cancer cells were used to
subsequent experimentation.
Apoptosis analysis
Apoptosis analysis of ovarian tumor cells was
performed using flow cytometry. Human ovarian tumor cells
(5×106) were cultured in 6-well plates with paclitaxel
(2.0 mg/ml; Sigma-Aldrich; Merck KGaA) for 48 h to achieve the
maximal apoptosis rate. Ovarian tumor cells were harvested at 48 h
post-treatment by trypsinization. Ovarian tumor cells were
subsequently washed in cold PBS and adjusted to 1×106
cells/ml with PBS. Following double staining with fluorescein
isothiocyanate (FITC)-Annexin V and propidium iodide using the FITC
Annexin V Apoptosis Detection kit I (BestBio Biotechnology,
Shanghai, China), cells were analyzed using a FACScan®
flow cytometer equipped with Cell Quest software (version 3.3; BD
Biosciences, San Jose, CA, USA), according to manufacturer's
instructions, to detect apoptosis in ovarian tumor cells. All
experiments were performed in triplicate.
MTT cytotoxicity assays
Ovarian cancer cells were incubated
(1×103) with paclitaxel or PBS in 96-well plates for 48
h in triplicate. Subsequently, 20 µl MTT (5 mg/ml) in PBS solution
was added to each well, the plate was further incubated for 4 h.
Most of the medium was removed and 100 µl dimethyl sulfoxide was
added into the wells to solubilize the crystals. The OD was
measured using an ELISA reader (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) at wavelength of 450 nm.
Histological, immunohistochemical and
immunofluorescence staining analyses
Ovarian tumor tissues were fixed in situ
overnight in 10% buffered formalin. The fixed tissues were cut
mid-sagittal and being embedded in paraffin (4 µm thickness) using
standard protocols. Hematoxylin and eosin staining was used to
visualize the area of myocardial infarction after treatment with
matrine. Immunohistochemical staining was performed using an
avidin-biotin-peroxidase technique. Tumor sections (4 µm) were
deparaffinized in xylene, dehydrated through graded ethanol and
treated with 0.3% hydrogen peroxide in methanol for 30 min at 37°C.
Paraffin-embedded ovarian normal tissue and tumor sections were
prepared and epitope retrieval was performed at 95°C for 15 min for
further analysis. The paraffin sections were treated with hydrogen
peroxide (3%) for 10–15 min, which subsequently was blocked with a
blocking solution (5% skim milk powder) for 10–15 min at 37°C.
Subsequently, the sections were incubated in goat anti-human
anti-Snai2 (1:1,000, cat. no. ab187109), Twist (1:1,000, cat. no.
ab50887), ZEB1 (1:1,000, cat. no. ab71286), LKB1 (1:1,000, cat. no.
ab15095), SIK1 (1:1,000, cat. no. ab64428), phosphorylated (p-)LKB1
(1:1,000, cat. no. ab63473), p-SIK1 (1:1,000, cat. no. ab217809),
TGF-β (1:1,000, cat. no. ab31013) and VIM (1:1,000, cat. no.
ab137321) at 4°C for 12 h. All sections were washed three times and
incubated with secondary rabbit anti-goat antibodies (1:2,000, cat.
no. ab150117; Abcam) for 1 h at 37°C. For immunofluorescence,
ovarian tumor cells were stained with goat anti-human vascular
endothelial growth factor and angiopoietin-1 antibodies.
Additionally, a terminal deoxynucleotidyl transferase dUTP nick end
labeling assay was performed using a Peroxidase Apoptosis Detection
kit (Chemicon; EMD Millipore). All sections were observed in six
random fields in the confocal microscope at magnification, ×40
(Nikon E400, Nikon Corporation, Tokyo, Japan).
Analysis of the cell cycle
To analyze the effects of LKB1 overexpression on the
cell cycle stage of ovarian cancer cells, flow cytometry was
performed. Exponentially, culturing ovarian cancer cells
(1×106) or LKB1 overexpression were cultured for 24 h at
37°C. Cells were washed and trypsinized and rinsed with
phosphate-buffered saline (PBS). All cells were fixed in 75%
ice-cold ethanol for 5 min and then washed with PBS three times.
The fixed cells were washed with RNase A (20 µg ml/l, Fermentas;
Thermo Fisher Scientific, Inc.) and stained with propidium iodide
(20 µg ml/l, Sigma-Aldrich; Merck KGaA) for 10 min at 37°C. The
percentages of cells in G1 phase were analyzed using BD FACSCalibur
(Becton Dickinson; BD Biosciences, San Jose, CA, USA).
Cell invasion and migration
assays
Ovarian tumor cells subjected to different
treatments (LKB1 overexpression or TGF-β inhibition) were used to
analyze invasion and migration. Migration and invasion assays in
ovarian tumor cells were conducted in a 24-well MEM culture plate
with chamber inserts (BD Biosciences) with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.) for 12 h at 37°C. For
migration assays, 1×103 cells/well ovarian tumor cells
were placed into the upper chamber with a non-coated membrane. For
the invasion assays, cells (1×103 cells/well) were
placed into the upper chamber with a Matrigel-coated membrane.
Matrigel were fixed with 4% formaldehyde and stained with
4′,6-diamidino-2-phenylindole as well as counted in 6 random fields
under a microscope. Invasion and migration were calculated in at
least three randomly stained fields under a light microscope (Nikon
E400; Nikon Corporation).
