Introduction
Liver fibrosis occurs in response to chronic liver
injuries, including hepatitis B and C, alcohol consumption, fatty
liver disease, cholestasis and autoimmune hepatitis (1). Activated hepatic stellate cells
(HSCs) are the primary source of extracellular matrix (ECM) in the
liver. Inhibiting the activation and promoting the apoptosis of
HSCs would be an ideal therapeutic strategy to limit liver fibrosis
(2). In recent decades, studies
have revealed that the senescence of activated HSCs reduces liver
fibrosis. The senescent HSCs have reduced viability, downregulated
ECM component production, increased levels of ECM-degrading enzymes
and enhanced immune surveillance (3). This indicates that promoting the
senescence of activated HSCs may be an effective strategy in liver
fibrosis therapy.
Senescence is a cellular process that irreversibly
arrests the proliferation of damaged cells and induces the
secretion of the inflammatory mediators IL-6, IL-8 and TNF-α which
are part of a larger senescence associated secretory phenotype
(SASP) (4). Cytokines have various
roles in the development and/or promotion of premature cellular
senescence (5). IL-6 regulates
premature cellular senescence by activating its downstream signal
transducer and activator of transcription3 (STAT3) and chemokine
IL-8 has been demonstrated to reinforce the senescence process of
cells (6). The inflammatory
cytokine TNF-α promotes the premature senescence of rat nucleus
pulposus cells via the PI3K/Akt signaling pathway (7). Interleukin-10 (IL-10) is a cytokine
that downregulates the pro-inflammatory response and has a
modulatory effect on hepatic fibrogenesis (8). Previous studies have demonstrated
that IL-10 attenuates carbon tetrachloride- and porcine
serum-induced liver fibrosis (9,10).
However, to the best of the author's knowledge, the role of IL-10
in activating HSC senescence has not yet been reported. The aim of
present study was to obtain high-purity and yield primary rat HSCs
to evaluate the effects of IL-10 on the senescence of activated
HSCs and to examine the underlying mechanisms. The present study
may provide a new insight on anti-fibrotic therapy of IL-10.
Materials and methods
Animals
Fifteen male Sprague-Dawley rats (12–16-week-old)
were provided by the Shanghai Experimental Animal Center (Shanghai,
China). The rats were housed at 22±2°C with 55±5% humidity and a 12
h alternating light/dark cycle. Rats had access to water and food
ad libitum. The present study was approved by the Animal
Care Committee of Fujian Medical University (Fuzhou, China) and
experiments were performed in accordance with the Guidelines on
Animal Experiments in Fujian Medical University (Fuzhou,
China).
Isolation and culture of primary rat
HSCs
Rat HSC isolation was performed as previously
described (11). Rat livers were
perfused in situ and HSCs were isolated by density gradient
centrifugation. Rat livers were perfused via the portal vein with
D-Hanks buffer (PYG-0079, Boster Biological Technology, Wuhan,
China) and subsequently with 0.075% type I collagenase for 10 min
(Worthington Biochemical Corporation, Lakewood, NJ, USA). Livers
were further digested by shaking for 20 min at 200 rpm and 37°C.
Cell suspension was filtered to remove undigested debris and Hanks
buffer was added to make up the solution to 50 ml. This was
centrifuged at 50 × g for 5 min at room temperature to remove
residual hepatocytes. Supernatant was subsequently transferred to a
clean 50 ml tube and centrifuged at 500 × g for 5 min at 4°C. Cell
pellets were resuspended in 12% Histodenz (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) and centrifuged by density gradient for
HSC isolation. The newly isolated HSCs were seeded in 6-well plates
(2×106 cells/well) and cultivated in Dulbecco's modified
Eagle's medium (DMEM; Hyclone; GE Healthcare Life Sciences, Logan,
UT, USA) supplemented with 20% fetal bovine serum (FBS; Hyclone; GE
Healthcare Life Sciences) at 37°C in an environment containing 5%
CO2 for 48 h. Following seeding, HSCs were cultured in
DMEM (Hyclone; GE Healthcare Life Sciences) supplemented with 10%
FBS (Hyclone; GE Healthcare Life Sciences) and 1% penicillin and 1%
streptomycin.
Identification of primary rat HSCs
Cell viability and yield
An equal volume (0.1 ml) of DMEM-suspended HSCs were
mixed with 0.4% Trypan blue solution. An aliquot of the mixture was
analyzed in a Neubauer chamber following 5 min at room temperature
and viable and non-viable cells within four big squares in the four
corners were counted under a phase contrast microscopy
(magnification, ×100) The cell viability (%) was calculated as
follows: Total viable cells (unstained)/total cells (stained and
unstained) ×100%.
Autofluorescence detection of
HSCs
Primary HSCs were seeded in 6-well plates
(2×105 cells/well) and cultured in DMEM (Hyclone; GE
Healthcare Life Sciences) supplemented with 10% FBS (Hyclone; GE
Healthcare Life Sciences). Vitamin A lipid droplet autofluorescence
in primary rat HSCs was detected at a wavelength of 328 nm using a
fluorescence microscope (magnification, ×200; Nikon Corporation,
Tokyo, Japan).
Oil Red O staining
Cells were fixed in 4% paraformaldehyde for 15 min
at room temperature, washed with ice cold PBS and stained with 0.3%
oil red O solution for 10 min at the room temperature. Stained
slides were washed and counterstained with hematoxylin Staining
Solution (Beyotime Institute of Biotechnology, shanghai, China) for
30 sec at room temperature and washed again in PBS. Positive cells
were observed from ten random fields under a phase contrast
microscope (magnification, ×200).
Immunocytochemistry assay
Cells were fixed in 4% paraformaldehyde for 15 min
at room temperature and washed twice with ice cold PBS, and then
permeabilized with 0.3% Triton X-100 for 15 min. Endogenous
peroxidase was removed with 30 ml/l H2O2.
