Introduction
Gliomas, including glioblastoma multiforme (GBM),
have a poor prognosis and high rate of relapse (1). The most common type of glial neoplasm
is astrocytic tumor, approximately 80% of which are glioblastomas
with common genetic defects (2).
Gliomas can be treated by open surgery, chemotherapy, and/or
radiotherapy. However, no effective treatments are available,
resulting in a mean survival duration of <15 months (3).
Angiogenesis is a common feature of cancer and is
mediated by upregulation of angiogenic genes (4). Angiogenesis leads to the development
of large twisting vessels (5). The
disordered vasculature of GBM leads to a diminished blood supply,
which reduces the effectiveness of chemotherapy and radiotherapy
(6). Anti-angiogenic approaches
reportedly enhance the efficacy of chemotherapeutics (7). Bevacizumab (BEV), a recombinant
protein that inhibits VEGF-A, is a promising candidate
anti-angiogenesis drug (8). The
efficacy of BEV cotreatment for GBM patients with temozolomide
(TMZ) and radio/chemotherapy has been studied (9). A clinical study of avastin (AVAglio)
in GBM patients indicated that cotreatment with BEV and avastin
improved the progression-free survival duration (PFS, 4.4 months)
while radiotherapy with BEV cotreatment showed no significant
benefit for survival (10).
Therefore, there is an urgent need to develop drugs other than TMZ
and anti-VEGF agents for GBM patients.
TRIO and F-actin-binding protein (TrioBP) was
identified as a novel Trio-interacting protein by screening of a
skeletal muscle cDNA library (11). TrioBP contains an N-terminal
pleckstrin homology domain and a C-terminal coiled-coil region. The
interaction of TrioBP-1 with F-actin stabilizes the structure of
the latter (11). Subsequent
studies of the structure of TrioBP genes revealed that the TrioBP-4
and TrioBP-5 isoforms are required for hearing (12,13).
Linkage analysis of families has identified several mutations in
the 22q13 region designated DFNB28, which spans 34 genes, including
TrioBP-1 (14). TrioBP-1 is
ubiquitously expressed, whereas TrioBP-4 and TrioBP-5 are expressed
mainly in the eye and ear. Mutations in TrioBP-4 and TrioBP-5 have
been linked to a form of autosomal recessive nonsyndromic deafness.
All of the TrioBP mutations in DFNB28 that result in deafness are
located in exon 6 of TrioBP and affect only the TrioBP-4 and
TrioBP-5 isoforms (15). Because
the siRNA-induced decrease in Trio expression inhibits the
migration of glioblastoma cells (16), TrioBP, as a Trio-interacting
protein, might be involved in the development of gliomas.
Therefore, the role of TrioBP in glioblastoma was evaluated to
enable the development of novel chemotherapeutics. Taken together,
our results indicate that TrioBP has potential as a candidate
therapeutic agent for glioblastoma. An enhanced understanding of
the role of TrioBP in glioblastoma should provide important
information for the management of cancer.
Materials and methods
Antibodies and reagents
Anti-TrioBP antibodies were purchased from Novus
Biologicals, LLC (Littleton, CO, USA) and anti-actin antibodies
were from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
Horseradish peroxidase-conjugated anti-mouse IgG or anti-rabbit IgG
secondary antibodies were purchased from Komabiotech (Seoul,
Korea). The siRNA against human TrioBP were synthesized by
Intergrated DNA technologies. The siRNA sequences for TrioBP were
the following; 5′-TCCCAGCAGAACCATCCAACAAGAGAA-3′.
Cell culture and transfection
The glioblastoma cells (U87, U25, and U343-MG) were
maintained in medium (RPMI) supplemented with 10% FBS, 25 mM HEPES
(Thermo Fisher Scientific, Inc., Waltham, MA, USA), 1%
Antibiotics-Antimycotics (Thermo Fisher Scientific, Inc.). U87 and
U251-MG cells were transiently transfected with 30 nM control siRNA
or TrioBP siRNA by using Neon Transfection System (Thermo Fisher
Scientific, Inc.).
Patient samples
The study was approved by the Hospital Institutional
Review Board (approval number CNUH 2013-11-006) according to the
Declaration of Helsinki at Chungnam National University Hospital
(Daejeon, Korea), and written informed consent was obtained from
each patient before surgery. Normal brain tissue samples were
obtained from cadavers alternatively, from autopsy of surrounding
normal brain of glioblastoma patient who underwent surgery.
Immunoblot analysis
The immunoblot analysis was performed as the
described previously (17,18). Briefly, cells were placed on ice
and extracted with lysis buffer containing 50 mM Tris-HCl, pH 7.5,
1% v/v Nonidet P-40, 120 mM NaCl, 25 mM sodium fluoride, 40 mM
β-glycerol phosphate, 0.1 mM sodium orthovanadate, 1 mM
phenylmethylsulfonyl fluoride, 1 mM benzamidine, and 2 µM
microcystin-LR. Lysates were centrifuged for 15 min at 12,000 × g.
The cell extracts were resolved by 10–15% SDS-PAGE, and transferred
to Immobilon-P membranes (EMD Millipore, Billerica, MA, USA). The
filters were blocked for 1 h in 1X Tri-buffered saline buffer (TBS;
140 mM NaCl, 2.7 mM KCl, 250 mM Tris-HCl, pH 7.4), containing 5%
skimmed milk and 0.2% Tween-20, followed by an overnight incubation
with the anti-TrioBP and anti-actin antibodies diluted 1,000-fold
at 4°C. The secondary antibody was horseradish
peroxidase-conjugated anti-mouse IgG or anti-rabbit IgG (Koma
Biotech, Seoul, Korea), diluted 5,000-fold in the blocking buffer.
