Introduction
Hyperthyroidism is a pathological syndrome,
characterized by high levels of circulating thyroid hormone (TH).
Thyrotoxicosis is diagnosed by high serum concentrations of
thyroxine (T4) and triiodothyronine (T3), and low serum
concentrations of thyrotropin, additionally termed
thyroid-stimulating hormone. Symptoms of overt hyperthyroidism
include heat intolerance, palpitations, anxiety, fatigue, weight
loss and muscle weakness, which are associated with lipid
metabolism disorders (1).
Accumulating evidence has demonstrated the important roles of TH in
regulating lipid metabolism, with a number of genes involved in
lipid metabolism in the liver, an important TH target tissue, being
directly regulated by TH (2). The
effects of TH on lipid metabolism in the liver are associated with
a number of genes involved in lipogenesis and fatty acid oxidation
(3). However, the underlying
mechanisms associating TH with altered lipid metabolism remain to
be elucidated.
MicroRNAs (miRNAs/miRs) are a class of small,
noncoding, single-stranded RNA molecules containing ~22–25
nucleotides, that are able to bind to the 3′untranslated region
(3′UTR) of their target mRNA, leading to translational inhibition
or mRNA degradation (4). miRNAs
are important regulators of biological processes, including
metabolism, cell growth, apoptosis and carcinogenesis (5,6).
Global miRNA and mRNA expression profiling has revealed a potential
regulatory role for miRNAs in response to TH in the developing
mouse liver (7). miR-1, miR-206,
miR-133a and miR-133b expression was observed to be significantly
increased in the livers of hypothyroid mice, while their expression
was decreased in the livers of hyperthyroid mice (8). In addition, a number of miRNAs have
been reported to be involved in cholesterol efflux and hepatic
lipid metabolism (9). Therefore,
miRNAs may serve a role in lipometabolism regulated by TH in
hyperthyroidism.
The present study aimed to investigate the effects
of increased TH on serum miR-206 expression in patients with
hyperthyroidism. The role of the miRNA miR-206 in lipid metabolism
was assessed in cultured human hepatoblastoma HepG2 cells in
response to T3, with either overexpression or inhibition of
endogenous miR-206 expression. It was hypothesized that miR-206
expression may be altered by TH and has an important role in
T3-regulated lipid metabolism in liver cells.
Materials and methods
Patients
From October 2013 to March 2014 a total of 22
subjects recruited from October 2013 to March 2014 took part in the
present study. A total of 12 patients diagnosed with
hyperthyroidism were recruited from the Department of Endocrinology
and 10 healthy subjects were recruited from the Physical
Examination Center at The 82nd Hospital of Chinese People's
Liberation Army (CPLA). Patients who had diabetes mellitus or other
endocrine diseases, or with a body mass index >30
kg/m2, were excluded from taking part in the study.
Patient details are shown in Table
I. Patients taking any drugs known to influence TH metabolism
were additionally excluded. The ethics committee of The 82nd
Hospital of CPLA approved the present study. Written informed
consent was obtained from all subjects.
 | Table I.Clinical characteristics of study
subjects. |
Table I.
Clinical characteristics of study
subjects.
|
| Patients with
hyperthyroidism | Healthy subjects | P-value | P-value summary |
|---|
| No. subjects
(female) | 12
(8) | 10
(8) | – | – |
| Age, years | 49.0±7.5 | 42.7±12.1 |
0.1512 | ns |
| FT3, pg/ml |
9.9±0.5 |
3.5±0.1 | <0.0001 | – |
| FT4, ng/dl |
5.3±0.5 |
1.3±0.0 | <0.0001 | – |
| ALT, U/l | 23.8±1.9 | 18.6±2.1 |
0.0827 | ns |
| AST, U/l | 25.3±1.7 | 23.4±2.5 |
0.5143 | ns |
| TG, mmol/l | 1.22±0.31 | 1.71±0.50 |
0.3768 | ns |
| TC, mmol/l | 3.31±0.25 | 3.49±0.43 |
0.7080 | ns |
Clinical characteristics analysis
The clinical characteristics of the study population
include in serum free triiodothyronine (FT3), serum free thyroxine
(FT4), alanine aminotransferase (ALT), aspartate aminotransferase
(AST), triglyceride content (TG) and total cholesterol (TC). These
were measured by Olympus AU640 automatic biochemical analyzer
(Olympus Corporation, Tokyo, Japan).