They were incubated with mouse anti-rat α-smooth muscle actin
(α-SMA, 1:400, cat. no. BM0002; Boster Biological Technology) and
rabbit anti-rat desmin (1:200, cat. no. BM0036; Boster Biological
Technology) primary antibodies in a humidified chamber overnight at
4°C. Cells were treated with instant Polink-2 plus®
Polymer HRP Detection System (cat. no. PV-9001/PV-9002; ZSGB-BIO;
OriGene Technologies, Inc.) and incubated in a buffer solution
containing 3,3-diaminobenzidine tetrahydrochloride (DAB) to produce
a brown reaction product. Finally, cells were counterstained with
hematoxylin Staining Solution (C0107; Beyotime Institute of
Biotechnology) dehydrated, plated on coverslips and visualized
under a phase contrast microscopy (magnification, ×200). Counting
the total 200 cells on the cover glass, the purity of hepatic
stellate cells (%) = Desmin positive cells/total (positive and
negative) ×100%. the degree of HSC activation (%) = a-SMA positive
cells/total (positive and negative) ×100%.
Primary rat HSCs culture protocol
Primary rat HSCs were seeded in 6-well plates
(2×105 cells/well) and cultured in DMEM (Hyclone; GE
Healthcare Life Sciences) supplemented with 10% FBS (Hyclone; GE
Healthcare Life Sciences). When cells reached 60–70% confluence,
the culture medium was replaced with serum-free DMEM for 24 h and
subsequently divided into the control and IL-10 treatment groups.
Control group HSCs were cultured with DMEM supplemented with 10%
FBS, whereas cells in the IL-10 treatment group were cultured with
increasing concentrations of IL-10 (10, 20 or 40 ng/ml; PeproTech,
Inc., Rocky Hill, NJ, USA) for a further 24 h at 37°C.
Primary rat HSCs senescence
analysis
The Senescence β-Galactosidase Staining kit (C0602;
Beyotime Institute of Biotechnology) was used to detect the
senescence-associated β-galactosidase (SA-β-Gal) activity of
activated HSCs. Briefly, HSCs treated with or without IL-10 were
fixed with the staining fixative for 15 min at room temperature,
washed with PBS and stained overnight with X-Gal solution at 37°C.
Cells were washed twice with PBS and incubated with
4′,6-diamidino-2-phenlindole dihydrochloride (0.5 µg/ml; Roche
Diagnostics, Indianapolis, IN, USA) for 10 min. SA-β-Gal-positive
cells were detected in at least three fields with a phase contrast
microscope (magnification, ×200) using Image-Pro Plus software
(version 6.0; Media Cybernetics, Inc., Rockville, MD, USA).
Cell viability assay
The Cell Counting kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) was used to examine cell
viability. Primary rat HSCs were seeded in 96-well plates
(3×103 cells/well) in DMEM (Hyclone; GE Healthcare Life
Sciences) supplemented with 10% FBS (Hyclone; GE Healthcare Life
Sciences) for 24 h and subsequently cultured without serum for 16 h
prior to culture with either DMEM supplemented with 10% FBS alone
or with increasing concentrations of IL-10 for 24 h. The medium was
replaced with fresh DMEM and 10 µl CCK-8 was added to each well for
2 h. The absorbency was determined with a microplate reader
(Epoch™; BioTek Instruments, Inc., Winooski, VT, USA) at
a wavelength of 450 nm. The absorbance values were normalized by
subtracting the blank values obtained from untreated cells.
Apoptosis assay
The FITC Annexin V Apoptosis Detection kit (cat. no.
556547; BD Biosciences, San Jose, CA, USA) was used to analyze the
apoptosis of activated primary rat HSCs. Activated primary rat HSCs
following treatment with or without IL-10 were washed twice with
cold PBS and suspended in 1X Binding Buffer at a concentration of
1×106 cells/ml. Subsequently, 100 µl of the solution
(1×105 cells) were added to 5 ml culture and add 5 µl of
FITC Annexin V and 5 µl PI for 15 min at room temperature in the
dark, 400 µl of 1X Binding Buffer were added to each tube and
analyzed using a FACScan cytometer (BD Accuri C6; BD
Biosciences).
ELISA assay
Primary rat HSCs were treated with or without IL-10
for 24 h. Cell culture supernatant was collected and assayed for
IL-6 (cat. no. R6000B), IL-8 (cat. no. D8000C) and tumour necrosis
factor (TNF)-α (cat. no. RTA00) expression levels using the
respective ELISA kits, according to the manufacturer's protocol
(R&B Systems, Minneapolis, MN, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from HSCs using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) and reverse transcribed to cDNA using a
Prime Script Reverse Transcription System (Takara Biotechnology
Co., Ltd., Dalian, China) according to the manufacturer's protocol.
cDNA samples were amplified using the SYBR-Green master mix kit
(Takara Biotechnology Co., Ltd.). The primer sequences used were as
follows: p53 forward, 5′-CTCCTCTCCCCAGCAAAAG-3′ and reverse,
5′-CCTGCTGTCTCCTGACTCCT-3′; p21 forward, 5′-TGTGGTAGTTGGAGCTGGTG-3′
and reverse, 5′-TGACCTGCTGTGTCGAGAAT-3′; and β-actin forward,
5′-GGCATCCTGACCCTGAAGTA-3′ and reverse, 5′-AGGCATACAGGGACAACACA-3′.
The PCR cycling conditions were as follows: 95°C for 2 min and 45
cycles of 95°C for 15 sec, 63°C for 15 sec, 72°C for 20 sec. All
procedures were performed in triplicate. Alterations in mRNA
expression were calculated relative to the expression control
β-actin using the 2−ΔΔCq method (12).