The detection of protein expression was visualized by enhanced
chemiluminescence, according to the manufacturer's instructions
(Thermo Fisher Scientific, Inc.).
Reverse transcription-quantitative
reverse transcription-polymerase chain reaction (RT-qPCR)
Total RNA was extracted from frozen tissue samples
or from cells using the PureHelix RNA Extraction Solution
(Nanohelix, South Korea). The cDNA was synthesized from total RNA
with the SuperScript III First-Strand Synthesis System for RT-qPCR
(Invitrogen; Thermo Fisher Scientific, Inc.). The RT-qPCR
measurement of individual cDNAs was performed using SYBR-Green dye
to measure duplex DNA formation with the StepOne Plus real-time PCR
system (Invitrogen; Thermo Fisher Scientific, Inc.) and normalized
to the expression of glyceraldehyde 3-phosphate dehydrogenase
(GAPDH) RNA. The following primers were used in the RT-qPCR
(forward, reverse); human TrioBP:
F-5′-TCCAAGGTCTCCCTTAGTACA/R-5′-GTGGGACTGGACTTGCTA; human GAPDH:
F-5′-TCGACAGTCAGCCGCATCTTCTTT/R-5′-TACGACCAAATCCGTTGACTCCGA.
RNA sequencing and RNA-Seq data
analysis
Total RNA of U87-MG, U251-MG and normal brain was
extracted using TRIzol reagent (Invitrogen) following the
manufacturer's procedures. The total RNA quantity and purity were
analysis of Bioanalyzer 2100 and RNA 6000 Nano LabChip Kit (Agilent
Technologies, Inc., Santa Clara, CA, USA). Roughly 10 µg of total
RNA was subjected to isolate Poly (A) mRNA with poly-T oligo
attached magnetic beads (Invitrogen; Thermo Fisher Scientific,
Inc.). Following purification, the mRNA is fragmented into small
pieces using divalent cations under raised temperature. Then the
cleaved RNA fragments were reverse-transcribed to create the final
cDNA library in accordance with the protocol for the mRNA-Seq
sample preparation kit (Illumina, Inc., San Diego, CA, USA). The
average insert size for the paired-end libraries was 300 bp (± 50
bp). Next we performed the paired-end sequencing on an Illumina
Hiseq 2000 system at Macrogen (Seoul, Korea) following the vendor's
recommended protocol. For each sample, sequenced reads were aligned
to the UCSC human reference genome (19) using the Tophat package (20), which initially removes a portion of
the reads based on quality information accompanying each read and
then maps the reads to the reference genome. FPKM (fragments per
kilobase of exon per million fragments mapped) were calculated to
compare the expression level of TrioBP mRNA variants in each
sample.
Confocal imaging analysis and indirect
immunofluorescence
U251-MG cells were grown on glass coverslips until
they were 50–70% confluent. After 24 h, the cells were fixed in 4%
paraformaldehyde at room temperature for 10 min and permeabilized
in 0.2% Triton X-100 for 5 min at room temperature. Then cells were
incubated in blocking buffer containing 5% bovine serum albumin
(Sigma-Aldrich; Merck KGaA) in 1X TBS for 1 h at 37°C. The rabbit
polyclonal anti-TrioBP was diluted 200-fold for primary antibody
and incubated for overnight. The secondary antibody,
FITC-conjugated anti-rabbit antibody (BD Biosciences, Franklin
Lakes, NJ, USA) was used. After appropriate rinsing, cover slips
were mounted with Vectashield (Vector Laboratories, Inc.,
Burlingame, CA, USA) and visualized using a Zeiss confocal
microscope (Zeiss AG, Oberkochen, Germany).
Immunohistochemistry
The analysis of immunohistochemistry was performed
as the described previously (21).
A human cancer tissue array slide with paraffin sections was
purchased from Bio Max (US Biomax Inc; Thermo Fisher Scientific,
Inc.). Histostain-Plus kits (Zymed Laboratories Inc., San
Francisco, CA, US) were used in accordance with the manufacturer's
instructions for the immunohistochemistry of tissue array. Briefly,
paraffin sections were deparaffinized with xylene and rehydrated in
a graded series of ethanol. The slide was submerged in peroxidase
quenching solution for 10 min. After it was washed twice with PBS
for 5 min, it was added with 2 drops of Reagent A for blocking and
incubated for 30 min. Following two washes with PBS, the primary
antibody was applied at 4°C for overnight. Then biotinylated
secondary antibody, Reagent B, was added after rinsing with PBS. It
was incubated at room temperature for 1 h. It was rinsed with PBS
and dropped with enzyme conjugated Reagent C. After it was washed
with PBS, DAB chromogen, and a mixture of Reagent D1, D2, and D3,
it was dropped, and signals were observed with a florescence
microscope. Then the reaction was stopped with distilled water, and
pictures were taken with a microscope.