Statistical analysis
All data are presented as the mean ± standard error
of triplicate samples, and analyses were performed using Prism 6.0
software (GraphPad Software, Inc., La Jolla, CA, USA). Statistical
differences between experimental groups were analyzed using
Student's t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Analysis of the expression levels of
EMT proteins, LKB1 and SIK1 in ovarian tumor tissues
The expression levels of EMT molecules, LKB1 and
SIK1 were measured in ovarian tumor tissues. As presented in
Fig. 1A, expression of the EMT
pathway components Snai2, Twist and ZEB1 was upregulated in ovarian
tumor tissues compared with normal ovarian tissues. The results in
Fig. 1B and C demonstrated that
the expression and phosphorylation levels of LKB1 and SIK1 were
downregulated in ovarian tumor tissues compared with normal ovarian
tissues. It was additionally observed that the expression levels of
TGF-β and VIM were markedly increased in ovarian tumor tissues
compared with normal ovarian tissues (Fig. 1D). The results of the present study
demonstrated that the knockdown of LKB1 expression promoted TGF-β
and Snai1 expression in ovarian tumor cells (Fig. 1E). Additionally, it was observed
that the knockdown of LKB1 promoted the apoptosis resistance of
ovarian tumor cells treated with paclitaxel (Fig. 1F). The present results suggested
that the expression levels of EMT, LKB1 and SIK1 were increased in
ovarian tumor tissues, which may be associated with the aberrant
growth and aggressiveness of ovarian tumor cells.
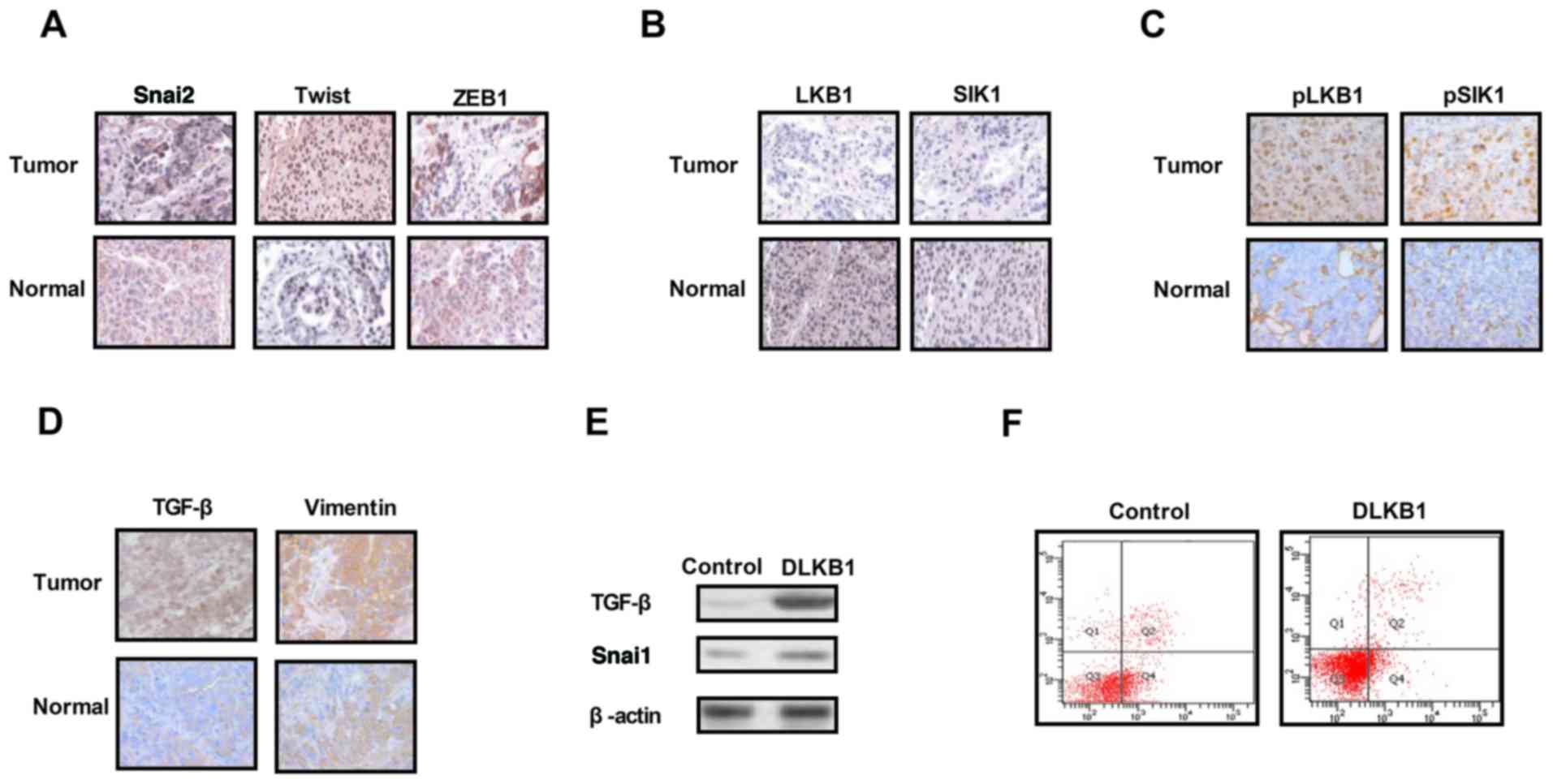 | Figure 1.Expression levels of TGF-β, EMT, LKB1
and SIK1 in ovarian tumor tissues. (A) Expression levels of EMT
components including Snai2, Twist and ZEB1 between ovarian tumor
tissues and normal ovarian tissues. (B) Expression levels of LKB1
and SIK1 between ovarian tumor tissues and normal ovarian tissues.