Cell culture and treatment
Human hepatoblastoma (HepG2) cell line is known to
be misidentified, originally thought to be a hepatocellular
carcinoma cell line but shown to be from an hepatoblastoma
(10). HepG2 is a perpetual cell
line, which was derived from the liver tissue of a 15-year-old
Caucasian American male with a well-differentiated hepatocellular
carcinoma. HepG2 cells are a suitable in vitro model system
for the study of polarized human hepatocytes. HepG2 cells and their
derivatives are also used as a model system for studies of liver
metabolism and toxicity of xenobiotics. HepG2 cells were cultured
in Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin, and 100 µg/ml streptomycin. For treatment with T3
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), cells were seeded
in 12-well culture plates at a density of 3×105
cells/well, and maintained for 2 days incubated at 37°C in a
humidified chamber containing 5% CO2. Following
overnight incubation in serum-free DMEM, cells were treated with
100 nM T3 for 12, 24 or 36 h. For the investigation of triglyceride
synthesis, the medium was supplied with a 0.25 mM free fatty acid
(FFA) cocktail composed of lauric acid, myristic acid, linoleic
acid, oleic acid and arachidonic acids (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany).
Transient transfections
HepG2 cells, grown to 70–80% confluence, were
transiently transfected with miR-206 mimic (sense,
5′-UGGAAUGUAAGGAAGUGUGUGG-3′ and antisense,
5′-ACACACUUCCUUACAUUCCAUU-3′) or miR-206 inhibitor
(5′-CCACACACUUCCUUACAUUCCA-3′), or a corresponding mimic negative
control inhibitor (5′-UUCUCCGAACGUGUCACGUTT-3′; Shanghai GenePharma
Co., Ltd., Shanghai, China), using Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol. All the mimic and inhibitor containing FAM reporter
fluorophores were transfected at 10 nM. Transfection efficiency was
observed using an inverted fluorescent microscope (original
magnification, ×400; Leica Microsystems GmbH, Wetzlar, Germany),
and evaluated using the reverse transcription-quantitative
polymerase chain reaction (RT-qPCR). The mixtures were added to the
culture, and the culture medium was changed after 8 h, and cultured
in DMEM supplemented with 10% fetal bovine serum for 36 h further
for subsequent experiments.
RT-qPCR quantification of miR-206
expression
Cell/serum total RNA was extracted using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) or QIAzol
reagent (Applied Biosystems; Thermo Fisher Scientific, Inc.),
followed by isolation and purification using a miRNeasy Mini
kit/miRNeasy serum kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.). Subsequently, first-strand cDNA was synthesized
by RT using a TaqMan miRNA RT kit (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The expression of miR-206 was analyzed by
qPCR using TaqMan Universal Master Mix II in the ABI 7500 Sequence
Detection System (Applied Biosystems; Thermo Fisher Scientific,
Inc.). All primers were provided in the TaqMan® MicroRNA
Assays (cat. no. 4427975; Thermo Fisher Scientific, Inc.). PCR
conditions were as follows: 50°C for 2 min, 95°C for 2 min, and
then 40 cycles of 94°C for 15 sec and 60°C for 1 min. U6 small
nucleolar RNA was used as an internal standard for normalization.
miR-206 expression was calculated using the comparative
quantification cycle (Cq) method (2−ΔCq; ΔCq =
CqmiR-206 - CqU6), the Cq values were
converted into RQ via the 2-ΔΔCq method (11), incorporating the calculated
amplification efficiency for each primer pair.
Oil Red o staining
Oil Red O stock solution was prepared in isopropanol
(0.25 g/100 ml) and heated to 100°C for 10 min. HepG2 cells grown
to 70–80% confluence were washed with PBS and fixed using 4%
paraformaldehyde for 30 min at room temperature. Cells were soaked
in filtered 0.6% (w/v) Oil Red O solution (60% Oil Red O stock
solution, 40% water) for 20 min at room temperature. Cells were
washed with water to remove unbound dye and visualized using light
microscopy (original magnification, ×400).
Statistical analysis
Each sample was analyzed in triplicate. All data are
presented as the mean ± standard error of the mean. Student's
t-test was used to analyze the differences between groups. The
correlation between T4 parameters and miR-206 expression levels was
analyzed using the Pearson product-moment correlation coefficient
method. Comparisons among multiple groups were statistically
performed using one-way analysis of variance followed by Dunnett's
test. Two-tailed P<0.05 was considered to indicate a
statistically significant difference. The data were analyzed using
SPSS software version 17.0 (SPSS, Inc., Chicago, IL, USA).
Results
Serum miR-206 levels in patients with
hyperthyroidism
The clinical characteristics of the study population
are presented in Table I. Patients
with hyperthyroidism exhibited increased serum free T3 (FT3) and
free T4 (FT4) concentrations compared with the control
subjects.