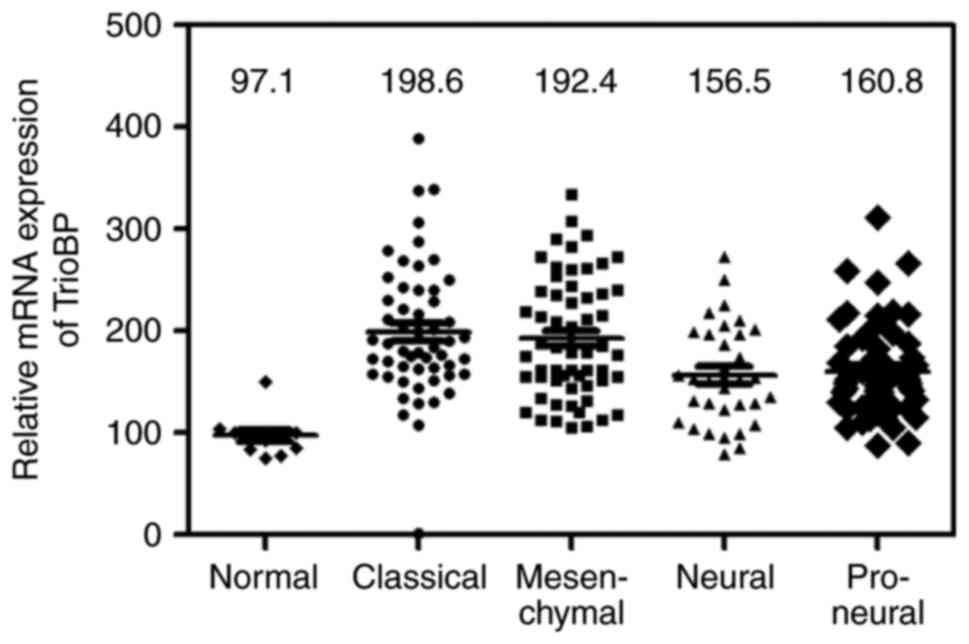
Bioinformatics data set
Glioma data sets and corresponding clinical data
were downloaded from the publicly available databases (213 cases
from the TCGA GBM dataset (http://www.betastasis.com/glioma/tcga_gbm/)). Normal;
n=11, Classical; n=54, Mesenchy-mal; n=58, Neural; n=33, Proneural;
n=57.
Real-time assay for cell proliferation
and migration
Real-time assay for cell proliferation and migration
were measured using an xCELLigence RTCA DP system (Roche
Diagnostics, Indianapolis, IN, USA), which monitors cellular events
in real-time without the incorporation of labels. Briefly, cells
were placed into well of an E-plate 16 (for proliferation; U87-MG
2×104, U251-MG 1.2×104 cells) or
Matrigel-coating (Matrigel: serum free media, 1:40) of the upper
chamber of CIM-plate 16 (for migration; U87-MG 2×104,
U251-MG 1.2×104 cells) and incubated for indicated
times.
Statistical analysis
Data are expressed as the mean ± SD from at least
three separate experiments performed triplicate. The differences
between groups were analyzed using a Student's t-test and P<0.05
was considered significant, and P<0.01 was highly significant
compared with corresponding control values. Statistical analyses
were carried out using SPSS software ver. 13.0 (SPSS, Inc.,
Chicago, IL, USA). Other statistical analysis performed using
analysis of variance (ANOVA) and Tukey's post hoc tests, using the
GraphPad Prism 5 software (GraphPad Software, Inc., La Jolla, CA,
USA). Differences were considered significant if P<0.05,
P<0.01, and P<0.001.
Results
TrioBP expression is high in
glioblastoma tissue
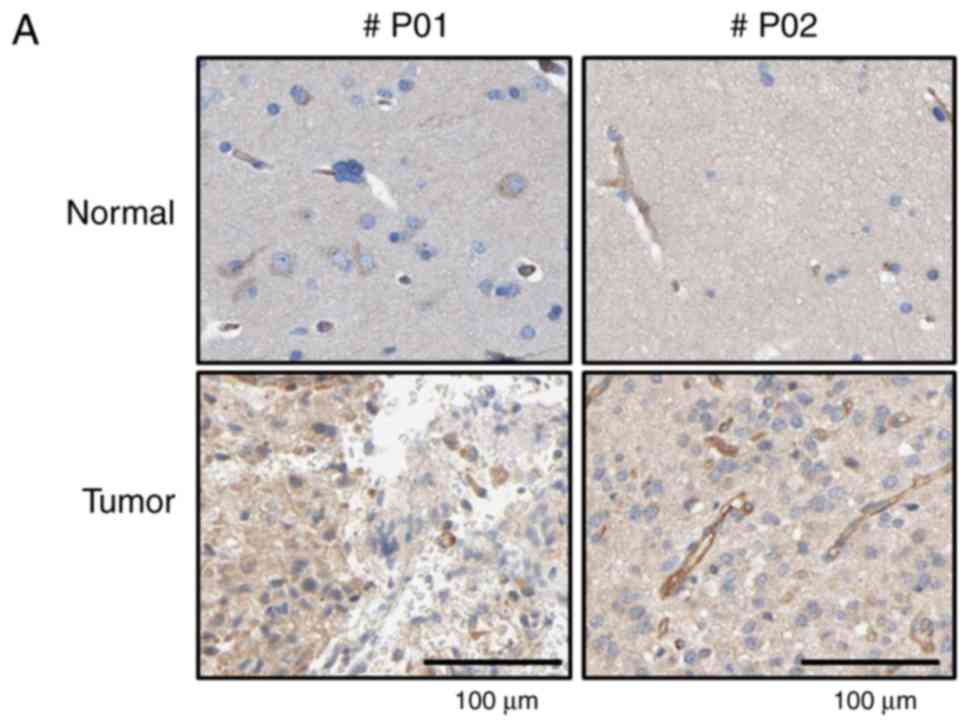
Immunohistochemical (IHC) analysis using a tissue
array and anti-TrioBP antibody showed stronger signals in brain
tumor tissues than in surrounding normal tissues (Fig. 1A). Total lysates of normal and
cancerous tissues from GBM patients were subjected to
immunoblotting using an anti-TrioBP antibody. TrioBP levels in GBM
tissues were higher than those in the surrounding normal tissues
(Fig. 1B), suggesting that TrioBP
expression is upregulated in glioblastoma patients.