(C) Phosphorylation levels of LKB1 and SIK1 in ovarian tumor
tissues compared with normal ovarian tissues. (D) Expression levels
of TGF-β and vimentin in ovarian tumor tissues. (E) LKB1 knockdown
suppressed Snai1 expression in ovarian tumor cells. (F) LKB1
knockdown decreased the apoptotic resistance of ovarian tumor cells
treated with paclitaxel. TGF-β, transforming growth factor-β;
Snai2, zinc-finger protein SNAI2; EMT, epithelial-mesenchymal
transition; LKB1, liver kinase B1; SIK1, salt-inducible kinase 1;
Twist, Twist-related protein 1; ZEB1, zinc finger E-box-binding
homeobox 1; DLKB1, downregulated LKB1. Magnification, ×40. |
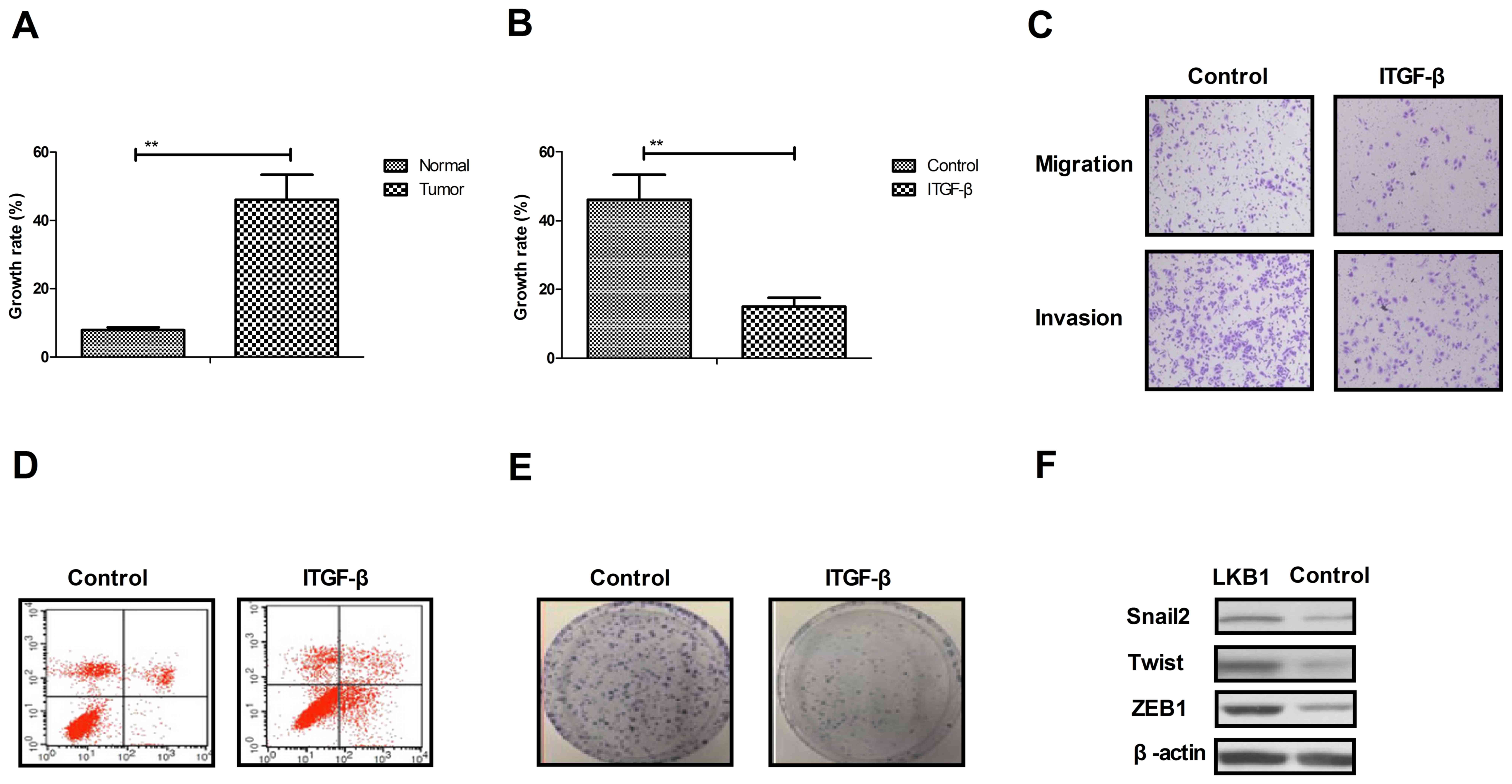
Analysis of growth and apoptotic
resistance of ovarian tumor cells following inhibition of TGF-β
expression
As presented in Fig.
2A, the growth rate of ovarian tumor cells isolated from
clinical patients was increased compared with normal ovarian cells.
Proliferation and migration assays demonstrated that growth and
aggressiveness was inhibited by the inhibition of TGF-β expression
in ovarian tumor cells (Fig. 2B and
C). Apoptosis experiments demonstrated that the inhibition
TGF-β expression by siRNA promoted apoptotic sensitivity in cells
treated with paclitaxel for 48 h (Fig.
2D). It was observed that the inhibition of TGF-β expression by
siRNA inhibited the proliferation of ovarian tumor cells compared
to si-vector-transfected cells (Fig.
2E). Additionally, it was observed that the inhibition of TGF-β
expression suppressed the expression levels of Snai2, Twist and
ZEB1, as determined by western blotting (Fig. 2F). The results of the present study
suggested that TGF-β expression may be associated with the growth
and apoptotic resistance of ovarian tumor cells.
LKB1 upregulation stimulates SIK1
expression and inhibits the EMT signaling pathway in ovarian tumor
cells
The present study further analyzed the influences of
LKB1 on TGF-β expression and the EMT signaling pathway in ovarian
tumor cells. The results in Fig.
3A demonstrated that SIK1 expression was promoted by LKB1
upregulation in ovarian tumor cells, as determined by
immunofluorescence analysis. Western blotting demonstrated that
TGF-β expression was decreased by LKB1 (Fig. 3B). The apoptosis assay indicated
that the apoptosis sensitivity of ovarian tumor cells was increased
following upregulation of LKB1 (Fig.