Western blot analysis
HSCs were washed twice with PBS and lysed with
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology) and blocked in a phenylmethylsulfonyl fluoride
(Beyotime Institute of Biotechnology) and PhosSTOP (Roche
Diagnostics) cocktail on ice. The supernatant was obtained by
centrifugation at 12,000 × g for 15 min at 4°C. Protein
concentration was determined by the enhanced BCA protein assay kit
(Beyotime Institute of Biotechnology). Equal amounts (40 µg) of
protein were separated on a 10% SDS-polyacrylamide gel and
transferred onto nitrocellulose membranes. Following blocking with
5% skim milk (cat. no. 1172GR500; BioFroxx GmbH, Einhausen,
Germany; www.neofroxx.com/en/home/) in Tris-buffered
saline/Tween-20 for 1 h at room temperature, the membranes were
incubated with the primary antibodies (p53 (cat. no. 2524, 1:1,000;
CST Biological Reagents, Co., Ltd., Shanghai, China), GAPDH (cat.
no. 2118, 1:1,000; CST Biological Reagents, Co., Ltd.), and p21
(cat. no. ab109199, 1:1,000; Abcam, Cambridge, UK) at 4°C
overnight, followed by incubation with horseradish
peroxidase-conjugated homologous secondary antibody (cat. no.
ZB-2301/ZB2305, 1:3,000; ZSGB-BIO; OriGene Technologies, Inc.,
Beijing, China) for 1 h at room temperature. The signals were
visualized using an enhanced chemiluminescence kit (Santa Cruz
Biotechnology, Inc., Dallas, TX, USA). The band density was
determined by densitometry, and quantified using
ChemiDoc™ Touch Imageing System with Image
Lab™ Touch Software 5.2 version. (Bio-Rad Laboratories,
Inc., Hercules, CA, USA).
Statistical analysis
Data are expressed as the mean ± standard error.
Statistical analysis was performed with one-way analysis of
variance followed by Tukey's Multiple Comparison Test, using SPSS
software, version 13.0 (SPSS, Inc., Chicago, IL, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
Isolation and identification of
primary rat HSCs
To examine the effects of IL-10 on primary rat HSC
senescence, isolation and culture of the HSCs was conducted. The
collagenase perfusion and density-gradient centrifugation method
(13) was used to isolate a
high-purity and high-yield of quiescent rat HSCs. These were
visible as a thin white layer in the interface between the
Histodenz solution and the overlay with DMEM following isolation
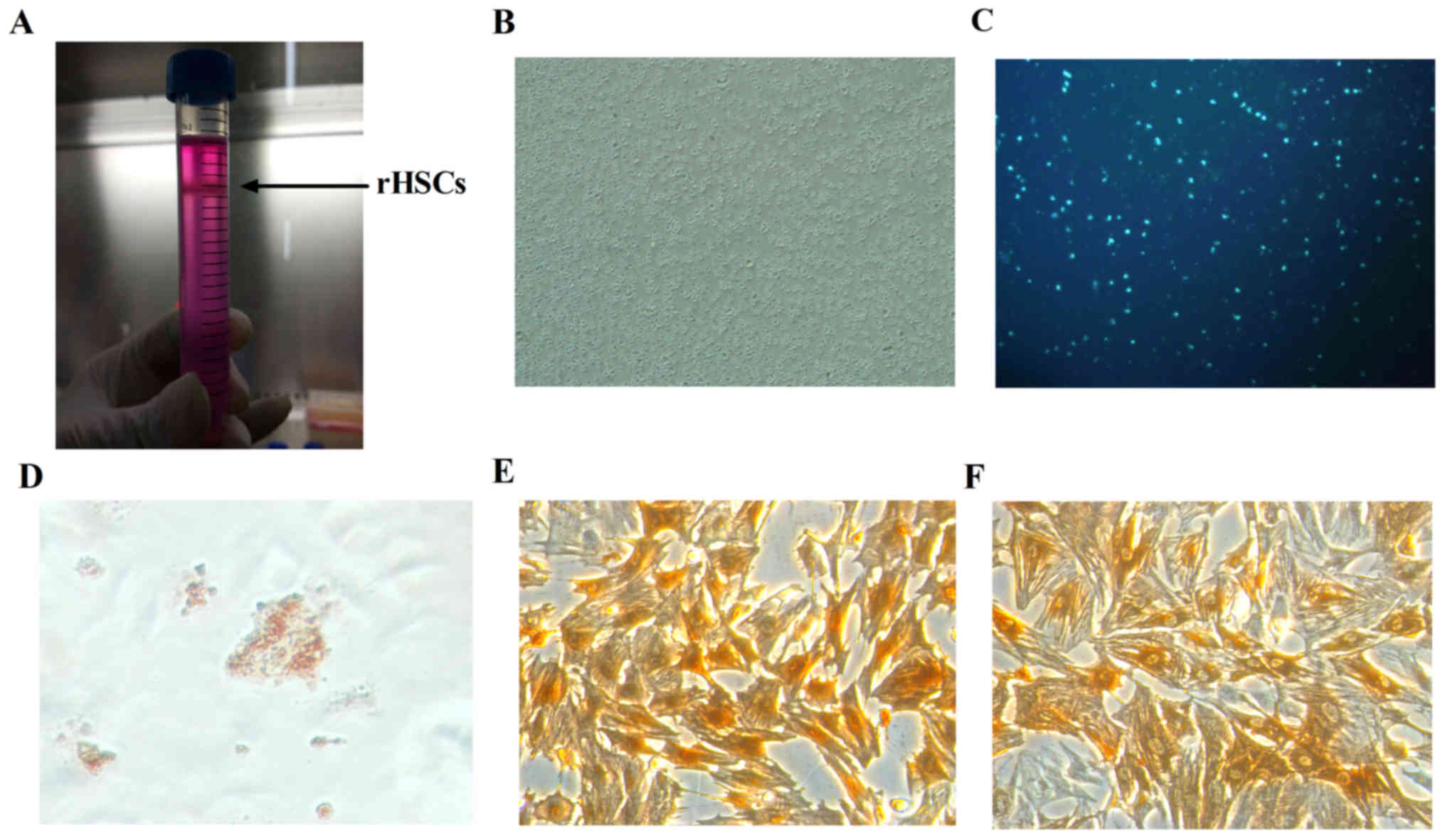
from density-gradient centrifugation (Fig. 1A). A total of 2–5×107
cells/rat were harvested. Cell viability as detected by Trypan blue
staining was >95%. Phase contrast microscopy revealed that the
morphology of plated HSCs was round and bright (Fig. 1B).