TrioBP expression is elevated in
glioblastoma cell lines
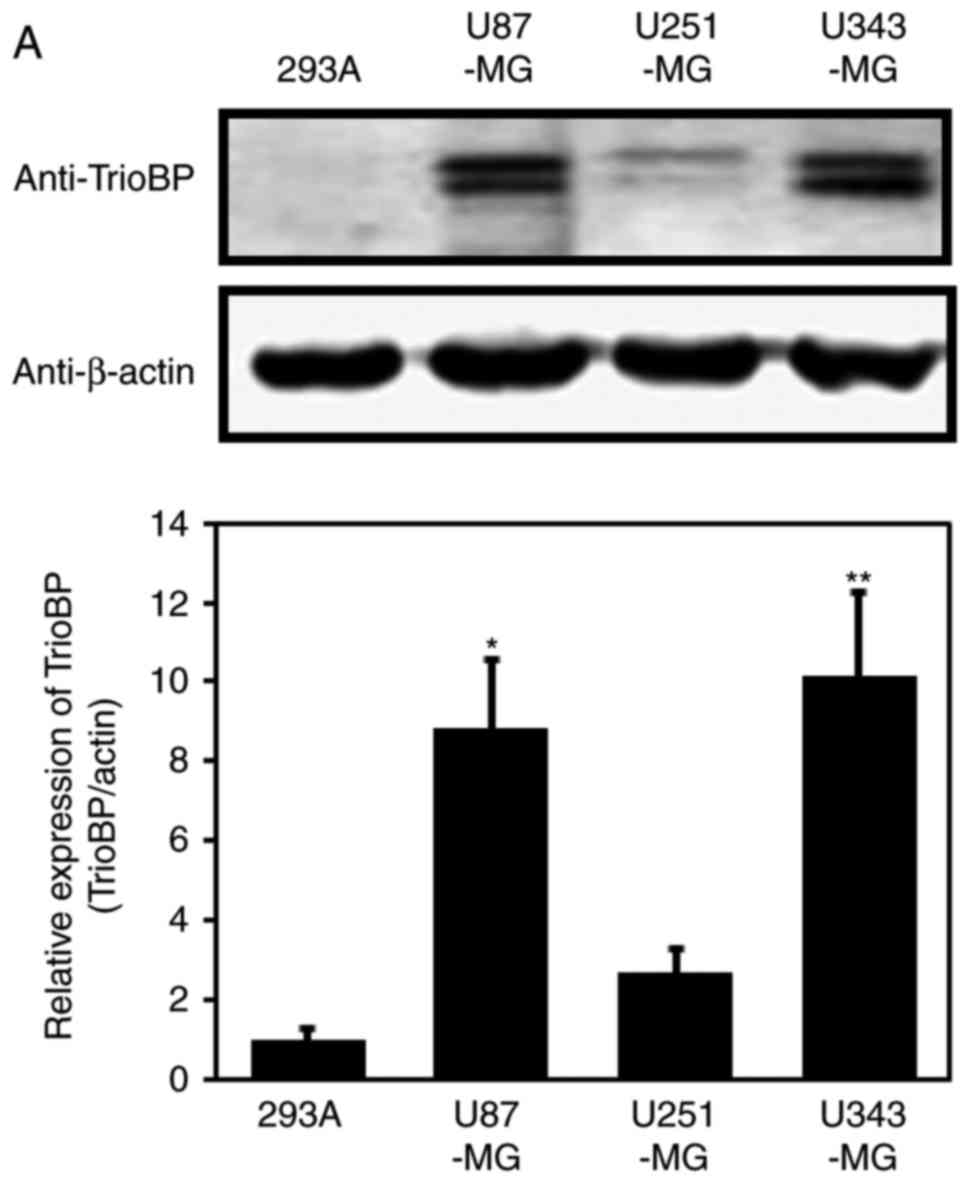
Based on the high TrioBP expression in samples from
GBM patients (Fig. 1), TrioBP
expression in glioblastoma cell lines was assessed by
immunoblotting. TrioBP expression was significantly increased in
U87-MG and U343-MG cells than in the other cell lines (Fig. 2A). Reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
showed that the mRNA level of TrioBP was increased in U343-MG, and
U87-MG cells compared with that in the controls (Fig. 2B).
