Introduction
Vascular smooth muscle cells (VSMCs) are cellular
components of the medial layer of the artery wall adjacent to
vascular endothelial cells (VECs). VSMCs serve a number of roles
under normal physiological conditions (1,2).
Proliferation and migration of VSMCs occurs during the early stage
of atherosclerotic lesion formation and post angioplasty restenosis
(3). Numerous cell types are
involved in atherogenesis and a complex network of transcription
factors and proteins is involved in this process (4).
VECs can regulate the contractile phenotype of VSMCs
through modulation of miR-206/ADP-ribosylation factor 6 and
sodium/calcium exchanger 1/exosome (2). When stimulated by endothelial injury
signals, VSMCs migrate into intima and undergo a proliferative
phenotypic switch (5). Macrophages
contribute to pathogenesis of restenosis via secretion of growth
factors, cytokines and chemokines, which induce proliferation and
activation of VSMCs (4).
Endothelial dysfunction is an early contributor and predictor of
atherosclerosis (AS) (6,7). Endothelial dysfunction results in the
earliest detectable alterations during the development of
atherosclerotic lesions (6,7). The
VECs-VSMCs co-culture model has been widely adopted for the
investigation of AS (2,8,9).
Data presented by a previous study supports the
hypothesis that inflammation contributes to the initiation,
progression and plaque rupture of AS (10). AS is dependent on innate immune
responses and involves activation of Toll-like receptors (TLRs).
TLRs are pattern-recognition receptors and a part of the
interleukin-1 receptor/Toll-like receptor superfamily. Activation
of TLRs leads to expression of inflammatory proteins that may lead
to acute coronary syndrome (11–13).
Different subtypes of TLRs are involved in a number of aspects of
the inflammatory response.
TLR4 exhibits a high level of expression in aortic
tissues from patients who have undergone coronary artery bypass
graft surgery and is positively correlated with Gensini score
(14). Kapelouzou et al
(15) demonstrated that the TLR4
mRNA expression level was markedly upregulated in a hyperlipidemic
rabbit model and was associated with the progression of AS. Studies
in knockout mice demonstrated that the TLR4/nuclear factor-κB
(NF-κB) signaling pathway contributes to chronic unpredictable mild
stress-induced AS via activation of pro-inflammatory cytokines
(16). The TLR4/NF-κB signaling
pathway also contributes to the early stage intimal foam cell
accumulation at lesion-prone aortic sites (12). In vitro, TLR4 mediates
hyperglycemia-induced inflammation and dysregulation of the
endothelial glycocalyx levels in human macrovascular aortic ECs
(17). TLR4 inhibitor CLI-095 can
suppress the progression of AS by reducing macrophage foam cell
formation (18). Yang et al
(19) hypothesized that TLR4
signaling promoted a pro-inflammatory phenotype in VSMCs. Positive
feedback regulation of VSMCs proliferation is mediated by the
TLR4/Rac family small GTPase 1/Akt serine/threonine kinase
signaling pathway (20).
Furthermore, the TLR4/NF-κB signaling pathway can increase the
expression of matrix metalloproteinase-9 mRNA and protein levels in
order to stimulate migration of human aortic VSMCs (21).
7-Difluoromethoxy-5,4′-dimethoxy-genistein (DFMG) is
a novel active chemical molecule. Our previous studies indicated
that DFMG was more efficient compared with genistein (GEN) in
reducing the risk of cardiovascular disease by inhibition of
oxidative damage and/or inhibition of the TLR4 signaling pathway
(22,23). The authors of the present study,
also previously reported that DFMG could protect VECs through
inhibition of the mitochondrial apoptotic pathway (24,25).
Furthermore, the preventive effect of DFMG was more apparent
compared with the therapeutic effect within apolipoprotein
E−/−mice induced by a cholesterol-rich diet (26).
The present study investigated whether TLR4 has a
role in abnormal proliferation and migration of VSMCs induced by
lysophosphatidylcholine (LPC)-injured VECs. The mechanism
underlying DFMG-mediated protection of VSMCs from VECs-stimulated
dysfunction in vitro has also been investigated. The present
study aimed to determine the activity of DFMG on the proliferation
and migration of VSMCs using a non-contact co-culture model with
LPC-injured VECs.
Materials and methods
Reagents and plasmids
LPC was purchased from Sigma-Aldrich (Merck KGaA,
Darmstadt, Germany) and dissolved in PBS. DFMG (purity, >99%)
was synthesized as previously described (27) and dissolved in dimethyl sulfoxide
(Amresco, LLC, Solon, OH, USA). DFMG was subsequently
filter-sterilized. Human interleukin-6 (IL-6) and tumor necrosis
factor-α (TNF-α) ELISA kits were purchased from Shanghai ExCell
Biology, Inc. (EH004-96 and EH009-96; Shanghai, China). The Cell
Counting kit-8 (CCK-8) was purchased from Vazyme (Piscataway, NJ,
USA). MegaTran1.0 transfection reagent, p-green fluorescent protein
(GFP)-V-RS-TLR4-short hairpin (sh)RNA and pCMV6-AC-GFP-TLR4-cDNA
plasmids were obtained from OriGene Technologies, Inc. (Rockville,
MD, USA). Anti-TLR4 and anti-β-actin antibodies were obtained from
Abgent, Inc. (ASC10194 and AM1021B, San Diego, CA, USA). All
secondary antibodies were supplied by ComWin Biotech Co. Ltd.
(CW0156S and CW0102S; Beijing, China).
Cell culture
Human umbilical vein EC and human aortic SMC cell
lines were obtained from the China Center for Type Culture
Collection (Wuhan, China). Both cell lines were incubated in a
humidified incubator at 37°C with 5% CO2 and cultured in
Dulbecco's modified Eagle's medium (DMEM), supplemented with 10%
fetal bovine serum (FBS), 100 IU/ml penicillin G and 100 µg/ml
streptomycin. Cell culture reagents were obtained from Biological
Industries Israel Beit-Haemek Ltd. (Beit HaEmek, Israel).
Determination of IL-6 and TNF-α
levels
VECs were pretreated with LPC (10, 20, 30, 40 and 50
µM) at 37°C for 24 h; the levels of IL-6 and TNF-α in supernatants
were quantified using human IL-6 and TNF-α ELISA kits
(aforementioned) according to the manufacturer's protocol. A total
of 100 µl serially-diluted standard samples and/or supernatant
samples were added into the microplate and incubated at 37°C for 1
h. The plate was further incubated with 100 µl antibody solution
against IL-6 or TNF-α at 37°C for an additional 1 h. Subsequently,
100 µl horseradish peroxidase-conjugated secondary antibodies were
added and the plate was incubated at 37°C for 30 min. The samples
were washed four times with 100 µl PBS containing Tween-20 (PBST)
prior to incubation with secondary antibodies. Finally, the plates
were incubated with 100 µl substrate in the dark at 37°C for 15
min. The absorbance was measured at a wavelength of 450 nm with a
spectrophotometer (Bio-Rad Laboratories, Inc., Hercules, CA,
USA).