3C). In addition, the results demonstrated that the expression
of important regulatory factors in the EMT pathway, E-cad, VIM,
Snail2 and γH2AX, was downregulated by LKB1 upregulation in ovarian
tumor cells (Fig. 3D). Growth,
migration and invasion assays demonstrated that growth and
aggressiveness was inhibited following overexpression of LKB1
expression in ovarian tumor cells (Fig. 3E and F). The present findings
suggested that activation of the LKB1 signaling pathway may inhibit
the TGF-β-mediated EMT pathway and decrease growth, aggressiveness
and apoptosis resistance in ovarian carcinoma cells.
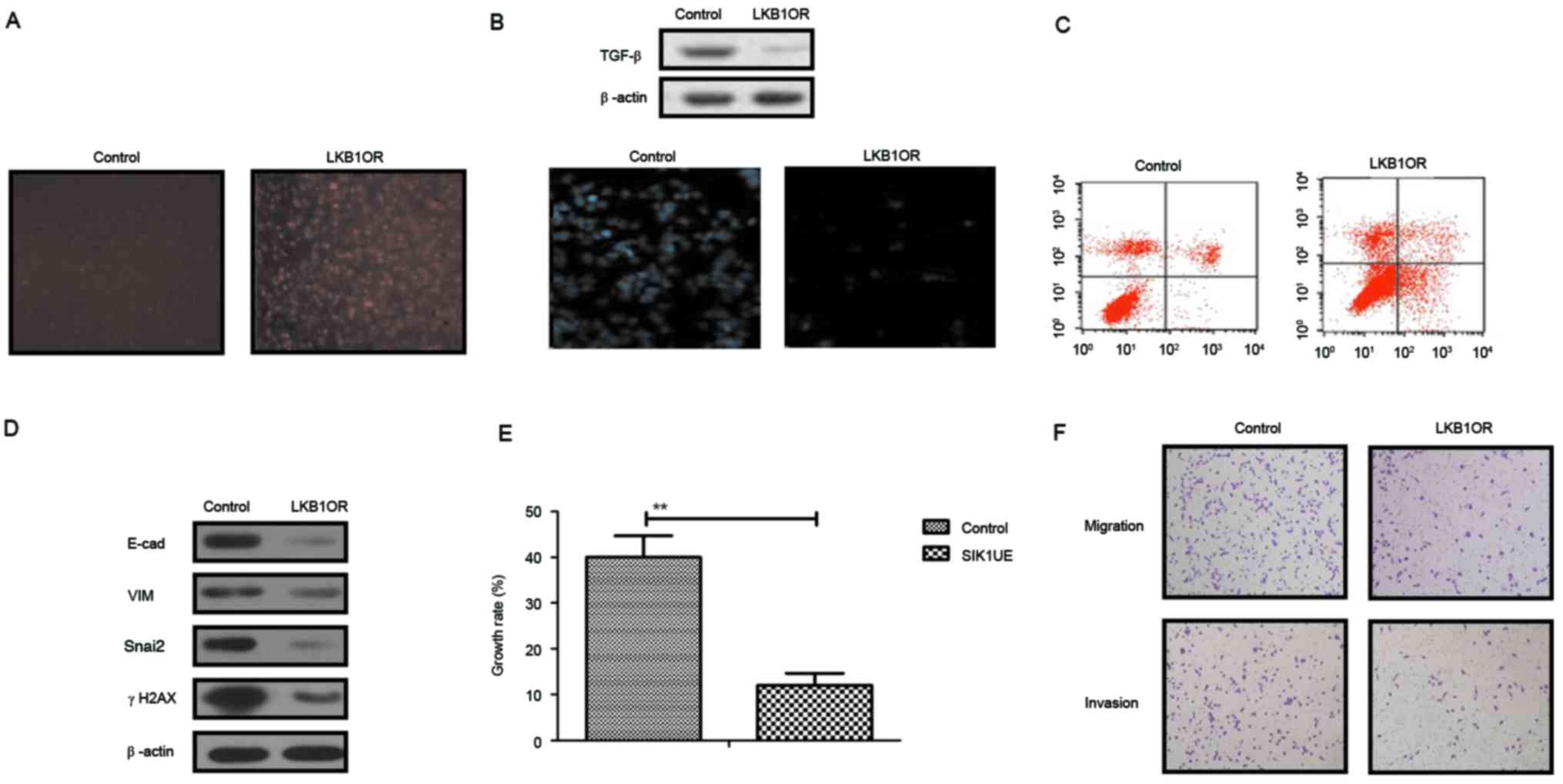 | Figure 3.Effects of LKB1 on SIK1 expression
and the EMT signaling pathway in ovarian tumor cells. (A) SIK1
expression was promoted by LKB1 upregulation in ovarian tumor
cells, as determined by immunofluorescence. (B) LKB1 overexpression
suppressed TGF-β expression in ovarian tumor cells. (C) LKB1
upregulation promoted apoptotic sensitivity in ovarian tumor cells
treated with paclitaxel. (D) LKB1 overexpression decreased E-cad,
VIM, Snai2 and γH2AX expression in ovarian tumor cells. (E)
Upregulated SIK1 suppressed the growth of ovarian tumor cells
compared with the control. (F) LKB1 overexpression inhibited the
aggressiveness of ovarian tumor cells. **P<0.01. LKB1, liver
kinase B1; SIK1, salt-inducible kinase 1; Snai2, zinc-finger
protein SNAI2; VIM, vimentin; E-cad, E-cadherin; TGF-β,
transforming growth factor-β; γH2AX, γ-histone H2AX; LKB1OR, LKB1
overexpression; SIK1UE, SIK1 upregulation. |
LKB1 overexpression inhibits cell
cycle and apoptosis resistance-associated protein expression in
ovarian tumor cells
The present study investigated the effects of LKB1
on cells survival- and apoptosis resistance-associated protein
expression in ovarian tumor cells. The results demonstrated that
LKB1 overexpression inhibited the survival and cell cycle
progression of tumor cells treated with paclitaxel, compared with
control cells (Fig. 4A and B).
Western blotting demonstrated that the expression of Bcl-2, Bax and
p53 was inhibited by LKB1 overexpression in ovarian tumor cells
compared with control cells (Fig.