Upregulation of TrioBP mRNA in U251-MG
and U87-MG cells
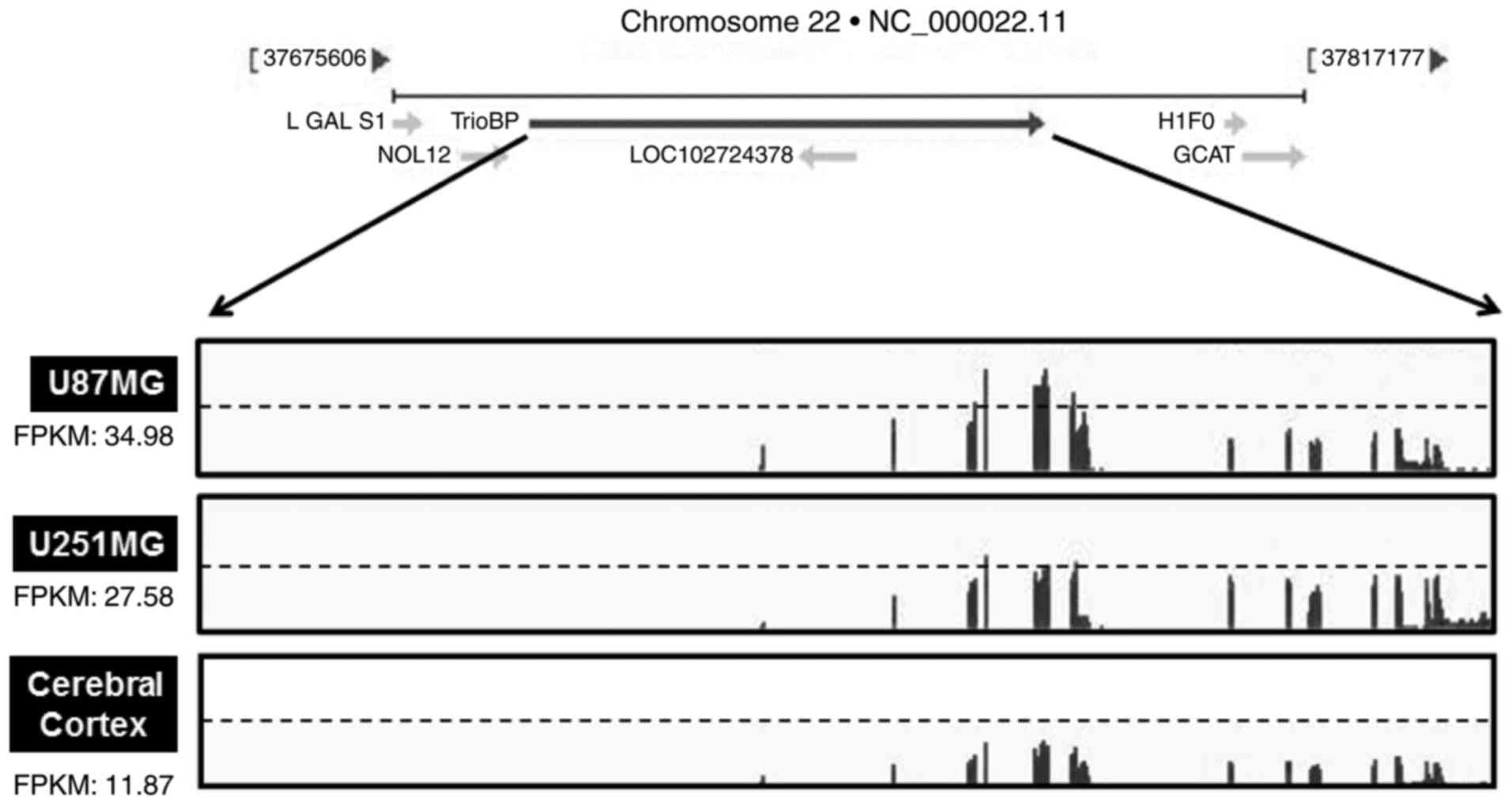
As TrioBP expression was elevated in glioblastoma
cells (Fig. 2A and B), the TrioBP
mRNA levels in glioblastoma cell lines were determined by
transcriptome profiling. Total RNA was prepared from U251-MG (low
TrioBP expression) and U87-MG (high TrioBP expression) glioblastoma
cells. Next, mRNA was split into small fragments to produce a cDNA
library. The number of fragments per kilobase of exon per million
fragments mapped (FPKM) was considered to be representative of the
TrioBP mRNA level. FPKM values were greater in U87-MG cells (34.98)
and U251-MG cells (27.58) than in cerebral cortex cells (11.87)
(Fig. 3), demonstrating that
TrioBP transcription is enhanced in glioblastoma cells.
TrioBP knockdown suppresses the
proliferation and migration of U251-MG and U87-MG cells
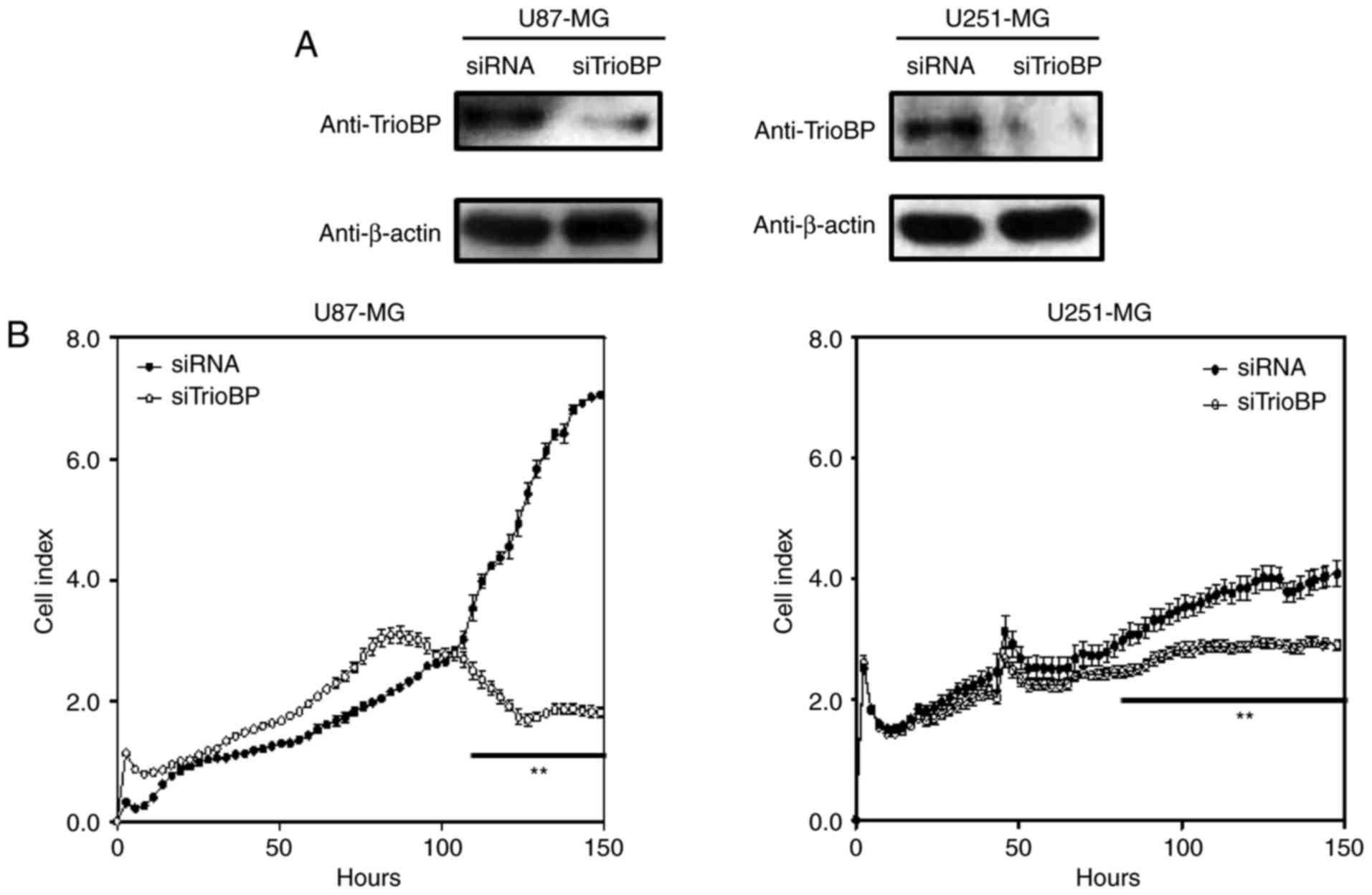
To investigate the roles of TrioBP in cancer cells,