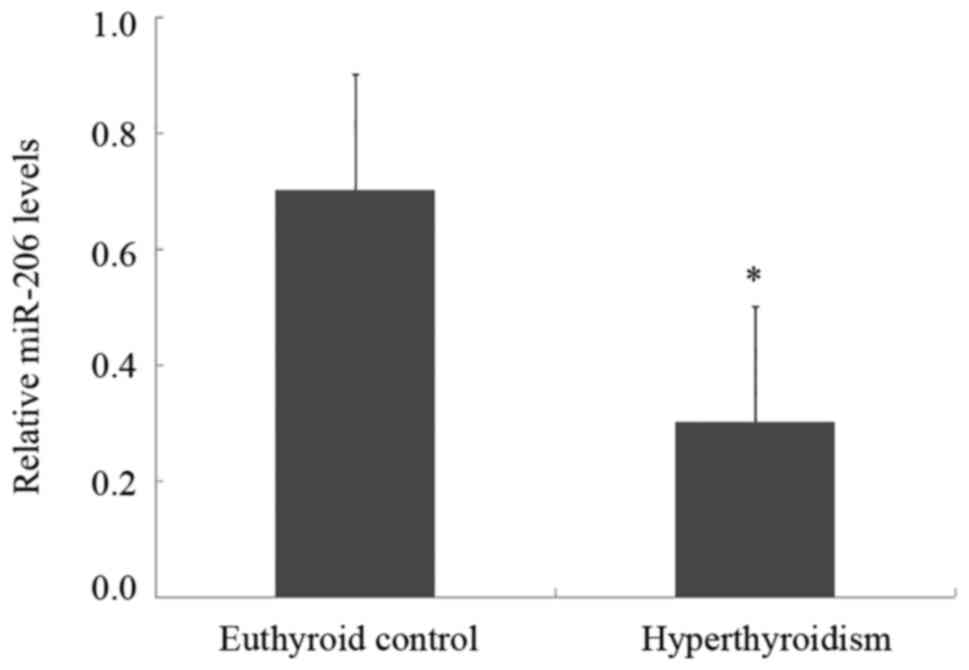
The RT-qPCR analysis demonstrated that the serum
miR-206 levels in patients with hyperthyroidism were significantly
decreased compared with euthyroid control subjects (Fig. 1). The present study further
analyzed the correlation between TH and miR-206 expression in
patients with hyperthyroidism. No apparent correlation was observed
between miR-206 expression and serum FT3 or FT4 levels (data not
shown).
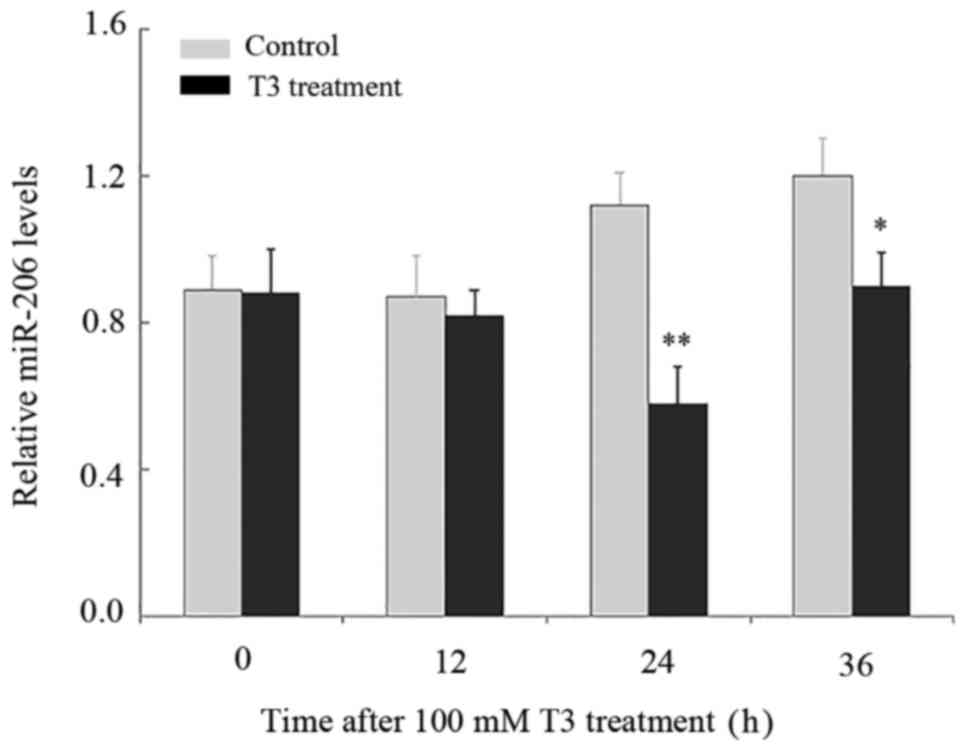
T3 downregulates miR-206 expression in
HepG2 cells
In order to clarify the effect of TH on miR-206
expression, HepG2 cells were treated with 100 nM T3 for 12, 24 or
36 h, with miR-206 expression examined using RT-qPCR analysis. The
expression of miR-206 was significantly decreased 24 h subsequent
to the treatment with T3, compared with HepG2 cells without T3
(Fig. 2). The expression of
miR-206 remained reduced at 36 h following the addition of T3
(Fig. 2).