Establishment of the co-culture
model
Co-culture of VECs and VSMCs was performed using a
Transwell chamber with a 10 µm-thick porous membrane. The membrane
contained 0.4 µm pores in a Transwell cell culture insert (Corning
Incorporated, Corning, NY, USA). This prevented cells from direct
contact. Briefly, VSMCs (1×105 cells/well) were seeded
in 6-well plates and cultured for 24 h prior to co-culture with
pretreated and/or transfected VECs. VECs (1×105
cells/well) were individually cultured on the insert filters for 24
h, then pretreated with DFMG (0.3, 1.0 and 3.0 µM) for 12 or 24 h
prior to treatment with LPC for 24 h, or pretreated following
transfection for 48 h. Subsequently, VECs (1×105
cells/well) with inserts were added to the upper chamber of the
Transwell system (VSMCs at the bottom). Both cells were maintained
in DMEM with 10% FBS at 37°C for 24 h. A light microscope was used
to observe migratory cells in lower chamber; magnification,
×200.
VSMCs viability assay
The CCK-8 assay was used in order to detect cell
viability of VSMCs. VSMCs were co-cultured with VECs treated with
different interventions for 24 h. VSMCs were digested and cultured
in 100 µl medium in 96-well plates (1×104 cells/well)
with four replicates for 24 h. Subsequently, the medium was
replaced and 10 µl CCK-8 solution was added to each well for an
additional 2 h incubation at 37°C. The absorbance was measured at a
wavelength of 450 nm. Cell viability was calculated based on the
optical density values of each group.
VSMCs wound healing (migration)
assay
The wound healing assay was carried out as
previously described (28). Prior
to co-culture, VSMCs were seeded on 6-well plates and scratched
using a 200 µl pipette tip in order to generate a cell-free area.
PBS was used to rinse cell debris three times and cells were
incubated in FBS-free media in order to prevent proliferation.
Phase contrast images of cells were captured at 0 h. Furthermore,
pretreated and/or transfected VECs with inserts were added to the
upper chamber of the Transwell system. Following 24 h incubation at
37°C, images of the same plates and cells were captured. Cell-free
areas were quantified using Adobe Photoshop CS6 software (Adobe
Systems, Inc., San Jose, CA, USA). The migration was expressed as
percentage coverage.
VECs transfection
Transfection of VECs with TLR4 shRNA and cDNA was
performed out MegaTran 1.0, a non-lipid polymer-based transfection
reagent with relatively low toxicity on the transfected cells. VECs
were seeded on 6-well plates and cultured until 50–70% confluence
prior to transfection. 3 µg TLR4 shRNA, cDNA and corresponding
negative control (only with GFP gene) plasmid (3 µg) was diluted in
300 µl DMEM and vortexed gently. A total of 9 µl MegaTran 1.0 was
added to the diluted DNA and vortexed immediately for 10 sec.
Samples were incubated for 10 min at room temperature and the
MegaTran 1.0/DNA mixture was added to each well (already containing
2 ml DMEM). The plate was shaken gently in order to achieve an even
distribution of the complexes and incubated at 37°C for 4 h. The
expression of TLR4 was detected at 24 to 48 h following
transfection.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the cells using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA), according to the manufacturer's protocol. To quantify the
mRNA, 500 ng RNA from each sample were reverse transcribed into
complementary DNA (cDNA) at 50°C for 15 min, then 85°C for 5 sec
using HiScript Q RT SuperMix for qPCR (Vazyme Biotech Co., Ltd.,
Nanjing, China). qPCR was carried out using SYBR Premix
ExTaq™ (Vazyme) and a 7500 fast qPCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). Cycling conditions
consisted of an initial incubation at 95°C for 5 min, then 40
cycles of 10 sec at 95°C and 30 sec at 60°C, final extension for 15
sec at 95°C and 1 min at 60°C. The samples were run in triplicate.
The 2−ΔΔCq method was used to analyze relative gene
expression (29). GAPDH was used
as an endogenous housekeeping gene control. The qPCR primer
sequences are listed in Table
I.
 | Table I.Primer sequences used for qPCR. |
Table I.
Primer sequences used for qPCR.
| Gene | Primer sequence
(5′-3′) | Product size
(bp) |
|---|
| GAPDH | F:
CAGGAGGCATTGCTGATGAT | 138 |
|
| R:
GAAGGCTGGGGCTCATTT |
|
| TLR4 | F:
CCGAGGCCATTATGCTATGT | 141 |
|
| R:
TCCCTTCCTCCTTTTCCCTA |
|
Western blotting
Cells were digested and subsequently lysed on ice
using radioimmunoprecipitation assay lysis buffer containing 1%
phenylmethyl sulfonyl fluoride. Samples were centrifuged at 12,000
× g at 4°C for 15 min. Protein concentration was determined with a
Bicinchoninic Acid protein assay kit (Beijing Solario Science &
Technology Co., Ltd., Beijing, China). A total of 25 µg of each
protein lysate was separated by 10% SDA-PAGE and transferred onto
polyvinylidene difluoride membranes (EMD Millipore, Billerica, MA,
USA) and blocked with 5% skimmed milk in PBST (PBS containing 0.05%
Tween-20) at 37°C for 1 h. The membranes were incubated with rabbit
anti-TLR4 antibody (1:1,000 dilution) and mouse anti-β-actin
antibody (1:10,000 dilution) on a shaking platform at 4°C
overnight. Following washing of the membranes with PBST, goat
anti-rabbit antibody (1:10,000 dilution) and goat anti-mouse
antibody (1:10,000 dilution) were incubated with the membranes for
1 h at room temperature. The membranes were washed with PBST three
times and protein expression was visualized by Pierce ECL Western
Blotting Substrate (Pierce; Thermo Fisher Scientific, Inc.). The
protein expression was quantified using densitometry and analyzed
using GelPro software (version 3.2; Media Cybenetics, Inc.,
Rockville, MD, USA). β-actin expression was used as an internal
control.
Statistical analysis
Data are presented as the mean ± standard deviation
from three independent experiments. SPSS software (version 17.0;
SPSS, Inc., Chicago, IL, USA) and Graph Pad Prism 5 (GraphPad
Software, Inc., La Jolla, CA, USA) were used for all statistical
analyses. Differences between groups were examined by Student's
t-test for two groups and one-way analysis of variance followed by
a least significant difference test for multiple comparisons.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Establishment of an injured VECs-VSMCs
co-culture model
Initially different concentrations of LPC were used
in order to treat VECs and detect the secretion of IL-6 and TNF-α.