U251-MG and U87-MG cells were transfected with TrioBP siRNA and
their proliferation and migration were evaluated by Excelligence.
TrioBP expression was significantly reduced (Fig. 4A). siRNA-mediated knockdown of
TrioBP inhibited the proliferation of U87-MG cells (left) and
U251-MG cells (right) (Fig. 4B).
The reduction in proliferation of U87-MG cells (left) was greater
than that of U251-MG cells (right). In addition, the migration of
U87-MG and U251-MG cells was suppressed (Fig. 4C). Therefore, TrioBP may play
important roles in cancer cells.
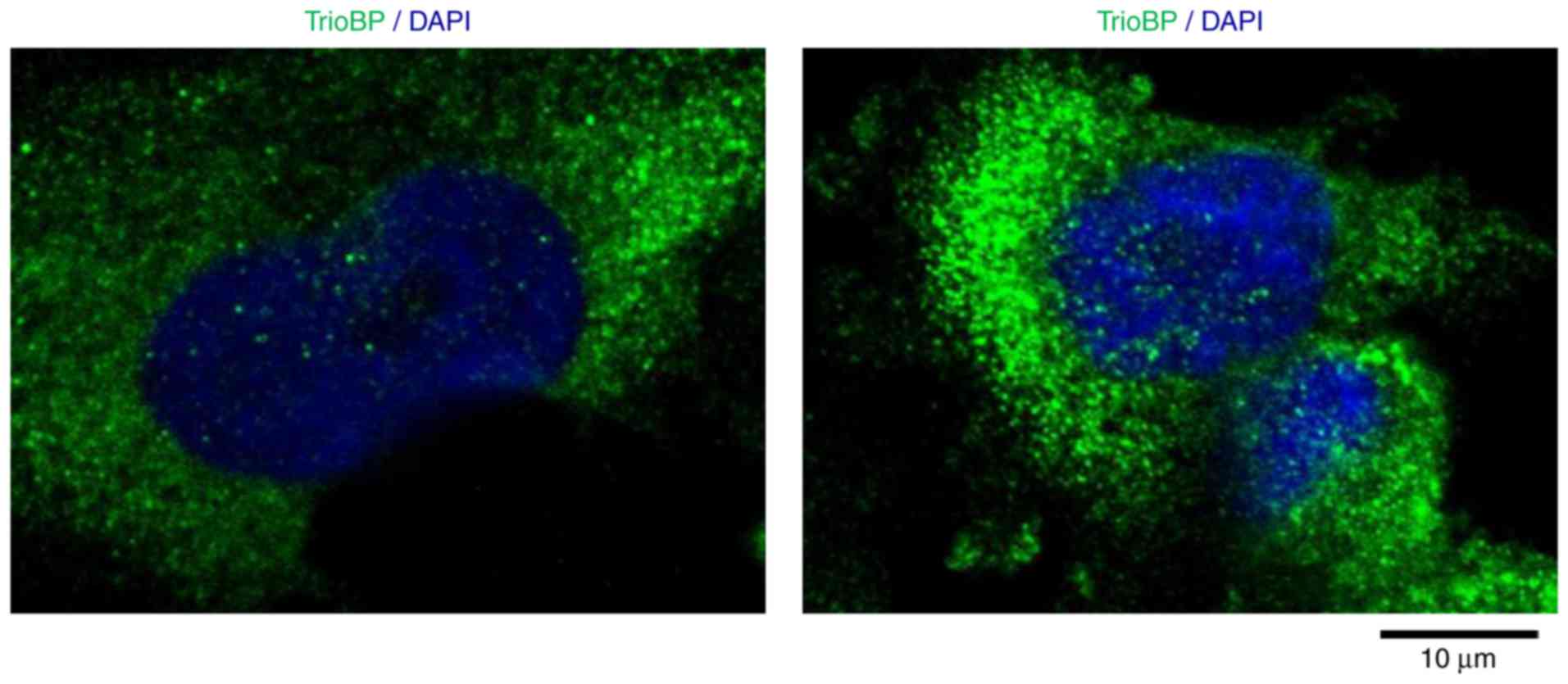
Cytoplasmic localization of TrioBP in
U251-MG cells
To investigate further its roles in GBM cells, the
intracellular localization of TrioBP in U251-MG cells was
evaluated. Interestingly, TrioBP was distributed in a punctate
pattern, possibly associated with the cytoskeleton and cytosol
(Fig. 5), indicating that
localization to the cytosol is important for its function in
glioblastoma cells.