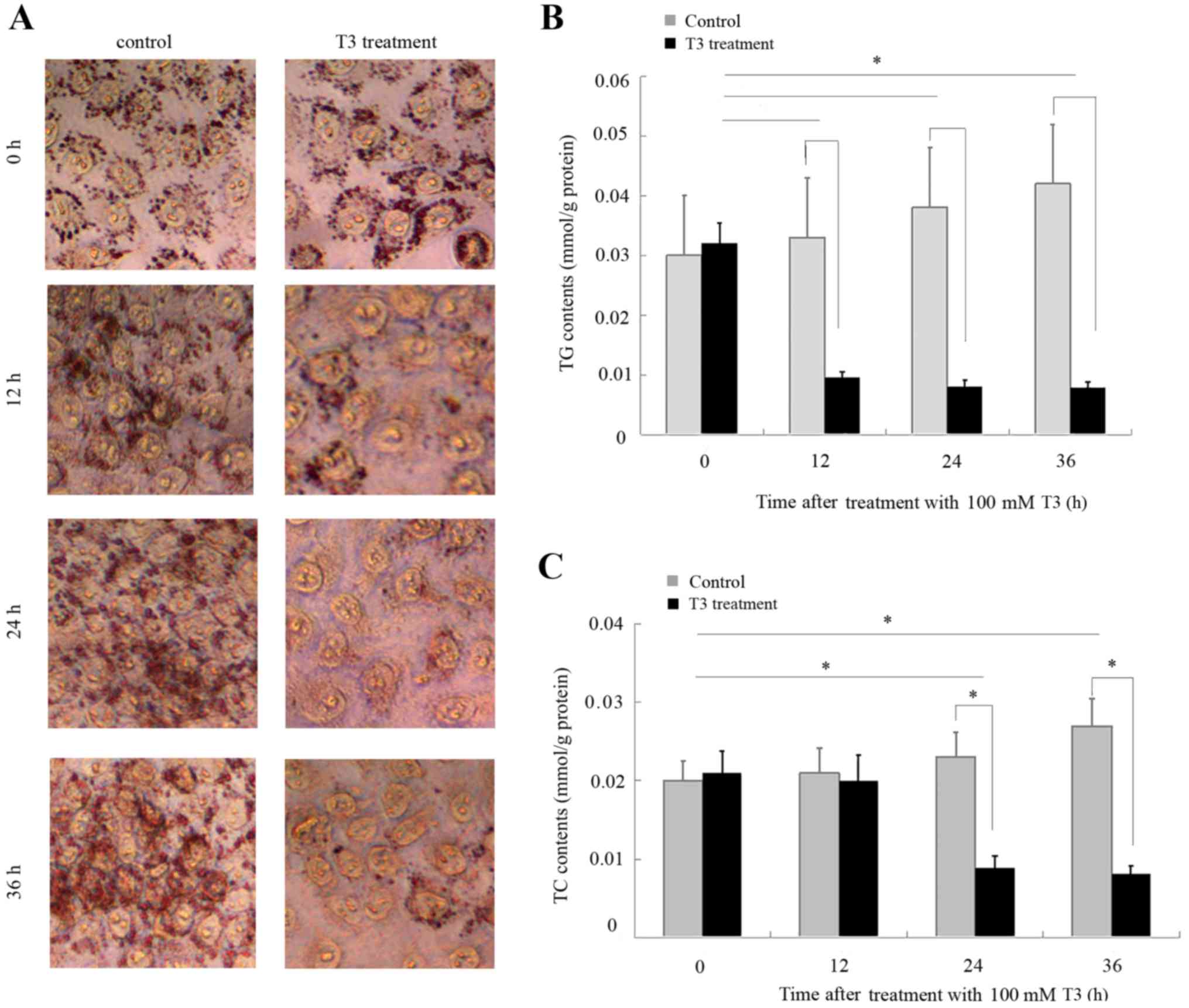
Effects of T3 on TG and TC synthesis
in HepG2 cells
To validate the effects of T3 on lipid synthesis,
HepG2 cells were cultured in DMEM supplemented with a 0.25 mM FFA
cocktail and treated with 100 nM T3 for 12, 24 or 36 h. Oil Red O
staining demonstrated that T3 suppressed lipid accumulation in
HepG2 cells (Fig. 3A). The TG was
significantly reduced 12 h subsequent to treatment with T3
(Fig. 3B), while the TC content
was significantly reduced 24 h subsequent to treatment with T3
(Fig. 3C). The TG and TC content
of the HepG2 cells remained reduced 36 h following treatment with
T3.