The results indicated that LPC induced VECs injury in a
dose-dependent manner. LPC (20–50 µM) significantly promoted the
secretion of IL-6 and TNF-α (Fig. 1A
and B). An increase in proliferation and migration of VSMCs was
observed that was caused by co-culture with LPC-injured VECs
(Fig. 1C-E). Consequently, 30 µM
LPC was selected in order to produce the injured VECs-VSMCs
co-culture model.
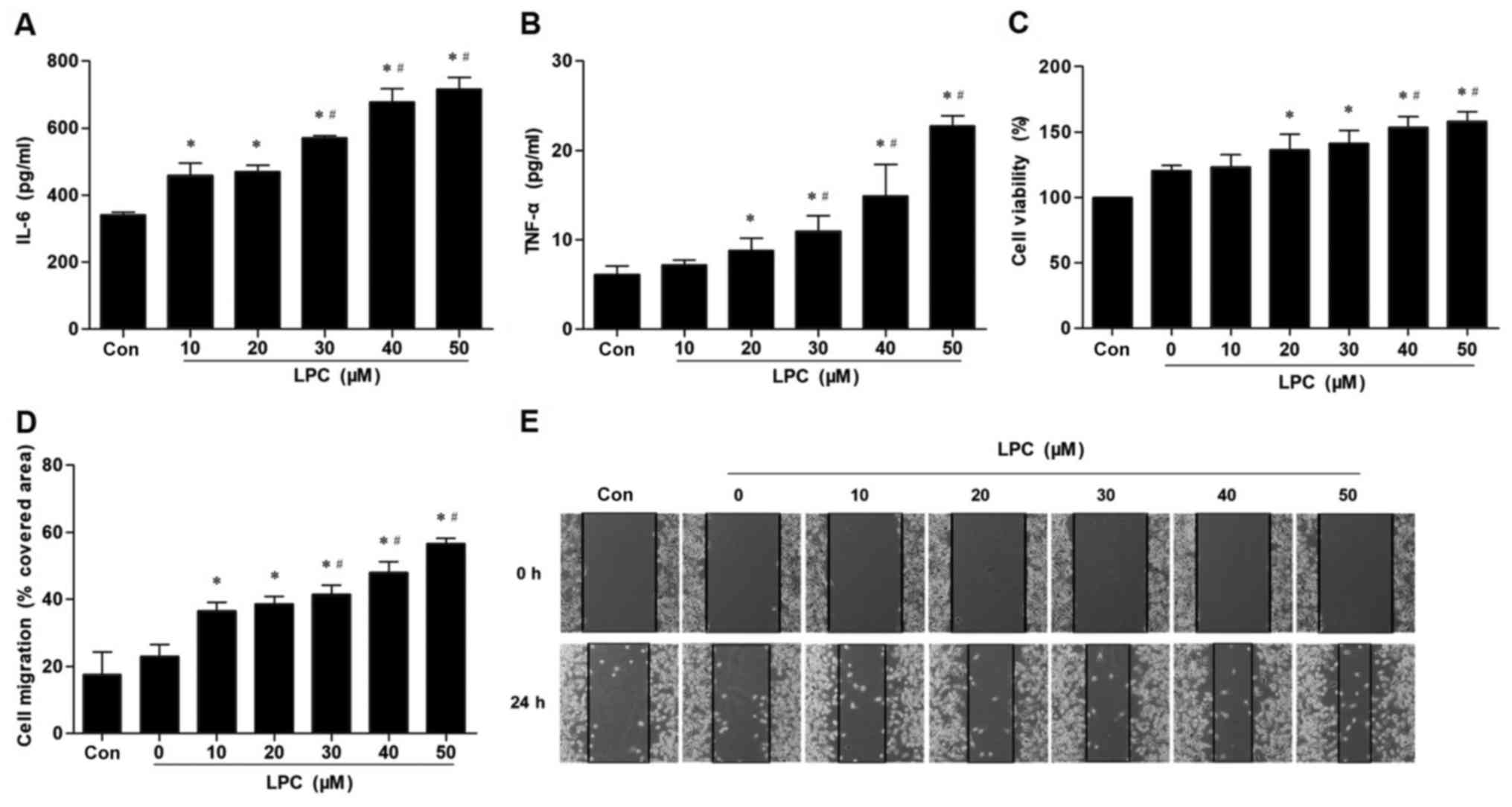 | Figure 1.Effect of LPC on secretion of IL-6
and TNF-α in VECs, and proliferation and migration of VSMCs induced
by co-culture with injured VECs. LPC (20, 30, 40 and 50 µM)
significantly increased the levels of (A) IL-6 and (B) TNF-α in
VECs-conditioned medium, in a concentration-dependent manner.
Pre-treatment of VECs with LPC (20, 30, 40 and 50 µM) significantly
enhanced VMSC (C) survival and (D) migratory ability demonstrated
by (E) wound healing induced by co-culture with injured VECs. Data
are presented as the mean ± standard deviation of three independent
experiments. *P<0.05 vs. Con. #P<0.05 vs. 10 and
20 µM LPC. LPC, lysophosphatidylcholine; IL, interleukin; TNF-α,
tumor necrosis factor-α; Con, control. |
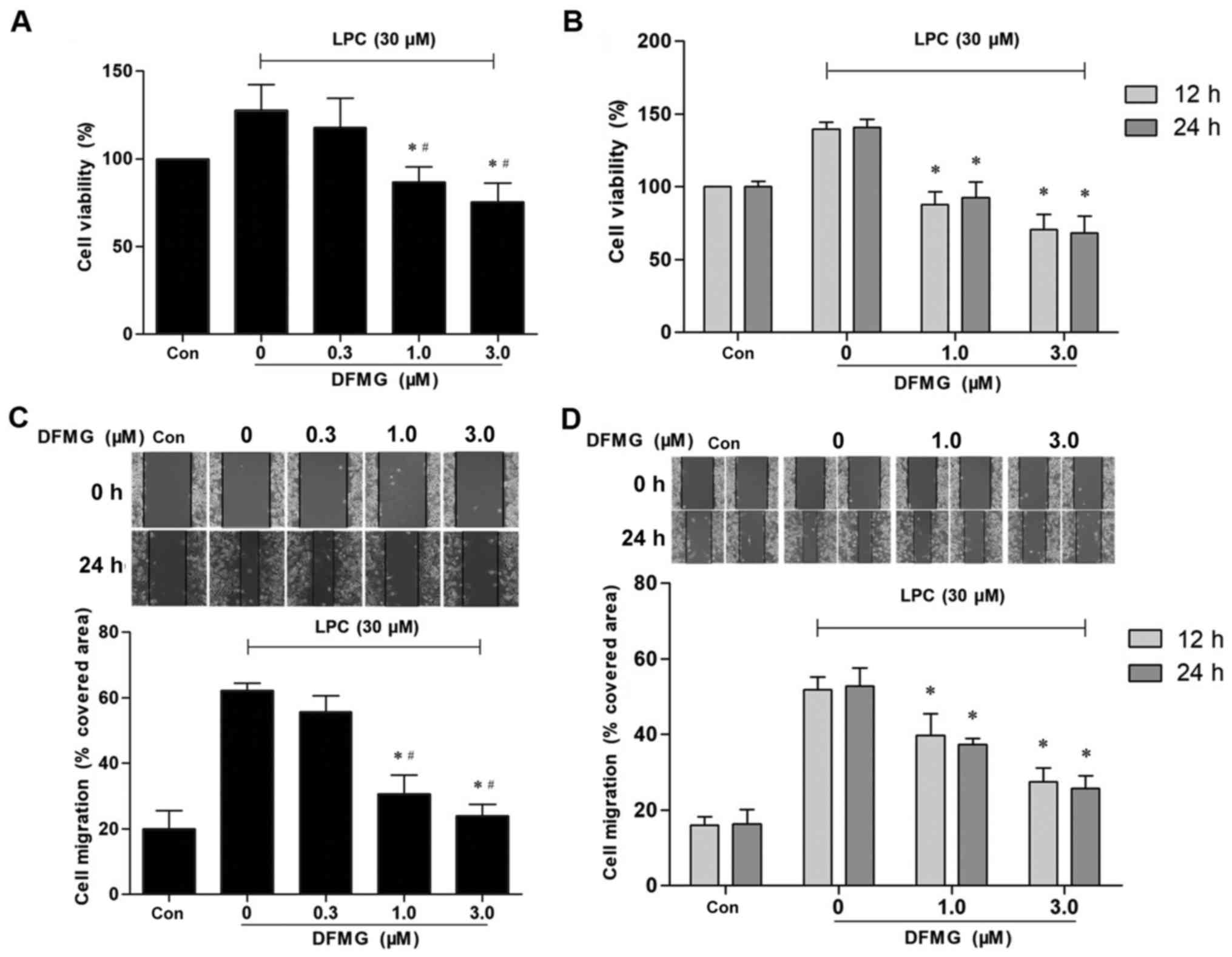
DFMG reverses the proliferation and
migration of VSMCs induced by co-culture with injured VECs
To determine whether DFMG protects against
proliferation and migration of VSMCs induced by co-culture with
LPC-injured VECs, VECs were pretreated with various concentrations