A typical feature of HSCs is the presence of
cytoplasmic, triacylglycerol rich droplets (14). In the present study, rat HSCs
emitted a striking, rapidly fading blue-green autofluorescence when
excited by ultraviolet light at 328 nm (Fig. 1C). Quiescent HSCs may be identified
with the Oil Red O stain. The retinoid droplets of HSCs cultured
for 72 h were observed to be bright red under the light microscope
(Fig. 1D). HSCs were identified
using desmin, a reliable HSC marker (15) and α-SMA, an activated HSC marker.
HSCs were cultured for 7 days and immunocytochemical analysis
demonstrated that desmin (Fig. 1E)
and α-SMA (Fig. 1F) were expressed
in the HSC cytoplasm. The purity of the isolated HSCs was 93% and
the degree of HSC activation was 96%.
IL-10 promotes the senescence of
primary rat HSCs in vitro
The most widely used assay for cell senescence is
the histochemical detection of β-galactosidase activity at pH 6.0,
which is known as SA-β-Gal (4). In
the present study, the effect of IL-10 on the senescence of primary
rat HSCs was examined by SA-β-Gal staining. Cells were incubated in
serum-free medium for 24 h, followed by incubation with different
concentrations of IL-10 for a further 24 h prior to staining. The
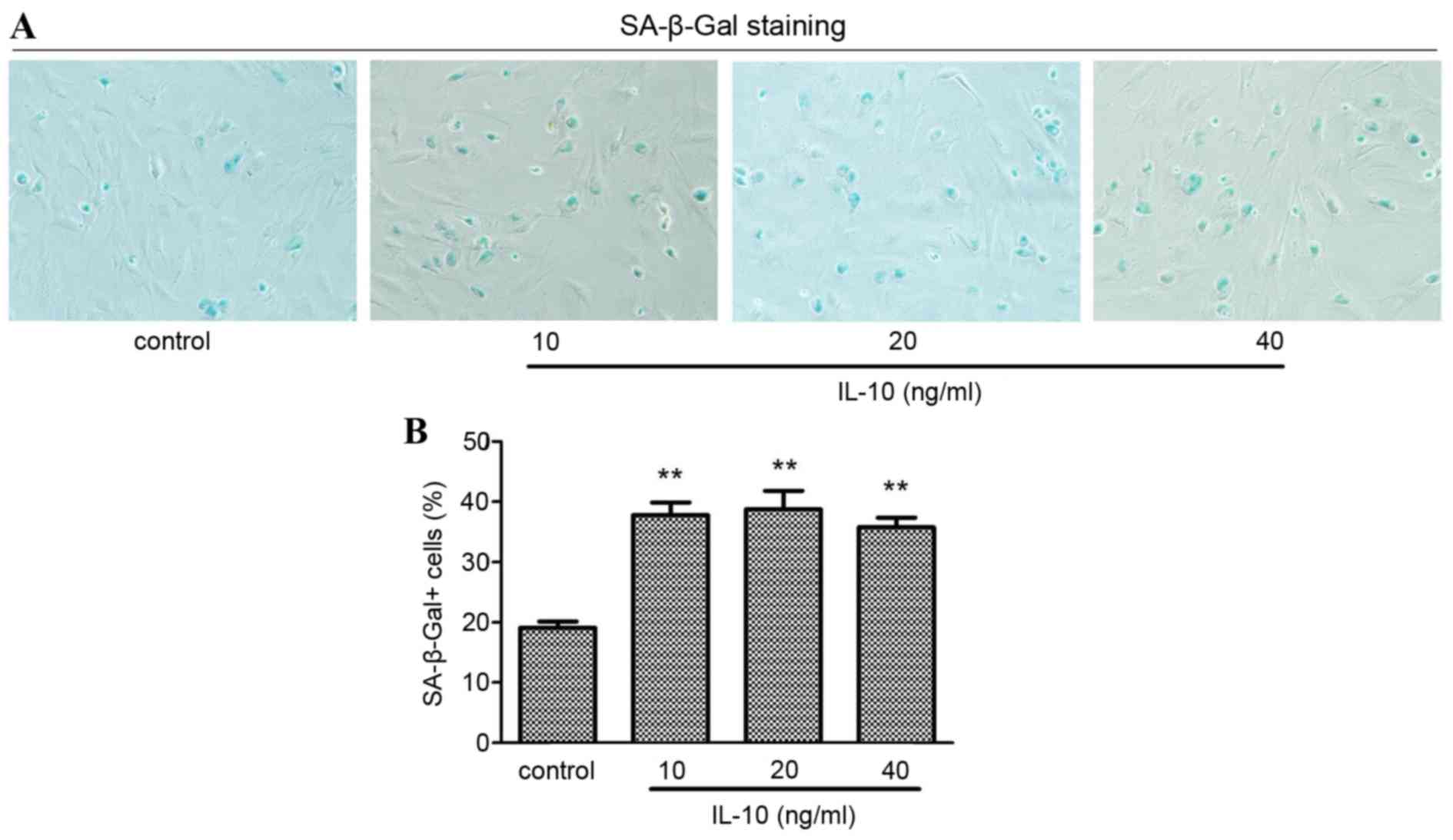
senescent primary rat HSCs cytoplasm was stained blue (Fig. 2A). The number of SA-β-Gal positive
cells was significantly increased in HSCs treated in all IL-10
groups compared with the cells in the control group (P<0.01).
The number of SA-β-Gal positive cells peaked in the 20 ng/ml IL-10
treatment group and subsequently decreased (Fig. 2B).