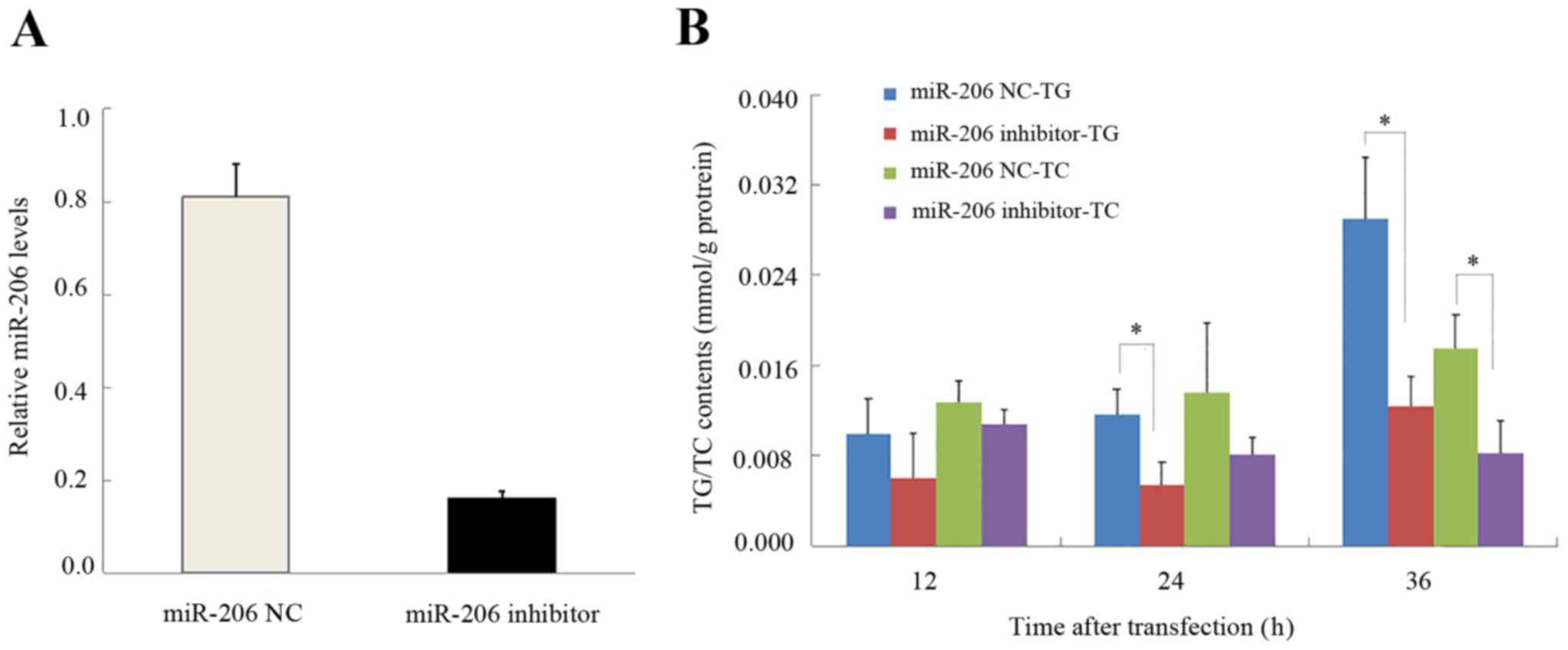
Effects of miR-206 on TG and TC
composition in HepG2 cells
The transfection of HepG2 cells with miR-206
inhibitor confirmed that the inhibitor effectively knocked down
miR-206 expression (Fig. 4A). The
addition of the miR-206 inhibitor reduced the TG at 24 h, and TG
and TC content at 36 h in HepG2 cells (Fig. 4B).