of DFMG for various durations prior to treatment with LPC (30 µM).
A significant decrease in the proliferation and migration of VSMCs
was observed (Fig. 2).
Proliferation and migration of VSMCs decreased following treatment
with ≥1.0 µM DFMG (Fig. 2A and C).
Cell viability of the control group was set as 100%. VECs were
pretreated with 1.0 µM DFMG and the relative cell viability of
VSMCs was ~86.7%, whereas the corresponding healing rate was
~30.73% (Fig. 2A and C).
Proliferation and migration of VSMCs was markedly reduced following
pretreatment of VECs for 12 h with 1.0 or 3.0 µM DFMG (Fig. 2B and D). The longer incubation time
(24 h) demonstrated no significant effect on migration and
proliferation of VSMCs (Fig. 2B and
D). Thus, 1.0 µM DFMG was selected as the optimum concentration
for the pretreatment of VECs for 12 h.
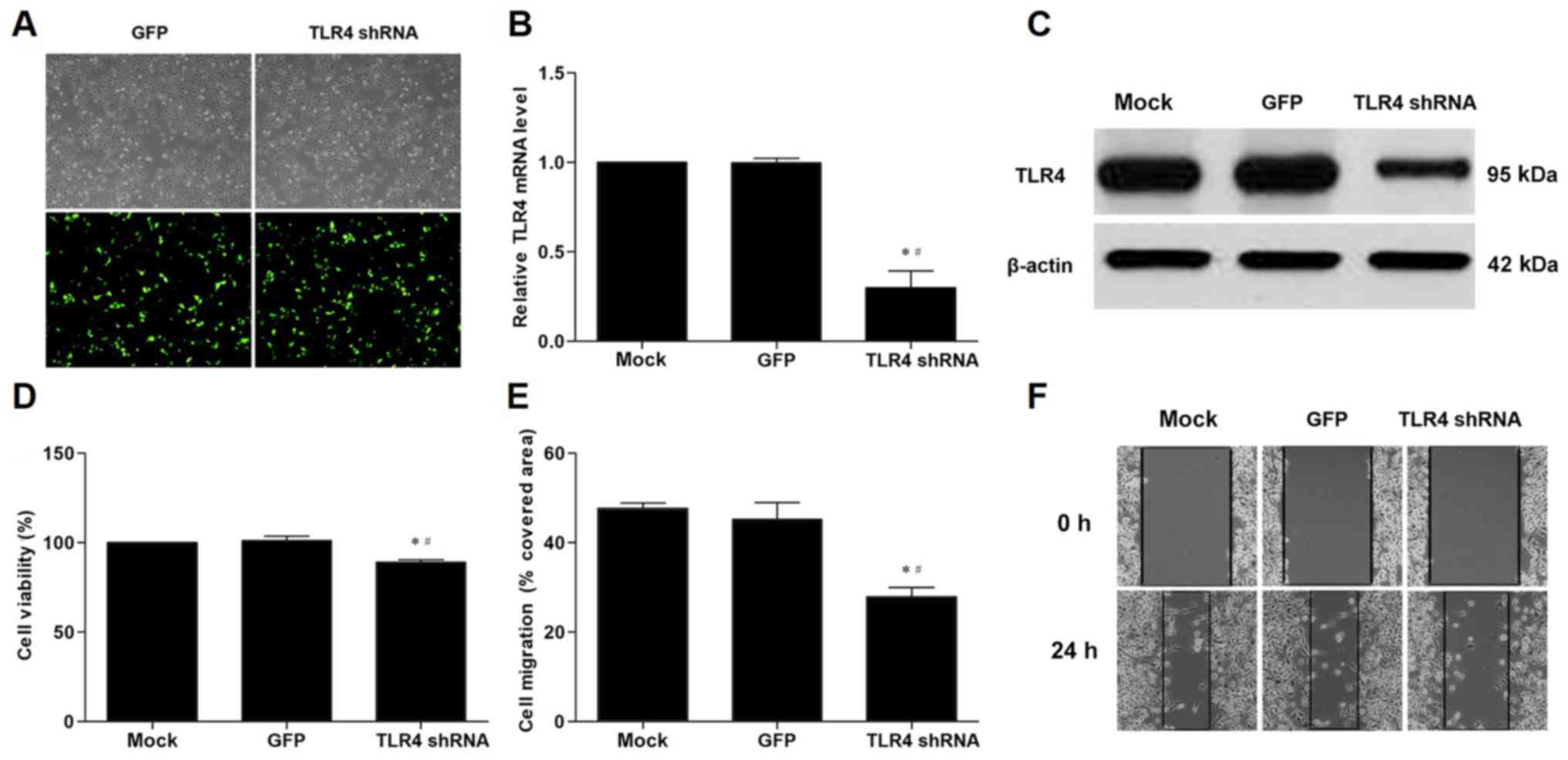
Proliferation and migration of VSMCs
co-cultured with VECs is inhibited by transfection with TLR4
shRNA
To further investigate the role of TLR4,
transfection of TLR4 shRNA was used in order to silence TLR4 gene
expression in VECs. VECs transfected with TLR4 shRNA exhibited
significantly decreased expression of TLR4 mRNA and protein levels
compared with those transfected with the GFP gene (Fig. 3A-C). TLR4 shRNA significantly
inhibited the proliferation and migration of VSMCs co-cultured with
VECs compared with the GFP control (Fig. 3D-F).