IL-10 promotes apoptosis and decreases
the viability of activated primary rat HSCs
To investigate if IL-10 promoted primary rat HSCs
senescence by decreasing viability or promoting apoptosis, flow
cytometry analysis and a CCK-8 assay were performed, respectively.
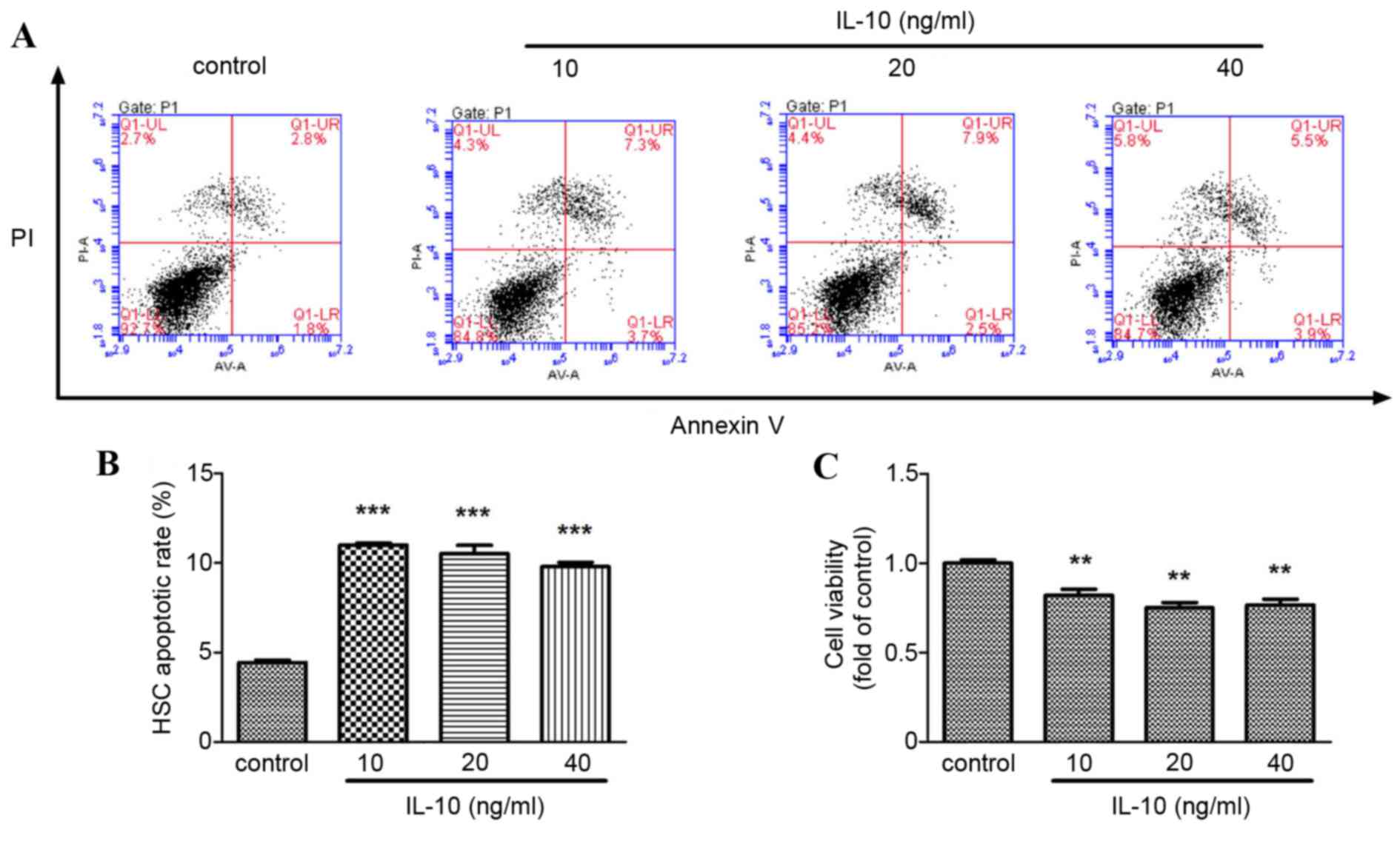
The apoptotic rate of primary rat HSCs increased significantly in
the IL-10 treatment groups compared with the control group
(P<0.001; Fig. 3A and B) and
HSC viability decreased significantly in the IL-10 treatment groups
compared with the control group (P<0.01; Fig. 3C).