4C). However, caspase-3, caspase-8 and Apaf-1 expression was
increased by LKB1 overexpression in ovarian tumor cells compared
with control cells (Fig. 4D). The
expression of aggressiveness-associated proteins FN and RNF-8 was
decreased by LKB1 overexpression in ovarian tumor cells (Fig. 4E). Additionally, the results
demonstrated that the expression levels of vascular endothelial
growth factor (VEGF) and angiotensin-1 (Ang-1) were downregulated
by LKB1 overexpression, as determined by immunofluorescence
analysis (Fig. 4F). The present
results suggested that LKB1 overexpression may inhibit
aggressiveness- and apoptosis resistance-associated protein
expression in ovarian tumor cells.
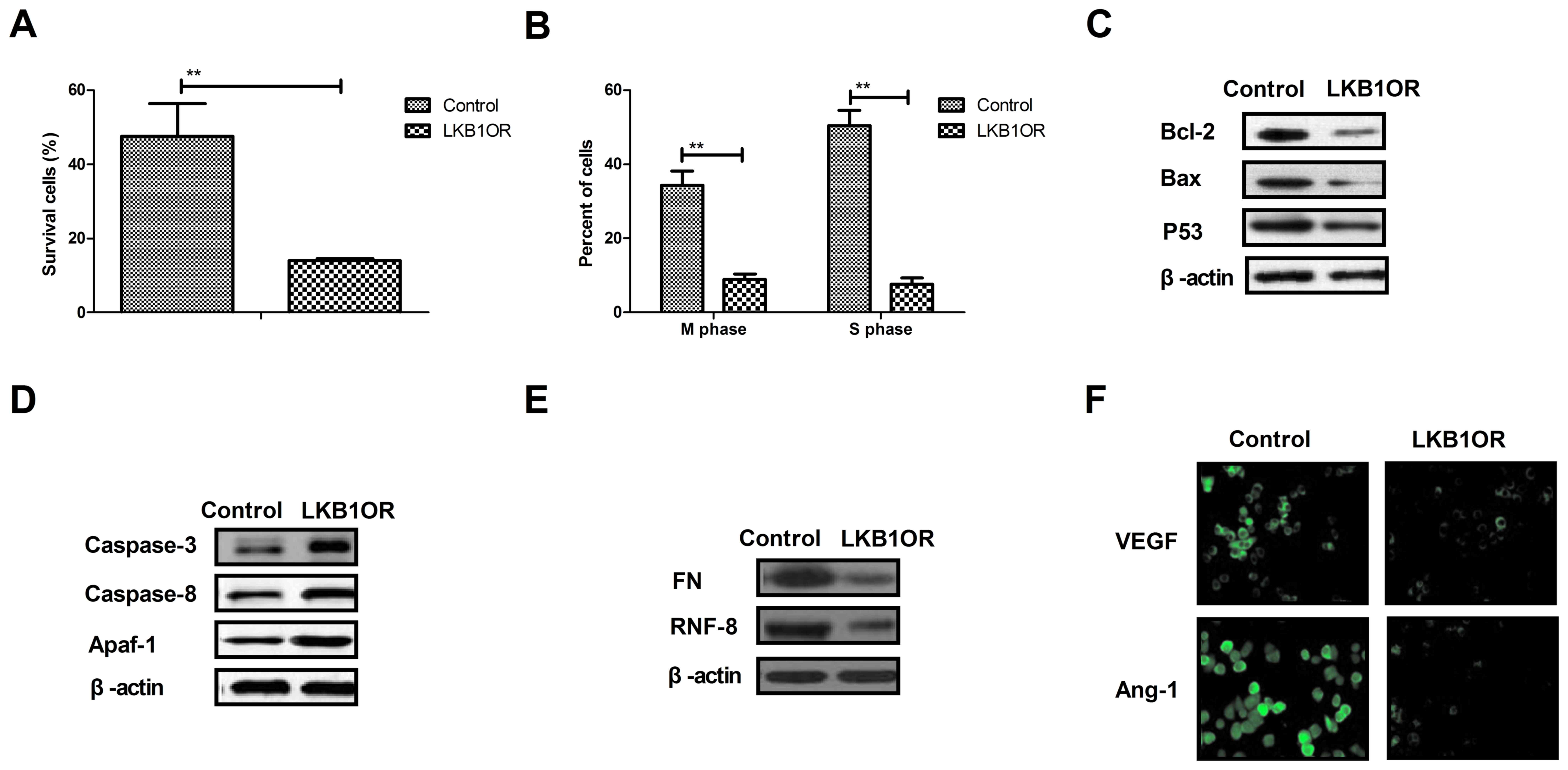 | Figure 4.Effects of LKB1 overexpression on
cell cycle- and apoptosis resistance-associated protein expression
in ovarian tumor cells. (A) LKB1 overexpression decreased the
survival rate of ovarian tumor cells treated with paclitaxel. (B)
LKB1 overexpression promoted the cell cycle arrest in ovarian tumor
cells treated with paclitaxel. (C) LKB1 overexpression decreased
the expression of Bcl-2, Bax and p53 in ovarian tumor cells, as
determined by western blotting. (D) LKB1 overexpression promoted
the expression of caspase-3, caspase-8 and Apaf-1 in ovarian tumor
cells, as determined by western blotting. (E) LKB1 overexpression
decreased the expression of FN and RNF-8 in ovarian tumor cells.
(F) LKB1 upregulation downregulated the expression levels of VEGF
and Ang-1 in ovarian tumor cells. **P<0.01. LKB1, liver kinase
B1; LKB1OR, LKB1 overexpression; Bcl-2, apoptosis regulator Bcl-2;
Bax, apoptosis regulator BAX; p53, cellular tumor antigen p53;
Apaf-1, apoptotic protease-activating factor 1; FN, fibronectin;
RNF-8, E3 ubiquitin-protein ligase RNF-8; VEGF, vascular
endothelial growth factor; Ang-1, angiotensin-1. |
Discussion
Ovarian cancer is associated with poor prevention,
intractable malignancy and a poor prognosis (24). Traditional treatments, including
radiotherapy, chemotherapy and surgery, are limited to palliative
approaches for patients with advanced ovarian cancer (25,26).