Bioinformatic analysis of the TCGA GBM
cohort
A bioinformatic analysis was performed on the TCGA
GBM cohort. Similar to the above results, TrioBP expression was
increased in classical (n=54), mesenchymal (n=58), neural (n=33),
and proneural (n=57) glioblastomas compared with the level in
normal controls (n=11; one-way analysis of variance, P<0.0001;
Fig. 6).
Discussion
GBM is an aggressive type of brain tumor with a poor
prognosis despite the use of multiple therapies. Molecular
targeting may facilitate the development of effective treatment
strategies for GBM (1). Therefore,
novel biomarkers and candidate therapeutics for glioblastomas are
urgently needed.
In this study, TrioBP was found to have potential as
a marker of GBM, as its expression was elevated in a GBM cell line
and samples from GBM patients. Each of nine mutations in TrioBP
leads to hearing loss, from prelingual severe to profound
nonsyndromic deafness (12,13).
TrioBP interacts with Trio, which has rho guanine nucleotide
exchange factor (GEF) activity due to its kinase domain (11), resulting in inhibition of the
function of Trio in cell adhesion (22). RhoGEFs, downstream effectors of Rho
GTPases, stimulate the invasiveness and survival of glioma cells
(23). TrioBP-1 also interacts
with F-actin, leading to the stabilization of F-actin structures
(11). The characteristics of its
binding partners (Trio and F-actin) (23,24)
suggest a role for TrioBP in tumorigenesis. However, the function
of TrioBP in glioblastoma had not been investigated. Therefore, we
determined whether TrioBP participates in the progression of brain
tumors, including GBM. Unexpectedly, TrioBP expression was high in
glioblastoma tissue (Fig. 1A).
This was confirmed by immunoblotting of total cell lysates from
normal and cancerous tissues from GBM patients sampled during
surgery (Fig. 1B). TrioBP
expression was also high in U87-MG and U343-MG cells (Fig. 2A). RT-qPCR analysis revealed that
TrioBP mRNA levels are elevated in U343-MG, and U87-MG cells
(Fig. 2B) compared with that in
293A cells. Furthermore, the FPKM of TrioBP mRNA level was elevated
in U251-MG and U87-MG cells compared with that in normal brain
cells (Fig. 3). TrioBP-specific
siRNA in human mesenchymal stem cells suppresses cell proliferation
and migration by decreasing F-actin levels (25). Moreover, TrioBP isoforms 4 and 5
reportedly promote the motility of pancreatic cancer cells
(26). Indeed, the proliferation
and migration (which are characteristics of GBM cells) of
TrioBP-knockdown U87-MG and U251-MG cells were also inhibited
(Fig. 4).
To investigate further its roles in GBM, the
intracellular localization of TrioBP in U251-MG cells was
evaluated. Interestingly, TrioBP was distributed in a punctate
pattern, possibly associated with the cytoskeleton and cytosol
(Fig. 5), indicating that
localization to the cytosol is important for its function in
glioblastoma cells. A bioinformatic analysis of the TCGA GBM cohort
showed that TrioBP expression was high in all GBMs tested (Fig. 6), suggesting that TrioBP expression
is correlated with the clinical parameters of gliomas. In
conclusion, TrioBP is related to glioblastoma and may have
potential for predicting the prognosis of glioblastoma
patients.
Acknowledgements
This study was financially supported by research
fund of Chungnam National University in 2015 (Jongsun Park) and by
the National Research Foundation of Korea (NRF) grant funded by the
Korea Government (MEST; NRF-2012M3A9B6055302, NRF-2014R1A1A3050752,
NRF-2015R1A2A2A01003597 and NRF-2015R1D1A3A01015694).
References
|
1
|
Glaser T, Han I, Wu L and Zeng X: Targeted
nanotechnology in glioblastoma multiforme. Front Pharmacol.
8:1662017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Arif SH, Pandith AA, Bhat AR, Ramzan AU,
Malik Nk, Chibber SS, Wani AA, Tabasum R and Altaf Kirmani A: EGFR
and PTEN Gene Mutation Status in Glioblastoma Patients and their
Prognostic Impact on Patient's Survival. J Carcinog Mutagen.