IL-10 promotes the expression of
senescence-associated proteins p53 and p21
p53 and p21 have a key role in cellular senescence,
responding to a range of cellular damage signals (4). To examine the role of p53 and p21 in
IL-10-induced senescence, p53 and p21 expression was detected in
the absence or presence of IL-10 by RT-qPCR and western blot
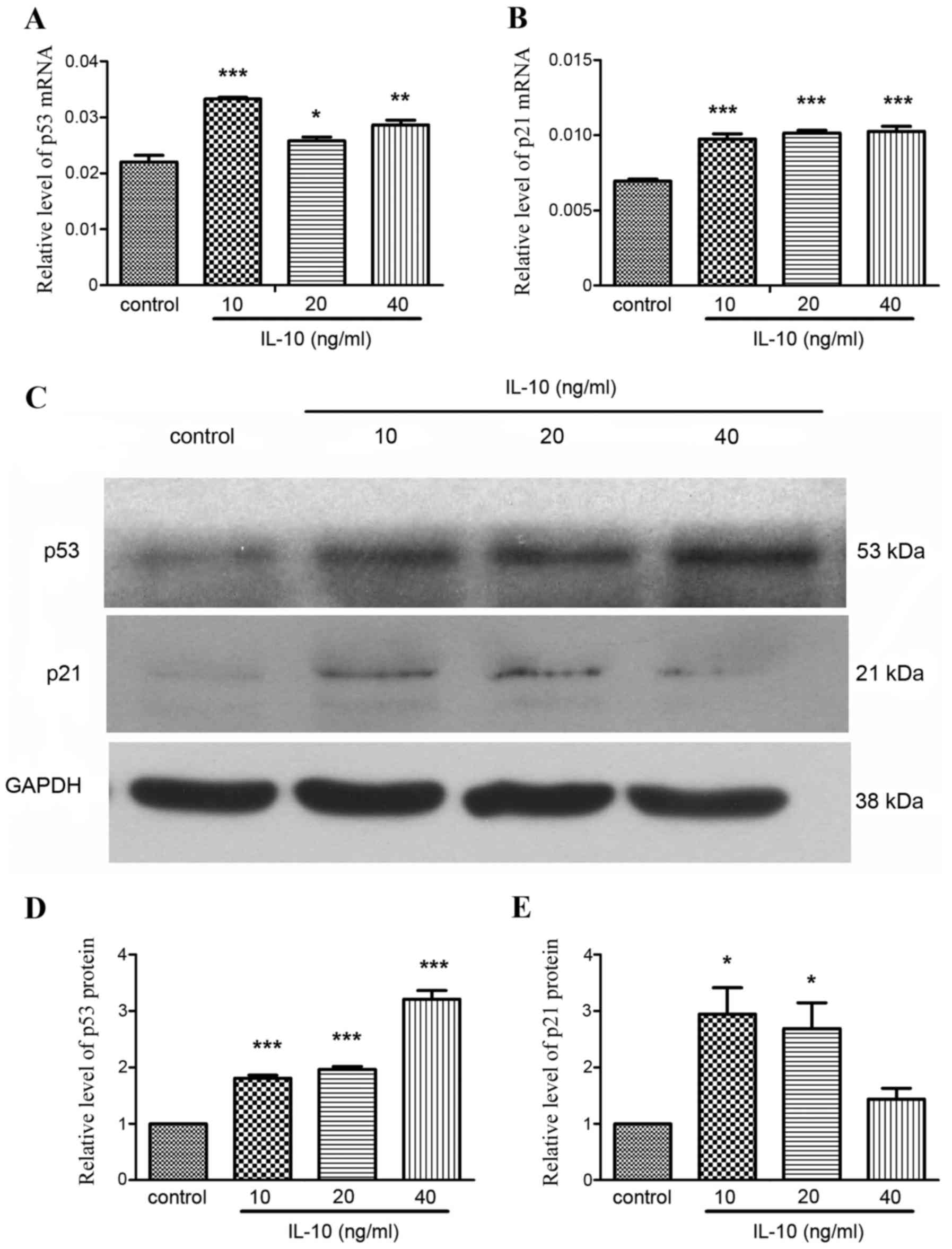
analysis. The expression of p53 mRNA was significantly increased in
the IL-10 treatment groups compared with the control group
(P<0.05; Fig. 4A), particularly
in the 10 ng/ml group (P<0.001). p21 mRNA expression was also
significantly increased in all IL-10 treatment groups (P<0.001;
Fig. 4B). Western blot analysis
demonstrated that that p53 and p21 protein expression was increased
in the IL-10 treatment groups (Fig.
4C). Relative quantification of protein expression revealed
that p53 expression increased in a dose-dependent manner (Fig. 4D). Relative expression of p21 was
highest in the 10 and 20 ng/ml IL-10 treatment groups (Fig. 4E).
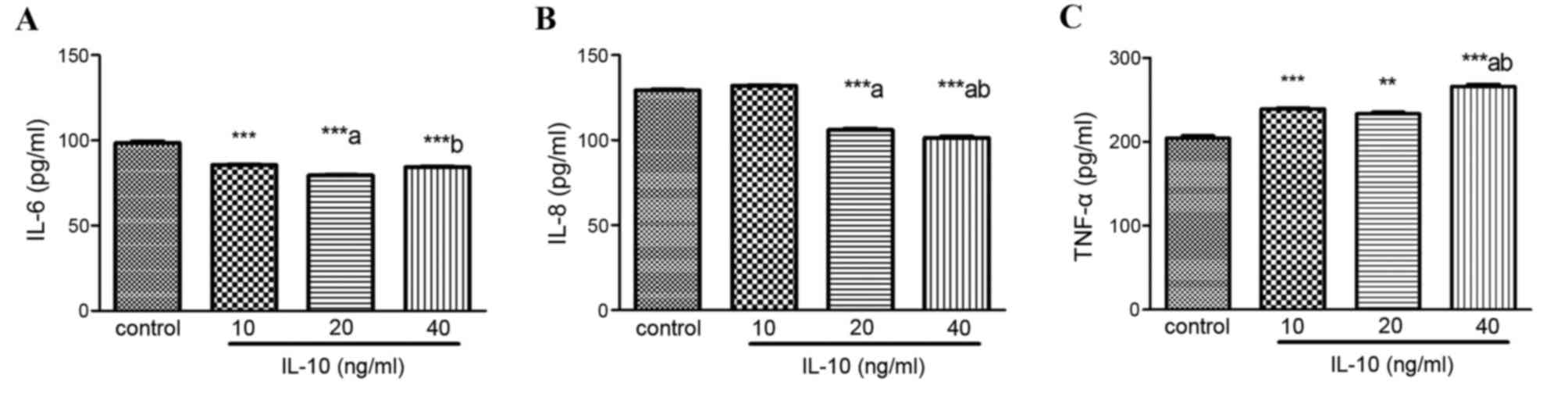
IL-10 regulates the expression of
senescence-associated inflammatory cytokines
Senescent cells secrete a number of inflammatory
cytokines and chemokines that reinforce and propagate senescence in
an autocrine and paracrine manner (4). The present study detected the
expression of IL-6, IL-8 and TNF-α in HSCs treated with increasing
concentrations of IL-10 for 24 h. Compared with the control, IL-6
expression was markedly inhibited by IL-10 in a dose-dependent
manner (P<0.001; Fig. 5A). IL-8
levels were also significantly decreased in the 20 and 40 ng/ml
IL-10 groups (P<0.001; Fig.