Although novel therapeutic agents and protocols for patients with
ovarian cancer have been proposed in previous reports, the
mortality and survival rates remain poor due to an increased rate
of recurrence and metastasis following surgical resection (27). It has been reported that intrinsic
and acquired resistance to anticancer treatments has been
recognized to be a notable impediment to favorable outcomes in the
clinic (28,29). The present study investigated the
growth and aggressiveness of clinical ovarian cancer cells and
analyzed the potential molecular mechanisms of apoptotic resistance
in cells treated with a chemotherapeutic drug. Previous studies
have suggested that ovarian cells may survive exposure to
chemotherapeutic drug treatment and may display cancer stem cell
and EMT-positive phenotypes (30,31).
Consistent with the results in the published literature, an
additional previous study demonstrated that the apoptotic
resistance of ovarian cancer was induced by the EMT phenotype and
expression of EMT-associated proteins (32). Notably, the present findings
suggested that the inhibition of LKB1-SIK1 may reverse apoptotic
resistance in ovarian cancer cells through the TGF-β-mediated EMT
signaling pathway.
Previous studies have indicated that the development
of anti-angiogenic drugs for the treatment of patients with ovarian
cancer is an emerging field of oncology hoping to enter the
preclinical stage of clinical trials (33–35).
Additionally, many patients with advanced ovarian cancer respond
poorly to traditional treatments or experience limited benefit from
these treatments (36). The
present study demonstrated an association between the EMT and SIK1
signaling pathways in ovarian carcinoma cells. EMT leads to
cellular heterogeneity and supports tumor engraftment, which has
been associated with underlying tumor heterogeneity and growth,
metastasis and progression of ovarian cancer (37). Tang et al (38) demonstrated that inhibiting
vasculogenic mimicry formation by reducing EMT may contribute to
tumor apoptosis in ovarian cancer. In the present study, the
results indicated that key regulatory factors in the EMT signaling
pathway were upregulated in clinical ovarian tumor tissues,
resulting in rapid growth and aggressiveness of tumor cells.
Inhibition of TGF-β expression led to an inhibition of growth and
aggressiveness, in addition to a promotion of apoptosis, in ovarian
carcinoma cells treated with paclitaxel. The present results
demonstrated that VEGF and Ang-1 were downregulated by LKB1
overexpression, suggesting that LKB1 may exert regulatory effects
on ovarian cancer cell growth.
A previous study demonstrated that the LKB1-SIK1
signaling pathway was suppressed in ovarian carcinoma cells
compared with normal ovarian cells, which led to activation of the
EMT signaling pathway (18). In
the present study, it was observed that inhibition of TGF-β
expression promoted paclitaxel-induced apoptosis and suppressed
tumor metastasis-associated protein expression (Snai2, Twist and
ZEB1) in ovarian tumor cells. Cha et al (39) suggested that the binding of
transcription elongation factor A protein 3 to TGF-β receptor I may
induce apoptosis regulated by the Smad- and mitogen-activated
protein kinase-dependent pathways in ovarian cancer cells. The
results of the present study indicated that the expression of
LKB1-SIK1 was suppressed in ovarian carcinoma cells, and that
increasing LKB1 expression was able to downregulate the expression
of TGF-β, and EMT and anti-apoptosis proteins, and to decrease
apoptotic resistance in ovarian carcinoma cells. A previous study
demonstrated that the knockdown of the LKB1 tumor suppressor gene
may potentially induce papillary serous ovarian cancer in the
ovarian surface epithelium (40).
An additional study indicated that suppression of the
LKB1-p53-p21/WAF1 pathway may promote the conversion of normal
ovarian cancer cells to cancer stem cells via regulation of miR-17
expression (41). Additionally, a
study reported that the restoration of SIK1 expression led to an
inhibition of proliferation, which further increased the
understanding of the pathogenesis and progression of ovarian cancer
(42). These previous reports
suggested that the expression levels of LKB1 may be decreased in
ovarian cancer cells. The results of the present study demonstrated
that the upregulation of LKB1 promoted SIK1 expression and markedly
suppressed the growth and aggressiveness of ovarian cancer cells.
Upregulation of LKB1 additionally promoted apoptosis in ovarian
carcinoma cells. The results also demonstrated that knockdown of
LKB1 further promoted the expression of TGF-β and EMT proteins,
which downregulated the chemosensitivity of ovarian carcinoma
cells. In addition, overexpression of LKB1 in ovarian carcinoma
cells increased the chemosensitivity of the cells, resulting in a
marked inhibition of migration and invasion. The results of the
present study demonstrated that LKB1 overexpression may serve an
inhibitory role in SIK1 and apoptosis resistance-associated protein
expression in ovarian tumor cells, indicating that LKB1 may be a
potential anticancer molecule.
In conclusion, the present study demonstrated that
the expression of LKB1 and SIK1 was downregulated, while TGF-β and
EMT protein expression levels were upregulated in clinical ovarian
tumor tissues and cells. The results of the present study
demonstrated that targeting the LKB1-SIK1 signal pathway may
suppress the TGF-β-mediated EMT pathway, which may provide a
therapeutic target for ovarian cancer. The apoptosis resistance of
ovarian cancer cells treated with paclitaxel was improved by
increasing LKB1 expression, resulting in the suppression of growth
and aggressiveness. The results of the present study additionally
indicated that inhibition of TGF-β markedly suppressed EMT- and
metastasis-associated protein expression in ovarian cancer cells.