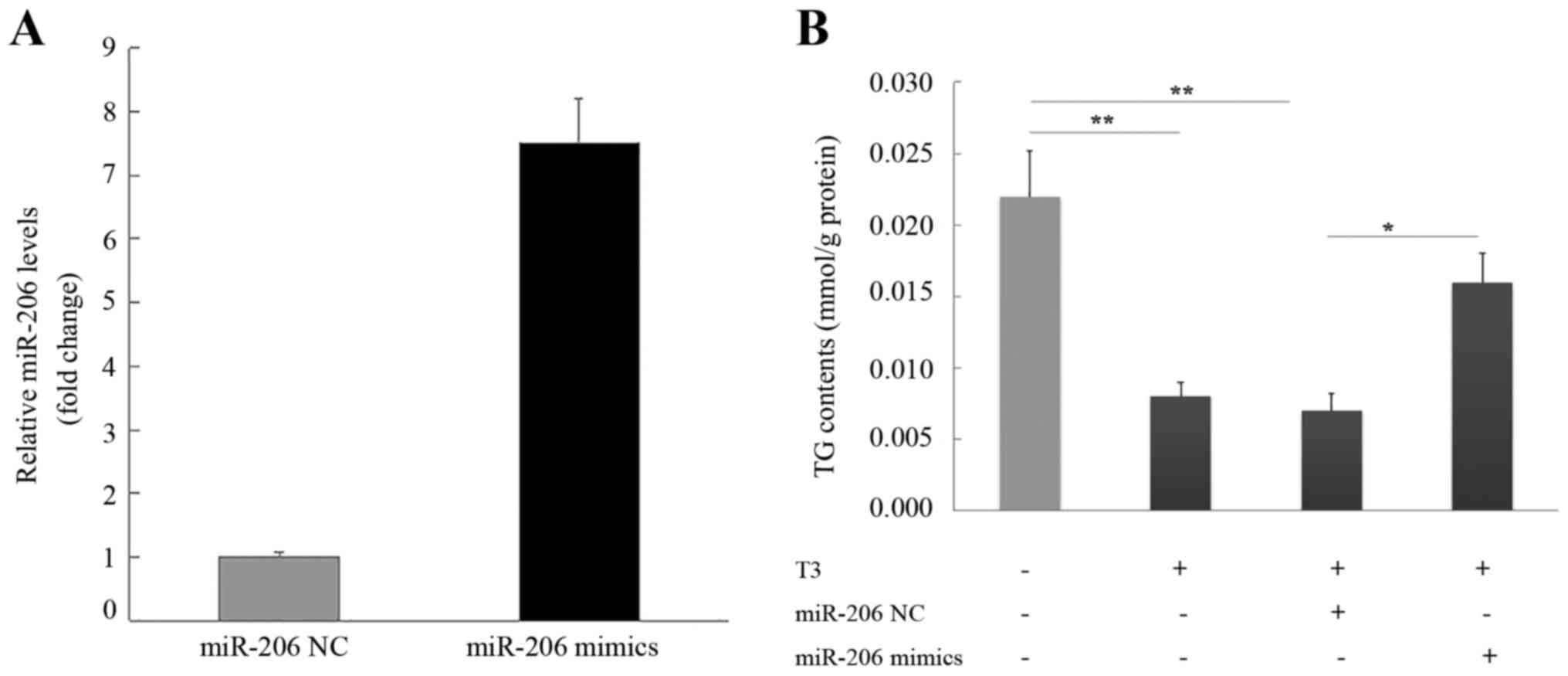
miR-206 partially rescues the TG
reduction induced by treatment with T3 in HepG2 cells
The transfection of HepG2 cells with the miR-206
mimic or mimic negative control confirmed that the miR-206 mimic
increased miR-206 expression (Fig.
5A). HepG2 cells treated with T3 exhibited a reduced TG content
that was restored with miR-206 overexpression (Fig. 5B). The addition of mimic negative
control did not alter TG in T3-treated cells.
Discussion
Appropriate TH levels are important for normal
growth, development and metabolism in adults (12). It is well established that TH
status is associated with body weight and energy expenditure
(13). Hyperthyroidism, excess TH,
promotes a hypermetabolic state characterized by increased resting
energy expenditure and weight loss. Serum TG and TC are typically
decreased in patients with hyperthyroidism. TH exerts direct and
indirect actions on the regulation of TG and TC production,
disposal and efflux (12,14,15),
primarily through specific nuclear receptor-mediated T3 actions
that modulate gene transcription. However, accumulating evidence
has indicated that non-genomic mechanisms may be involved in the TH
regulation of metabolism (16–18).
Reports have indicated that miRNAs have a potential role in
mediating the action of TH (7,19).
The results of the present study demonstrated that miR-206
expression was significantly decreased in patients with
hyperthyroidism compared with euthyroid subjects. Therefore,
understanding the association between TH and miR-206 may further
the understanding of the mechanisms underlying metabolic
dysfunction in patients with hyperthyroidism.
The liver is an important target of TH in the
regulation of energy metabolism and physiology (20,21).
Hyperthyroidism frequently results in liver dysfunction, and
hypothyroidism is associated with non-alcoholic fatty liver
disease. Previous studies have suggested that T3 has the ability to
reduce levels of lipids, including TC and TG (22,23).