5B). Conversely, TNF-α levels significantly increased in IL-10
treatment groups, particularly at 40 ng/ml (P<0.001; Fig. 5C).
Discussion
HSCs contribute to liver fibrosis and activated HSCs
are considered the primary cellular target for anti-fibrotic
therapies in all liver disease etiologies (16). Primary HSCs are a key tool in
investigating the mechanisms underlying liver fibrosis and the
efficacy of anti-fibrotic therapies. In the present study, primary
rat HSCs were isolated by collagenase perfusion and
density-gradient centrifugation and the effects of IL-10 on the
senescence of activated HSCs was examined. Non-toxic and chemically
near-inert components are ideal for density gradient centrifugation
and it has been demonstrated that Histodenz is a suitable choice of
medium (13). Therefore, Histodenz
was used to isolate rat HSCs in the present study. A high yield
(2–6×107 cells/rat) of HSCs was obtained through 12%
Histodenz density-gradient centrifugation and desmin staining
determined that the HSCs were >90% pure. There are various steps
that may be taken to increase the yield, purity and viability of
HSCs. DMEM overlay must be added slowly, rats must weight >500
g, undigested debris after filter not to triturate and the time
taken to isolate and digest the cells in vitro must be as
short as possible (15). The
present study demonstrated that freshly isolated HSCs stained
positive for retinoid droplets and expressed desmin. Following
culture and passage for 7 days, cells were activated and expressed
α-SMA. These results are consistent with previous reports (17) and lay the foundation for further
study on the effect of IL-10 on HSC senescence.
The inhibition and removal of activated HSCs is a
key strategy in the treatment of liver fibrosis (18). A previous study suggested that
activated HSC senescence limits the extent of fibrosis following
hepatic injury and that senescent HSCs lose the ability to produce
collagen and proliferate (1).
Cytokines may promote cellular senescence (5). IL-22 is a member of the IL-10 family
that has been demonstrated to induce HSC senescence and reduce
liver fibrosis in mice (19).
Additionally, results of the author's previous study revealed that
IL-10 attenuates porcine serum-induced liver fibrosis by inhibiting
HSC activation (10). In a model
of carbon tetrachloride-induced liver fibrosis, SA-β-Gal activity
is detected in the fibrotic liver and SA-β-Gal positive cell
accumulation in the area of fibrotic scar tissue occurs at an
increased rate in IL-10 treated rats compared with untreated liver
sections (Huang et al, unpublished data), suggesting that
the anti-fibrotic effect of IL-10 may be associated with activated
HSC senescence. The present study demonstrated that the senescence
of HSCs increased following treatment with IL-10 in
vitro.
Cellular senescence is a form of irreversible cell
cycle arrest that occurs in response to cellular stressors,
including DNA damage, oxidative stress, oncogene activation,
chromatin disruption and telomere erosion (4). Senescent cells differ from other
non-dividing cells. They have SA-β-Gal activity, an absence of
proliferative markers and express tumor suppressors and cell cycle
inhibitors (4). The most widely
used assay for senescence is the histochemical detection of
SA-β-Gal activity (4). The present
study demonstrated that IL-10 treatment increased the activity of
SA-β-Gal in HSCs. Senescent HSCs may inhibit HSC viability
(20). The current study also
revealed that IL-10 treatment not only limited HSC viability,
however also promoted HSC apoptosis.
The primary mechanisms that control cell senescence
are p53-dependent and retinoblastoma-dependent (5). The p53-dependent pathway has been
recognized as one of the most important signal transduction
pathways for the induction of cell senescence (4). p53 promotes senescence by
transactivating genes including p21, microRNA-34 and B cell
lymphoma-2 associated X, apoptosis regulator which result in cell
cycle progression and viability inhibition (21). Previous studies have demonstrated
that p53-p21 signaling pathway activation may induce HSC senescence
and inhibit HSC viability (18,20,22).
p53 knockout mice exhibit severe hepatic fibrosis due to the
inhibition of HSC senescence (23). The present study investigated if
IL-10 induced HSC senescence through the upregulation of the
p53-p21 signaling pathway. The results demonstrated that p53 and
p21 mRNA and protein expression was significantly increased
following IL-10 treatment, suggesting that the p53-p21 dependent
pathway has an important role in IL-10-induced HSC senescence.
Cellular senescence not only decreases the viability
of damaged cells, however also affects the microenvironment through
the secretion of various pro-inflammatory cytokines, chemokines and
proteases. This is termed the senescence-associated secretory
phenotype (SASP) (24). SASP
factors include IL-1α, IL-1β, IL-6, IL-8 and TNF-α (6,25).
IL-6, IL-8 and TNF-α have been demonstrated to reinforce the
processes of senescence (6,7). The
upregulation of inflammatory cytokine expression makes HSCs
susceptible to apoptosis through natural killer cell-mediated
apoptosis, which limits the progression of fibrosis (3). To the best of the author's knowledge,
the ability of IL-10 to promote activated HSC senescence though the
upregulation of SASP has not yet been investigated. Therefore, the
present study detected IL-6, IL-8, and TNF-α expression in
activated HSCs following IL-10 treatment. The results revealed that
IL-10 inhibited the expression of IL-6 and IL-8 and increased the
expression of TNF-α. Li et al (7) demonstrated that TNF-α may promote the
premature senescence of rat nucleus pulposus cells. The mechanism
of IL-6, IL-8 and TNF-α in IL-10-induced HSC senescence requires
further study.
In conclusion, the present study demonstrated that
primary rat HSCs may be successfully isolated at a high yield,
viability and purity by the collagenase perfusion and
density-gradient centrifugation method. IL-10 was revealed to
promote the senescence of activated HSCs by upregulating the
expression of p53 and p21.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81600486), the
Fujian Province Nature Scientific Foundation of China (grant no.