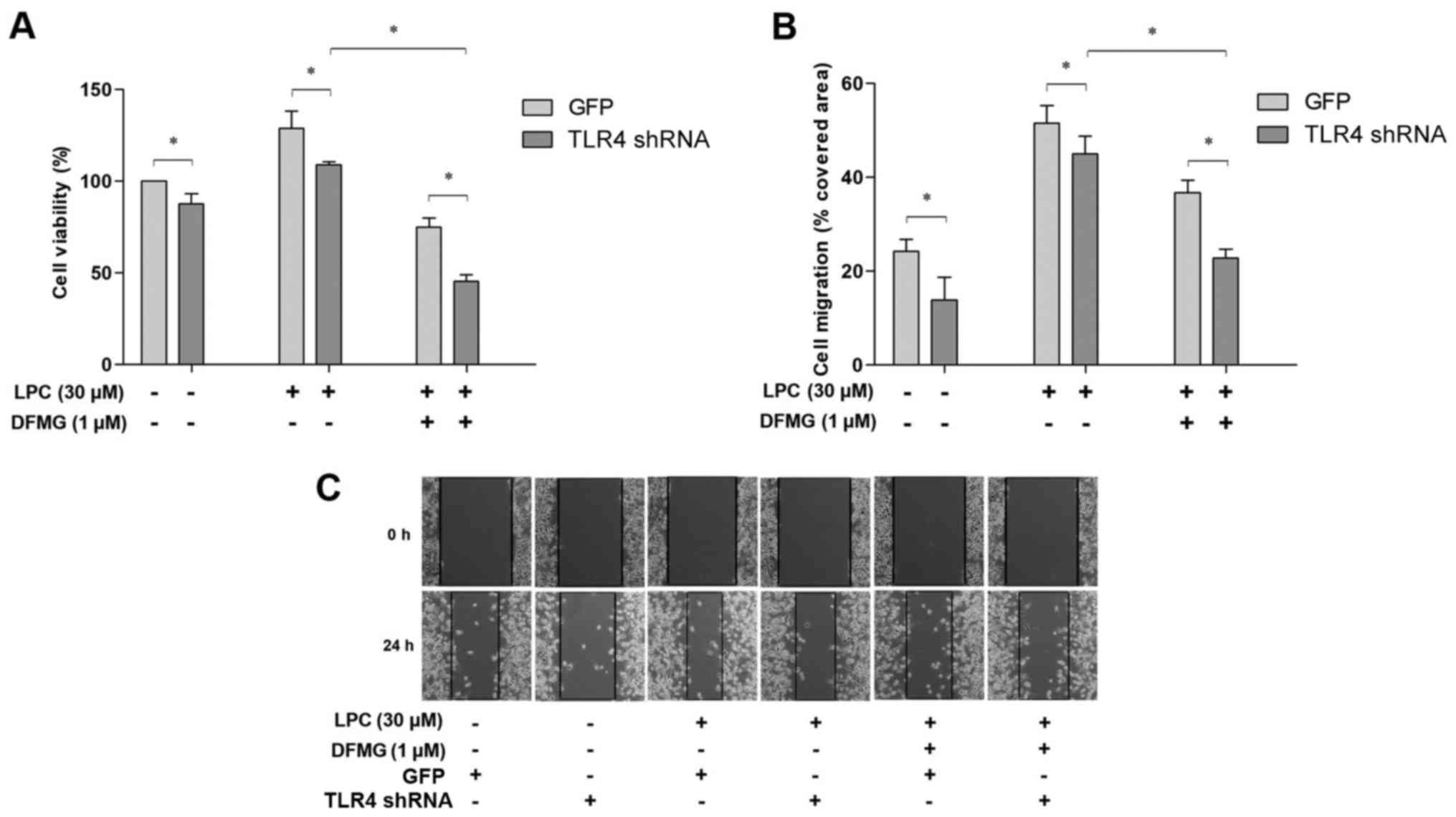
Proliferation and migration of VSMCs
co-cultured with LPC-injured VECs is inhibited by DFMG treatment
and TLR4 shRNA transfection
The effects of DFMG in combination with TLR4 shRNA
transfection of VECs were investigated in order to determine
whether DFMG could reverse the effects of LPC-injured VECs on the
proliferation and migration of VSMCs. The results indicated that
DFMG and TLR4 shRNA suppressed proliferation and migration of
VSMCs, and the combination of these interventions was more
effective than TLR4 knockdown alone (Fig. 4). DFMG and TLR4 shRNA exhibited a
synergistic effect in promoting the stability of VSMCs.