Notably, it was suggested that enhanced LKB1-SIK1 signaling may
inhibit the TGF-β-mediated EMT signaling pathway and the
chemoresistance of ovarian cancer cells, which may subsequently
contribute to limited metastatic potential, suggesting that
targeting the LKB1-SIK1-TGF-β-EMT signaling pathways may be a
promising therapeutic option for promoting the chemosensitivity and
inhibiting the growth and metastasis of ovarian cancer cells.
References
|
1
|
Heidemann LN, Hartwell D, Heidemann CH and
Jochumsen KM: The relation between endometriosis and ovarian cancer
- a review. Acta Obstet Gynecol Scand. 93:20–31. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Petrillo M, Legge F, Ferrandina G,
Monterisi A, Pedone Anchora L and Scambia G: Fertility-sparing
surgery in ovarian cancer extended beyond the ovaries: A case
report and review of the literature. Gynecol Obstet Invest. 77:1–5.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kobayashi O, Sugiyama Y, Cho H, Tsuburaya
A, Sairenji M, Motohashi H and Yoshikawa T: Clinical and
pathological study of gastric cancer with ovarian metastasis. Int J
Clin Oncol. 8:67–71. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Raavé R, de Vries RB, Massuger LF, van
Kuppevelt TH and Daamen WF: Drug delivery systems for ovarian
cancer treatment: A systematic review and meta-analysis of animal
studies. PeerJ. 3:e14892015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gallardo-Rincón D, Espinosa-Romero R,
Muñoz WR, Mendoza-Martínez R, Villar-Álvarez SD, Oñate-Ocaña L,
Isla-Ortiz D, Márquez-Manríquez JP, Apodaca-Cruz Á and
Meneses-García A: Epidemiological overview, advances in diagnosis,
prevention, treatment and management of epithelial ovarian cancer
in Mexico. Salud Publica Mex. 58:302–308. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ganesan P, Kumar L, Hariprasad R, Gupta A,
Dawar R and Vijayaraghavan M: Improving care in ovarian cancer: The
role of a clinico-pathological meeting. Natl Med J India.
21:225–227. 2008.PubMed/NCBI
|
|
7
|
Jin F, Li HS, Zhao L, Wei YJ, Zhang H, Guo
YJ, Pang R, Jiang XB and Zhao HY: Expression of anti-apoptotic and
multi-drug resistance-associated protein genes in cancer stem cell
isolated from TJ905 glioblastoma multiforme cell line. Zhonghua Yi
Xue Za Zhi. 88:2312–2316. 2008.(In Chinese). PubMed/NCBI
|
|
8
|
Hamada S, Masamune A, Miura S, Satoh K and
Shimosegawa T: miR-365 induces gemcitabine resistance in pancreatic
cancer cells by targeting the adaptor protein SHC1 and
pro-apoptotic regulator BAX. Cell Signal. 26:179–185. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shiota M, Yokomizo A and Naito S:
Pro-survival and anti-apoptotic properties of androgen receptor
signaling by oxidative stress promote treatment resistance in
prostate cancer. Endocr Relat Cancer. 19:R243–R253. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang TM, Barbone D, Fennell DA and
Broaddus VC: Bcl-2 family proteins contribute to apoptotic
resistance in lung cancer multicellular spheroids. Am J Respir Cell
Mol Biol. 41:14–23. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zaffaroni N, Pennati M, Colella G, Perego
P, Supino R, Gatti L, Pilotti S, Zunino F and Daidone MG:
Expression of the anti-apoptotic gene survivin correlates with
taxol resistance in human ovarian cancer. Cell Mol Life Sci.
59:1406–1412. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xing H, Weng D, Chen G, Tao W, Zhu T, Yang
X, Meng L, Wang S, Lu Y and Ma D: Activation of
fibronectin/PI-3K/Akt2 leads to chemoresistance to docetaxel by
regulating survivin protein expression in ovarian and breast cancer
cells. Cancer Lett. 261:108–119. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liguang Z, Peishu L, Hongluan M, Hong J,
Rong W, Wachtel MS and Frezza EE: Survivin expression in ovarian
cancer. Exp Oncol. 29:121–125. 2007.PubMed/NCBI
|
|
14
|
Cheng JC, Auersperg N and Leung PC:
TGF-beta induces serous borderline ovarian tumor cell invasion by
activating EMT but triggers apoptosis in low-grade serous ovarian
carcinoma cells. PLoS One. 7:e424362012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Song J: EMT or apoptosis: A decision for
TGF-beta. Cell Res. 17:289–290. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gal A, Sjöblom T, Fedorova L, Imreh S,
Beug H and Moustakas A: Sustained TGF beta exposure suppresses Smad
and non-Smad signalling in mammary epithelial cells, leading to EMT
and inhibition of growth arrest and apoptosis. Oncogene.
27:1218–1230. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chorna I, Bilyy R, Datsyuk L and Stoika R:
Comparative study of human breast carcinoma MCF-7 cells differing
in their resistance to doxorubicin: effect of ionizing radiation on
apoptosis and TGF-beta production. Exp Oncol. 26:111–117.
2004.PubMed/NCBI
|
|
18
|
Yao YH, Cui Y, Qiu XN, Zhang LZ, Zhang W,
Li H and Yu JM: Attenuated LKB1-SIK1 signaling promotes
epithelial-mesenchymal transition and radioresistance of non-small
cell lung cancer cells. Chin J Cancer. 35:502016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kruger W, Jung R, Kröger N, Gutensohn K,
Fiedler W, Neumaier M, Jänicke F, Wagener C and Zander AR:
Sensitivity of assays designed for the detection of disseminated
epithelial tumor cells is influenced by cell separation methods.