2016J01462) and the Key Clinical Specialty Discipline Construction
Program of Fujian and Nation, P.R. China.
References
|
1
|
Seki E and Brenner DA: Recent advancement
of molecular mechanisms of liver fibrosis. J Hepatobiliary Pancreat
Sci. 22:512–518. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ramachandran P, Iredale JP and Fallowfield
JA: Resolution of liver fibrosis: Basic mechanisms and clinical
relevance. Semin Liver Dis. 35:119–131. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Krizhanovsky V, Yon M, Dickins RA, Hearn
S, Simon J, Miething C, Yee H, Zender L and Lowe SW: Senescence of
activated stellate cells limits liver fibrosis. Cell. 134:657–667.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Muñoz-Espín D and Serrano M: Cellular
senescence: From physiology to pathology. Nat Rev Mol Cell Bio.
15:482–496. 2014. View
Article : Google Scholar
|
|
5
|
Kojima H, Inoue T, Kunimoto H and Nakajima
K: IL-6-STAT3 signaling and premature senescence. JAKSTAT.
2:e257632014.
|
|
6
|
Kuilman T, Michaloglou C, Vredeveld LC,
Douma S, van Doorn R, Desmet CJ, Aarden LA, Mooi WJ and Peeper DS:
Oncogene-induced senescence relayed by an interleukin-dependent
inflammatory network. Cell. 133:1019–1031. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li P, Gan Y, Xu Y, Song L, Wang L, Ouyang
B, Zhang C and Zhou Q: The inflammatory cytokine TNF-α promotes the
premature senescence of rat nucleus pulposus cells via the PI3K/Akt
signaling pathway. Sci Rep. 7:429382017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hung KS, Lee TH, Chou WY, Wu CL, Cho CL,
Lu CN, Jawan B and Wang CH: Interleukin-10 gene therapy reverses
thioacetamide-induced liver fibrosis in mice. Biochem Biophys Res
Commun. 336:324–331. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang YH, Shi MN, Zheng WD, Zhang LJ, Chen
ZX and Wang XZ: Therapeutic effect of interleukin-10 on
CCl4-induced hepatic fibrosis in rats. World J Gastroenterol.
12:1386–1391. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang YH, Chen YX, Zhang LJ, Chen ZX and
Wang XZ: Hydrodynamics-based transfection of rat interleukin-10
gene attenuates porcine serum-induced liver fibrosis in rats by
inhibiting the activation of hepatic stellate cells. Int J Mol Med.
34:677–686. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shi MN, Huang YH, Zheng WD, Zhang LJ, Chen
ZX and Wang XZ: Relationship between transforming growth factor
beta1 and anti-fibrotic effect of interleukin-10. World J
Gastroenterol. 12:2357–2362. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Weiskirchen R and Gressner AM: Isolation
and culture of hepatic stellate cells. Methods Mol Med. 117:99–113.
2005.PubMed/NCBI
|
|
14
|
Chang W, Yang M, Song L, Shen K, Wang H,
Gao X, Li M, Niu W and Qin X: Isolation and culture of hepatic
stellate cells from mouse liver. Acta Biochim Biophys Sin
(Shanghai). 46:291–298. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mederacke I, Dapito DH, Affò S, Uchinami H
and Schwabe RF: High-yield and high-purity isolation of hepatic
stellate cells from normal and fibrotic mouse livers. Nat Protoc.
10:305–315. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mederacke I, Hsu CC, Troeger JS, Huebener
P, Mu X, Dapito DH, Pradere JP and Schwabe RF: Fate tracing reveals
hepatic stellate cells as dominant contributors to liver fibrosis
independent of its aetiology. Nat Commun. 4:28232013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim KH, Chen CC, Monzon RI and Lau LF:
Matricellular protein CCN1 promotes regression of liver fibrosis
through induction of cellular senescence in hepatic myofibroblasts.
Mol Cell Biol. 33:2078–2090. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jin H, Lian N, Zhang F, Bian M, Chen X,
Zhang C, Jia Y, Lu C, Hao M, Yao S, et al: Inhibition of YAP
signaling contributes to senescence of hepatic stellate cells
induced by tetramethylpyrazine. Eur J Pharm Sci. 96:323–333. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kong X, Feng D, Wang H, Hong F, Bertola A,
Wang FS and Gao B: Interleukin-22 induces hepatic stellate cell
senescence and restricts liver fibrosis in mice. Hepatology.
56:1150–1159. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen J, Xu T, Zhu D, Wang J, Huang C, Lyu
L, Hu B, Sun W and Duan Y: Egg antigen p40 of Schistosoma japonicum
promotes senescence in activated hepatic stellate cells by
activation of the STAT3/p53/p21 pathway. Cell Death Dis.
7:e23152016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
He L, He X, Lowe SW and Hannon GJ:
microRNAs join the p53 network-another piece in the
tumour-suppression puzzle. Nat Rev Cancer. 7:819–822. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ahsan MK and Mehal WZ: Activation of
adenosine receptor A2A increases HSC proliferation and inhibits
death and senescence by down-regulation of p53 and Rb. Front
Pharmacol. 5:692014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kong D, Zhang F, Zhang Z, Lu Y and Zheng
S: Clearance of activated stellate cells for hepatic fibrosis
regression: Molecular basis and translational potential. Biomed
Pharmacother. 67:246–250. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang Z, Yao Z, Zhao S, Shao J, Chen A,
Zhang F and Zheng S: Interaction between autophagy and senescence
is required for dihydroartemisinin to alleviate liver fibrosis.
Cell Death Dis. 8:e28862017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Braumüller H, Wieder T, Brenner E, Aßmann
S, Hahn M, Alkhaled M, Schilbach K, Essmann F, Kneilling M,
Griessinger C, et al: T-helper-1-cell cytokines drive cancer into
senescence. Nature. 494:361–365. 2013. View Article : Google Scholar : PubMed/NCBI
|