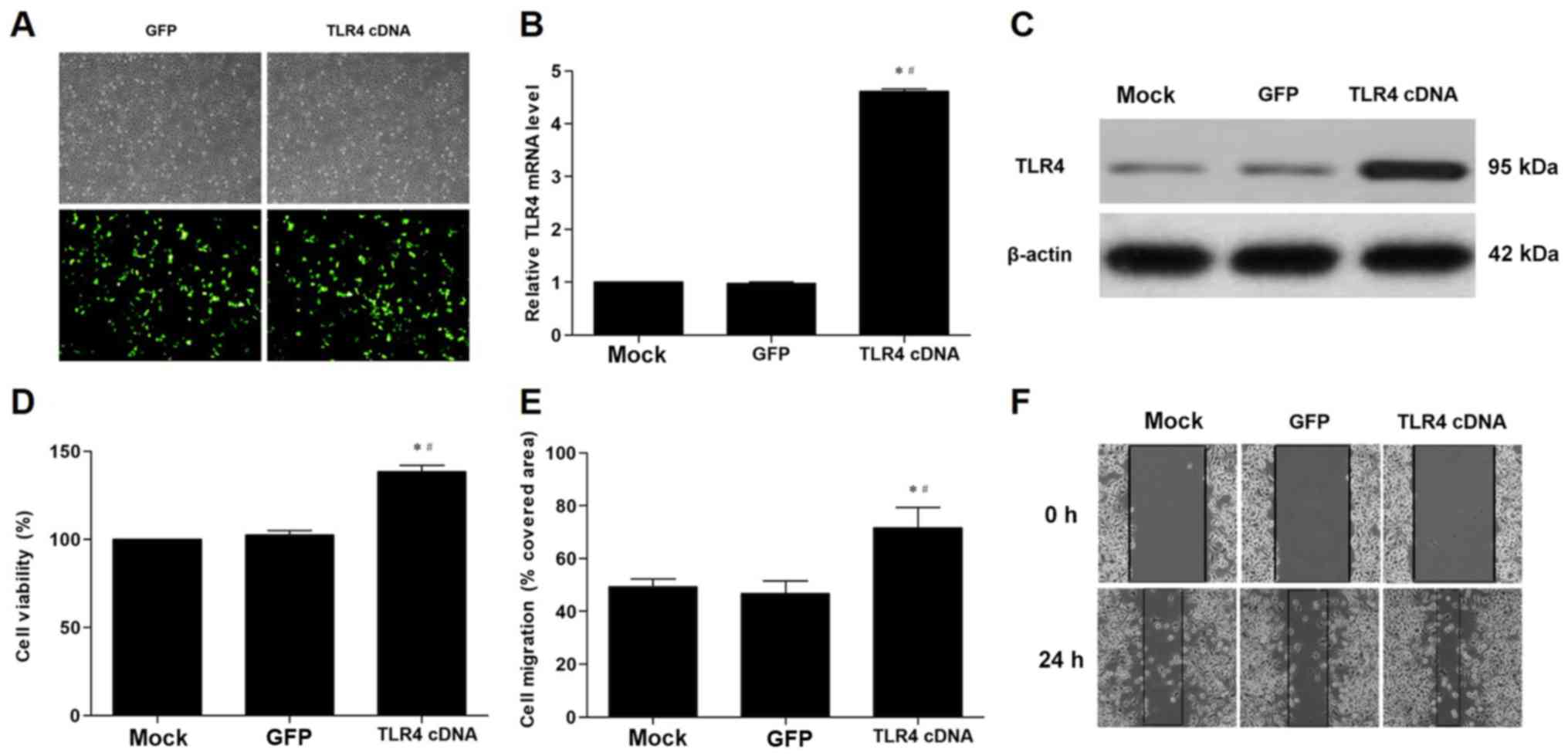
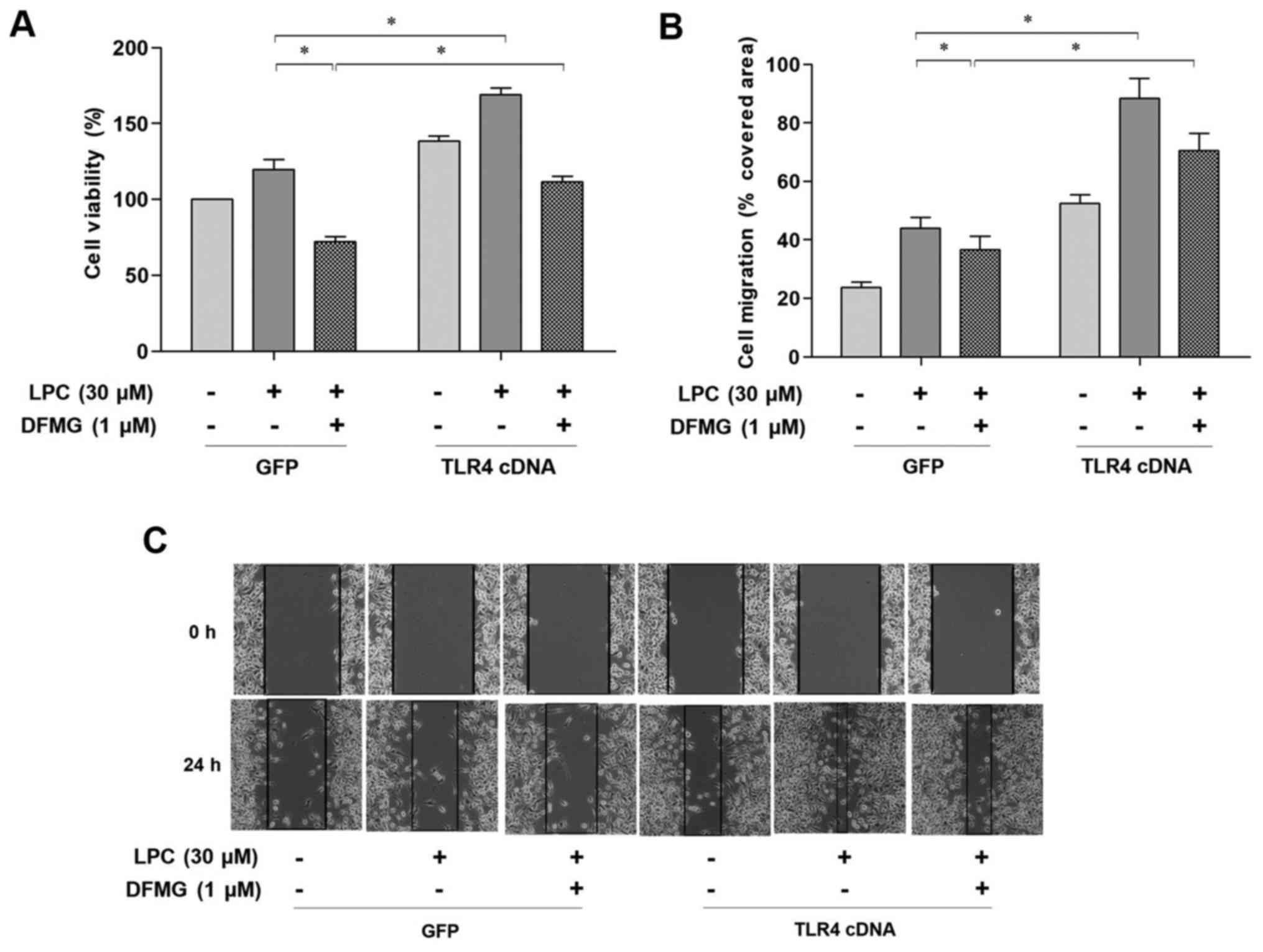
TLR4 overexpression in VECs promotes
proliferation and migration of VSMCs induced by co-culture with
VECs
TLR4 cDNA significantly increased the expression of
TLR4 mRNA and protein in VECs compared with the cells transfected
with the GFP gene (Fig. 5A-C).
Furthermore, transfection of the cells with TLR4 cDNA significantly
enhanced the proliferation and migration of VSMCs in the co-culture
model compared with the GFP control (Fig. 5D-F).
TLR4 cDNA transfection of VECs
enhances injured VEC-induced proliferation and migration of VSMCs,
which in turn attenuates the protective effect of DFMG
Combined treatment with DFMG and TLR4 cDNA
transfection of VECs was performed in order to investigate whether
TLR4 was involved in the DFMG mediated anti-migratory and
anti-proliferative effect in VSMCs. The results indicated that TLR4
cDNA transfection significantly attenuated the inhibitory effects
of DFMG on the proliferation and migration of VSMCs (Fig. 6).