The results of the present study demonstrated that prolonged
treatment with T3 led to reduced TG and TC content in HepG2 cells.
In addition, it was observed that that treatment with T3 resulted
in a downregulation of miR-206 expression. Increasing evidence has
suggested that miRNAs are involved in the post-transcriptional
regulation of genes in lipid metabolism (24,25).
Previous studies have indicated that the regulatory role of miR-206
in TC handling is tissue-specific (26,27).
It was demonstrated in the present study that the inhibition of
endogenous miR-206 expression in HepG2 cells resulted in decreased
intracellular TG and TC content. These results indicated that
miR-206 may be involved in the TH-mediated regulation of lipid
metabolism in HepG2 cells.
In order to confirm that miR-206 has a role in the
TH-mediated regulation of lipid metabolism, HepG2 cells were
transfected with an miR-206 mimic prior to being treated with T3.
Overexpression of miR-206 in HepG2 cells increased lipid
accumulation and attenuated the T3-induced downregulation of TG. A
previous study demonstrated that expression of miR-206 increased in
the liver of hypothyroid mice compared with controls (7). Given that TH deficiency is associated
with an increased incidence of hepatic steatosis, these results
suggested that miR-206 may serve an important role in the lipid
dysregulation in the liver associated with hyperthyroidism and
hypothyroidism.
The present study has limitations, in that the
miR-206 levels were only determined in the serum of hyperthyroid
patients and not in the liver. In addition, the use of HepG2 cells
provided an in vitro model of lipid metabolism in liver
cells and conclusions may not be directly applied to in vivo
liver physiology. However, the results of the present study
suggested that miRNAs may be a promising target for future
investigations into disorders of lipid metabolism in the liver in
cases of hyper- or hypothyroidism. Future studies, including the
use of mouse models of hyperthyroidism, are required for in
vivo investigation into the effects of miR-206 on lipid
metabolism in the liver in a physiological context. Further larger
studies may be used to study the effects of hyper- or
hypothyroidism on miR-206 expression in human subjects.
In conclusion, the results of the present study
demonstrated that patients with hyperthyroidism exhibited decreased
serum miR-206 expression levels and that miR-206 was involved in
the T3-mediated regulation of lipid metabolism in HepG2 cells. The
results of the present study provided novel insights into the
molecular mechanisms underlying TH regulation of lipid metabolism.
Whether miR-206 is affected in the liver in patients with
hyperthyroidism remains to be elucidated. Further experiments are
required to clarify whether miR-206 has the potential to be either
a biomarker or therapeutic target in hyperthyroidism.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from the
Medical and Scientific Innovations Project of Chinese People's
Liberation Army (grant no. 14SM030).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
YJZ, HW and YG collected data. YJZ, CZ and NZ
conducted the experiments. YJZ, NZ, WK, RL, and XX analysed the
data. CZ and YPZ wrote the paper. YPZ and XX designed the
study.
Ethics approval and consent to
participate
The Ethics Committee of the 82nd Hospital of CPLA
approved the present study. Written informed consent was obtained
from all subjects.
Consent for publication
Written informed consent was obtained from all
subjects.
Competing interests
All authors declared that they have no conflict of
interest with regard to this work.
References
|
1
|
Cooper D: Hyperthyroidism. Lancet.
362:459–468. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Boelaert K and Franklyn JA: Thyroid
hormone in health and disease. J Endocrinol. 187:1–15. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Oppenheimer JH, Schwartz HL, Lane JT and
Thompson MP: Functional relationship of thyroid hormone-induced
lipogenesis, lipolysis and thermogenesis in rat. J Clin Invest.
87:125–132. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kong YW, Cannell IG, de Moor CH, Hill K,
Garside PG, Hamilton TL, Meijer HA, Dobbyn HC, Stoneley M, Spriggs
KA, et al: The mechanism of micro-rna-mediated translation
repression is determined by the promoter of the target gene. Proc
Nail Acad Sci USA. 105:pp. 8866–8871. 2008; View Article : Google Scholar
|
|
5
|
Chen XM: MicroRNA signatures in liver
diseases. World J Gastroentero. 15:1665–1672. 2009. View Article : Google Scholar
|
|
6
|
Lynn FC: Meta-regulation: microRNA
regulation of glucose and lipid metabolism. Trends Endocrinol
Metab. 20:452–459. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dong H, Paquette M, Williams A, Zoeller
RT, Wade M and Yauk C: Thyroid hormone may regulate mRNA abundance
in liver by acting on micrornas. PLoS One. 5:e121362010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Visser WE, Heemstra KA, Swagemakers SM,
Ozgür Z, Corssmit EP, Burggraaf J, VanIjcken WF, vanderSpek PJ,
Smit JW and Visser TJ: Physiological thyroid hormone levels
regulate numerous skeletal muscle transcripts. J Clin Endocr Metab.