Clin Chem. 46:435–436. 2000.PubMed/NCBI
|
|
20
|
Zhang S, Balch C, Chan MW, Lai HC, Matei
D, Schilder JM, Yan PS, Huang TH and Nephew KP: Identification and
characterization of ovarian cancer-initiating cells from primary
human tumors. Cancer Res. 68:4311–4320. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wai-Hoe L, Wing-Seng L, Ismail Z and
Lay-Harn G: SDS-PAGE-based quantitative assay for screening of
kidney stone disease. Biol Proced Online. 11:145–160. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Scherer O, Maeß MB, Lindner S, Garscha U,
Weinigel C, Rummler S, Werz O and Lorkowski S: A procedure for
efficient non-viral siRNA transfection of primary human monocytes
using nucleofection. J Immunol Methods. 422:118–124. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Renshaw A and Elsheikh TM: A validation
study of the Focalpoint GS imaging system for gynecologic cytology
screening. Cancer Cytopathol. 121:737–738. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kobayashi M, Chiba A, Izawa H, Yanagida E,
Okamoto M, Shimodaira S, Yonemitsu Y, Shibamoto Y, Suzuki N and
Nagaya M: DC-vaccine study group at the Japan Society of Innovative
Cell Ther: The feasibility and clinical effects of dendritic
cell-based immunotherapy targeting synthesized peptides for
recurrent ovarian cancer. J Ovarian Res. 7:482014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ebell MH, Culp MB and Radke TJ: A
systematic review of symptoms for the diagnosis of ovarian cancer.
Am J Prev Med. 50:384–394. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Barrett CL, DeBoever C, Jepsen K, Saenz
CC, Carson DA and Frazer KA: Systematic transcriptome analysis
reveals tumor-specific isoforms for ovarian cancer diagnosis and
therapy. Proc Natl Acad Sci USA. 112:pp. E3050–E3057. 2015;
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Duda K, Cholewa H, Łabuzek K,
Boratyn-Nowicka A and Okopień B: Novel strategies of ovarian cancer
treatment. Pol Merkur Lekarski. 39:337–342. 2015.(In Polish).
PubMed/NCBI
|
|
28
|
Monteith GR: Prostate cancer cells alter
the nature of their calcium influx to promote growth and acquire
apoptotic resistance. Cancer Cell. 26:1–2. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Moody SE, Schinzel AC, Singh S, Izzo F,
Strickland MR, Luo L, Thomas SR, Boehm JS, Kim SY, Wang ZC and Hahn
WC: PRKACA mediates resistance to HER2-targeted therapy in breast
cancer cells and restores anti-apoptotic signaling. Oncogene.
34:2061–2071. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Haslehurst AM, Koti M, Dharsee M, Nuin P,
Evans K, Geraci J, Childs T, Chen J, Li J, Weberpals J, et al: EMT
transcription factors snail and slug directly contribute to
cisplatin resistance in ovarian cancer. BMC Cancer. 12:912012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Thériault BL, Shepherd TG, Mujoomdar ML
and Nachtigal MW: BMP4 induces EMT and Rho GTPase activation in
human ovarian cancer cells. Carcinogenesis. 28:1153–1162. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Takai M, Terai Y, Kawaguchi H, Ashihara K,
Fujiwara S, Tanaka T, Tsunetoh S, Tanaka Y, Sasaki H, Kanemura M,
et al: The EMT (epithelial-mesenchymal-transition)-related protein
expression indicates the metastatic status and prognosis in
patients with ovarian cancer. J Ovarian Res. 7:762014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lili LN, Matyunina LV, Walker LD, Wells
SL, Benigno BB and McDonald JF: Molecular profiling supports the
role of epithelial-to-mesenchymal transition (EMT) in ovarian
cancer metastasis. J Ovarian Res. 6:492013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ford CE, Jary E, Ma SS, Nixdorf S,
Heinzelmann-Schwarz VA and Ward RL: The Wnt gatekeeper SFRP4
modulates EMT, cell migration and downstream Wnt signalling in
serous ovarian cancer cells. PLoS One. 8:e543622013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huang RY, Chung VY and Thiery JP:
Targeting pathways contributing to epithelial-mesenchymal
transition (EMT) in epithelial ovarian cancer. Curr Drug Targets.
13:1649–1653. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mao Y, Xu J, Li Z, Zhang N, Yin H and Liu
Z: The role of nuclear β-catenin accumulation in the Twist2-induced
ovarian cancer EMT. PLoS One. 8:e782002013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jiang H, Lin X, Liu Y, Gong W, Ma X, Yu Y,
Xie Y, Sun X, Feng Y, Janzen V and Chen T: Transformation of
epithelial ovarian cancer stemlike cells into mesenchymal lineage
via EMT results in cellular heterogeneity and supports tumor
engraftment. Mol Med. 18:1197–1208. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tang J, Wang J, Fan L, Li X, Liu N, Luo W,
Wang J and Wang Y and Wang Y: cRGD inhibits vasculogenic mimicry
formation by down-regulating uPA expression and reducing EMT in
ovarian cancer. Oncotarget. 7:24050–24062. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cha Y, Kim DK, Hyun J, Kim SJ and Park KS:
TCEA3 binds to TGF-beta receptor I and induces Smad-independent,
JNK-dependent apoptosis in ovarian cancer cells. Cell Signal.
25:1245–1251. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tanwar PS, Mohapatra G, Chiang S, Engler
DA, Zhang L, Kaneko-Tarui T, Ohguchi Y, Birrer MJ and Teixeira JM:
Loss of LKB1 and PTEN tumor suppressor genes in the ovarian surface
epithelium induces papillary serous ovarian cancer. Carcinogenesis.
35:546–553. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu T, Qin W, Hou L and Huang Y:
MicroRNA-17 promotes normal ovarian cancer cells to cancer stem
cells development via suppression of the LKB1-p53-p21/WAF1 pathway.
Tumour Biol. 36:1881–1893. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen JL, Chen F, Zhang TT and Liu NF:
Suppression of SIK1 by miR-141 in human ovarian cancer cell lines
and tissues. Int J Mol Med. 37:1601–1610. 2016. View Article : Google Scholar : PubMed/NCBI
|