Discussion
During the pathogenesis of atherosclerosis, the
endothelial dysfunction is considered to be an initial step.
Various risk factors lead to structural and functional alteration
in the vascular endothelium, including lipid deposition into the
intima, infiltration of inflammatory cytokines (IL-6, IL-8, TNF-α),
proliferation and migration of VSMCs, synthesis of extracellular
matrix, and the formation of foam cells and plaques (30). Oxidation and enzymatic modification
of low-density lipoproteins (LDL) leads to synthesis of LPC
(31). LPC is a component of
oxidized-LDL (ox-LDL) that can stimulate ox-LDL to cause AS
(32). In the present study, LPC
stimulated secretion of IL-6 and TNF-α in VECs, and promoted the
proliferation and migration of VSMCs in a co-culture model. These
processes are crucial to neointima formation (33).
TLR4 is involved in the inflammatory reaction.
Recently, the interaction between the TLR4 transduction pathway and
AS has been extensively investigated (34–37).
In the present study, a transient transfection was used in order to
modulate the expression of TLR4 mRNA and protein in VECs. The
results of the present study indicated that TLR4 cDNA promoted the
proliferation and migration of VSMCs in a co-culture model, while
TLR4 shRNA exerted the opposite effect. Furthermore, the results of
the present study further supported the role of TLR4 signaling in
the proliferation and migration of VSMCs induced by co-culture with
LPC-injured VECs.
Genistein, the lead compound of DFMG, is a natural
isoflavone present in legumes and dentate plants. Genistein can
alleviate inflammatory damage by altering TLR4/NF-κB signaling
(22,23). In the present study, VECs were
pretreated with DFMG prior to treatment with LPC and subsequently
co-cultured with VSMCs. The results demonstrated that the
proliferation and migration of VSMCs were decreased, which was
dependent on the concentration of DFMG. In addition, the
combination of treatment with DFMG and TLR4 shRNA transfection of
VECs significantly suppressed the proliferation and migration of
VSMCs induced by the co-culture with LPC-injured VECs. Inhibition
of TLR4 signaling in VECs restrained abnormal proliferation and
migration of VSMCs. TLR4 cDNA transfection of VECs enhanced
proliferation and migration of VSMCs that were induced by
co-culture with injured VECs, and attenuated the effects of
DFMG.
In conclusion, the results of the present study
demonstrated that DFMG effectively inhibited proliferation and
migration of VSMCs induced by co-culture with LPC-injured VECs via
regulation of the TLR4 signaling pathway. DFMG may be useful as a
novel agent for the prevention of AS. However, the present study
has certain limitations, including that the effects of DFMG on
VSMCs have not been investigated in vivo. Furthermore,
regulation of the TLR4/myeloid differentiation primary response 88
and TLR4/Toll-like receptor adaptor molecule 1 pathways by DFMG
during this process has not been investigated. Consequently,
understanding of detailed effects of DFMG on the TLR4 signaling
pathway in AS requires further investigation.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from the
Natural Science Foundation of China (grant no. 81370382).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
XF designed the study and prepared the manuscript.
LC and HH performed the experiments. YZ analysed the data. JC
contributed to the interpretation of some of the data and revised
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AS
|
atherosclerosis
|
|
DFMG
|
7-difluoromethoxy-5,4′-dimethoxy-genistein
|
|
IL-6
|
interleukin-6
|
|
LPC
|
lysophosphatidylcholine
|
|
TLR4
|
Toll-like receptor 4
|
|
TNF-α
|
tumor necrosis factor-α
|
|
VECs
|
vascular endothelial cells
|
|
VSMCs
|
vascular smooth muscle cells
|
References
|
1
|
Heidari M, Mandato CA and Lehoux S:
Vascular smooth muscle cell phenotypic modulation and the
extracellular matrix. Artery Res. 9:14–18. 2015. View Article : Google Scholar
|
|
2
|
Lin X, He Y, Hou X, Zhang Z, Wang R and Wu
Q: Endothelial cells can regulate smooth muscle cells in
contractile phenotype through the miR-206/ARF6&NCX1/exosome
axis. PLoS One. 11:e01529592016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Afzal TA, Luong LA, Chen D, Zhang C, Yang
F, Chen Q, An W, Wilkes E, Yashiro K, Cutillas PR, et al: NCK
associated protein 1 modulated by miRNA-214 determines vascular
smooth muscle cell migration, proliferation, and neointima
hyperplasia. J Am Heart Assoc. 5(pii): e0046292016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ostriker A, Horita HN, Poczobutt J,
Weiser-Evans MC and Nemenoff RA: Vascular smooth muscle
cell-derived transforming growth factor-β promotes maturation of
activated, neointima lesion-like macrophages. Arterioscler Thromb
Vasc Biol. 34:877–886. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Owens GK, Kumar MS and Wamhoff BR:
Molecular regulation of vascular smooth muscle cell differentiation
in development and disease. Physiol Rev. 84:767–801. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gimbrone MA Jr and García-Cardeña G:
Endothelial cell dysfunction and the pathobiology of
atherosclerosis. Circ Res. 118:620–636. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ding H, Li D, Zhang Y, Zhang T, Zhu H, Xu
T, Luo Y and Wang C: Luteolin inhibits smooth muscle cell migration
and proliferation by attenuating the production of Nox4, p-Akt and
VEGF in endothelial cells. Curr Pharm Biotechnol. 14:1009–1015.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xia Y, Bhattacharyya A, Roszell EE, Sandig
M and Mequanint K: The role of endothelial cell-bound Jagged1 in
Notch3-induced human coronary artery smooth muscle cell
differentiation. Biomaterials. 33:2462–2472. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou J, Li YS, Nguyen P, Wang KC, Weiss A,
Kuo YC, Chiu JJ, Shyy JY and Chien S: Regulation of vascular smooth
muscle cell turnover by endothelial cell-secreted microRNA-126:
Role of shear stress. Circ Res. 113:40–51. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li Y, Guo Y, Chen Y, Wang Y, You Y, Yang
Q, Weng X, Li Q, Zhu X, Zhou B, et al: Establishment of an
interleukin-1β-induced inflammation-activated endothelial
cell-smooth muscle cell-mononuclear cell co-culture model and
evaluation of the anti-inflammatory effects of tanshinone IIA on
atherosclerosis. Mol Med Rep. 12:1665–1676. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Satoh S, Yada R, Inoue H, Omura S, Ejima
E, Mori T, Takenaka K, Kawamura N, Numaguchi K, Mori E, et al:
Toll-like receptor-4 is upregulated in plaque debris of patients
with acute coronary syndrome more than Toll-like receptor-2. Heart
Vessels. 31:1–5. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Roshan MH, Tambo A and Pace NP: The role
of TLR2, TLR4, and TLR9 in the pathogenesis of atherosclerosis. Int
J Inflam. 2016:15328322016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lim S and Park S: Role of vascular smooth
muscle cell in the inflammation of atherosclerosis. BMB Rep.