94:3487–3496. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Moore KJ, Rayner KJ, Suárez Y and
Fernández-Hernando C: The role of microRNAs in cholesterol efflux
and hepatic lipid metabolism. Annu Rev Nutr. 31:49–63. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
López-Terrada D, Cheung SW, Finegold MJ
and Knowles BB: Hep g2 is a hepatoblastoma-derived cell line. Hum
Pathol. 40:1512–1515. 2009. View Article : Google Scholar
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Brent GA: Mechanisms of thyroid hormone
action. J Clin Invest. 122:3035–3043. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Iwen KA, Schröder E and Brabant G: Thyroid
hormones and the metabolic syndrome. Eur Thyroid J. 2:83–92. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Webb P: Thyroid hormone receptor and lipid
regulation. Curr Opin Invest Drugs. 11:1135–1142. 2010.
|
|
15
|
Shoemaker TJ, Kono T, Mariash CN and
Evansmolina C: Thyroid hormone analogues for the treatment of
metabolic disorders: New potential for unmet clinical needs? Endocr
Pract. 18:954–964. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cheng SY, Leonard JL and Davis PJ:
Molecular aspects of thyroid hormone actions. Endocr Rev.
31:139–170. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cordeiro A, deSouza LL, Oliveira LS,
Faustino LC, Santiago LA, Bloise FF, Ortiga-Carvalho TM, Almeida NA
and Pazos-Moura CC: Thyroid hormone regulation of Sirtuin 1
expression and implications to integrated responses in fasted mice.
J Endocrinol. 216:181–193. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sinha RA, You SH, Zhou J, Siddique MM, Bay
BH, Zhu X, Privalsky ML, Cheng SY, Stevens RD, Summers SA, et al:
Thyroid hormone stimulates hepatic lipid catabolism via activation
of autophagy. J Clin Invest. 122:2428–2438. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yap CS, Sinha RA, Ota S, Katsuki M and Yen
PM: Thyroid hormone negatively regulates CDX2 and SOAT2 mRNA
expression via induction of miRNA-181d in hepatic cells. Biochen
Bioph Res Commun. 440:635–639. 2013. View Article : Google Scholar
|
|
20
|
Ståhlberg N, Merino R, Hernández LH,
Fernández-Pérez L, Sandelin A, Engström P, Tollet-Egnell P, Lenhard
B and Flores-Morales A: Exploring hepatic hormone actions using a
compilation of gene expression profiles. BMC Physiol. 5:82005.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dong H, Yauk CL, Williams A, Lee A,
Douglas GR and Wade MG: Hepatic gene expression changes in
hypothyroid juvenile mice: Characterization of a novel negative
thyroid-responsive element. Endocrinology. 148:3932–3940. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shibata A, Kawakami Y, Kimura T, Miyazawa
T and Nakagawa K: α-tocopherol attenuates the triglyceride- and
cholesterol-lowering effects of rice bran tocotrienol in rats fed a
western diet. J Agr Food Chem. 64:5361–5366. 2016. View Article : Google Scholar
|
|
23
|
Goldberg IJ, Huang LS, Huggins LA, Yu S,
Nagareddy PR, Scanlan TS and Ehrenkranz JR: Thyroid hormone reduces
cholesterol via a non-LDL receptor-mediated pathway. Endocrinology.
153:5143–5149. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Vickers KC, Sethupathy P, Baran-Gale J and
Remaley AT: Complexity of microRNA function and the role of isomiRs
in lipid homeostasis. J Lipid Res. 54:1182–1191. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vickers KC, Shoucri BM, Levin MG, Wu H,
Pearson DS, Osei-Hwedieh D, Collins FS, Remaley AT and Sethupathy
P: MicroRNA-27b is a regulatory hub in lipid metabolism and is
altered in dyslipidemia. Hepatology. 57:533–542. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhong D, Huang G, Zhang Y, Zeng Y, Xu Z,
Zhao Y, He X and He F: MicroRNA-1 and microRNA-206 suppress
LXRα-induced lipogenesis in hepatocytes. Cell Signal. 25:1429–1437.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Vinod M, Chennamsetty I, Colin S, Belloy
L, De Paoli F, Schaider H, Graier WF, Frank S, Kratky D, Staels B,
et al: MIR-206 controls LXRα expression and promotes LXR-mediated
cholesterol efflux in macrophages. Biochim Biophys Acta.
1841:827–835. 2014. View Article : Google Scholar : PubMed/NCBI
|