6:2182015.
|
|
3
|
Virk SM, Gibson RM, Quinones-Mateu ME and
Barnholtz-Sloan JS: Identification of variants in primary and
recurrent glioblastoma using a cancer-specific gene panel and whole
exome sequencing. PLoS One. 10:e01241782015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bergers G and Benjamin LE: Tumorigenesis
and the angiogenic switch. Nat Rev Cancer. 3:401–410. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wesseling P, Ruiter DJ and Burger PC:
Angiogenesis in brain tumors; pathobiological and clinical aspects.
J Neurooncol. 32:253–265. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Das S and Marsden PA: Angiogenesis in
glioblastoma. N Engl J Med. 369:1561–1563. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Carmeliet P and Jain RK: Molecular
mechanisms and clinical applications of angiogenesis. Nature.
473:298–307. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Norden AD, Drappatz J and Wen PY: Novel
anti-angiogenic therapies for malignant gliomas. Lancet Neurol.
7:1152–1160. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chinot OL, Wick W, Mason W, Henriksson R,
Saran F, Nishikawa R, Carpentier AF, Hoang-Xuan K, Kavan P, Cernea
D, et al: Bevacizumab plus radiotherapy-temozolomide for newly
diagnosed glioblastoma. N Engl J Med. 370:709–722. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gilbert MR, Dignam JJ, Armstrong TS, Wefel
JS, Blumenthal DT, Vogelbaum MA, Colman H, Chakravarti A, Pugh S,
Won M, et al: A randomized trial of bevacizumab for newly diagnosed
glioblastoma. N Engl J Med. 370:699–708. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Seipel K, O'Brien SP, Iannotti E, Medley
QG and Streuli M: Tara, a novel F-actin binding protein, associates
with the Trio guanine nucleotide exchange factor and regulates
actin cytoskeletal organization. J Cell Sci. 114:389–399.
2001.PubMed/NCBI
|
|
12
|
Shahin H, Walsh T, Sobe T, Abu Sa'ed J,
Abu Rayan A, Lynch ED, Lee MK, Avraham KB, King MC and Kanaan M:
Mutations in a novel isoform of TRIOBP that encodes a
filamentous-actin binding protein are responsible for DFNB28
recessive nonsyndromic hearing loss. Am J Hum Genet. 78:144–152.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Riazuddin S, Khan SN, Ahmed ZM, Ghosh M,
Caution K, Nazli S, Kabra M, Zafar AU, Chen K, Naz S, et al:
Mutations in TRIOBP, which encodes a putative
cytoskeletal-organizing protein, are associated with nonsyndromic
recessive deafness. Am J Hum Genet. 78:137–143. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lenz DR and Avraham KB: Hereditary hearing
loss: From human mutation to mechanism. Hear Res. 281:3–10. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kitajiri S, Sakamoto T, Belyantseva IA,
Goodyear RJ, Stepanyan R, Fujiwara I, Bird JE, Riazuddin S,
Riazuddin S, Ahmed ZM, et al: Actin-bundling protein TRIOBP forms
resilient rootlets of hair cell stereocilia essential for hearing.
Cell. 141:786–798. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Salhia B, Tran NL, Chan A, Wolf A, Nakada
M, Rutka F, Ennis M, McDonough WS, Berens ME, Symons M and Rutka
JT: The guanine nucleotide exchange factors trio, Ect2 and Vav3
mediate the invasive behavior of glioblastoma. Am J Pathol.
173:1828–1838. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim HB and Yoo BS: Propolis inhibits
UVA-induced Apoptosis of human keratinocyte HaCaT cells by
scavenging ROS. Toxicol Res. 32:345–351. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li Y, Park J, Piao L, Kong G, Kim Y, Park
KA, Zhang T, Hong J, Hur GM, Seok JH, et al: PKB-mediated PHF20
phosphorylation on Ser291 is required for p53 function in DNA
damage. Cell Signal. 25:74–84. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bioinformatics UG: The International Human
Genome Sequencing Consortium. http://genome.ucsc.eduAccessed. 20:20162015.
|
|
20
|
Kim D and Salzberg S: TopHat 2.1.0.
https://ccb.jhu.edu/software/tophat/index.shtmlJuly
29–2015
|
|
21
|
Na CH, Hong JH, Kim WS, Shanta SR, Bang
JY, Park D, Kim HK and Kim KP: Identification of protein markers
specific for papillary renal cell carcinoma using imaging mass
spectrometry. Mol Cells. 38:624–629. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
van Rijssel J and van Buul JD: The many
faces of the guanine-nucleotide exchange factor trio. Cell Adh
Migr. 6:482–487. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fortin Ensign SP, Mathews IT, Symons MH,
Berens ME and Tran NL: Implications of Rho GTPase signaling in
glioma cell invasion and tumor progression. Front Oncol. 3:2412013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li A, Dawson JC, Forero-Vargas M, Spence
HJ, Yu X, König I, Anderson K and Machesky LM: The actin-bundling
protein fascin stabilizes actin in invadopodia and potentiates
protrusive invasion. Curr Biol. 20:339–345. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yun SP, Ryu JM, Jang MW and Han HJ:
Interaction of profilin-1 and F-actin via a β-arrestin-1/JNK
signaling pathway involved in prostaglandin E(2)-induced human
mesenchymal stem cells migration and proliferation. J Cell physiol.
226:559–571. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bao J, Wang S, Gunther LK, Kitajiri S, Li
C and Sakamoto T: The actin-bundling protein TRIOBP-4 and −5
promotes the motility of pancreatic cancer cells. Cancer Lett.
356:367–373. 2015. View Article : Google Scholar : PubMed/NCBI
|