47:1–7. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xing S, Zheng F, Zhang W, Wang D and Xing
Q: Relationship between toll-like receptor 4 levels in aorta and
severity of atherosclerosis. J Int Med Res. 42:958–965. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kapelouzou A, Giaglis S, Peroulis M,
Katsimpoulas M, Moustardas P, Aravanis CV, Kostakis A, Karayannakos
PE and Cokkinos DV: Overexpression of toll-like receptors 2, 3, 4
and 8 is correlated to the vascular atherosclerotic process in the
hyperlipidemic rabbit model: The effect of statin treatment. J Vasc
Res. 54:156–169. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tang YL, Jiang JH, Wang S, Liu Z, Tang XQ,
Peng J, Yang YZ and Gu HF: TLR4/NF-κB signaling contributes to
chronic unpredictable mild stress-induced atherosclerosis in
ApoE-/- mice. PLoS One. 10:e01236852015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pahwa R, Nallasamy P and Jialal I:
Toll-like receptors 2 and 4 mediate hyperglycemia induced
macrovascular aortic endothelial cell inflammation and perturbation
of the endothelial glycocalyx. J Diabetes Complications.
30:563–572. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang XQ, Wan HQ, Wei XJ, Zhang Y and Qu P:
CLI-095 decreases atherosclerosis by modulating foam cell formation
in apolipoprotein E-deficient mice. Mol Med Rep. 14:49–56. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang X, Coriolan D, Murthy V, Schultz K,
Golenbock DT and Beasley D: Proinflammatory phenotype of vascular
smooth muscle cells: Role of efficient Toll-like receptor 4
signaling. Am J Physiol Heart Circ Physiol. 289:H1069–H1076. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang D, Li D, Cao L, Wang L, Zhu S, Xu T,
Wang C and Pan D: Positive feedback regulation of proliferation in
vascular smooth muscle cells stimulated by lipopolysaccharide is
mediated through the TLR 4/Rac1/Akt pathway. PLoS One.
9:e923982014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li H, Xu H and Liu S: Toll-like receptors
4 induces expression of matrix metalloproteinase-9 in human aortic
smooth muscle cells. Mol Biol Rep. 38:1419–1423. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhou X, Yuan L, Zhao X, Hou C, Ma W, Yu H
and Xiao R: Genistein antagonizes inflammatory damage induced by
β-amyloid peptide in microglia through TLR4 and NF-κB. Nutrition.
30:90–95. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ma W, Ding B, Yu H, Yuan L, Xi Y and Xiao
R: Genistein alleviates β-amyloid-induced inflammatory damage
through regulating Toll-like receptor 4/nuclear factor κB. J Med
Food. 18:273–279. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu F, Cao JG, Li C, Tan JS and Fu XH:
Protective effects of 7-difluoromethyl-5,4′-dimethoxygenistein
against human aorta endothelial injury caused by lysophosphatidyl
choline. Mol Cell Biochem. 363:147–155. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu S, Li L, Zhang J, Huang H, Yang S, Ren
C, Fu X and Zhang Y: 7-difluoromethyl-5, 4′-dimethoxygenistein
reverses LPC-induced apoptosis of HUVE-12 cells through regulating
mitochondrial apoptosis pathway. Curr Signal Transd Ther. 9:50–58.
2014. View Article : Google Scholar
|
|
26
|
Zhang Y, Li L, You J, Cao J and Fu X:
Effect of 7-difluoromethyl-5, 4′-dimethoxygenistein on aorta
atherosclerosis in hyperlipidemia ApoE(−/−) mice induced by a
cholesterol-rich diet. Drug Des Devel Ther. 7:233–242. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fu XH, Wang L, Zhao H, Xiang HL and Cao
JG: Synthesis of genistein derivatives and determination of their
protective effects against vascular endothelial cell damages caused
by hydrogen peroxide. Bioorg Med Chem Lett. 18:513–517. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen L, DeWispelaere A, Dastvan F, Osborne
WR, Blechner C, Windhorst S and Daum G: Smooth muscle-alpha actin
inhibits vascular smooth muscle cell proliferation and migration by
inhibiting Rac1 activity. PLoS One. 11:e01557262016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cibor D, Domagala-Rodacka R, Rodacki T,
Jurczyszyn A, Mach T and Owczarek D: Endothelial dysfunction in
inflammatory bowel diseases: Pathogenesis, assessment and
implications. World J Gastroenterol. 22:1067–1077. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sevastou I, Kaffe E, Mouratis MA and
Aidinis V: Lysoglycerophospholipids in chronic inflammatory
disorders: The PLA(2)/LPC and ATX/LPA axes. Biochim Biophys Acta.
1831:42–60. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Voight BF, Peloso GM, Orho-Melander M,
Frikke-Schmidt R, Barbalic M, Jensen MK, Hindy G, Hólm H, Ding EL,
Johnson T, et al: Plasma HDL cholesterol and risk of myocardial
infarction: A mendelian randomisation study. Lancet. 380:572–580.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang X, Hu G, Gao X, Wang Y, Zhang W,
Harmon EY, Zhi X, Xu Z, Lennartz MR, Barroso M, et al: The
induction of yes-associated protein expression after arterial
injury is crucial for smooth muscle phenotypic modulation and
neointima formation. Arterioscler Thromb Vasc Biol. 32:2662–2669.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Luo H, Wang J, Qiao C, Ma N, Liu D and
Zhang W: Pycnogenol attenuates atherosclerosis by regulating lipid
metabolism through the TLR4-NF-κB pathway. Exp Mol Med.
47:e1912015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee GL, Wu JY, Tsai CS, Lin CY, Tsai YT,
Lin CS, Wang YF, Yet SF, Hsu YJ and Kuo CC: TLR4-activated
MAPK-IL-6 axis regulates vascular smooth muscle cell function. Int
J Mol Sci. 17(pii): E13942016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hosseini H, Li Y, Kanellakis P, Tay C, Cao
A, Liu E, Peter K, Tipping P, Toh BH, Bobik A and Kyaw T: Toll-like
receptor (TLR)4 and MyD88 are essential for atheroprotection by
peritoneal B1a B cells. J Am Heart Assoc. 5(pii): e0029472016.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ding Y, Subramanian S, Montes VN,
Goodspeed L, Wang S, Han C, Teresa AS III, Kim J, O'Brien KD and
Chait A: Toll-like receptor 4 deficiency decreases atherosclerosis
but does not protect against inflammation in obese low-density
lipoprotein receptor deficient mice. Arterioscler Thromb Vasc Biol.
32:1596–1604. 2012. View Article : Google Scholar : PubMed/NCBI
|